Antimicrobial Properties of Apis mellifera’s Bee Venom

 ,
,  , , ,
, , ,  ,
,
Abstract
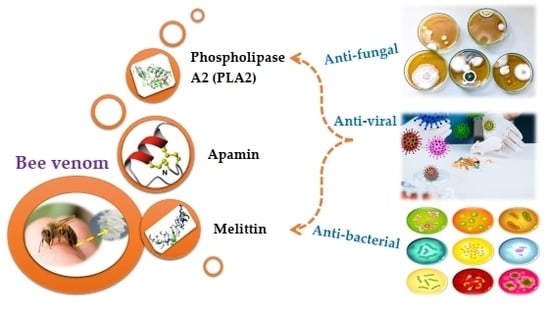
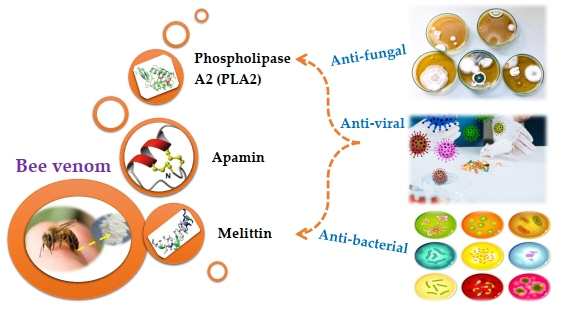
:
1. Introduction
2. Antimicrobial Properties of Bee Venom and Mode of Action for the Venom and its Derived Compounds
2.1. Antibacterial
2.2. Anti-Viral
2.3. Anti-fungal
3. Concluding Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- Rehman, K.; Fiayyaz, F.; Khurshid, M.; Sabir, S. Antibiotics and Antimicrobial Resistance: Temporal and Global Trends in the Environment; Elsevier Inc.: Amsterdam, The Netherlands, 2020; Chapter 2; ISBN 9780128188828. [Google Scholar]
- Memariani, H.; Memariani, M.; Shahidi-Dadras, M.; Nasiri, S.; Akhavan, M.M.; Moravvej, H. Melittin: From honeybees to superbugs. Appl. Microbiol. Biotechnol. 2019, 103, 3265–3276. [Google Scholar] [CrossRef]
- Shin, S.; Ye, M.; Choi, S.; Park, K. The effects of melittin and apamin on airborne Fungi-Induced chemical mediator and extracellular matrix production from nasal polyp fibroblasts. Toxicon 2017, 9, 384. [Google Scholar] [CrossRef] [Green Version]
- Leandro, L.F.; Mendes, C.A.; Casemiro, L.A. Antimicrobial activity of apitoxin, melittin and phospholipase A 2 of honey bee (Apis mellifera) venom against oral pathogens. An. Acad. Bras. Cienc. 2015, 87, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Han, S.M.; Kim, J.M.; Hong, I.P.; Woo, S.O.; Kim, S.G.; Jang, H.R.; Pak, S.C. Antibacterial activity and antibiotic-enhancing effects of honeybee venom against methicillin-resistant Staphylococcus aureus. Molecules 2016, 21, 79. [Google Scholar] [CrossRef] [Green Version]
- Socarras, K.M.; Theophilus, P.A.S.; Torres, J.P.; Gupta, K.; Sapi, E. Antimicrobial activity of bee venom and melittin against borrelia burgdorferi. Antibiotics 2017, 6, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yacoub, T.; Rima, M.; Karam, M.; Fajloun, J.M.S. Antimicrobials from venomous animals: An oeriew. Molecules 2020, 25, 2402. [Google Scholar] [CrossRef]
- De Barros, E.; Gonçalves, R.M.; Cardoso, M.H.; Santos, N.C.; Franco, O.L.; Cândido, E.S. Snake venom cathelicidins as natural antimicrobial peptides. Front. Pharmacol. 2019, 10, 1415–1427. [Google Scholar] [CrossRef] [Green Version]
- Das Neves, R.C.; Mortari, M.R.; Schwartz, E.F.; Kipnis, A.; Junqueira-Kipnis, A.P. Antimicrobial and antibiofilm effects of peptides from venom of social wasp and scorpion on multidrug-resistant Acinetobacter baumannii. Toxins 2019, 11, 216. [Google Scholar] [CrossRef] [Green Version]
- Samy, R.P.; Gopalakrishnakone, P.; Thwin, M.M.; Chow, T.K.V.; Bow, H.; Yap, E.H.; Thong, T.W.J. Antibacterial activity of snake, scorpion and bee venoms: A comparison with purified venom phospholipase A2 enzymes. J. Appl. Microbiol. 2007, 102, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Flávia, A.; Pereira, M.; Albano, M.; Cristina, F.; Alves, B.; Fernanda, B.; Teles, M.; Furlanetto, A.; Mores, V.L. Influence of apitoxin and melittin from Apis mellifera bee on Staphylococcus aureus strains. Microb. Pathog. 2020, 141, 104011. [Google Scholar]
- Wang, L.; Zhao, X.; Zhu, C.; Zhao, Y.; Liu, S.; Xia, X.; Liu, X.; Zhang, H.; Xu, Y.; Hang, B.; et al. The antimicrobial peptide MPX kills Actinobacillus pleuropneumoniae and reduces its pathogenicity in mice. Vet. Microbiol. 2020, 243, 108634. [Google Scholar] [CrossRef] [PubMed]
- Baracchi, D.; Francese, S.; Turillazzi, S. Beyond the antipredatory defence: Honey bee venom function as a component of social immunity. Toxicon 2011, 58, 550–557. [Google Scholar] [CrossRef] [Green Version]
- Pak, S.C. An introduction to the Toxins special issue on “Bee and wasp venoms: Biological characteristics and therapeutic application”. Toxins 2016, 8, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellik, Y. Bee Venom: Its potential use in alternative medicine. Anti-Infect. Agents 2015, 13, 3–16. [Google Scholar] [CrossRef]
- AL-Ani, I.; Zimmermann, S.; Reichling, J.; Wink, M. Pharmacological synergism of bee venom and melittin with antibiotics and plant secondary metabolites against multi-drug resistant microbial pathogens. Phytomedicine 2015, 22, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Janik, J.E.; Wania-Galicia, L.; Kalauokalani, D. Bee stings—A remedy for postherpetic neuralgia? A case report. Reg. Anesth. Pain Med. 2007, 32, 533–535. [Google Scholar] [CrossRef]
- Dong, J.; Ying, B.; Huang, S.; Ma, S.; Long, P.; Tu, X.; Yang, W.; Wu, Z.; Chen, W.; Miao, X. High-Performance liquid chromatography combined with intrinsic fluorescence detection to analyse melittin in individual honeybee (Apis mellifera) venom sac. J. Chromatogr. B 2015, 1002, 139–143. [Google Scholar] [CrossRef]
