Combination of Extrusion and Fermentation with Lactobacillus plantarum and L. uvarum Strains for Improving the Safety Characteristics of Wheat Bran

 ,
,  ,
, 
 ,
, 
 and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Acidity and Microbiological Parameters of Wheat Bran
2.2. Influence of the Different Treatments on Fructose, Glucose, Sucrose, and Maltose Concentration in Processed Wheat Bran
2.3. Free Amino Acid and Biogenic Amine Content in Wheat Bran Samples
2.4. Influence of the Different Treatments on Mycotoxin Concentration in Wheat Bran
3. Conclusions
4. Materials and Methods
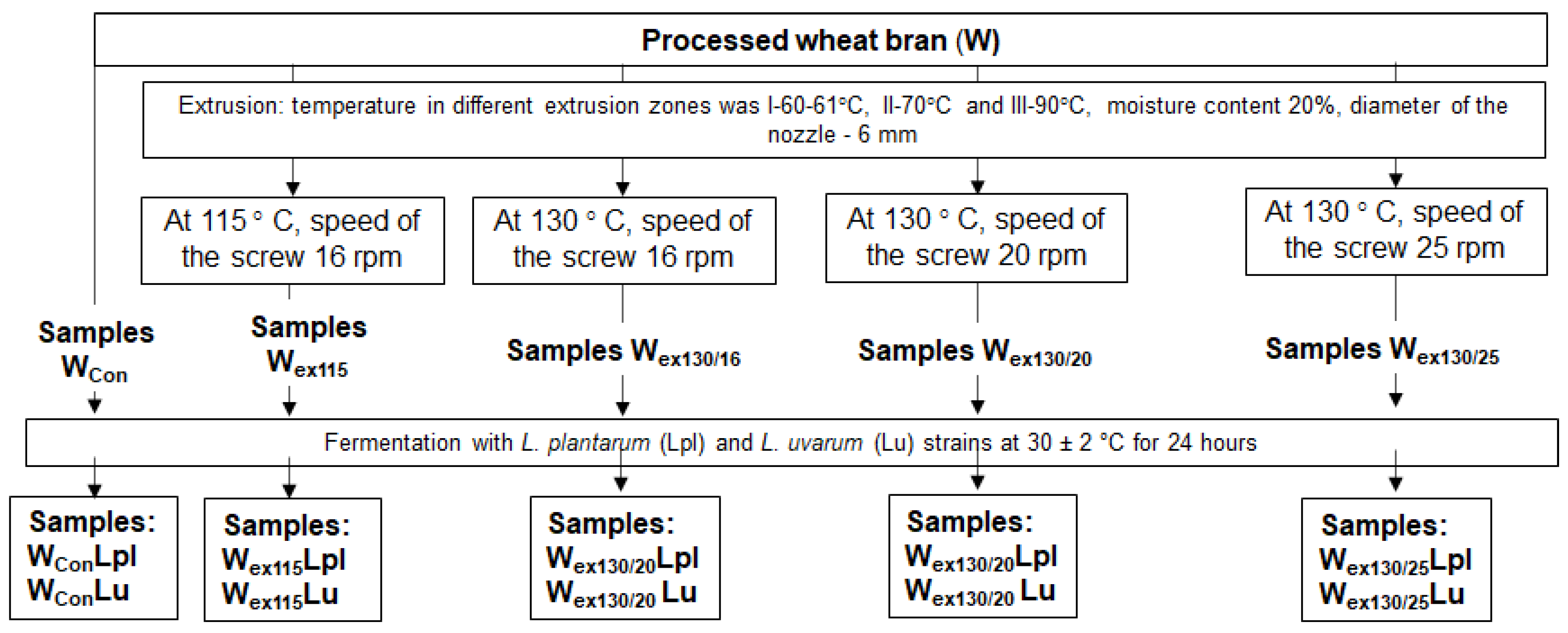
4.1. Processing of Wheat Bran
4.2. Evaluation of Acidity and Microbiological Characteristics of the Wheat Bran Samples
4.3. Analysis of the Sugars in Processed Wheat Bran
4.4. Determination of Free Amino Acids and Biogenic Amines Content in Wheat Bran Samples
4.5. High-Performance Liquid Chromatography Coupled to Triple Quadrupole Mass Spectrometry (HPLC- MS/MS) for Mycotoxin Analysis
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prueckler, M.; Siebenhandl-Ehn, S.; Apprich, S.; Hoeltinger, S.; Haas, C.; Schmid, E.; Kneifel, W. Wheat Bran-Based Biorefinery 1: Composition of Wheat Bran and Strategies of Functionalization. LWT-Food Sci. Technol. 2014, 56, 211–221. [Google Scholar] [CrossRef]
- Soukoulis, C.; Aprea, E. Cereal Bran Fractionation: Processing Techniques for the Recovery of Functional Components and their Applications to the Food Industry. Recent Pat. Food Nutr. Agric. 2012, 4, 61–77. [Google Scholar] [CrossRef]
- Ferri, M.; Happel, A.; Zanaroli, G.; Bertolini, M.; Chiesa, S.; Commisso, M.; Guzzo, F.; Tassoni, A. Advances in Combined Enzymatic Extraction of Ferulic Acid from Wheat Bran. New Biotechnol. 2020, 56, 38–45. [Google Scholar] [CrossRef]
- Fritsch, C.; Staebler, A.; Happel, A.; Cubero Márquez, M.A.; Aguiló-Aguayo, I.; Abadias, M.; Gallur, M.; Cigognini, I.M.; Montanari, A.; López, M.J. Processing, Valorization and Application of Bio-Waste Derived Compounds from Potato, Tomato, Olive and Cereals: A Review. Sustainability 2017, 9, 1492. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, L.; Phillips, F.; O’sullivan, K.; Walton, J. Wheat Bran: Its Composition and Benefits to Health, a European Perspective. Int. J. Food Sci. Nutr. 2012, 63, 1001–1013. [Google Scholar] [CrossRef] [Green Version]
- Deroover, L.; Tie, Y.; Verspreet, J.; Courtin, C.M.; Verbeke, K. Modifying Wheat Bran to Improve its Health Benefits. Crit. Rev. Food Sci. Nutr. 2020, 60, 1104–1122. [Google Scholar] [CrossRef]
- Rosa, N.N.; Barron, C.; Gaiani, C.; Dufour, C.; Micard, V. Ultra-Fine Grinding Increases the Antioxidant Capacity of Wheat Bran. J. Cereal Sci. 2013, 57, 84–90. [Google Scholar] [CrossRef]
- Mosharraf, L.; Kadivar, M.; Shahedi, M. Effect of Hydrothermaled Bran on Physicochemical, Rheological and Microstructural Characteristics of Sangak Bread. J. Cereal Sci. 2009, 49, 398–404. [Google Scholar] [CrossRef]
- Aktas-Akyildiz, E.; Masatcioglu, M.T.; Köksel, H. Effect of Extrusion Treatment on Enzymatic Hydrolysis of Wheat Bran. J. Cereal Sci. 2020, 93, 102941. [Google Scholar] [CrossRef]
- Mao, M.; Wang, P.; Shi, K.; Lu, Z.; Bie, X.; Zhao, H.; Zhan, C.; Lv, F. Effect of Solid State Fermentation by Enterococcus Faecalis M2 on Antioxidant and Nutritional Properties of Wheat Bran. J. Cereal Sci. 2020, 94, 102997. [Google Scholar] [CrossRef]
