Antioxidant and Cytoprotective Properties of Cyanobacteria: Potential for Biotechnological Applications



 and
and
Abstract
:1. Introduction
2. Results
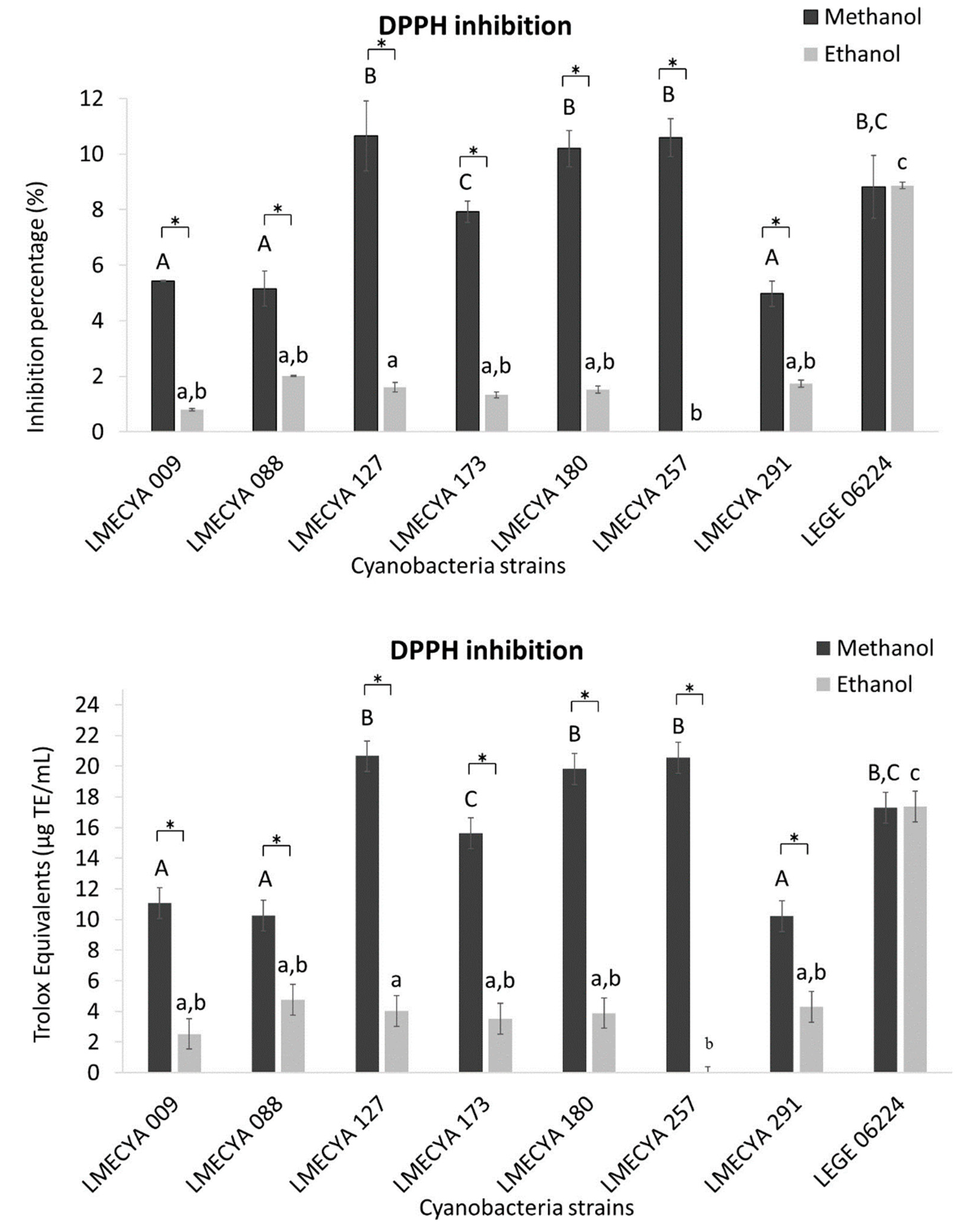
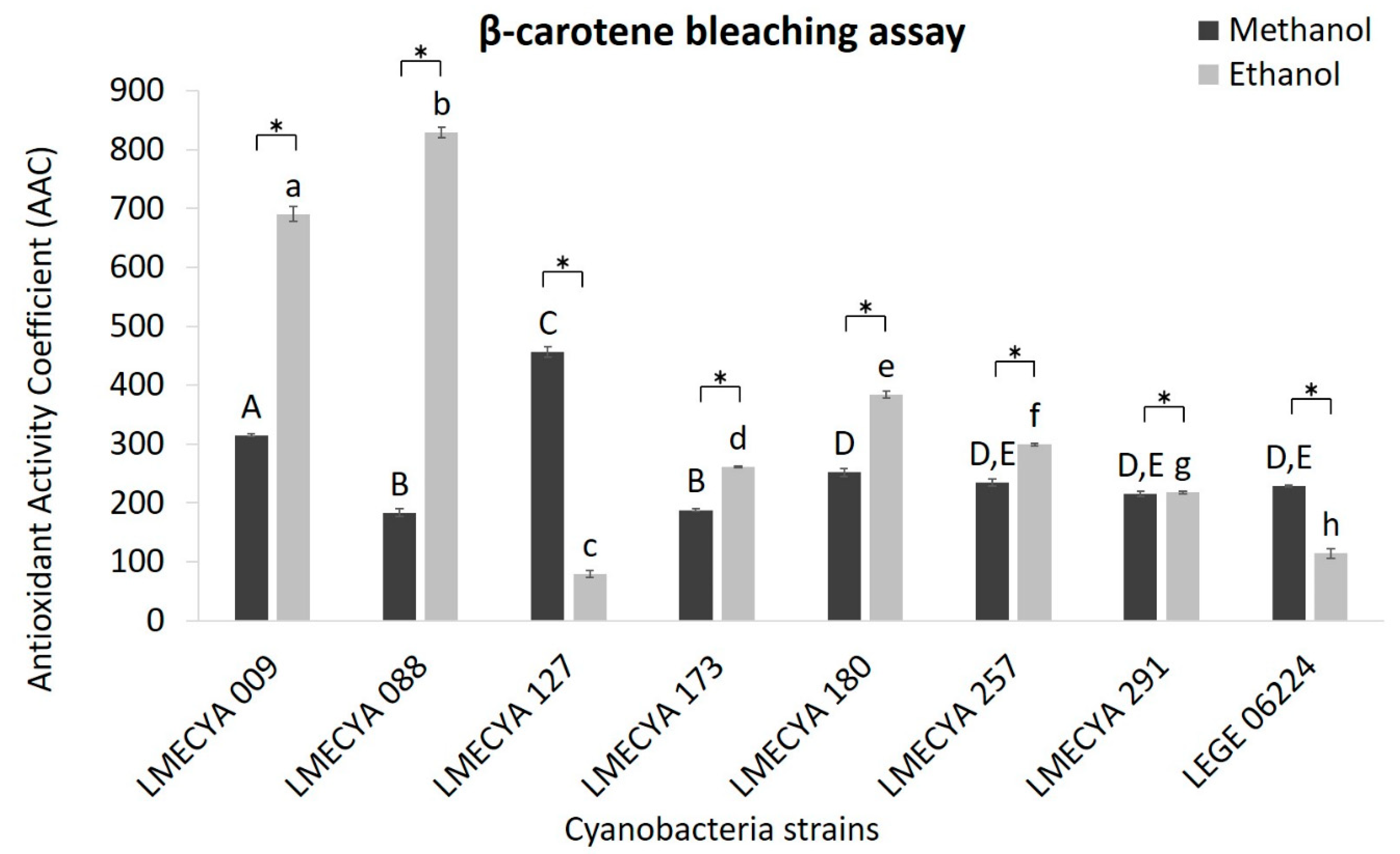
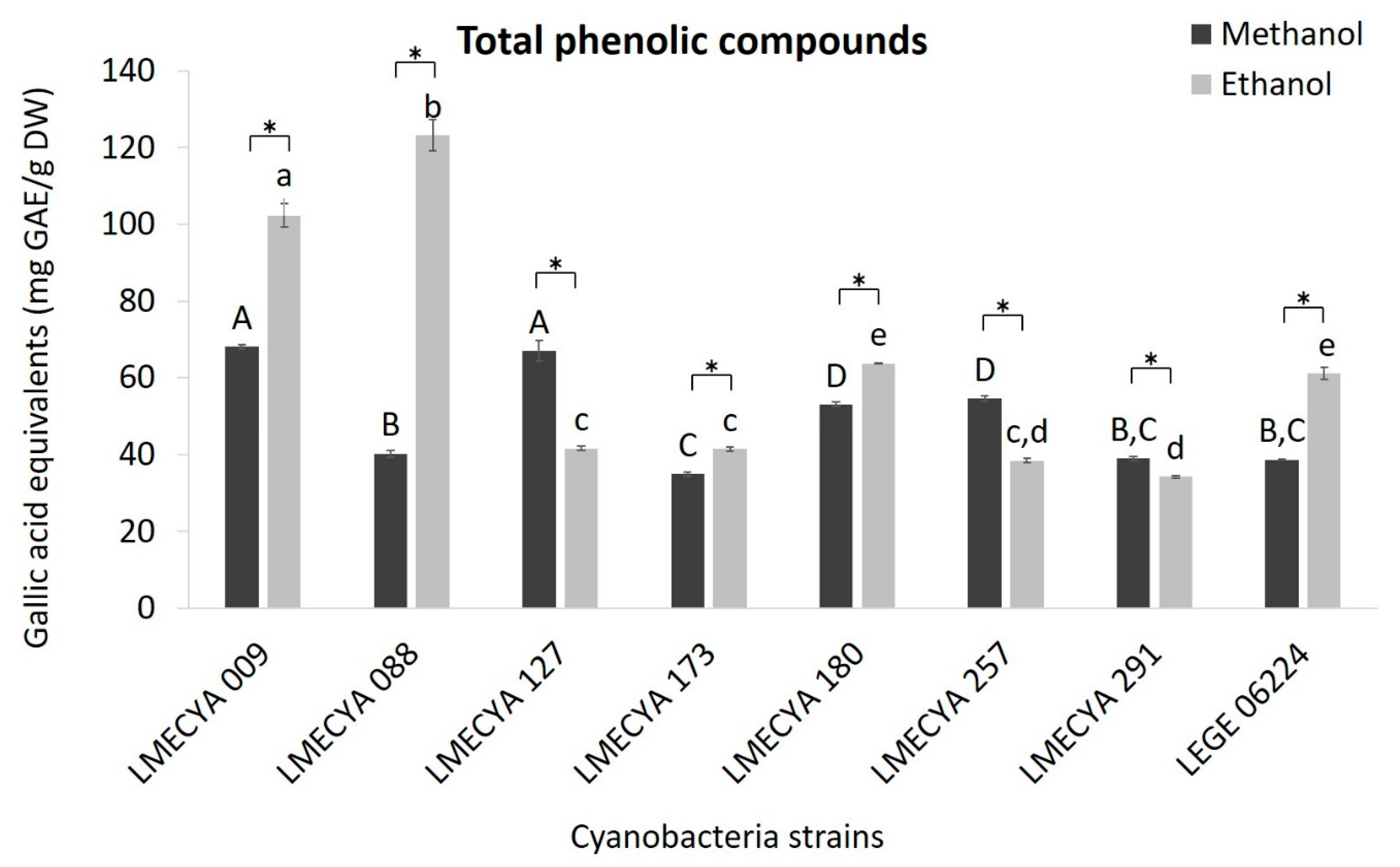
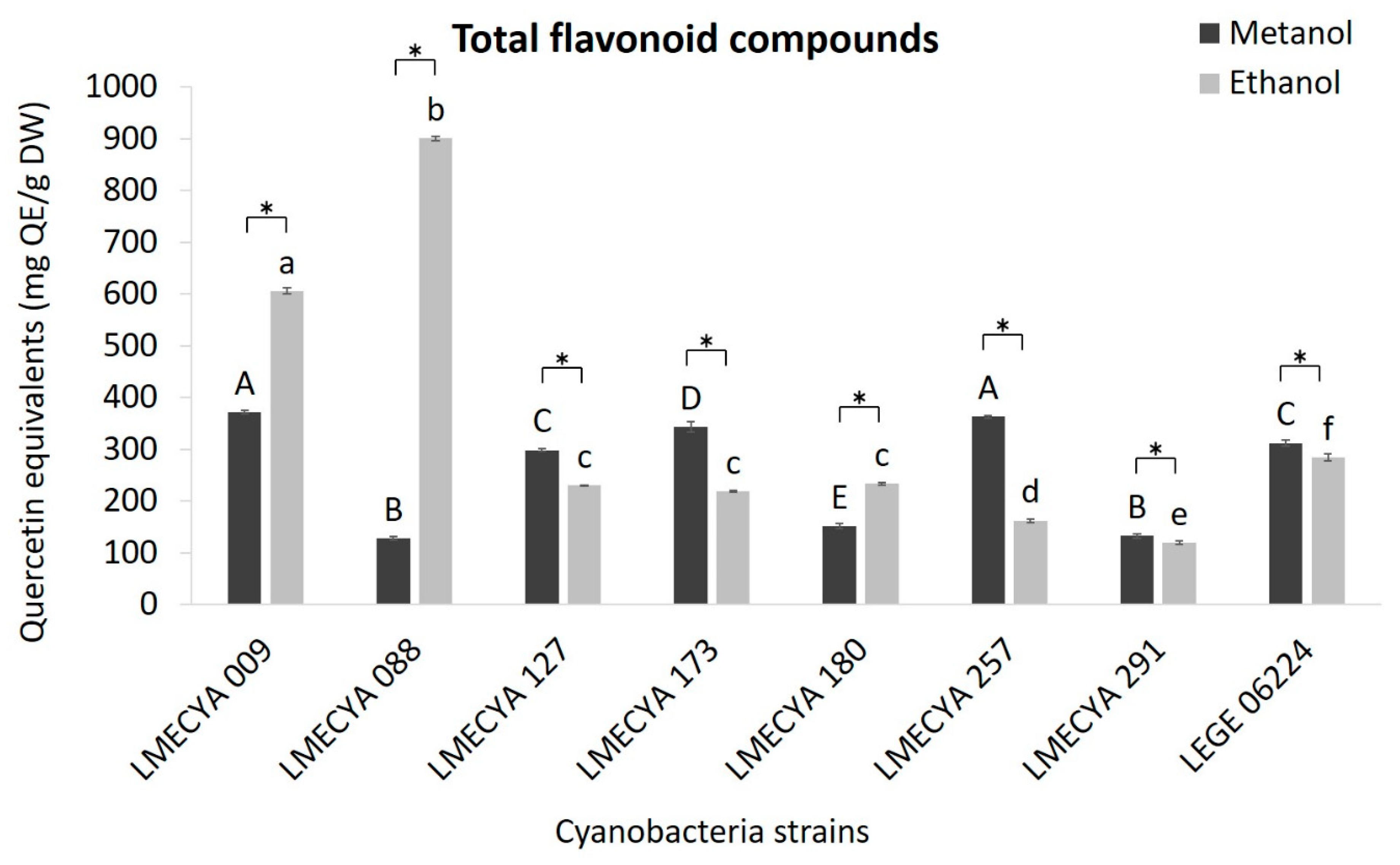
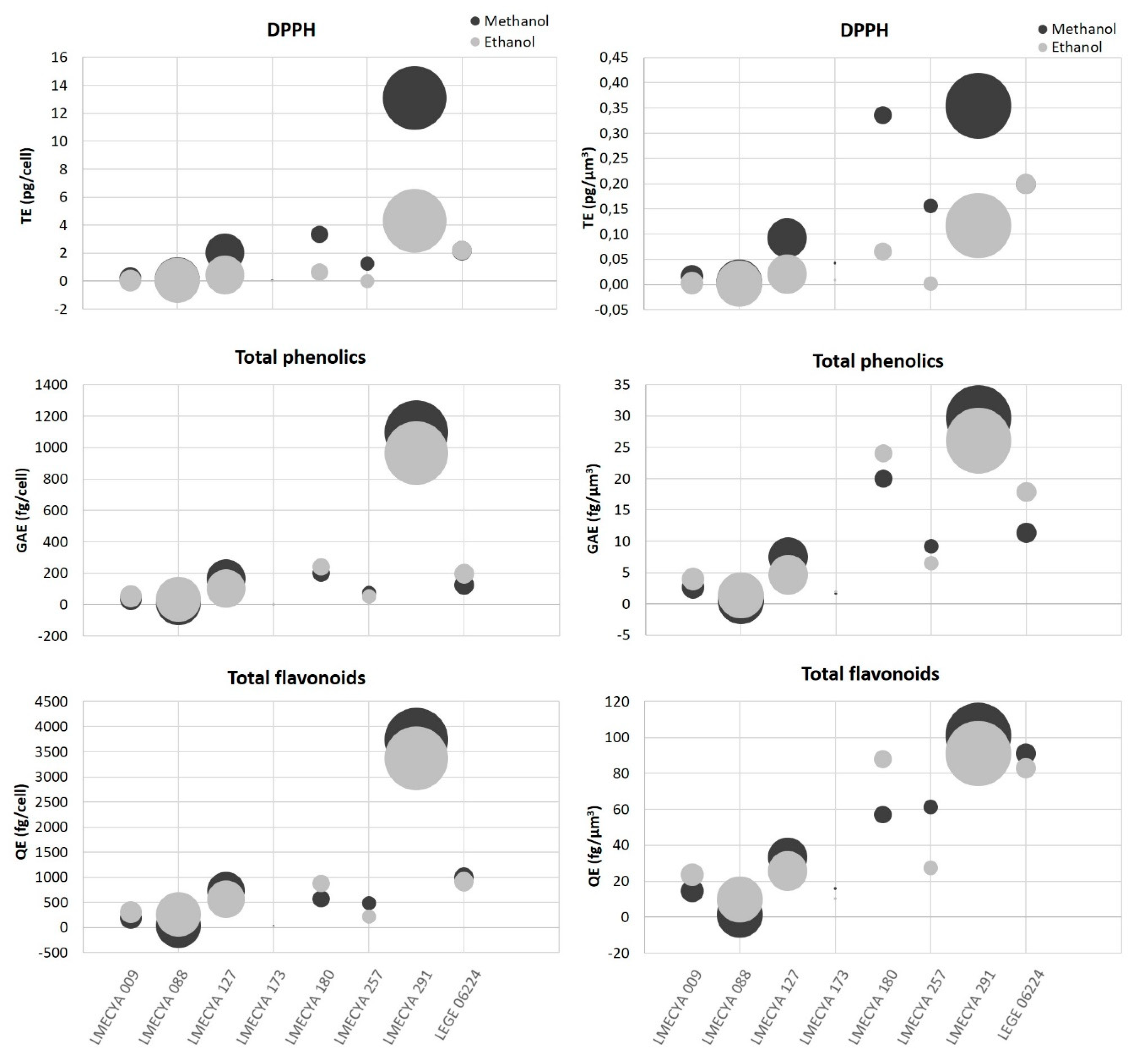
2.1. Antioxidant Profile of Cyanobacterial Strains
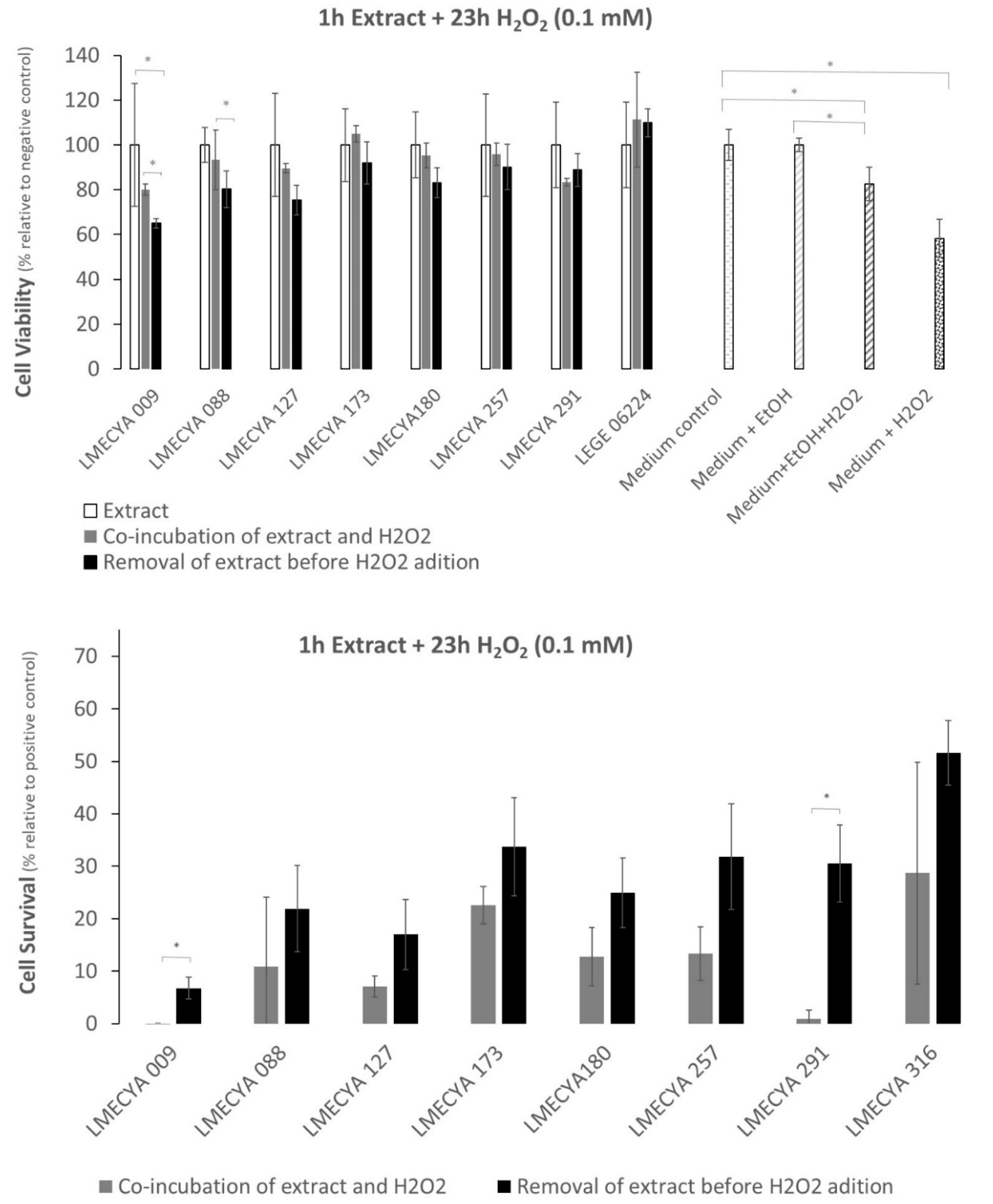
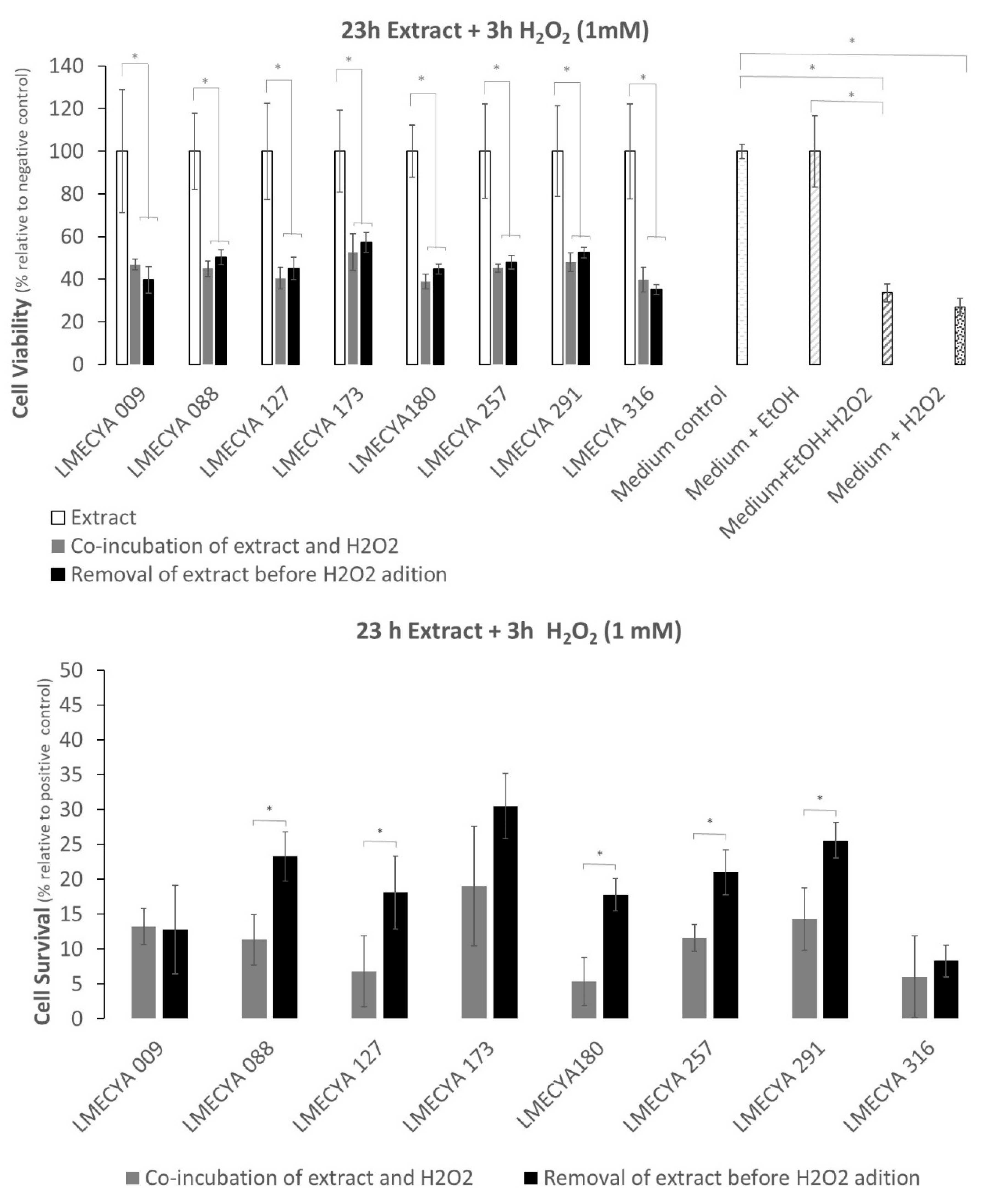
2.2. Protective Effect of Cyanobacterial Extracts on HEK293T Cell Line against H2O2-Induced Cytotoxicity
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Cyanobacterial Strains Maintenance and Biomass Production