- Huh, J.; Kang, J.W.; Nam, D.; Baek, Y.; Choi, D. Melittin suppresses VEGFA-induced tumor growth by blocking VEGFR-2 and the COX-2-mediated MAPK signaling pathway. J. Nat. Prod. 2012, 75, 1922–1929. [Google Scholar] [CrossRef]
- Buku, A.; Reibman, J.; Pistelli, A.; Blandina, P.; Gazis, D. Mast cell degranulating (MCD) peptide analogs with reduced ring structure. J. Protein Chem. 1992, 11, 275–280. [Google Scholar] [CrossRef]
- Mourelle, D.; Brigatte, P.; Bringanti, L.D.B.; De Souza, B.M.; Arcuri, H.A.; Gomes, P.C.; Baptista-Saidemberg, N.B.; Ruggiero Neto, J.; Palma, M.S. Hyperalgesic and edematogenic effects of Secapin-2, a peptide isolated from Africanized honeybee (Apis mellifera) venom. Peptides 2014, 59, 42–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Y.; Yang, X.X.; Sheng, Y.X.; Zhang, J.L.; Yu, D.Q. A novel peptide from Apis mellifera and solid-phase synthesis of its analogue. Chin. Chem. Lett. 2012, 23, 1161–1164. [Google Scholar] [CrossRef]
- Gauldie, J.; Hanson, J.M.; Shipolini, R.A.; Vernon, C.A. The structures of some peptides from bee venom. Eur. J. Biochem. 1978, 83, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Shkenderov, S.; Koburova, K. Adolapin—A newly isolated analgetic and anti-inflammatory polypeptide from bee venom. Toxicon 1982, 20, 317–321. [Google Scholar] [CrossRef]
- Drici, M.D.; Diochot, S.; Terrenoire, C.; Romey, G.; Lazdunski, M. The bee venom peptide tertiapin underlines the role of I(KACh) in acetylcholine-induced atrioventricular blocks. Br. J. Pharmacol. 2000, 131, 569–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgieva, D.; Greunke, K.; Genov, N.; Betzel, C. 3-D Model of the bee venom acid phosphatase: Insights into allergenicity. Biochem. Biophys. Res. Commun. 2009, 378, 711–715. [Google Scholar] [CrossRef]
- Rybak-Chmielewska, H.; Szczęsna, T. HPLC study of chemical composition of honeybee (Apis mellifera L.) venom. J. Apic. Sci. 2004, 48, 103–109. [Google Scholar]
- Shipolini, R.A.; Callewaert, G.L.; Cottrell, R.C.; Doonan, S.; Vernon, C.A.; Banks, B.E. Phospholipase A from bee venom. Eur. J. Biochem. 1971, 20, 459–468. [Google Scholar] [CrossRef]
- Blank, S.; Seismann, H.; Bockisch, B.; Braren, I.; Cifuentes, L.; McIntyre, M.; Rühl, D.; Ring, J.; Bredehorst, R.; Ollert, M.W.; et al. Identification, recombinant expression, and characterization of the 100 kDa high molecular weight Hymenoptera venom allergens Api m 5 and Ves v 3. J. Immunol. 2010, 184, 5403–5413. [Google Scholar]
- Kettner, A.; Hughes, G.J.; Frutiger, S.; Astori, M.; Roggero, M.; Spertini, F.; Corradin, G. Api m 6: A new bee venom allergen. J. Allergy Clin. Immunol. 2001, 107, 914–920. [Google Scholar] [CrossRef]
- Georgieva, D.; Greunke, K.; Betzel, C. Three-dimensional model of the honeybee venom allergen Api m 7: Structural and functional insights. Mol. Biosyst. 2010, 6, 1056–1060. [Google Scholar] [CrossRef]
- Blank, S.; Seismann, H.; Michel, Y.; Mcintyre, M.; Cifuentes, L.; Braren, I.; Grunwald, T.; Darsow, U.; Ring, J.; Bredehorst, R.; et al. Api m 10, a genuine A. mellifera venom allergen, is clinically relevant but underrepresented in therapeutic extracts. Eur. J. Allergy Clin. Immunol. 2011, 66, 1322–1329. [Google Scholar] [CrossRef] [PubMed]
- Blank, S.; Bantleon, F.I.; McIntyre, M.; Ollert, M.; Spillner, E. The major royal jelly proteins 8 and 9 (Api m 11) are glycosylated components of Apis mellifera venom with allergenic potential beyond carbohydrate-based reactivity. Clin. Exp. Allergy 2012, 42, 976–985. [Google Scholar] [CrossRef] [PubMed]
- Peiren, N.; de Graaf, D.C.; Vanrobaeys, F.; Danneels, E.L.; Devreese, B.; Van Beeumen, J.; Jacobs, F.J. Proteomic analysis of the honey bee worker venom gland focusing on the mechanisms of protection against tissue damage. Toxicon 2008, 52, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Pickets, J.A.; Williams, I.H.; Martin, A.P. (Z)-11-Eicosen-1-ol, an important new pheromonal component from the sting of the honey bee, Apis mellifera L. (Hymenoptera, Apidae). J. Chem. Ecol. 1982, 8, 163–175. [Google Scholar] [CrossRef]
- Kocyigit, A.; Guler, E.M.; Kaleli, S. Anti-inflammatory and antioxidative properties of honey bee venom on freund’s complete adjuvant-induced arthritis model in rats. Toxicon 2019, 161, 4–11. [Google Scholar] [CrossRef]
- Sobral, F.; Sampaio, A.; Falcão, S.; João, M.; Queiroz, R.P.; Ricardo, C. Chemical characterization, antioxidant, anti-inflammatory and cytotoxic properties of bee venom collected in Northeast Portugal Filipa. Food Chem. Toxicol. 2016, 94, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Varanda, E.A.; Tavares, D.C. Radioprotection: Mechanisms and radioprotective agents including honeybee venom. J. Venom. Anim. Toxins 1998, 4, 5–21. [Google Scholar] [CrossRef]