- Hole, A.S.; Rud, I.; Grimmer, S.; Sigl, S.; Narvhus, J.; Sahlstrøm, S. Improved Bioavailability of Dietary Phenolic Acids in Whole Grain Barley and Oat Groat Following Fermentation with Probiotic Lactobacillus Acidophilus, Lactobacillus Johnsonii, and Lactobacillus Reuteri. J. Agric. Food Chem. 2012, 60, 6369–6375. [Google Scholar] [CrossRef] [PubMed]
- Nisa, K.; Rosyida, V.; Nurhayati, S.; Indrianingsih, A.; Darsih, C.; Apriyana, W. Total Phenolic Contents and Antioxidant Activity of Rice Bran Fermented with Lactic Acid Bacteria. In IOP Conference Series: Earth and Environmental Science, Proceedings of the 2nd International Conference on Natural Products and Bioresource Sciences—2018, Tangerang, Indonesia, 1–2 November 2018; IOP Publishing: Bristol, UK, 2018; Volume 251, p. 012020. [Google Scholar]
- Oliveira, P.M.; Zannini, E.; Arendt, E.K. Cereal Fungal Infection, Mycotoxins, and Lactic Acid Bacteria Mediated Bioprotection: From Crop Farming to Cereal Products. Food Microbiol. 2014, 37, 78–95. [Google Scholar] [CrossRef]
- Barac, A. Mycotoxins and human disease. In Clinically Relevant Mycoses; Presterl, E., Ed.; Springer International Publishing: New York, NY, USA, 2019; pp. 213–225. [Google Scholar]
- Patel, P.K.; Sharma, A.; Bhoomika, B.P.; Yadav, A. Mycotoxins and Animal Health: A Short Note. Pharma Innov. 2019, 8, 43–48. [Google Scholar]
- Liu, Y.; Yamdeu, J.H.G.; Gong, Y.Y.; Orfila, C. A Review of Postharvest Approaches to Reduce Fungal and Mycotoxin Contamination of Foods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1521–1560. [Google Scholar] [CrossRef]
- Tola, M.; Kebede, B. Occurrence, Importance and Control of Mycotoxins: A Review. Cogent Food Agric. 2016, 2, 1191103. [Google Scholar] [CrossRef]
- Duarte, S.; Pena, A.; Lino, C. A Review on Ochratoxin A Occurrence and Effects of Processing of Cereal and Cereal Derived Food Products. Food Microbiol. 2010, 27, 187–198. [Google Scholar] [CrossRef]
- Bartkiene, E.; Lele, V.; Ruzauskas, M.; Domig, K.J.; Starkute, V.; Zavistanaviciute, P.; Bartkevics, V.; Pugajeva, I.; Klupsaite, D.; Juodeikiene, G. Lactic Acid Bacteria Isolation from Spontaneous Sourdough and their Characterization Including Antimicrobial and Antifungal Properties Evaluation. Microorganisms 2020, 8, 64. [Google Scholar] [CrossRef] [Green Version]
- Arena, M.; de Nadra, M.M. Biogenic Amine Production by Lactobacillus. J. Appl. Microbiol. 2001, 90, 158–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsanhoty, R.M.; Ramadan, M.F. Genetic Screening of Biogenic Amines Production Capacity from some Lactic Acid Bacteria Strains. Food Control 2016, 68, 220–228. [Google Scholar] [CrossRef]
- Özogul, F.; Hamed, I. The Importance of Lactic Acid Bacteria for the Prevention of Bacterial Growth and their Biogenic Amines Formation: A Review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1660–1670. [Google Scholar] [CrossRef]
- Wójcik, W.; Łukasiewicz, M.; Puppel, K. Biogenic Amines: Formation, Action and Toxicity—A Review. J. Sci. Food Agric. 2020. [Google Scholar] [CrossRef]
- Xiang, H.; Sun-Waterhouse, D.; Waterhouse, G.I.; Cui, C.; Ruan, Z. Fermentation-Enabled Wellness Foods: A Fresh Perspective. Food Sci. Hum. Wellness. 2019, 8, 203–243. [Google Scholar] [CrossRef]
- Komesu, A.; de Oliveira, J.A.R.; da Silva Martins, L.H.; Maciel, M.R.W.; Maciel Filho, R. Lactic Acid Production to Purification: A Review. BioResources 2017, 12, 4364–4383. [Google Scholar] [CrossRef]
- Bianchetti, D.G.; Amelio, G.S.; Lava, S.A.; Bianchetti, M.G.; Simonetti, G.D.; Agostoni, C.; Fossali, E.F.; Milani, G.P. D-Lactic Acidosis in Humans: Systematic Literature Review. Pediatr. Nephrol. 2018, 33, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Cubas-Cano, E.; González-Fernández, C.; Ballesteros, M.; Tomás-Pejó, E. Biotechnological Advances in Lactic Acid Production by Lactic Acid Bacteria: Lignocellulose as Novel Substrate. Biofuel Bioprod. Biorefin. 2018, 12, 290–303. [Google Scholar] [CrossRef]