5.3. Preparation of Cyanobacterial Extracts
5.4. Evaluation of Antioxidant Potential of Cyanobacterial Extract by Chemical Methods
5.4.1. DPPH Scavenging Method
5.4.2. β-carotene Bleaching Assay
5.4.3. Determination of Total Phenolic Compounds
5.4.4. Determination of Total Flavonoid Compounds
5.5. Evaluation of the Protective Effect of Cyanobacterial Extracts against Hydrogen Peroxide Cytotoxicity in HEK293T Cell Line
5.5.1. HEK293T Cell Line Maintenance and Exposure to Cyanobacteria Extracts and Hydrogen Peroxide
5.5.2. MTT Assay
5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dvořák, P.; Casamatta, D.A.; Hašler, P.; Jahodářová, E.; Norwich, A.R.; Poulíčková, A. Diversity of the Cyanobacteria. In Modern Topics in the Phototrophic Prokaryotes; Hallenbeck, P., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 3–46. [Google Scholar] [CrossRef]
- Dvořák, P.; Poulíčková, A.; Hašler, P.; Belli, M.; Casamatta, D.A.; Papini, A. Species concepts and speciation factors in cyanobacteria, with connection to the problems of diversity and classification. Biodivers. Conserv. 2015, 24, 739–757. [Google Scholar] [CrossRef] [Green Version]
- Ehling-Schulz, M.; Scherer, S. UV protection in cyanobacteria. Eur. J. Phycol. 1999, 34, 329–338. [Google Scholar] [CrossRef]
- Wada, N.; Sakamoto, T.; Matsugo, S. Mycosporine-Like Amino Acids and Their Derivatives as Natural Antioxidants. Antioxidants 2015, 4, 603–646. [Google Scholar] [CrossRef]