- Han, S.; Yeo, J.; Baek, H.; Lin, S.M.; Meyer, S.; Molan, P. Postantibiotic effect of purified melittin from honeybee (Apis mellifera) venom against Escherichia coli and Staphylococcus aureus. J. Asian Nat. Prod. Res. 2009, 11, 796–804. [Google Scholar] [CrossRef]
- Han, S.M.; Lee, K.G.; Yeo, J.H.; Baek, H.J.; Park, K. Antibacterial and anti-inflammatory effects of honeybee (Apis mellifera) venom against acne-inducing bacteria. J. Med. Plants Res. 2010, 4, 459–464. [Google Scholar]
- McGhee, S.; Visovksy, C.; Zambroski, C.; Finnegan, A. Lyme disease: Recognition and management for emergency nurses. Emerg. Nurse 2018, 28. [Google Scholar] [CrossRef]
- Arteaga, V.; Lamas, A.; Regal, P.; Vázquez, B.; Manuel, J.; Cepeda, A.; Manuel, C. Antimicrobial activity of apitoxin from Apis mellifera in Salmonella enterica strains isolated from poultry and its effects on motility, biofilm formation and gene expression. Microb. Pthogenesis 2019, 137, 103771–103776. [Google Scholar] [CrossRef] [PubMed]
- Jung, B.; Lee, J.; Park, S.; Hyun, P.; Park, J.; Suh, G. Immunoprophylactic effects of administering honeybee (Apis melifera ) venom spray against salmonella gallinarum in broiler chicks. J. Vet. Sci. 2013, 75, 1287–1295. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.H.; Jang, A.Y.; Lin, S.; Lim, S.; Kim, D.; Park, K.; Han, S.M.; Yeo, J.H.; Seo, H.S. Melittin, a honeybee venom-derived antimicrobial peptide, may target methicillin-resistant Staphylococcus aureus. Mol. Med. Rep. 2015, 12, 6483–6490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamasbi, E.; Mularski, A.; Separovic, F. Model membrane and cell studies of antimicrobial activity of melittin analogues. Curr. Top. Med. Chem. 2016, 16, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Picoli, T.; Peter, C.M.; Zani, J.L.; Waller, S.B.; Lopes, M.G.; Boesche, K.N.; D’Ávila Vargas, G.; de Oliveira Hübner, S.; Fischer, G. Melittin and its potential in the destruction and inhibition of the biofilm formation by Staphylococcus aureus, Escherichia coli and Pseudomonas aeruginosa isolated from bovine milk. Microb. Pathog. 2017, 112, 57–62. [Google Scholar] [CrossRef]
- Lubke, L.L.; Garon, C.F. The antimicrobial agent melittin exhibits powerful in vitro inhibitory effects on the lyme disease spirochete. Clin. Infect. Dis. 1997, 25, S48–S51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarre, W.W.; Schneewind, O. Surface proteins of gram-positive bacteria and mechanisms of their targeting to the cell wall envelope. Microbiol. Mol. Biol. Rev. 1999, 63, 174–229. [Google Scholar] [CrossRef] [Green Version]
- Lüderitz, O.; Freudenberg, M.A.; Galanos, C.; Lehmann, V.; Rietschel, E.T.; Shaw, D.H. Lipopolysaccharides of gram-negative bacteria. Curr. Top. Membr. Transp. 1982, 17, 79–151. [Google Scholar]
- Jamasbi, E.; Batinovic, S.; Sharples, R.A.; Sani, M.A.; Robins-Browne, R.M.; Wade, J.D.; Separovic, F.; Hossain, M.A. Melittin peptides exhibit different activity on different cells and model membranes. Amino Acids 2014, 46, 2759–2766. [Google Scholar] [CrossRef]
- Park, D.; Jung, J.W.; Lee, M.O.; Lee, S.Y.; Kim, B.; Jin, H.J.; Kim, J.; Ahn, Y.J.; Lee, K.W.; Song, Y.S.; et al. Functional characterization of naturally occurring melittin peptide isoforms in two honey bee species, Apis mellifera and Apis cerana. Peptides 2014, 53, 185–193. [Google Scholar] [CrossRef]
- Akbari, R.; Hakemi, M.; Ali, V.; Hossein, H.; Jean, A.; Sabatier, M. Action mechanism of melittin—derived antimicrobial peptides, MDP1 and MDP2, de novo designed against multidrug resistant bacteria. Amino Acids 2018, 50, 1231–1243. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Singh, A.K.; Wu, X.; Lyu, Y.; Bhunia, A.K.; Narsimhan, G. Characterization of antimicrobial activity against Listeria and cytotoxicity of native melittin and its mutant variants. Colloids Surf. B Biointerfaces 2016, 143, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Dosler, S.; Gerceker, A.A. In vitro activities of antimicrobial cationic peptides; melittin and nisin, alone or in combination with antibiotics against Gram-positive bacteria. J. Chemother. 2012, 24, 137–143. [Google Scholar] [CrossRef]
- Giacometti, A.; Cirioni, O.; Kamysz, W.; D’Amato, G.; Silvestri, C.; Del Prete, M.S.; Łukasiak, J.; Scalise, G. Comparative activities of cecropin A, melittin, and cecropin A–melittin peptide CA (1–7) M (2–9) NH2 against multidrug-resistant nosocomial isolates of Acinetobacter baumannii. Peptides 2003, 24, 1315–1318. [Google Scholar] [CrossRef]