- Saeed, A.H.; Salam, A.I. Current Limitations and Challenges with Lactic Acid Bacteria: A Review. Food Nutr. Sci. 2013, 4, 73–87. [Google Scholar] [CrossRef] [Green Version]
- Le Lay, C.; Mounier, J.; Vasseur, V.; Weill, A.; Le Blay, G.; Barbier, G.; Coton, E. In Vitro and in Situ Screening of Lactic Acid Bacteria and Propionibacteria Antifungal Activities Against Bakery Product Spoilage Molds. Food Control 2016, 60, 247–255. [Google Scholar] [CrossRef]
- Arte, E.; Rizzello, C.G.; Verni, M.; Nordlund, E.; Katina, K.; Coda, R. Impact of Enzymatic and Microbial Bioprocessing on Protein Modification and Nutritional Properties of Wheat Bran. J. Agric. Food Chem. 2015, 63, 8685–8693. [Google Scholar] [CrossRef]
- Prückler, M.; Lorenz, C.; Endo, A.; Kraler, M.; Dürrschmid, K.; Hendriks, K.; da Silva, F.S.; Auterith, E.; Kneifel, W.; Michlmayr, H. Comparison of Homo-and Heterofermentative Lactic Acid Bacteria for Implementation of Fermented Wheat Bran in Bread. Food Microbiol. 2015, 49, 211–219. [Google Scholar] [CrossRef]
- Sieuwerts, S.; Bron, P.A.; Smid, E.J. Mutually Stimulating Interactions between Lactic Acid Bacteria and Saccharomyces Cerevisiae in Sourdough Fermentation. LWT-Food Sci. Technol. 2018, 90, 201–206. [Google Scholar] [CrossRef]
- Jiddere, G.; Filli, K.B. The Effect of Feed Moisture and Barrel Temperature on the Essential Amino Acids Profile of Sorghum Malt and Bambara Groundnut Based Extrudates. Int. J. Food Process. Technol. 2015, 6, 1. [Google Scholar]
- Spaggiari, M.; Ricci, A.; Calani, L.; Bresciani, L.; Neviani, E.; Dall’Asta, C.; Lazzi, C.; Galaverna, G. Solid State Lactic Acid Fermentation: A Strategy to Improve Wheat Bran Functionality. LWT-Food Sci. Technol. 2020, 118, 108668. [Google Scholar] [CrossRef]
- Zhao, H.; Guo, X.; Zhu, K. Impact of Solid State Fermentation on Nutritional, Physical and Flavor Properties of Wheat Bran. Food Chem. 2017, 217, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Osen, R.; Toelstede, S.; Eisner, P.; Schweiggert-Weisz, U. Effect of High Moisture Extrusion Cooking on Protein–protein Interactions of Pea (Pisum Sativum L.) Protein Isolates. Int. J. Food Sci. Tech. 2015, 50, 1390–1396. [Google Scholar] [CrossRef]
- Leonard, W.; Zhang, P.; Ying, D.; Fang, Z. Application of Extrusion Technology in Plant Food Processing Byproducts: An overview. Compr. Rev. Food Sci. Food Saf. 2020, 19, 218–246. [Google Scholar] [CrossRef]
- Ruiz-Capillas, C.; Herrero, A.M. Impact of Biogenic Amines on Food Quality and Safety. Foods 2019, 8, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costantini, A.; Pietroniro, R.; Doria, F.; Pessione, E.; Garcia-Moruno, E. Putrescine Production from Different Amino Acid Precursors by Lactic Acid Bacteria from Wine and Cider. Int. J. Food Microbiol. 2013, 165, 11–17. [Google Scholar] [CrossRef] [PubMed]
- del Rio, B.; Redruello, B.; Linares, D.M.; Ladero, V.; Ruas-Madiedo, P.; Fernandez, M.; Martin, M.C.; Alvarez, M.A. The Biogenic Amines Putrescine and Cadaverine show in Vitro Cytotoxicity at Concentrations that can be found in Foods. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Mantis, F.; Tsachev, I.; Sabatakou, O.; Burriel, A.; Vacalopoulos, A.; Ramantanis, S. Safety and Shelf-Life of Widely Distributed Vacuum Packed, Heat Treated Sausages. Bulg. J. Vet. Med. 2005, 8, 245–254. [Google Scholar]
