Antifungal Activity of Quinofumelin against Fusarium graminearum and Its Inhibitory Effect on DON Biosynthesis

 , and
, and
Abstract
:1. Introduction
2. Results
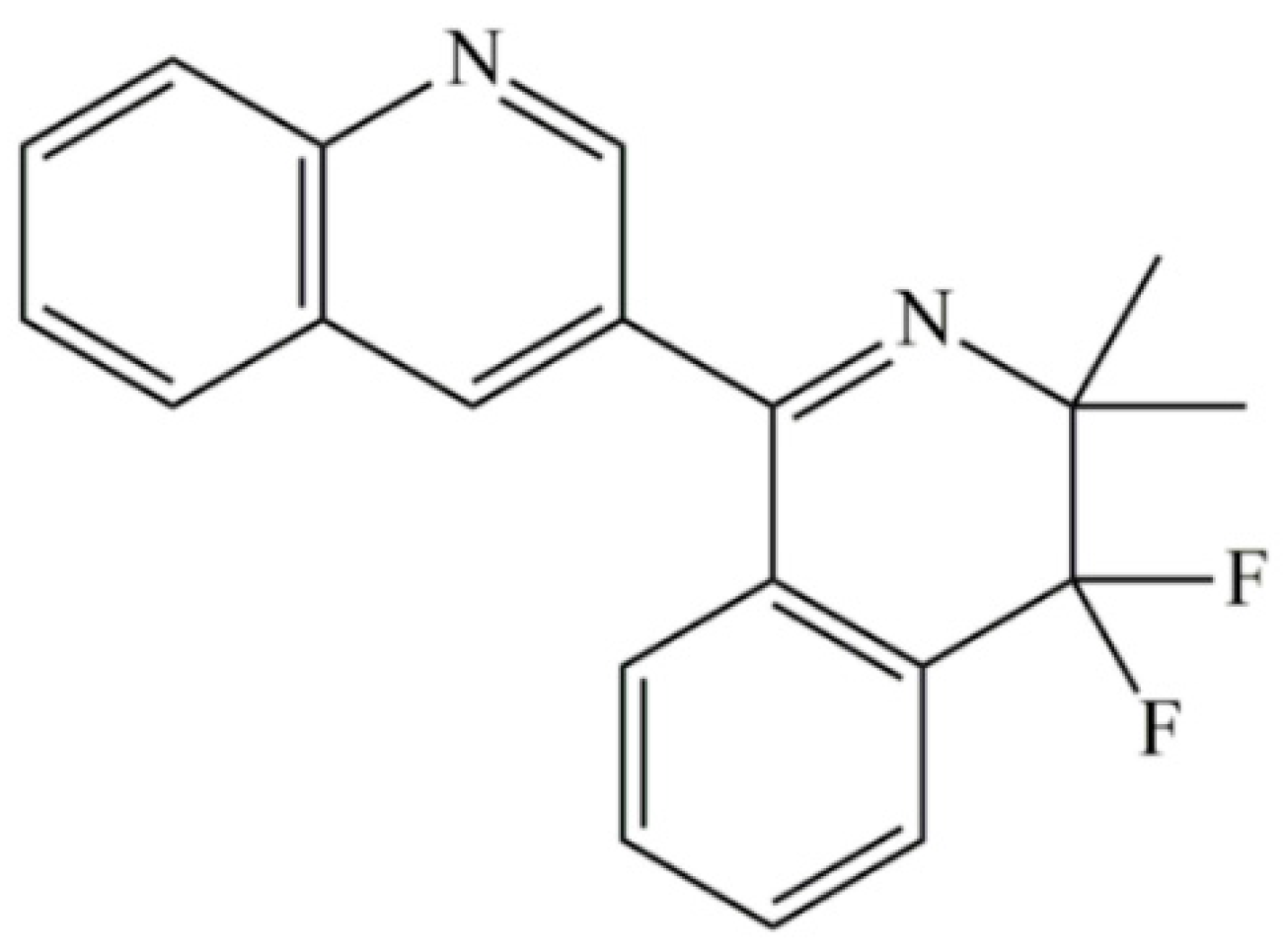
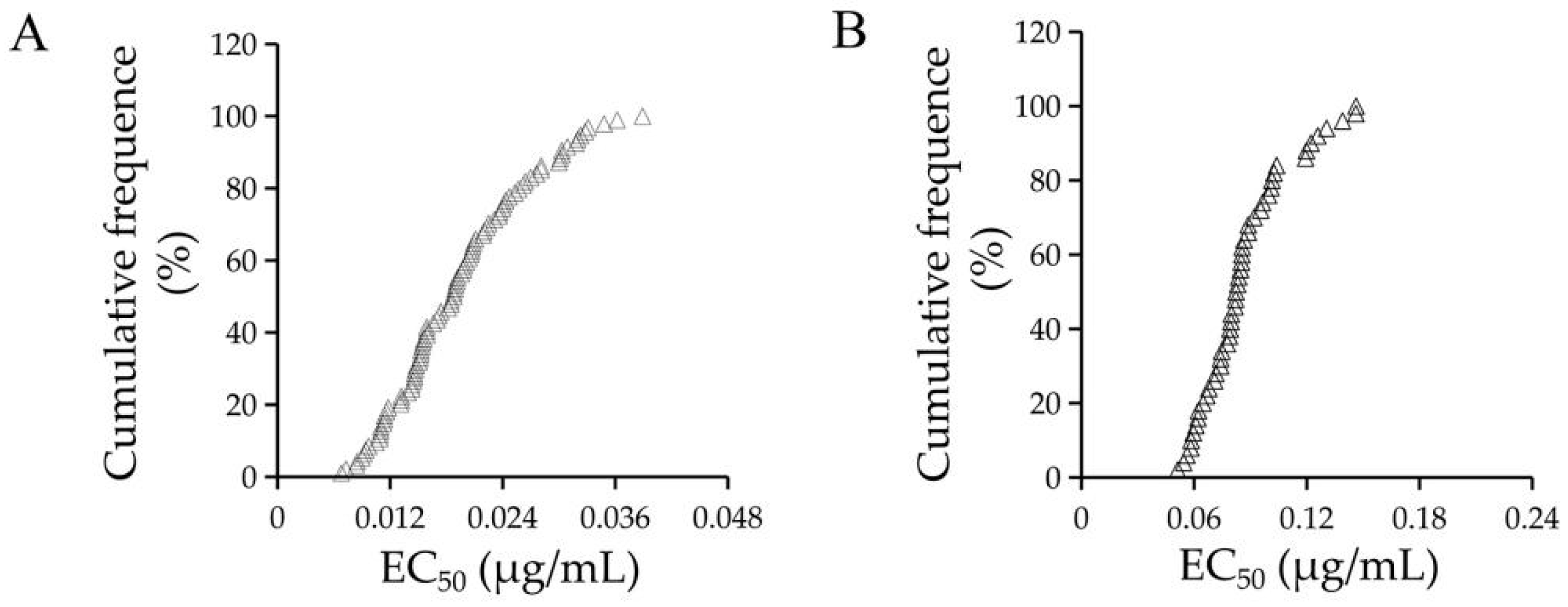
2.1. The Sensitivity of F. graminearum to Quinofumelin
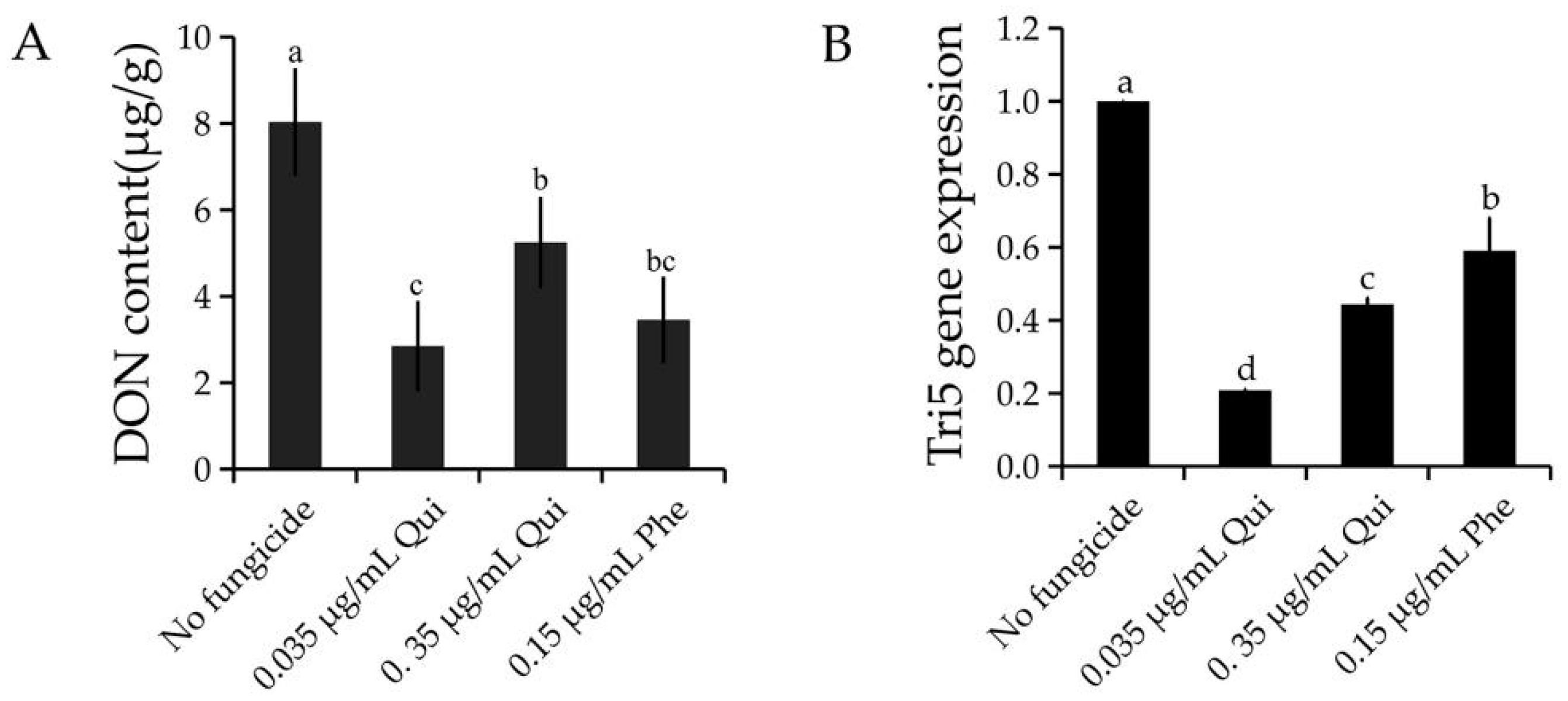
2.2. Quinofumelin Reduced DON Biosynthesis and TRI5 Gene Expression in F. graminearum
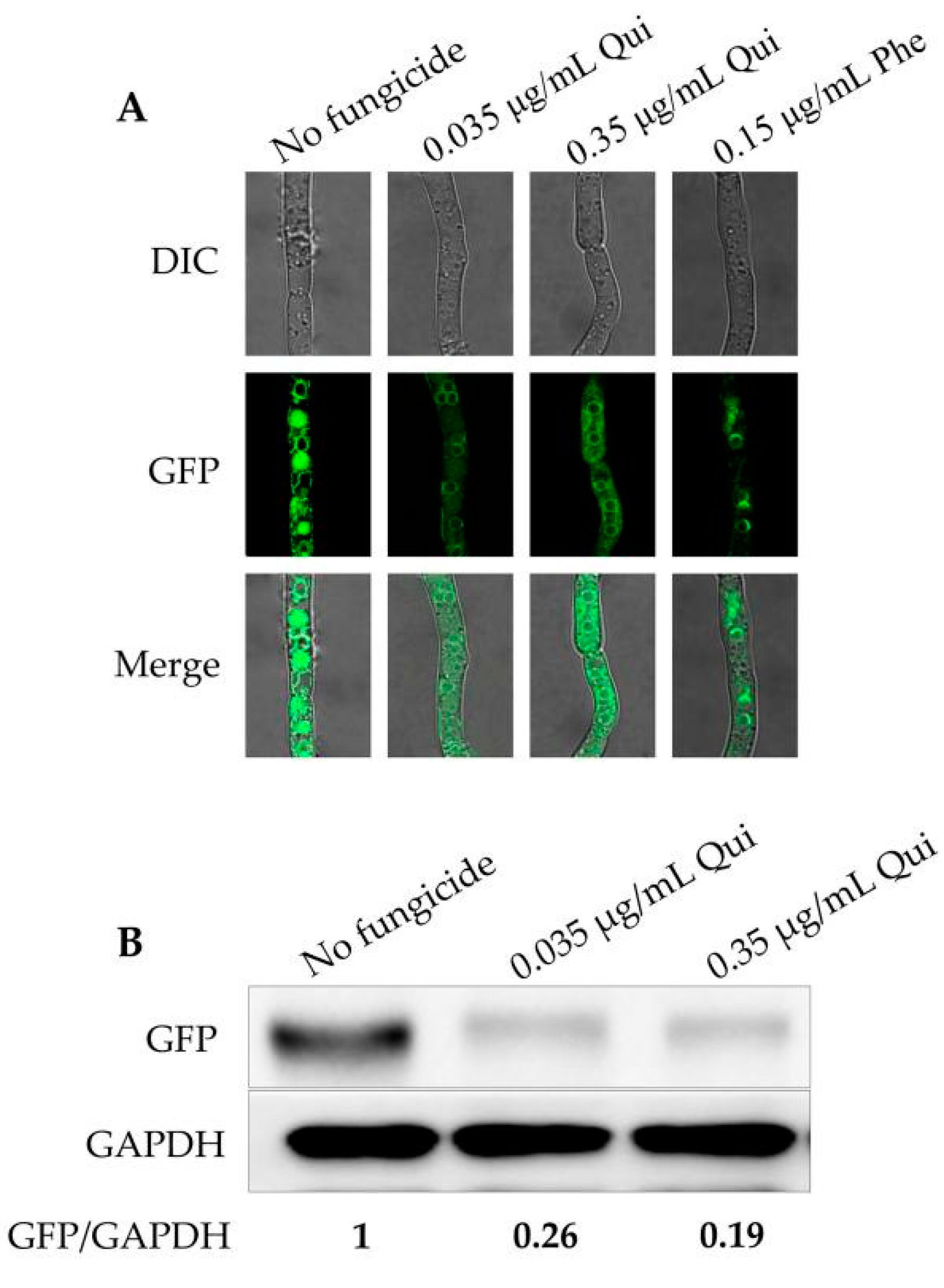
2.3. Quinofumelin Affects the Formation of Toxisomes and the Expression of TRI1 in the Translational Level in F. graminearum
2.4. Protective and Curative Activity of Quinofumelin against FHB
3. Discussion
4. Materials and Methods
4.1. Fungal Growth and Culture Conditions
4.2. Fungicides
4.3. Fungicide Sensitivity Test
4.4. DON Biosynthesis In Vitro
4.5. Formation of Toxisomes and Western Blotting Assays
4.6. Protective and Curative Activity of Quinofumelin on Wheat Coleoptiles
4.7. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- McMullen, M.; Jones, R.; Gallenberg, D. Scab of wheat and barley: A re-emerging disease of devastating impact. Plant Dis. 1997, 81, 1340–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.X.; Zhang, X.; Yao, J.B.; Cheng, S.H. Breeding for the resistance to Fusarium head blight of wheat in China. Front. Agric. Sci. Eng. 2019, 6, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Palazzini, J.; Fumero, V.; Yerkovich, N.; Barros, G.; Cuniberti, M.; Chulze, S. Correlation between Fusarium graminearum and deoxynivalenol during the 2012/13 wheat Fusarium head blight outbreak in Argentina. Cereal Res. Commun. 2015, 43, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.W.; Hao, Y.F.; Mergoum, M.; Bai, G.H.; Humphreys, G.; Cloutier, S.; Xia, X.C.; He, Z.H. Breeding wheat for resistance to Fusarium head blight in the Global North: China, USA, and Canada. Crop J. 2019, 7, 730–738. [Google Scholar] [CrossRef]
- McMullen, M.; Bergstrom, G.; De Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; Van Sanford, D. A unified effort to fight an enemy of wheat and barley: Fusarium head blight. Plant Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audenaert, K.; Vanheule, A.; Hofte, M.; Haesaert, G. Deoxynivalenol: A major player in the multifaceted response of Fusarium to its environment. Toxins 2014, 6, 1–19. [Google Scholar] [CrossRef] [Green Version]
- De Walle, J.V.; Sergent, T.; Piront, N.; Toussaint, O.; Schneider, Y.J.; Larondelle, Y. Deoxynivalenol affects in vitro intestinal epithelial cell barrier integrity through inhibition of protein synthesis. Toxicol. Appl. Pharm. 2010, 245, 291–298. [Google Scholar] [CrossRef]
- Figueroa, M.; Hammond-Kosack, K.E.; Solomon, P.S. A review of wheat diseases-a field perspective. Mol. Plant Pathol. 2018, 19, 1523–1536. [Google Scholar] [CrossRef]