- Akbari, R.; Hakemi-vala, M.; Pashaie, F.; Bevalian, P.; Hashemi, A.; Bagheri, K.P. Highly synergistic effects of melittin with conventional antibiotics against multidrug-resistant isolates of acinetobacter baumannii and pseudomonas aeruginosa. Microb. Drug Resist. 2018, 25, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-T.; Hwang, J.-Y.; Sung, M.-S.; Je, S.-Y.; Bae, D.-R.; Han, S.-M.; Lee, S.-H. The minimum inhibitory concentration (MIC) of bee venom against bacteria isolated from pigs and chickens. Korean J. Vet. Serv 2006, 29, 19–26. [Google Scholar]
- Fernández, N.J.; Porrini, M.P.; Podaza, E.A.; Damiani, N.; Gende, L.B.; Eguaras, M.J. A scientific note on the first report of honeybee venom inhibiting Paenibacillus larvae growth. Apidologie 2014, 45, 719–721. [Google Scholar] [CrossRef]
- Zolfagharian, H.; Mohajeri, M.; Babaie, M. Bee venom (Apis mellifera) an effective potential alternative to gentamicin for specific bacteria strains: Bee venom an effective potential for bacteria. J. Pharmacopunct. 2016, 19, 225–230. [Google Scholar] [CrossRef]
- Frangieh, J.; Salma, Y.; Haddad, K.; Mattei, C.; Legros, C.; Fajloun, Z.; El Obeid, D. First characterization of the venom from Apis mellifera syriaca, a honeybee from the middle east region. Toxins 2019, 11, 191. [Google Scholar] [CrossRef] [Green Version]
- Vila-Farres, X.; Garcia de la Maria, C.; López-Rojas, R.; Pachón, J.; Giralt, E.; Vila, J. In vitro activity of several antimicrobial peptides against colistin-susceptible and colistin-resistant Acinetobacter baumannii. Clin. Microbiol. Infect. 2012, 18, 383–387. [Google Scholar] [CrossRef] [Green Version]
- Pashaei, F.; Bevalian, P.; Akbari, R.; Bagheri, K.P. Single dose eradication of extensively drug resistant Acinetobacter spp. in a mouse model of burn infection by melittin antimicrobial peptide. Microb. Pathog. 2019, 127, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Yokota, H.; Nagashima, Y.; Shiomi, K. Interaction of grammistins with lipids and their antibacterial activity. Fish. Sci. 2001, 67, 928–933. [Google Scholar] [CrossRef]
- Karyne, R.; Lechuga, G.C.; Souza, A.L.A.; das Silva Carvalho, J.P.R.; Bôas, M.H.S.V.; De Simone, S.G. Pan-drug resistant acinetobacter baumannii, but not other strains, are resistant to the bee venom peptide mellitin. Antibiotics 2020, 9, 178. [Google Scholar]
- Jamasbi, E.; Lucky, S.S.; Li, W.; Akhter, M.; Gopalakrishnakone, P.; Separovic, F. Effect of dimerized melittin on gastric cancer cells and antibacterial activity. Amino Acids 2018, 50, 1101–1110. [Google Scholar] [CrossRef]
- Asthana, N.; Yadav, S.P.; Ghosh, J.K. Dissection of antibacterial and toxic activity of melittin: A leucine zipper motif plays a crucial role in determining its hemolytic activity but not antibacterial activity. J. Biol. Chem. 2004, 279, 55042–55050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, S.; Li, W.; Zhang, L.; Zhang, Y.; Cao, B. Cecropin A—melittin mutant with improved proteolytic stability and enhanced antimicrobial activity against bacteria and fungi associated with gastroenteritis in vitro. Biochem. Biophys. Res. Commun. 2014, 451, 650–655. [Google Scholar] [CrossRef]
- Saugar, J.M.; Alarcón, T.; López-Hernández, S.; López-Brea, M.; Andreu, D.; Rivas, L. Activities of polymyxin B and cecropin A-melittin peptide CA (1–8) M (1–18) against a multiresistant strain of acinetobacter baumannii. Antimicrob. Agents Chemother. 2002, 46, 875–878. [Google Scholar] [CrossRef] [Green Version]
- Saravanan, R.; Bhunia, A.; Bhattacharjya, S. Micelle-bound structures and dynamics of the hinge deleted analog of melittin and its diastereomer: Implications in cell selective lysis by d-amino acid containing antimicrobial peptides. Biochim. Biophys. Acta-Biomembr. 2010, 1798, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Chen, S.; Li, S.; Yan, H.; Fan, Y.; Mi, H. Deletion of two C-terminal Gln residues of 12–26-residue fragment of melittin improves its antimicrobial activity. Peptides 2005, 26, 369–375. [Google Scholar] [CrossRef]
- Da Silva, C.M.; da Silva, D.L.; Modolo, L.V.; Alves, R.B.; de Resende, M.A.; Martins, C.V.B.; de Fátima, Â. Schiff bases: A short review of their antimicrobial activities. J. Adv. Res. 2011, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Soboksa, N.E.; Gari, S.R.; Hailu, A.B.; Alemu, B.M. Association between microbial water quality, sanitation and hygiene practices and childhood diarrhea in Kersa and Omo Nada districts of Jimma Zone, Ethiopia. PLoS ONE 2020, 15, e0229303. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Izopet, J.; Pavio, N.; Aggarwal, R.; Labrique, A.; Wedemeyer, H.; Dalton, H.R. Hepatitis E virus infection. Nat. Rev. Dis. Prim. 2017, 3, 17086–17101. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.K. Water-Associated Infectious Diseases; Springer Nature Singapore Pte Ltd.: Singapore, 2019; ISBN 9789811391972. [Google Scholar]