- Palumbo, R.; Crisci, A.; Venâncio, A.; Cortiñas Abrahantes, J.; Dorne, J.; Battilani, P.; Toscano, P. Occurrence and Co-Occurrence of Mycotoxins in Cereal-Based Feed and Food. Microorganisms 2020, 8, 74. [Google Scholar] [CrossRef] [Green Version]
- EC-European Commission. Commission Recommendation 2006/576/EC of 17 August 2006 on the Presence of Deoxynivalenol, Zearalenone, Ochratoxin A, T-2 and HT-2 and Fumonisins in Products Intended for Animal Feeding. Off. J. Eur. Union Lex 2006, 229, 7–9. [Google Scholar]
- Zhao, L.; Jin, H.; Lan, J.; Zhang, R.; Ren, H.; Zhang, X.; Yu, G. Detoxification of Zearalenone by Three Strains of Lactobacillus Plantarum from Fermented Food in Vitro. Food Control 2015, 54, 158–164. [Google Scholar] [CrossRef]
- Kaushik, G. Effect of Processing on Mycotoxin Content in Grains. Crit. Rev. Food Sci. Nutr. 2015, 55, 1672–1683. [Google Scholar] [CrossRef]
- Chlebicz, A.; Śliżewska, K. In Vitro Detoxification of Aflatoxin B1, Deoxynivalenol, Fumonisins, T-2 Toxin and Zearalenone by Probiotic Bacteria from Genus Lactobacillus and Saccharomyces Cerevisiae Yeast. Probiotics Antimicrob. Proteins 2020, 12, 289–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castells, M.; Marin, S.; Sanchis, V.; Ramos, A. Fate of Mycotoxins in Cereals during Extrusion Cooking: A Review. Food Addit. Contam. 2005, 22, 150–157. [Google Scholar] [CrossRef]
- Samar, M.; Neira, M.; Resnik, S.; Pacin, A. Effect of Fermentation on Naturally Occurring Deoxynivalenol (DON) in Argentinean Bread Processing Technology. Food Addit. Contam. 2001, 18, 1004–1010. [Google Scholar] [CrossRef] [PubMed]
- Scudamore, K.; Banks, J.; Guy, R. Fate of Ochratoxin A in the Processing of Whole Wheat Grain during Extrusion. Food Addit. Contam. 2004, 21, 488–497. [Google Scholar] [CrossRef]
- Khaneghah, A.M.; Fakhri, Y.; Sant’Ana, A.S. Impact of Unit Operations during Processing of Cereal-Based Products on the Levels of Deoxynivalenol, Total Aflatoxin, Ochratoxin A, and Zearalenone: A Systematic Review and Meta-Analysis. Food Chem. 2018, 268, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Bartkiene, E.; Lele, V.; Sakiene, V.; Zavistanaviciute, P.; Ruzauskas, M.; Bernatoniene, J.; Jakstas, V.; Viskelis, P.; Zadeike, D.; Juodeikiene, G. Improvement of the Antimicrobial Activity of Lactic Acid Bacteria in Combination with Berries/Fruits and Dairy Industry By-products. J. Sci. Food Agric. 2019, 99, 3992–4002. [Google Scholar] [CrossRef] [PubMed]
- Bartkiene, E.; Bartkevics, V.; Krungleviciute, V.; Juodeikiene, G.; Zadeike, D.; Baliukoniene, V.; Bakutis, B.; Zelvyte, R.; Santini, A.; Cizeikiene, D. Application of Hydrolases and Probiotic Pediococcus Acidilactici BaltBio01 Strain for Cereal by-Products Conversion to Bioproduct for Food/Feed. Int. J. Food Sci. Nutr. 2018, 69, 165–175. [Google Scholar] [CrossRef]
- Starkute, V.; Bartkiene, E.; Bartkevics, V.; Rusko, J.; Zadeike, D.; Juodeikiene, G. Amino Acids Profile and Antioxidant Activity of Different Lupinus Angustifolius Seeds After Solid State and Submerged Fermentations. J Food Sci. Technol. 2016, 53, 4141–4148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Gigirey, B.; Vieites Baptista de Sousa, J.M.; Villa, T.G.; Barros-Velazquez, J. Changes in Biogenic Amines and Microbiological Analysis in Albacore (Thunnus Alalunga) Muscle during Frozen Storage. J. Food Prot. 1998, 61, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Gunasekaran, Y.K.; Lele, V.; Sakiene, V.; Zavistanaviciute, P.; Zokaityte, E.; Klupsaite, D.; Bartkevics, V.; Guiné, R.P.; Bartkiene, E. Plant-based Proteinaceous Snacks: Effect of Fermentation and Ultrasonication on End-product Characteristics. Food Sci. Nutr. 2020, 8, 4746–4756. [Google Scholar] [CrossRef] [PubMed]