- Martel, I.J.; Poza, S.G.; Martel, G.R.; Rico, M.; Olivares, C.A.; Pinchetti, J.L.G. Phenolic Profile and Antioxidant Activity of Crude Extracts from Microalgae and Cyanobacteria Strains. J. Food Qual. 2017, 2017, 2924508. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.P.; Prabha, R.; Verma, S.; Meena, K.K.; Yandigeri, M. Antioxidant properties and polyphenolic content in terrestrial cyanobacteria. 3 Biotech 2017, 7, 134. [Google Scholar] [CrossRef] [Green Version]
- Babić, O.; Kovač, D.; Rašeta, M.; Šibul, F.; Svirčev, Z.; Simeunović, J. Evaluation of antioxidant activity and phenolic profile of filamentous terrestrial cyanobacterial strains isolated from forest ecosystem. J. Appl. Phycol. 2016, 28, 2333–2342. [Google Scholar] [CrossRef]
- Morone, J.; Alfeus, A.; Vasconcelos, V.; Martins, R. Revealing the potential of cyanobacteria in cosmetics and cosmeceuticals—A new bioactive approach. Algal Res. 2019, 41, 101541. [Google Scholar] [CrossRef]
- Singh, R.; Parihar, P.; Singh, M.; Bajguz, A.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Uncovering Potential Applications of Cyanobacteria and Algal Metabolites in Biology, Agriculture and Medicine: Current Status and Future Prospects. Front. Microbiol. 2017, 8, 515. [Google Scholar] [CrossRef] [Green Version]
- Andrade, M.A.; Lima, V.; Sanches-Silva, A.; Vilarinho, F.; Castilho, M.C.; Khwaldia, K.; Ramos, F. Pomegranate and grape by-products and their active compounds: Are they a valuable source for food applications? Trends Food Sci. Technol. 2019, 86, 68–84. [Google Scholar] [CrossRef]
- Yasin, D.; Zafaryab, M.; Ansari, S.; Ahmad, N.; Khan, N.F.; Zaki, A.; Rizvi, M.M.A.; Fatma, T. Evaluation of antioxidant and anti-proliferative efficacy of Nostoc muscorum NCCU-442. Biocatal. Agric. Biotechnol. 2019, 17, 284–293. [Google Scholar] [CrossRef]
- Bavini, J.M. Cyanobacteria in Skin Protection. Master’s Thesis, Faculdade de Ciências Universidade do Porto, Porto, Portugal, 2018. Available online: https://repositorio-aberto.up.pt/bitstream/10216/118610/2/311491.pdf (accessed on 30 April 2020).