- Memariani, H.; Memariani, M.; Moravvej, H.; Shahidi-Dadras, M. Melittin: A venom-derived peptide with promising anti-viral properties. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.B.; Lee, B.-H.; Nikapitiya, C.; Kim, J.-H.; Kim, T.-H.; Lee, H.-C.; Kim, C.G.; Lee, J.-S.; Kim, C.-J. Inhibitory effects of bee venom and its components against viruses in vitro and in vivo. J. Microbiol. 2016, 54, 853–866. [Google Scholar] [CrossRef]
- Kim, Y.-W.; Chaturvedi, P.; Chun, S.; Lee, Y.; Ahn, W. Honeybee venom possesses anticancer and antiviral effects by differential inhibition of HPV E6 and�E7 expression on cervical cancer cell line. Oncol. Rep. 2015, 33, 1675–1682. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-A.; Kim, Y.-M.; Hyun, P.-M.; Jeon, J.-W.; Park, J.-K.; Suh, G.-H.; Jung, B.-G.; Lee, B.-J. Honeybee (Apis mellifera) venom reinforces viral clearance during the early stage of infection with porcine reproductive and respiratory syndrome virus through the up-regulation of Th1-specific immune responses. Toxins 2015, 7, 1837–1853. [Google Scholar] [CrossRef] [Green Version]
- Fenard, D.; Lambeau, G.; Maurin, T.; Lefebvre, J.-C.; Doglio, A. A peptide derived from bee venom-secreted phospholipase A2 inhibits replication of T-cell tropic HIV-1 strains via interaction with the CXCR4 chemokine receptor. Mol. Pharmacol. 2001, 60, 341–347. [Google Scholar] [CrossRef] [Green Version]
- Fenard, D.; Lambeau, G.; Valentin, E.; Lefebvre, J.-C.; Lazdunski, M.; Doglio, A. Secreted phospholipases A 2, a new class of HIV inhibitors that block virus entry into host cells. J. Clin. Investig. 1999, 104, 611–618. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Aoki-Utsubo, C.; Kameoka, M.; Deng, L.; Terada, Y.; Kamitani, W.; Sato, K.; Koyanagi, Y.; Hijikata, M.; Shindo, K. Broad-spectrum antiviral agents: Secreted phospholipase A 2 targets viral envelope lipid bilayers derived from the endoplasmic reticulum membrane. Sci. Rep. 2017, 7, 15931. [Google Scholar] [CrossRef]
- Albiol Matanic, V.C.; Castilla, V. Antiviral activity of antimicrobial cationic peptides against Junin virus and herpes simplex virus. Int. J. Antimicrob. Agents 2004, 23, 382–389. [Google Scholar] [CrossRef]
- Baghian, A.; Kousoulas, K.G. Role of the Na+, K+ pump in herpes simplex type 1-induced cell fusion: Melittin causes specific reversion of syncytial mutants with the syn1 mutation to Syn+ (wild-type) phenotype. Virology 1993, 196, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Wachinger, M.; Kleinschmidt, A.; Winder, D.; von Pechmann, N.; Ludvigsen, A.; Neumann, M.; Holle, R.; Salmons, B.; Erfle, V.; Brack-Werner, R. Antimicrobial peptides melittin and cecropin inhibit replication of human immunodeficiency virus 1 by suppressing viral gene expression. J. Gen. Virol. 1998, 79, 731–740. [Google Scholar] [CrossRef]
- Kamal, S.A. In vitro study on the effect of bee venom on some cell lines and lumpy skin disease virus. J. Agric. Sci. Technol. A 2016, 6, 124–135. [Google Scholar]
- Fujii, G.; Horvath, S.; Woodward, S.; Eiserling, F.; Eisenberg, D. A molecular model for membrane fusion based on solution studies of an amphiphilic peptide from HIV gp41. Protein Sci. 1992, 1, 1454–1464. [Google Scholar] [CrossRef] [Green Version]
- Yasin, B.; Pang, M.; Turner, J.S.; Cho, Y.; Dinh, N.N.; Waring, A.J.; Lehrer, R.I.; Wagar, E.A. Evaluation of the inactivation of infectious herpes simplex virus by host-defense peptides. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-B. Antifungal activity of bee venom and sweet bee venom against clinically isolated candida albicans. J. Pharmacopunct. 2016, 19, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.R.; Kim, J.J.; Park, G.S.; Oh, S.M.; Han, C.S.; Lee, M.Y. The antifungal activity of bee venom against dermatophytes. J. Appl. Biol. Chem. 2012, 55, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Lee, D.G. Melittin triggers apoptosis in Candida albicans through the reactive oxygen species-mediated mitochondria/caspase-dependent pathway. FEMS Microbiol. Lett. 2014, 355, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Park, C.; Lee, D.G. Melittin induces apoptotic features in Candida albicans. Biochem. Biophys. Res. Commun. 2010, 394, 170–172. [Google Scholar] [CrossRef]
- Ali, E.M. Contributions of some biological activities of honey bee venom. J. Apic. Res. 2014, 53, 441–451. [Google Scholar] [CrossRef]