- Akoglu, H. User’s Guide to Correlation Coefficients. Turk. J. Emerg. Med. 2018, 18, 91–93. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Samples | pH | TTA, °N | Lactic Acid Content, g/100 g | LAB | M/Y | TBC | TEC | Fructose | Glucose | Sucrose | Maltose | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Duration of Fermentation, h | L(+) | D(−) | log10 CFU/g | g/100 g | ||||||||||
| 0 | 24 | 0 | 24 | |||||||||||
| WCon | 6.04 ± 0.01b | − | 0.10 ± 0.02a | − | − | − | 5.20 ± 0.12c | 4.26 ± 0.11b | 9.04 ± 0.14c | 5.69 ± 0.23c | 0.16 ± 0.04ab | 0.55 ± 0.06a | nd | 0.31 ± 0.03c |
| WConLpl | 3.49 ± 0.02a | 4.20 ± 0.20c | 0.065 ± 0.003a | 1.988 ± 0.015d | 8.50 ± 0.14d | 3.64 ± 0.10a | 9.55 ± 0.17d | nd | 0.19 ± 0.03b | 0.69 ± 0.02b | nd | 0.35 ± 0.02c | ||
| WConLu | 3.46 ± 0.01a | 4.30 ± 0.20c | 0.070 ± 0.004a | 1.868 ± 0.024d | 9.09 ± 0.15e | 4.85 ± 0.12d | 9.55 ± 0.16d | nd | 0.12 ± 0.01a | 0.75 ± 0.04c | nd | 0.32 ± 0.03c | ||
| Wex115 | 6.17 ± 0.02c | − | 0.10 ± 0.03a | − | − | − | 4.85 ± 0.09b | 4.20 ± 0.12b | 8.93 ± 0.10c | 4.92 ± 0.14b | nd | nd | 0.17 ± 0.02a | 0.48 ± 0.04d |
| Wex115/Lpl | 3.81 ± 0.02b | 5.50 ± 0.20e | 0.666 ± 0.011d | 0.899 ± 0.016c | 8.69 ± 0.13d | 4.18 ± 0.10b | 8.92 ± 0.12c | nd | 0.18 ± 0.01b | nd | nd | 0.15 ± 0.01b | ||
| Wex115/Lu | 3.91 ± 0.01b | 4.80 ± 0.10d | 0.237 ± 0.007b | 0.770 ± 0.012b | 8.65 ± 0.11d | 4.25 ± 0.09b | 8.90 ± 0.11c | nd | 0.11 ± 0.01a | nd | nd | 0.39 ± 0.02c | ||
| Wex130/16 | 6.13 ± 0.02c | − | 0.10 ± 0.02a | − | − | − | 3.17 ± 0.09a | 4.37 ± 0.10b | 8.70 ± 0.19b | 4.67 ± 0.16b | nd | nd | 0.42 ± 0.05c | 0.16 ± 0.02b |
| Wex130/16/Lpl | 4.02 ± 0.01b | 3.80 ± 0.10b | 0.524 ± 0.010d | 0.683 ± 0.009b | 8.43 ± 0.14d | 4.48 ± 0.08c | 8.65 ± 0.11b | nd | 0.12 ± 0.01a | nd | nd | nd | ||
| Wex130/16/Lu | 4.28 ± 0.01c | 3.30 ± 0.10a | 0.438 ± 0.003c | 0.317 ± 0.014a | 8.54 ± 0.09d | 4.39 ± 0.12b | 8.79 ± 0.12b | nd | nd | nd | 0.14 ± 0.01a | nd | ||
| Wex130/20 | 5.96 ± 0.01a | − | 0.20 ± 0.04b | − | − | − | 4.41 ± 0.11b | 4.20 ± 0.12b | 8.65 ± 0.11b | 4.79 ± 0.15b | nd | nd | 0.26 ± 0.03b | 0.10 ± 0.02a |
| Wex130/20/Lpl | 3.90 ± 0.01b | 5.50 ± 0.20e | 0.440 ± 0.014c | 0.385 ± 0.017a | 8.75 ± 0.11d | 4.25 ± 0.10b | 8.95 ± 0.13c | nd | 0.12 ± 0.01a | nd | nd | nd | ||
| Wex130/20/Lu | 4.02 ± 0.02b | 4.90 ± 0.10d | 0.449 ± 0.012c | 0.360 ± 0.023a | 8.62 ± 0.12d | 4.17 ± 0.13b | 8.74 ± 0.12b | nd | 0.11 ± 0.01a | nd | nd | nd | ||
| Wex130/25 | 5.91 ± 0.02a | − | 0.20 ± 0.03b | − | − | − | 5.34 ± 0.09c | 4.38 ± 0.19b | 8.46 ± 0.10a | 4.32 ± 0.14a | 0.11 ± 0.02a | nd | 0.81 ± 0.07d | 0.11 ± 0.01a |
| Wex130/25/Lpl | 4.22 ± 0.02c | 3.90 ± 0.10b | 0.423 ± 0.009c | 0.322 ± 0.014a | 8.46 ± 0.11d | 4.29 ± 0.07b | 8.63 ± 0.12ab | nd | nd | nd | nd | nd | ||
| Wex130/25/Lu | 4.20 ± 0.01c | 3.50 ± 0.10a | 0.275 ± 0.013b | 0.203 ± 0.007a | 8.79 ± 0.12d | 4.32 ± 0.07b | 8.84 ± 0.13b | nd | nd | nd | nd | nd | ||
| Samples | Asp | Glu | Asn | Ser | His | Gly | Thr | Arg | Ala | Tyr | Cys | Val | Met | Trp | Phe | Ile | Leu | Lys | Pro |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WCon | 0.43 ± 0.03a | 1.75 ± 0.09b | nd | 0.29 ± 0.03a | 0.12 ± 0.01a | 0.27 ± 0.02a | 0.25 ± 0.02a | 0.31 ± 0.03a | 0.24 ± 0.02a | 0.18 ± 0.01a | 0.34 ± 0.03a | 0.34 ± 0.03a | 0.12 ± 0.01a | 0.36 ± 0.03b | 0.28 ± 0.02ab | 0.40 ± 0.04b | 0.14 ± 0.01ab | 0.26 ± 0.02a | 0.50 ± 0.04d |