- Yuan, S.K.; Zhou, M.G. A major gene for resistance to carbendazim, in field isolates of Gibberella zeae. Can. J. Plant Pathol. 2005, 27, 58–63. [Google Scholar] [CrossRef]
- Duan, Y.B.; Zhang, X.K.; Ge, C.Y.; Wang, Y.; Cao, J.H.; Jia, X.J.; Wang, J.X.; Zhou, M.G. Development and application of loop-mediated isothermal amplification for detection of the F167Y mutation of carbendazim-resistant isolates in Fusarium graminearum. Sci. Rep. 2014, 4, 7094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Y.B.; Tao, X.; Zhao, H.H.; Xiao, X.M.; Li, M.X.; Wang, J.X.; Zhou, M.G. Activity of demethylation inhibitor fungicide metconazole on Chinese Fusarium graminearum species complex and its application in carbendazim-resistance management of Fusarium head blight in wheat. Plant Dis. 2019, 103, 929–937. [Google Scholar] [CrossRef]
- Duan, Y.B.; Li, M.X.; Zhao, H.H.; Lu, F.; Wang, J.X.; Zhou, M.G. Molecular and biological characteristics of laboratory metconazole-resistant mutants in Fusarium graminearum. Pestic. Biochem. Physiol. 2018, 152, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.H.; Duan, Y.B.; Zhou, M.G. Carbendazim-resistance associated beta (2)-tubulin substitutions increase deoxynivalenol biosynthesis by reducing the interaction between beta (2)-tubulin and IDH3 in Fusarium graminearum. Environ. Microbiol. 2020, 22, 598–614. [Google Scholar] [CrossRef] [PubMed]