- Lee, J.; Lee, D.G. Influence of the hydrophobic amino acids in the N- and C-terminal regions of pleurocidin on antifungal activity. J. Microbiol. Biotechnol. 2010, 20, 1192–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, S.; Gupta, A.; Karne, S.P.; Kamble, S.; Shinde, B. Anti-inflammatory activity of sting protein from Apis mellifera. Int. J. Life Sci. Sci. Res. 2017, 3, 914–919. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, W.R.; Kim, K.H.; An, H.J.; Chang, Y.C.; Han, S.M.; Park, Y.Y.; Pak, S.C.; Park, K.K. Effects of bee venom against Propionibacterium acnes-induced inflammation in human keratinocytes and monocytes. Int. J. Mol. Med. 2015, 35, 1651–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.M.; Pak, S.C.; Nicholls, Y.M.; Macfarlane, N. Evaluation of anti-acne property of purified bee venom serum in humans. J. Cosmet. Dermatol. 2016, 15, 324–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diwakar, L.; Ewan, P.; Huber, P.A.J.; Clark, A.; Nasser, S.; Krishna, M.T. The impact of national guidelines on venom immunotherapy practice in the United Kingdom. Clin. Exp. Allergy 2016, 46, 749–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safaeinejad, Z.; Nabiuni, M.; Nazari, Z. Potentiation of a novel palladium (II) complex lethality with bee venom on the human T-cell acute lymphoblastic leukemia cell line (MOLT-4). J. Venom. Anim. Toxins Incl. Trop. Dis. 2013, 19, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alizadehnohi, M.; Nabiuni, M.; Nazari, Z.; Safaeinejad, Z.; Irian, S. The synergistic cytotoxic effect of cisplatin and honey bee venom on human ovarian cancer cell line A2780cp. J. Venom Res. 2012, 3, 22–27. [Google Scholar]
- Trindade, R.A.; Kiyohara, P.K.; De Araujo, P.S.; da Costa Bueno, M.H. PLGA microspheres containing bee venom proteins for preventive immunotherapy. Int. J. Pharm. 2012, 423, 124–133. [Google Scholar] [CrossRef]

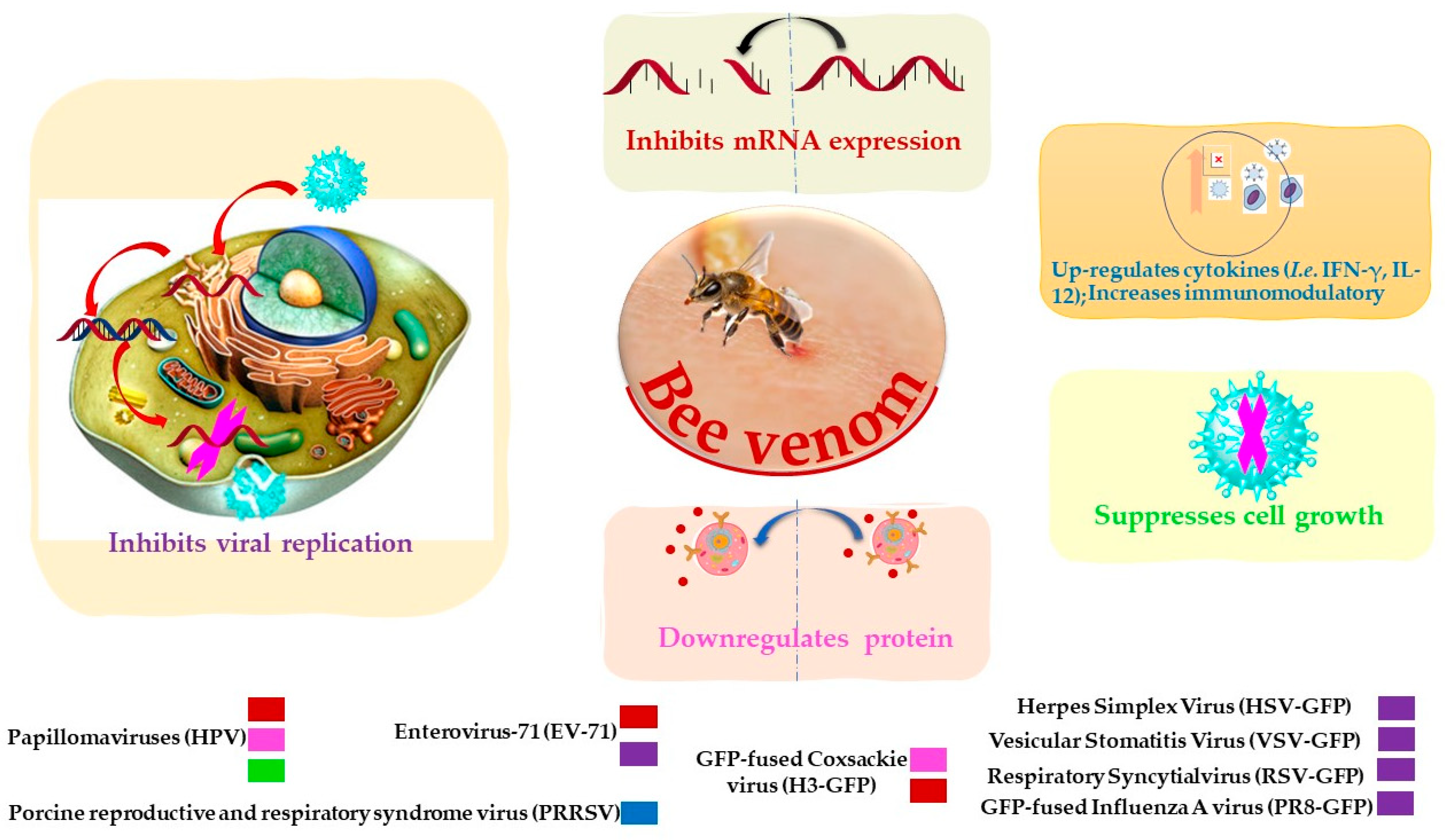

{kind=link}
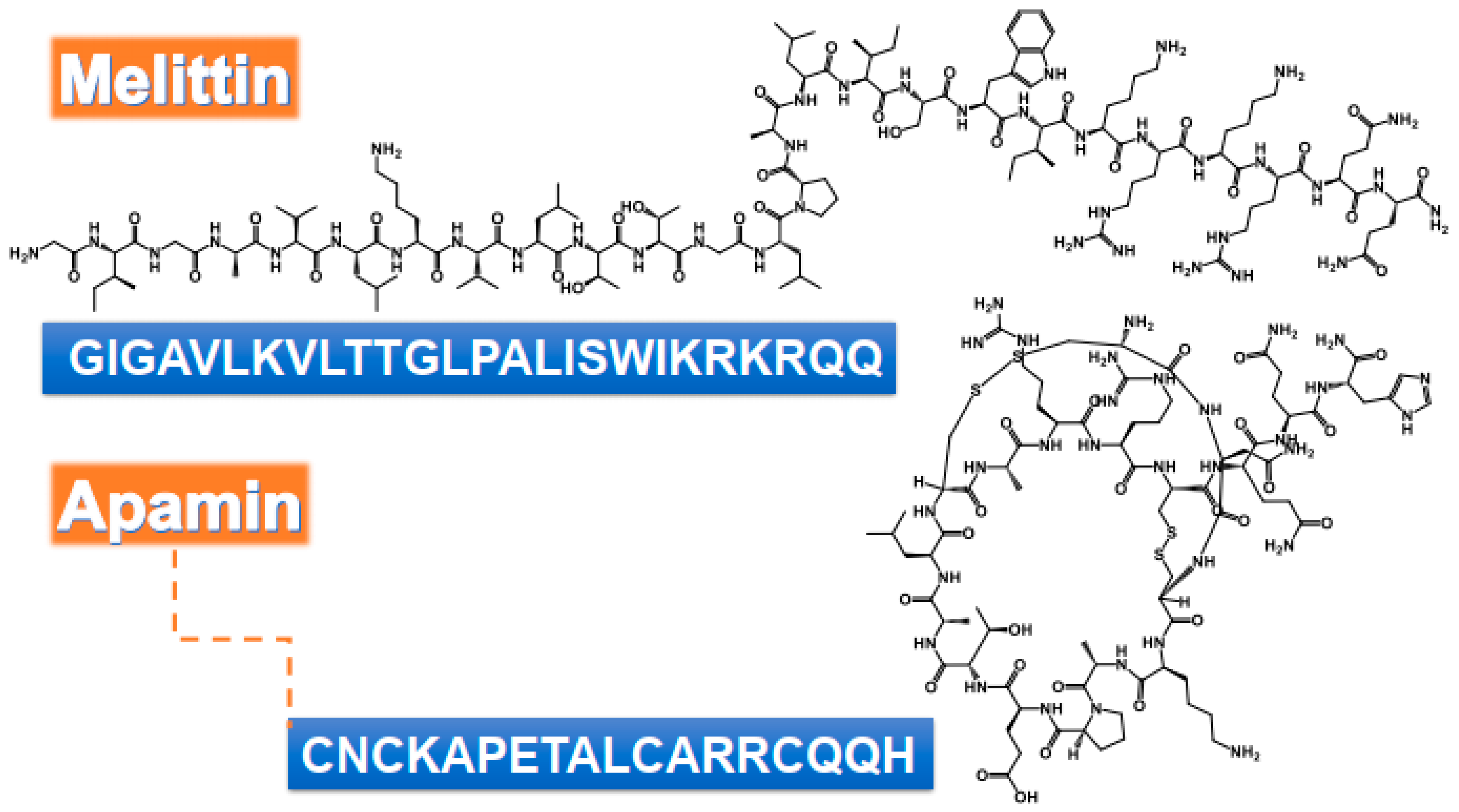{kind=link}
{kind=link}
| Bee Venom/Isolated Compounds | Organism | Method | Dose/Mode of Action | Reference |
|---|---|---|---|---|
| Bee venom | S. aureus | Disc diffusion | MIC 8 µg/mL | [57] |
| MBC 16 µg/mL | ||||
| S. aureus Gp | Disc-diffusion | At 100 µg/mL give inhibition zone 23.2 mm after 24 h | [10] | |
| MRSA CCARM 3366 | Broth microdilution | MIC 0.085 μg/mL | [5]. | |
| MBC 0.106 μg/ mL | ||||
| S. aureus CCARM 3708 | Broth microdilution | MIC 0.11 μg/mL | [5]. | |
| MBC 0.14 μg/mL | ||||
| MR S. aureus ATCC 33591 | Broth microdilution | MIC90% 7.2 μg/mL | [11] | |
| MBC90% 28.7 μg/mL | ||||
| PC: Cephalothin | ||||
| MIC90% 2 μg/mL | ||||
| MBC90% 2 μg/mL | ||||
| S. aureus enterotoxin ATCC 23235 | Broth microdilution | MIC 0.7 μg/mL | [11] | |
| PC: Cephalothin and Oxacillin | ||||
| MIC < 0.5 μg /mL | ||||
| S. hyicus | Disc diffusion | MIC 128 µg/mL | [57] | |
| MBC 128 µg/mL | ||||
| S. chromogenes | Disc diffusion | MIC 128 µg/mL | [57] | |
| MBC 128 µg/mL | ||||
| S. salivarius | Broth microdilution | MIC 20 µg/mL | [4] | |
| PC: Chlorhexidine digluconate | ||||
| MIC 0.9 µg/mL | ||||
| S. sanguinis | Broth microdilution | MIC 30 µg/mL | [4] | |
| PC: Chlorhexidine digluconate | ||||
| MIC 3.7 µg/mL | ||||
| S. sobrinus | Broth microdilution | MIC 40 µg/mL | [4] | |
| PC: Chlorhexidine digluconate | ||||
| MIC 0.9 µg/mL | ||||
| S. mitis | Broth microdilution | MIC 40 µg/mL | [4] | |
| PC: Chlorhexidine digluconate | ||||
| MIC 3.7 µg/mL | ||||
| S. mutans | Broth microdilution | MIC 20 µg/mL | [4] | |
| PC: Chlorhexidine digluconate | ||||
| MIC 0.9 µg/mL | ||||
| Klebsiella pneumonia | Broth microdilution | MIC 30 µg/mL for 24 h | [16] | |
| Bacillus subtilis | Broth microdilution | MIC 30 µg/mL for 24 h | [16] | |
| Paenibacillus larvae | Resazurin method | MIC 3.12 μg/mL | [58] | |
| MBC 4.16 μg/mL | ||||
| PC: Oxytetracycline | ||||
| MIC 0.63 μg/mL | ||||
| MBC 0.83 μg/mL | ||||
| E. faecalis | Broth microdilution | MIC 20 µg/mL | [4] | |
| PC: Chlorhexidine digluconate | ||||
| MIC 3.7 µg/mL | ||||
| L. casei | Broth microdilution | MIC 20 µg/mL | [4] | |
| PC: Chlorhexidine digluconate | ||||
| MIC 0.9 µg/mL | ||||
| Salmonella typhimurium | Disc-diffusion | Inhibition zone was 15.88 mm at 45 μg | [59] | |
| PC: Gentamicin | ||||
| Inhibition zone was 19 mm at 10 μg/mL | ||||
| E. coli | Disc-diffusion | At 45 μg inhibits 32.46 mm | [59] | |
| PC: Gentamicin | ||||
| At 10 μg/mL inhibits 20 mm | ||||
| P. aeruginosa | NR | The antibacterial activity was 38% at 50 µg/mL | [60] | |
| Borrelial spirochetes | Direct counting method | MIC 200 µg/mL | [6] | |
| PC: Doxycycline, cefoperazone, and daptomycin | ||||
| MIC 10 µg/mL | ||||
| Melittin | S. salivarius | Broth microdilution | MIC 10 µg/mL | [4] |
| E. faecalis | Broth microdilution | MIC 6 µg/mL | [4] | |
| L. casei | Broth microdilution | MIC 4 µg/mL | [4] | |
| S. sanguinis | Broth microdilution | MIC 10 µg/mL | [4] | |
| S. sobrinus | Broth microdilution | MIC 10 µg/mL | [4] | |
| S. mitis | Broth microdilution | MIC 10 µg/mL | [4] | |
| S. mutans | Broth microdilution | MIC 40 µg/mL | [4] | |