| WConLpl | 0.50 ± 0.04ab | 1.91 ± 0.12b | nd | 0.33 ± 0.02a | 0.13 ± 0.01a | 0.33 ± 0.03ab | 0.30 ± 0.03a | 0.37 ± 0.03b | 0.29 ± 0.02b | 0.19 ± 0.01a | 0.38 ± 0.03a | 0.43 ± 0.03b | 0.10 ± 0.01a | 0.36 ± 0.03b | 0.24 ± 0.02a | 0.45 ± 0.04b | 0.15 ± 0.01b | 0.27 ± 0.02a | 1.17 ± 0.12f |
| WConLu | 0.48 ± 0.03a | 2.05 ± 0.14bc | nd | 0.34 ± 0.03a | 0.16 ± 0.01a | 0.32 ± 0.03ab | 0.30 ± 0.03a | 0.37 ± 0.03b | 0.28 ± 0.02b | 0.21 ± 0.02a | 0.44 ± 0.04b | 0.43 ± 0.04b | 0.12 ± 0.01a | 0.39 ± 0.03b | 0.25 ± 0.02a | 0.48 ± 0.04b | 0.15 ± 0.01b | 0.31 ± 0.03a | 1.09 ± 0.07f |
| Wex115 | 0.44 ± 0.02a | 1.92 ± 0.10b | nd | 0.33 ± 0.03a | 0.13 ± 0.01a | 0.30 ± 0.03a | 0.29 ± 0.02a | 0.35 ± 0.03b | 0.31 ± 0.03b | 0.21 ± 0.02a | 0.40 ± 0.03b | 0.42 ± 0.04ab | 0.10 ± 0.01a | 0.37 ± 0.03b | 0.23 ± 0.02a | 0.46 ± 0.04b | 0.15 ± 0.01b | 0.30 ± 0.02a | 0.82 ± 0.07e |
| Wex115/Lpl | 0.45 ± 0.03a | 1.89 ± 0.11b | nd | 0.32 ± 0.02a | 0.14 ± 0.01a | 0.30 ± 0.03a | 0.30 ± 0.03ab | 0.33 ± 0.03ab | 0.26 ± 0.02b | 0.19 ± 0.01a | 0.35 ± 0.03a | 0.36 ± 0.03a | 0.11 ± 0.01a | 0.32 ± 0.03ab | 0.22 ± 0.02a | 0.42 ± 0.04b | 0.13 ± 0.01a | 0.29 ± 0.02a | 0.80 ± 0.06e |
| Wex115/Lu | 0.53 ± 0.04a | 2.13 ± 0.08c | nd | 0.37 ± 0.03b | 0.15 ± 0.01a | 0.36 ± 0.03b | 0.35 ± 0.03b | 0.39 ± 0.03b | 0.37 ± 0.03b | 0.21 ± 0.02a | 0.43 ± 0.04b | 0.46 ± 0.04b | 0.10 ± 0.01a | 0.40 ± 0.03b | 0.26 ± 0.02a | 0.50 ± 0.05b | 0.17 ± 0.01b | 0.38 ± 0.03b | 1.03 ± 0.08f |
| Wex130/16 | 0.42 ± 0.03a | 1.74 ± 0.14b | nd | 0.32 ± 0.03a | 0.11 ± 0.01a | 0.29 ± 0.02a | 0.29 ± 0.02a | 0.33 ± 0.03ab | 0.26 ± 0.02b | 0.21 ± 0.02a | 0.36 ± 0.03a | 0.35 ± 0.03a | 0.17 ± 0.01b | 0.32 ± 0.03ab | 0.22 ± 0.02a | 0.43 ± 0.04b | 0.14 ± 0.01ab | 0.29 ± 0.02a | 1.05 ± 0.07f |
| Wex130/16/Lpl | 0.47 ± 0.03a | 1.74 ± 0.13b | nd | 0.31 ± 0.03a | 0.10 ± 0.01a | 0.32 ± 0.03ab | 0.29 ± 0.02a | 0.32 ± 0.03ab | 0.33 ± 0.03b | 0.20 ± 0.01a | 0.36 ± 0.03a | 0.35 ± 0.03a | 0.18 ± 0.01b | 0.33 ± 0.03a | 0.22 ± 0.02a | 0.42 ± 0.04b | 0.15 ± 0.01b | 0.29 ± 0.02a | 1.08 ± 0.09f |
| Wex130/16/Lu | 0.53 ± 0.04a | 1.97 ± 0.15b | nd | 0.32 ± 0.03a | 0.11 ± 0.01a | 0.30 ± 0.03a | 0.30 ± 0.03ab | 0.35 ± 0.03b | 0.27 ± 0.02b | 0.20 ± 0.01a | 0.44 ± 0.03b | 0.40 ± 0.03ab | 0.17 ± 0.01b | 0.42 ± 0.04b | 0.32 ± 0.03b | 0.44 ± 0.04b | 0.14 ± 0.01ab | 0.28 ± 0.02a | 0.36 ± 0.03c |
| Wex130/20 | 0.45 ± 0.04a | 1.48 ± 0.10a | nd | 0.26 ± 0.02a | 0.14 ± 0.01a | 0.24 ± 0.02a | 0.27 ± 0.02a | 0.28 ± 0.02a | 0.22 ± 0.02a | 0.18 ± 0.01a | 0.40 ± 0.03b | 0.34 ± 0.03a | 0.11 ± 0.01a | 0.33 ± 0.03ab | 0.24 ± 0.02a | 0.34 ± 0.03a | 0.12 ± 0.01a | 0.34 ± 0.02b | 0.29 ± 0.02b |
| Wex130/20/Lpl | 0.50 ± 0.04a | 1.49 ± 0.11a | nd | 0.26 ± 0.02a | 0.12 ± 0.01a | 0.28 ± 0.02a | 0.28 ± 0.02a | 0.29 ± 0.02a | 0.25 ± 0.02a | 0.17 ± 0.01a | 0.45 ± 0.04b | 0.36 ± 0.03a | 0.12 ± 0.01a | 0.33 ± 0.03ab | 0.24 ± 0.02a | 0.34 ± 0.03a | 0.12 ± 0.01a | 0.35 ± 0.03b | 0.18 ± 0.01a |
| Wex130/20/Lu | 0.48 ± 0.02a | 1.55 ± 0.12a | nd | 0.27 ± 0.02a | 0.12 ± 0.01a | 0.27 ± 0.02a | 0.28 ± 0.02a | 0.29 ± 0.02a | 0.24 ± 0.02a | 0.18 ± 0.01a | 0.42 ± 0.04b | 0.36 ± 0.03a | 0.11 ± 0.01a | 0.33 ± 0.03ab | 0.24 ± 0.02a | 0.33 ± 0.03a | 0.11 ± 0.01a | 0.37 ± 0.03b | 0.24 ± 0.02b |
| Wex130/25 | 0.44 ± 0.03a | 1.43 ± 0.09a | nd | 0.26 ± 0.02a | 0.10 ± 0.01a | 0.24 ± 0.02a | 0.25 ± 0.02a | 0.27 ± 0.02a | 0.21 ± 0.02a | 0.19 ± 0.01a | 0.38 ± 0.03a | 0.32 ± 0.03a | 0.13 ± 0.01a | 0.32 ± 0.03ab | 0.24 ± 0.02a | 0.32 ± 0.03a | 0.10 ± 0.01a | 0.29 ± 0.02a | 0.27 ± 0.02b |
| Wex130/25/Lpl | 0.48 ± 0.03a | 1.48 ± 0.07a | nd | 0.25 ± 0.02a | 0.10 ± 0.01a | 0.26 ± 0.02a | 0.28 ± 0.02a | 0.28 ± 0.02a | 0.23 ± 0.02a | 0.18 ± 0.01a | 0.35 ± 0.03a | 0.34 ± 0.03a | 0.09 ± 0.01a | 0.29 ± 0.02a | 0.22 ± 0.02a | 0.32 ± 0.03a | 0.11 ± 0.01a | 0.35 ± 0.03b | 0.14 ± 0.01a |