- Li, H.K.; Diao, Y.M.; Wang, H.X.; Chen, C.J.; Ni, J.P.; Zhou, M.G. JS399-19, a new fungicide against wheat scab. Crop Prot. 2008, 27, 90–95. [Google Scholar] [CrossRef]
- Chen, Y.; Li, H.K.; Chen, C.J.; Zhou, M.G. Sensitivity of Fusarium graminearum to fungicide JS399-19: In vitro determination of baseline sensitivity and the risk of developing fungicide resistance. Phytoparasitica 2008, 36, 326–337. [Google Scholar] [CrossRef]
- Duan, Y.B.; Xiao, X.M.; Li, T.; Chen, W.W.; Wang, J.X.; Fraaije, B.A.; Zhou, M.G. Impact of epoxiconazole on Fusarium head blight control, grain yield and deoxynivalenol accumulation in wheat. Pestic. Biochem. Physiol. 2018, 152, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Liu, X.; Li, B.; Ma, Z. Characterization of sterol demethylation inhibitor-resistant isolates of Fusarium asiaticum and F. graminearum collected from wheat in China. Phytopathology 2009, 99, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, I.E.; Hollomon, D.W.; Gustafson, G.; Mitchell, J.C.; Longhurst, C.; Zhang, Z.; Gurr, S.J. Quinoxyfen perturbs signal transduction in barley powdery mildew (Blumeria graminis f.sp. hordei). Mol. Plant Pathol. 2003, 4, 177–186. [Google Scholar] [CrossRef]
- Hao, S.L.; Tian, J.F.; Xu, Y.; Fan, W.Y.; Jiang, A.R.; Liu, C.L. Synthesis and biological activity of Tebufloquin. Agrochemicals 2012, 51, 410–412. [Google Scholar]