| K. pneumonia | Broth microdilution | MIC 8 µg/mL throughout 24 h | [16] | |
| B. subtilis | Broth microdilution | MIC 6 µg/mL for 24 h | [16] | |
| Susceptible colistin- A. baumannii | Broth microdilution | MIC 4 mg/L after 24 h | [61] | |
| Acinetobacter spp. | Disc diffusion | Cell lysis | [62] | |
| Membranolytic effect | ||||
| MIC 0.5 µg/mL | ||||
| Colistin-resistant A. baumannii | Broth microdilution | MIC 2 mg/L after 24 h | [61] | |
| Listeria monocytogenes F4244 | Agar well diffusion | MIC 0.315 µg/mL | [53] | |
| MBC 3.263 µg/mL | ||||
| MR S. aureus ATCC 33591 | Broth microdilution | MIC90% 6.7 μg/mL | [11] | |
| MBC90% 26 μg/mL | ||||
| PC: Cephalothin | ||||
| MIC90% 2 μg/mL | ||||
| MBC90% 2 μg/mL | ||||
| S. aureus enterotoxin ATCC 23235 | Broth microdilution | MIC 3.6 μg/mL | [11] | |
| PC: Cephalothin and Oxacillin | ||||
| MIC <0.5 μg /mL | ||||
| S. aureus | Microtiter broth dilution | MIC 6.25 μg/mL | [63] | |
| B. spirochetes | Direct counting method | MIC 200 µg/mL | [6] | |
| PC: Doxycycline, cefoperazone, and daptomycin | ||||
| MIC 10 µg/mL | ||||
| A. baumannii ATCC 19606 | Broth microdilution | MIC 17µg/mL | [64] | |
| PC: Polymyxin | ||||
| MIC 0.25 µg/mL | ||||
| Imipenem: | ||||
| MIC ≤ 0.125 0.25 µg/mL | ||||
| A. baumannii 31852 (S) | Broth microdilution | MIC 20 µg/mL | [64] | |
| PC: Polymyxin | ||||
| MIC 0.25 µg/mL | ||||
| Imipenem: | ||||
| MIC 0.25 µg/mL | ||||
| A. baumannii 33677 (XDR) | Broth microdilution | MIC 31 µg/mL | [64] | |
| PC: Polymyxin | ||||
| MIC 0.25 µg/mL | ||||
| Imipenem: | ||||
| MIC 16 µg/mL | ||||
| A. baumannii 96734 (XDR) | Broth microdilution | MIC 45.5 µg/mL | [64] | |
| PC: Polymyxin | ||||
| MIC 0.25 µg/mL | ||||
| Imipenem: | ||||
| MIC 16 µg/mL | ||||
| Synthetic Melittin and Its Analogues | ||||
| Synthetic melittin | P. aeruginosa ATCC 47085 | Luria broth | MIC 12.1 µM | [65] |
| E. coli ATCC 29222 | Luria broth | MIC 13.2 µM | [65] | |
| E. coli DH5 | NR | MIC 3.9 µM | [66] | |
| PC: Tetracycline | ||||
| MIC 1.2 µM | ||||
| K. pneumoniae ATCC 13883 | Luria broth | MIC 14.9 µM | [65] | |
| A. baumannii ATCC 19606 | Luria broth | MIC 8.3 µM | [65] | |
| B. subtilis | NR | MIC 2 µM | [66] | |
| PC: Tetracycline | ||||
| MIC 0.2 µM | ||||
| S. aureus | NR | MIC 3.6 µM | [66] | |
| PC: Tetracycline | ||||
| MIC 4 µM | ||||
| Melittin Hybrid | ||||
| Cecropin A–melittin (CAM) | E. coli | Microtiter broth dilution | MIC 3.7 µg /mL | [67] |
| CAM-W | E. coli | Microtiter broth dilution | MIC 0.3 µg/mL | [67] |
| Cecropin A-melittin CA(1–8)M(1–18) | A. baumannii | Mueller-Hinton broth | MIC 2 µM | [68] |
| PC: Polymyxin B | ||||
| MIC 1 µM | ||||
| Mutant melittin I17K | L. monocytogenes F4244 | Agar well diffusion | MIC 0.814 µg/mL | [53] |
| MBC 7.412 µg/mL | ||||
| Mutant melittin G1I | L. monocytogenes F4244 | Agar well diffusion | MIC 0.494 µg/mL | [53] |
| MBC 5.366 µg/mL | ||||
| MM-1 | B. subtilis | NR | MIC 2.4 µM | [66] |
| PC: Tetracycline | ||||
| MIC 0.2 µM | ||||
| MM-2 | B. subtilis | NR | MIC 1.8 µM | [66] |
| PC: Tetracycline | ||||
| MIC 0.2 µM | ||||
| Mel-H | E. coli | Microtiter broth dilution | MIC 11.25 µM | [69] |
| P. aeruginosa ATCC27853 | Microtiter broth dilution | MIC 11.25 µM | [69] | |
| S. aureus ATCC25923 | Microtiter broth dilution | MIC 5.6 µM | [69] | |
| Mel(12–24) | B. subtilis | Broth microdilution | MIC 0.65 µg/mL | [70] |
| PC: Melittin | ||||
| MIC 0.18 µg/mL | ||||
| S. aureus | Broth microdilution | MIC 1.3 µg/mL | [70] | |
| PC: melittin | ||||
| MIC 0.72 µg/mL | ||||
| Phospholipase A2 | S. aureus Gp | Disc-diffusion | Hydrolysis of phospholipids | [10] |
| At 100 µg/mL inhibits 13.33 mm after 24 h | ||||
| L. casei | Broth microdilution | MIC 400 µg/mL | [4] | |
| Bee Venom/Isolated Compounds | Organism | Method | Dose/Mode of Action | Reference |
|---|---|---|---|---|
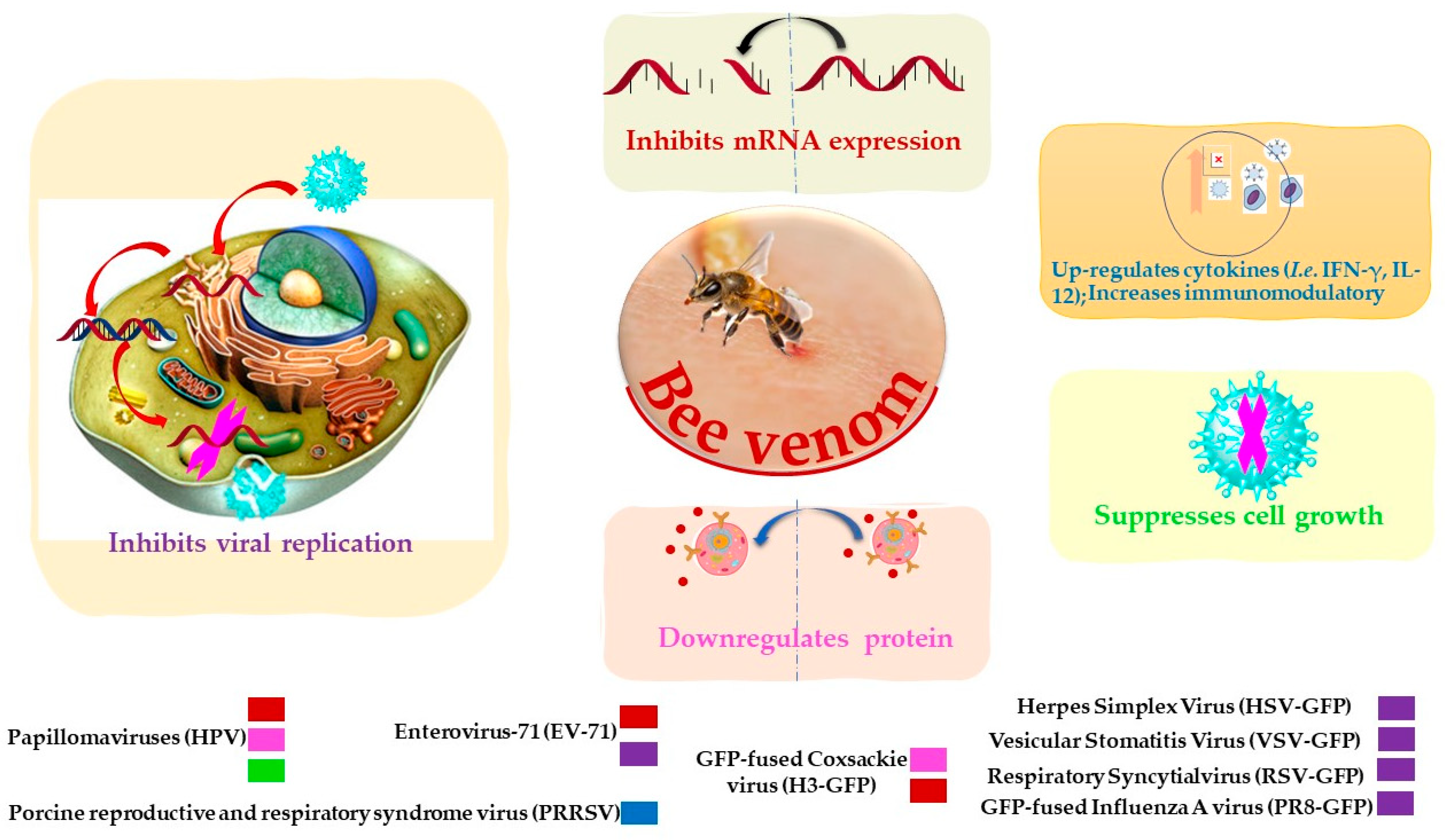
| Bee venom | Papillomaviruses (HPV16 E6) | Reverse transcription assay | Inhibits mRNA expression. | [77] |
| Suppresses cell growth. | ||||
| Downregulates protein. | ||||
| At 10 µg/mL inhibits 0.35 ± 0.06-fold after 24 h. | ||||
| Papillomaviruses (HPV16 E7) | Reverse transcription assay | Inhibits mRNA expression. | [77] | |
| Suppresses cell growth. | ||||
| Downregulates protein. | ||||
| At 10 µg/mL inhibits 0.44 ± 0.07-fold after 24 h. | ||||
| PRRSV | Enzyme-linked immunosorbent assay | Increases immunomodulatory against the virus. | [78] | |
| Significant up-regulate Th1 cytokines (IFN-γ and IL-12) and several types of immune cells. | ||||
| Vesicular stomatitis virus (VSV) | Plaque assay | Inhibits virus replication | [76] | |
| EC50 0.5 ± 0.06 μg/mL | ||||
| HSV | Plaque assay | Inhibits virus replication | [76] | |
| EC50 1.52 ± 0.11 μg/mL | ||||