| Wex130/25/Lu | 0.48 ± 0.04a | 1.47 ± 0.08a | nd | 0.26 ± 0.02a | 0.11 ± 0.01a | 0.26 ± 0.02a | 0.26 ± 0.02a | 0.27 ± 0.02a | 0.23 ± 0.02a | 0.17 ± 0.01a | 0.40 ± 0.03ab | 0.34 ± 0.03a | 0.13 ± 0.01a | 0.29 ± 0.02a | 0.22 ± 0.02a | 0.32 ± 0.03a | 0.11 ± 0.01a | 0.34 ± 0.03b | 0.24 ± 0.02b |
| Samples | PUT | CAD | HIST | SPRM | TYR | PHE | SPRMD |
|---|---|---|---|---|---|---|---|
| WCon | 91.3 ± 2.4a | 33.8 ± 2.0a | 9.2 ± 0.4a | 35.9 ± 2.5b | nd | nd | nd |
| WConLpl | 88.6 ± 3.7a | 48.9 ± 3.5b | nd | 31.8 ± 1.7b | nd | nd | nd |
| WConLu | 109.3 ± 5.2b | nd | nd | 26.2 ± 0.9a | nd | nd | nd |
| Wex115 | 105.1 ± 4.3b | nd | nd | 35.2 ± 2.6b | nd | nd | nd |
| Wex115/Lpl | 138.6 ± 5.4e | nd | nd | 44.2 ± 3.4c | nd | nd | nd |
| Wex115/Lu | 139.6 ± 6.1e | nd | nd | 34.9 ± 2.5b | nd | nd | nd |
| Wex130/16 | 134.2 ± 3.4e | nd | nd | 30.6 ± 2.1b | nd | nd | nd |
| Wex130/16/Lpl | 115.4 ± 5.4bc | nd | nd | 33.9 ± 1.9b | nd | nd | nd |
| Wex130/16/Lu | 122.1 ± 2.8cd | nd | nd | 31.6 ± 2.0b | nd | nd | nd |
| Wex130/20 | 150.6 ± 6.3f | nd | nd | 31.5 ± 0.7b | nd | nd | nd |
| Wex130/20/Lpl | 126.6 ± 2.9d | nd | nd | 31.2 ± 1.8b | nd | nd | nd |
| Wex130/20/Lu | 154.1 ± 4.7f | nd | nd | 32.5 ± 2.3b | nd | nd | nd |
| Wex130/25 | 107.0 ± 5.4b | nd | nd | 25.3 ± 2.1a | nd | nd | nd |
| Wex130/25/Lpl | 160.1 ± 7.2f | nd | nd | 33.6 ± 3.1b | nd | nd | nd |
| Wex130/25/Lu | 91.3 ± 2.1a | 33.8 ± 2.1a | 9.2 ± 0.3a | 35.9 ± 2.7b | nd | nd | nd |
| Samples | AOH | AME | 17-DMAG | 15-DON | DON | D3G | 15ACS | ENN A | ENN A1 | FB1 | FB2 | MEL | STC | OTB | OTA | T-2 | HT-2 | FUSX | Neo | AFB1 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WCon | 1.76 ± 0.13d | nd | nd | 50.18 ± 1.21e | 58.8 ± 0.69f | 3.93 ± 0.29h | 15.28± 1.17d | 5.31 ± 0.27c | 1.24 ± 0.08c | nd | nd | 0.3 ± 0.02d | nd | nd | 7.66 ± 0.37d | 1.81 ± 0.15c | 3.76 ± 0.38f | 7.66 ± 0.37d | nd | 3.2 ± 0.21d |
| WConLpl | 0.87 ± 0.07a | nd | nd | 36.77 ± 0.25c | 26.5 ± 0.17c | 0.61 ± 0.05c | 21.99 ± 0.21e | 2.29 ± 0.14b | 0.44 ± 0.03b | nd | nd | 0.19 ± 0.01bc | nd | nd | nd | 1.7 ± 0.15c | 0.27 ± 0.09a | nd | nd | nd |
| WConLu | 0.72 ± 0.04a | nd | nd | 41.73 ± 0.36d | 27 ± 0.12c | 0.8 ± 0.04d | 19.2 ± 0.12d | 1.75 ± 0.09a | 0.41 ± 0.03b | nd | nd | 0.22 ± 0.02c | 0.09 ± 0.01a | nd | nd | nd | 0.2 ± 0.06a | nd | nd | nd |
| Wex115 | 0.85 ±0.07a | nd | nd | 75.17 ± 0.63g | 45.1 ± 0.58e | 2.79 ± 0.17g | 1.2 ± 0.11a | 1.61 ± 0.12a | 0.35 ± 0.02b | nd | nd | 0.43 ± 0.03e | 0.11 ± 0.01a | nd | 1.68 ± 0.13c | 0.89 ± 0.04b | 2.85 ± 0.11e | 4.05 ± 0.21b | 0.1 ± 0.01b | 2.55 ± 0.19c |
| Wex115/Lpl | 0.9 ± 0.04a | 1.09 ± 0.09c | nd | 63.36 ± 0.29f | 45.8 ± 0.27e | 2.68 ± 0.12g | 1.68 ± 0.07b | 39.51 ± 0.05f | 9.65 ± 0.11g | nd | nd | 0.28 ± 0.02d | 0.33 ± 0.02b | 0.08 ± 0.01a | 2.92 ± 0.14d | 2.56 ± 0.13d | 0.61 ± 0.05b | nd | 0.05 ±0.01a | nd |
| Wex115/Lu | 0.81 ± 0.03a | 0.88 ± 0.06b | nd | 51.09 ± 0.34e | 53 ± 0.32f | 1.92 ± 0.09f | 1.8 ± 0.09b | 25.53 ± 0.07e | 6.22 ± 0.09f | nd | nd | 0.16 ± 0.01b | 0.29 ± 0.02b | 0.07 ± 0.01a | nd | nd | 1 ± 0.08d | nd | 0.06 ± 0.01a | nd |
| Wex130/16 | 1 ± 0.05b | 0.85 ± 0.04b | 0.34 ± 0.01a | 106.45 ± 2.67i | 32.9 ± 0.26d | 1.83 ± 0.12f | nd | 1.34 ± 0.28a | 0.25 ± 0.02a | nd | nd | 0.19 ± 0.01b | 0.7 ± 0.06c | nd | 0.63 ± 0.03b | 0.98 ± 0.06b | nd | 4.54 ± 0.27b | 0.08 ± 0.01ab | 1.7 ± 0.09b |