- Geethu, V.; Shamina, M. Antioxidant activity of cyanobacterium Nostoc spongiaeforme C. Agardh ex Born. & Flah. J. Algal Biomass Util. 2018, 9, 26–30. [Google Scholar]
- Hossain, M.F.; Ratnayake, R.R.; Meerajini, K.; Kumara, K.L.W. Antioxidant properties in some selected cyanobacteria isolated from freshwater bodies of Sri Lanka. Food Sci. Nutr. 2016, 4, 753–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajishamol, M.P.; Lekshmi, S.; Vijayalakshmy, K.C.; Saramma, A.V. Antioxidant activity of Cyanobacteria isolated from Cochin estuary. Indian J. Geo Mar. Sci. 2016, 45, 974–977. [Google Scholar]
- El-Aty, A.M.A.; Mohame, A.A.; Samhan, F.A. In vitro antioxidant and antibacterial activities of two fresh water Cyanobacterial species, Oscillatoria agardhii and Anabaena sphaerica. J. Appl. Pharm. Sci. 2014, 4, 69–75. [Google Scholar] [CrossRef]
- Sharathchandra, K.; Rajashekhar, M. Antioxidant activity in the four species of cyanobacteria isolated from a sulfur spring in the western ghats of karnataka. Int. J. Pharma Bio Sci. 2013, 4, 275–285. [Google Scholar]
- Li, H.B.; Cheng, K.W.; Wong, C.C.; Fan, K.W.; Chen, F.; Jiang, Y. Evaluation of antioxidant capacity and total phenolic content of different fractions of selected microalgae. Food Chem. 2007, 102, 771–776. [Google Scholar] [CrossRef]
- Singh, D.P.; Prabha, R.; Meena, K.K.; Sharma, L.S.; Sharma, A.K. Induced Accumulation of Polyphenolics and Flavonoids in Cyanobacteria under Salt Stress Protects Organisms through Enhanced Antioxidant Activity. Am. J. Plant Sci. 2014, 5, 726–735. [Google Scholar] [CrossRef]
- Zeeshan, M.; Suhail, S.; Biswas, D.; Farooqui, A.; Arif, J.M. Screening of Selected Cyanobacterial Strains for Phycochemical Compounds and Biological Activities in Vitro. Biochem. Cell. Arch. 2010, 10, 163–168. [Google Scholar]
- Hashtroudi, M.S.; Shariatmadari, Z.; Riahi, H.; Ghassempour, A. Analysis of Anabaena vaginicola and Nostoc calcicola from Northern Iran, as rich sources of major carotenoids. Food Chem. 2013, 136, 1148–1153. [Google Scholar] [CrossRef]
- Osuna-Ruiz, I.; López-Saiz, C.M.; Burgos-Hernández, A.; Velázquez, C.; Nieves-Soto, M.; Hurtado-Oliva, M.A. Antioxidant, antimutagenic and antiproliferative activities in selected seaweed species from Sinaloa, Mexico. Pharm. Biol. 2016, 54, 2196–2210. [Google Scholar] [CrossRef]
- Raja, R.; Hemaiswarya, S.; Arunkumar, K.; Carvalho, I.S. Antioxidant activity and lipid profile of three seaweeds of Faro, Portugal. Rev. Bras. Bot. 2016, 39, 9–17. [Google Scholar] [CrossRef]
- Chernane, H.; Mansori, M.; Latique, S.; El Kaoua, M. Evaluation of Antioxidant Capacity of Methanol Extract and Its Solvent Fractions Obtained From Four Moroccan Macro Algae Species. Eur. Sci. J. 2014, 10, 35–48. [Google Scholar] [CrossRef]
- Martins, C.; Vilarinho, F.; Sanches Silva, A.; Andrade, M.; Machado, A.V.; Castilho, M.C.; Sá, A.; Cunha, A.; Vaz, M.F.; Ramos, F. Active polylactic acid film incorporated with green tea extract: Development, characterization and effectiveness. Ind. Crops Prod. 2018, 123, 100–110. [Google Scholar] [CrossRef] [Green Version]
- Kedare, S.B.; Singh, R.P. Genesis and development of DPPH method of antioxidant assay. J. Food Sci. Technol. 2011, 48, 412–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Drug Administration (FDA). Substances GRAS in Food. In 21 CFR 182; Code of Federal Regulations; USA Federal Government: Washington, DC, USA, 2019. [Google Scholar]
- Martins, C.D.L.; Ramlov, F.; Carneiro, N.P.N.; Gestinari, L.M.; Santos, B.F.; Bento, L.M.; Lhullier, C.; Gouvea, L.; Bastos, E.; Horta, P.A.; et al. Antioxidant properties and total phenolic contents of some tropical seaweeds of the Brazilian coast. J. App. Phycol. 2013, 25, 1179–1187. [Google Scholar] [CrossRef]
- Lopes, G.; Clarinha, D.; Vasconcelos, V. Carotenoids from Cyanobacteria: A Biotechnological Approach for the Topical Treatment of Psoriasis. Microorganisms 2020, 8, 302. [Google Scholar] [CrossRef] [Green Version]
- Lakatos, M.; Bilger, W.; Büdel, B. Carotenoid composition of terrestrial cyanobacteria: Response to natural light conditions in open rock habitats in Venezuela. Eur. J. Phycol. 2001, 36, 367–375. [Google Scholar] [CrossRef]
- Palinska, K.A.; Deventer, B.; Hariri, K.; Lotocka, M. A taxonomic study on Phormidium–group (cyanobacteria) based on morphology, pigments, RAPD molecular markers and RFLP analysis of the 16S rRNA gene fragment. Fottea 2011, 11, 41–55. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Garcia, I.; Guil-Guerrero, J.L. Evaluation of the antioxidant activity of three microalgal species for use as dietary supplements and in the preservation of foods. Food Chem. 2008, 108, 1023–1026. [Google Scholar] [CrossRef]