| Coxsackie virus (H3) | Plaque assay | Inhibits mRNA expression | [76] | |
| Inhibits virus replication | ||||
| EC50 0.5 ± 0.04 μg/mL | ||||
| RSV | Plaque assay | Inhibits virus replication | [76] | |
| EC50 1.17 ± 0.09 μg/mL | ||||
| PR8 | Plaque assay | Inhibits virus replication. | [76] | |
| EC50 1.81 ± 0.08 μg/mL | ||||
| EV-71 | Plaque assay | Inhibits mRNA expression. | [76] | |
| Inhibits virus replication | ||||
| EC50 0.49 ± 0.02 μg/mL | ||||
| Lumpy skin disease virus (LSDV) | Agar gel precipitation test | At the dose 0.5 μg/mL | [85] | |
| Melittin | Immunodeficiency virus (HIV) | Lysis and fusion assays | Lytic and fusogenic | [86] |
| Herpes simplex (HSV-1) | Plaque assay Virus penetration assay | Inhibits cell fusion. | [83] | |
| Inhibits Na+, K+ pump activity. | ||||
| Inhibits virus adsorption and penetration to the cells. | ||||
| Immunodeficiency virus HIV-1 | Transient transfection Assays | Inhibits virus replication. | [84] | |
| Suppresses gene expression. | ||||
| Suppresses intracellular | ||||
| Protein and mRNA synthesis. | ||||
| Suppresses long terminal repeat (LTR) activity | ||||
| ID50 0.9–1.4 µM after 24 h. | ||||
| Arenavirus Junin (JV) | Plaque assay | Impedes the multiplication | [82] | |
| EC50 0.86 µM after 24 h. | ||||
| HSV-1 | Plaque assay | Impedes the multiplication | [82] | |
| EC50 1.35 µM after 24 h. | ||||
| Herpes simplex virus (HSV-2) | Plaque assay | Impedes the multiplication | [82] | |
| EC50 2.05 µM after 24 h. | ||||
| Herpes simplex virus 1 M (HSV-1 M) | Quantitative microplate assay | Viral inactivation at 100 µg/mL | [87] | |
| Herpes simplex virus 2 G (HSV-2 G) | Quantitative microplate assay | Viral inactivation at 100 µg/mL | [87] | |
| Phospholipase A2 (sPLA2) | Hepatitis C virus (HCV) | Plaque assay | IC50 117 ± 43 ng/mL after 24 h. | [81] |
| DENV | Plaque assay | IC50 183 ± 38 ng/mL after 24 h. | [81] | |
| JEV | Plaque assay | IC50 49 ± 13 ng/mL after 24 h. | [81] |
| Bee Venom/ Isolated Compounds | Organism | Method | Dose/Mode of Action | Reference |
|---|---|---|---|---|
| Bee venom | T. mentagrophytes | Broth dilution | At 0.63 ppm inhibits 92% After 1 h. | [89] |
| T. rubrum | Broth dilution | At 0.63 ppm inhibits 26% After 1 h. | [89] | |
| C. albicans | Disc diffusion | Prevents dimorphism MIC 40 µg/mL for 48 h. | [92] | |
| C. albicans | Broth microdilution | MIC 62.5–125 μg/mL for 24 h. | [88] | |
| Candida krusei | Broth microdilution | MIC 60 µg/mL throughout 48 h. | [16] | |
| A. alternate | NR | At 1 µg/mL inhibits 50% of interleukin (IL)-6 production. | [3] | |
| At 1 µg/mL inhibits 28.8% of interleukin (IL)-8 production. | [3] | |||
| Melittin | C. krusei | Broth microdilution | MIC 30 µg/mL for 48 h | [16] |
| C. albicans | NR | Disruptive the mitochondrial membrane. | [90] | |
| Apoptotic for 4 h | ||||
| Aspergillus flavus (KCTC 1375) | Microdilution method and MTT assay | MIC 1.25 µM | [93] | |
| PC: Amphotericin B: | ||||
| MIC 2.5 µM | ||||
| Fluconazole: MIC 10 µM | ||||
| Itraconazole: MIC 10 µM | ||||
| Malassezia furfur (KCTC 7744) | Microdilution method and MTT assay | MIC 1.25 µM | [93] | |
| PC: Amphotericin B: MIC 2.5 µM | ||||
| Fluconazole: MIC 5 µM | ||||
| Itraconazole: MIC 5 µM | ||||
| C. albicans (ATCC 90028) | Microdilution method and MTT assay | MIC 2.5 µM | [93] | |
| PC: Amphotericin B: MIC 5 µM | ||||
| Fluconazole: MIC 10 µM | ||||
| Itraconazole: MIC 10 µM | ||||
| Apamin | A. alternate | NR | At 1 µg/mL inhibits 42.6% of interleukin (IL)-6 production. | [3] |
| At 1 µg/mL inhibits 38.7% of interleukin (IL)-8 production. | [3] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Seedi, H.; Abd El-Wahed, A.; Yosri, N.; Musharraf, S.G.; Chen, L.; Moustafa, M.; Zou, X.; Al-Mousawi, S.; Guo, Z.; Khatib, A.; et al. Antimicrobial Properties of Apis mellifera’s Bee Venom. Toxins 2020, 12, 451. https://doi.org/10.3390/toxins12070451
El-Seedi H, Abd El-Wahed A, Yosri N, Musharraf SG, Chen L, Moustafa M, Zou X, Al-Mousawi S, Guo Z, Khatib A, et al. Antimicrobial Properties of Apis mellifera’s Bee Venom. Toxins. 2020; 12(7):451. https://doi.org/10.3390/toxins12070451
Chicago/Turabian StyleEl-Seedi, Hesham, Aida Abd El-Wahed, Nermeen Yosri, Syed Ghulam Musharraf, Lei Chen, Moustafa Moustafa, Xiaobo Zou, Saleh Al-Mousawi, Zhiming Guo, Alfi Khatib, and et al. 2020. "Antimicrobial Properties of Apis mellifera’s Bee Venom" Toxins 12, no. 7: 451. https://doi.org/10.3390/toxins12070451
APA StyleEl-Seedi, H., Abd El-Wahed, A., Yosri, N., Musharraf, S. G., Chen, L., Moustafa, M., Zou, X., Al-Mousawi, S., Guo, Z., Khatib, A., & Khalifa, S. (2020). Antimicrobial Properties of Apis mellifera’s Bee Venom. Toxins, 12(7), 451. https://doi.org/10.3390/toxins12070451