| Wex130/16/Lpl | 0.88 ± 0.04a | nd | 0.37 ± 0.03a | 24.24 ± 0.11b | 30.8 ± 0.19d | 1.13 ± 0.06e | 1.71 ± 0.14b | 11.48 ± 0.22d | 2.52 ± 0.11e | nd | nd | 0.07 ± 0.01a | 0.96 ± 0.05d | nd | nd | 2.41 ± 0.14d | 0.63 ± 0.05b | nd | nd | nd |
| Wex130/16/Lu | 0.95 ± 0.05ab | nd | 0.39 ± 0.02a | nd | 30.2 ± 0.15d | 0.98 ± 0.05d | 1.66 ± 0.11b | 10.18 ± 0.09d | 1.87 ± 0.06d | nd | nd | 0.06 ± 0.01a | 0.75 ± 0.04c | nd | 6.44 ± 0.21e | nd | 0.86 ± 0.06c | nd | 0.04 ±0.01a | nd |
| Wex130/20 | 1.8 ± 0.12d | 0.51 ± 0.03a | 0.79 ± 0.04b | 85.65 ± 1.17h | 24.4 ± 0.25c | 1.05 ± 0.09d | 1.31 ± 0.11a | 1.29 ± 0.14a | 0.25 ± 0.02a | 0.91 ± 0.07d | 0.07 ± 0.01 | 0.16 ± 0.02b | 1.76 ± 0.08g | nd | 0.29 ± 0.02a | 0.85 ± 0.05b | nd | 4.9 ± 0.15c | 0.06 ± 0.01a | 0.91 ± 0.04a |
| Wex130/20/Lpl | 0.86 ± 0.05a | nd | 0.86 ± 0.06b | 20.93 ± 0.12a | 21.1 ± 0.09b | 0.26 ± 0.02a | 2.18 ± 0.10c | 2.17 ± 0.09b | 0.42 ± 0.02b | 0.56 ± 0.05c | nd | 0.03 ± 0.01a | 2.11 ± 0.12h | nd | nd | nd | 1.02 ± 0.07d | nd | 0.04 ± 0.01a | nd |
| Wex130/20/Lu | 3.42 ± 0.23e | 0.63 ± 0.04a | 1.12 ± 0.09c | nd | 24.1 ± 0.10c | 0.3 ± 0.02a | 1.43 ± 0.12a | 2.21 ± 0.10b | 0.36 ± 0.03b | 0.41 ± 0.04b | nd | nd | 1.75 ± 0.16g | nd | nd | nd | 0.85 ± 0.05c | 3.52 ± 0.14a | 0.03 ± 0.01a | nd |
| Wex130/25 | 3.29 ± 0.17e | 3.45 ± 0.15e | 0.82 ± 0.04b | 83.45 ± 1.27h | 23.2 ± 0.13c | 1.11 ± 0.07e | nd | 1.29 ± 0.09a | 0.26 ± 0.02a | 0.07 ±0.01a | nd | 0.05± 0.01a | 1.96± 0.14gh | nd | 0.54 ± 0.03b | 1.48 ± 0.09c | nd | 4.35 ± 0.17b | 0.07 ± 0.01a | 0.92 ± 0.04a |
| Wex130/25/Lpl | 1.37 ± 0.10c | 1.01 ± 0.08c | 1.05 ± 0.07c | 18.82 ± 0.10a | 19.5 ± 0.12a | 0.51 ± 0.04b | 1.44 ± 0.13a | 1.58 ± 0.17a | 0.31 ± 0.02b | 0.04 ± 0.01a | nd | 0.04 ± 0.01a | 1.43 ± 0.11f | nd | nd | nd | 1.05 ± 0.04d | nd | 0.05 ± 0.01a | nd |
| Wex130/25/Lu | 1.31 ± 0.12c | 1.4 ± 0.09d | 0.78 ± 0.06b | nd | 19.9 ± 0.14a | 0.44 ± 0.02b | 1.81 ± 0.15b | 1.29 ± 0.09a | 0.26 ± 0.02a | 0.08 ± 0.01a | nd | 0.02 ± 0.01a | 1.27 ± 0.09e | nd | nd | nd | 1.17 ± 0.06d | nd | 0.05 ± 0.02a | nd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartkiene, E.; Zokaityte, E.; Lele, V.; Starkute, V.; Zavistanaviciute, P.; Klupsaite, D.; Cernauskas, D.; Ruzauskas, M.; Bartkevics, V.; Pugajeva, I.; et al. Combination of Extrusion and Fermentation with Lactobacillus plantarum and L. uvarum Strains for Improving the Safety Characteristics of Wheat Bran. Toxins 2021, 13, 163. https://doi.org/10.3390/toxins13020163
Bartkiene E, Zokaityte E, Lele V, Starkute V, Zavistanaviciute P, Klupsaite D, Cernauskas D, Ruzauskas M, Bartkevics V, Pugajeva I, et al. Combination of Extrusion and Fermentation with Lactobacillus plantarum and L. uvarum Strains for Improving the Safety Characteristics of Wheat Bran. Toxins. 2021; 13(2):163. https://doi.org/10.3390/toxins13020163
Chicago/Turabian StyleBartkiene, Elena, Egle Zokaityte, Vita Lele, Vytaute Starkute, Paulina Zavistanaviciute, Dovile Klupsaite, Darius Cernauskas, Modestas Ruzauskas, Vadims Bartkevics, Iveta Pugajeva, and et al. 2021. "Combination of Extrusion and Fermentation with Lactobacillus plantarum and L. uvarum Strains for Improving the Safety Characteristics of Wheat Bran" Toxins 13, no. 2: 163. https://doi.org/10.3390/toxins13020163
APA StyleBartkiene, E., Zokaityte, E., Lele, V., Starkute, V., Zavistanaviciute, P., Klupsaite, D., Cernauskas, D., Ruzauskas, M., Bartkevics, V., Pugajeva, I., Bērziņa, Z., Gruzauskas, R., Sidlauskiene, S., Santini, A., & Juodeikiene, G. (2021). Combination of Extrusion and Fermentation with Lactobacillus plantarum and L. uvarum Strains for Improving the Safety Characteristics of Wheat Bran. Toxins, 13(2), 163. https://doi.org/10.3390/toxins13020163