- Aydaş, S.B.; Ozturk, S.; Aslım, B. Phenylalanine ammonia lyase (PAL) enzyme activity and antioxidant properties of some cyanobacteria isolates. Food Chem. 2013, 136, 164–169. [Google Scholar] [CrossRef]
- Latifi, A.; Ruiz, M.; Zhang, C.C. Oxidative stress in cyanobacteria. FEMS Microbiol. Rev. 2009, 3, 258–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ručová, D.; Goga, M.; Matik, M.; Bačkor, M. Adaptations of cyanobacterium Nostoc commune to environmental stress: Comparison of morphological and physiological markers between European and Antarctic populations after rehydration. Czech Polar Rep. 2018, 8, 84–93. [Google Scholar] [CrossRef]
- Bian, Y.Y.; Guo, J.; Majeed, H.; Zhu, K.-X.; Guo, X.-N.; Peng, W.; Zhou, H.-M. Ferulic acid renders protection to HEK293 cells against oxidative damage and apoptosis induced by hydrogen peroxide. In Vitro Cell. Dev. Biol. Anim. 2015, 51, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Grauzdytė, D.; Pukalskas, A.; Viranaicken, W.; El Kalamouni, C.; Venskutonis, P.R. Protective effects of Phyllanthus phillyreifolius extracts against hydrogen peroxide induced oxidative stress in HEK293 cells. PLoS ONE 2018, 13, e0207672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badr, O.A.M.; EL-Shawaf, I.I.S.; El-Garhy, H.A.S.; Moustafa, M.M.A.; Ahmed-Farid, O.A. The potent therapeutic effect of novel cyanobacterial isolates against oxidative stress damage in redox rats. J. Appl. Microbiol. 2019, 126, 1278–1289. [Google Scholar] [CrossRef]
- Skulberg, R.; Skulberg, O.M. Forskning med algekulturer NIVAs kultursampling av alger. In Research with Algal Cultures; NIVA’s Culture Collection of Algae; Norsk Institutt for Vannforskning: Oslo, Norway, 1990. [Google Scholar]
- Le Gresley, M.; McDermott, G. Counting chamber methods for quantitative phytoplankton analysis haemocytometer, Palmer-Maloney cell and Sedgewick-Rafter cell. In Microscopic and Molecular Methods for Quantitative Phytoplankton Analyses; Karlson, B., Cusack, C., Bresnan, E., Eds.; IOC/UNESCO: Paris, France, 2010; pp. 25–27. [Google Scholar]
- Andrade, M.A.; Sanches-Silva, A.; Bonito, M.C.C.; Saraiva, M.; Ribeiro-Santos, R. Characterization of rosemary and thyme extracts for incorporation into a whey protein-based film. LWT Food Sci. Technol. 2018, 92, 497–508. [Google Scholar] [CrossRef]
- Kuskoski, E.M.; Asuero, A.G.; Morales, M.T.; Fett, R. Frutos tropicais silvestres e polpas de frutas congeladas: Atividade antioxidante, polifenóis e antocianinas. Ciência Rural 2006, 36, 1283–1287. [Google Scholar] [CrossRef] [Green Version]
- Erkan, N.; Ayranci, G.; Ayranci, E. Antioxidant activities of rosemary (Rosmarinus Officinalis L.) extract, blackseed (Nigella sativa L.) essential oil, carnosic acid, rosmarinic acid and sesamol. Food Chem. 2008, 110, 76–82. [Google Scholar] [CrossRef]
- Yoo, K.M.; Lee, C.H.; Lee, H.; Moon, B.; Lee, C.Y. Relative antioxidant and cytoprotective activities of common herbs. Food Chem. 2008, 106, 929–936. [Google Scholar] [CrossRef]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. Curr. Protoc. Immunol. 2001, 21, A–3B. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cyanobacterial Species | Strain Code | Strain Origin |
|---|---|---|
| Freshwater strains (ESSACC collection) | ||
| Dolichospermum flos-aquae | LMECYA 180 | Guadiana River/1999 |
| Aphanizomenon flos-aquae | LMECYA 088 | Montargil Reservoir/1999 |
| Aphanizomenon gracile | LMECYA 009 | Peneireiro Reservoir1996 |
| Leptolyngbya sp. | LMECYA 173 | Hydrothermal fountain /2003 |
| Microcystis aeruginosa | LMECYA 127 | Montargil reservoir/2000 |
| Nostoc sp. | LMECYA 291 | Garden soil /2013 |
| Planktothrix agardhii | LMECYA 257 | São Domingos Reservoir /2009 |
| WWTP strain (LEGE collection) | ||
| Planktothrix mougeotii | LEGE 06224 | Febros WWTP /2006 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerreiro, A.; Andrade, M.A.; Menezes, C.; Vilarinho, F.; Dias, E. Antioxidant and Cytoprotective Properties of Cyanobacteria: Potential for Biotechnological Applications. Toxins 2020, 12, 548. https://doi.org/10.3390/toxins12090548
Guerreiro A, Andrade MA, Menezes C, Vilarinho F, Dias E. Antioxidant and Cytoprotective Properties of Cyanobacteria: Potential for Biotechnological Applications. Toxins. 2020; 12(9):548. https://doi.org/10.3390/toxins12090548
Chicago/Turabian StyleGuerreiro, Adriana, Mariana A. Andrade, Carina Menezes, Fernanda Vilarinho, and Elsa Dias. 2020. "Antioxidant and Cytoprotective Properties of Cyanobacteria: Potential for Biotechnological Applications" Toxins 12, no. 9: 548. https://doi.org/10.3390/toxins12090548
APA StyleGuerreiro, A., Andrade, M. A., Menezes, C., Vilarinho, F., & Dias, E. (2020). Antioxidant and Cytoprotective Properties of Cyanobacteria: Potential for Biotechnological Applications. Toxins, 12(9), 548. https://doi.org/10.3390/toxins12090548