- FRAC Classification of Fungicides. Available online: https://www.frac.info/docs/default-source/publications/frac-mode-of-action-poster/frac-moa-poster-2021.pdf?sfvrsn=a6f6499a_2 (accessed on 16 April 2021).
- Tao, X.; Zhao, H.H.; Xu, H.R.; Li, Z.K.; Wang, J.X.; Song, X.S.; Zhou, M.G.; Duan, Y.B. Antifungal activity and biological characteristics of the novel fungicide quinofumelin against Sclerotinia sclerotiorum. Plant Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Alexander, N.J.; Proctor, R.H.; McCormick, S.P. Genes, gene clusters, and biosynthesis of trichothecenes and fumonisins in Fusarium. Toxin Rev. 2009, 28, 198–215. [Google Scholar] [CrossRef]
- Xu, C.; Li, M.; Zhou, Z.; Li, J.; Chen, D.; Duan, Y.; Zhou, M. Impact of five succinate dehydrogenase inhibitors on DON biosynthesis of Fusarium asiaticum, Causing Fusarium head blight in wheat. Toxins 2019, 11, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boenisch, M.J.; Blum, A.; Broz, K.L.; Gardiner, D.M.; Kistler, H.C. Nanoscale enrichment of the cytosolic enzyme trichodiene synthase near reorganized endoplasmic reticulum in Fusarium graminearum. Fungal Genet. Biol. 2019, 124, 73–77. [Google Scholar] [CrossRef]
- Liu, X.M.; Zheng, Z.T.; Li, B.; Cai, Y.Q.; Mao, X.W.; Zhou, M.G. A myosin passenger protein gene (FaSmy1) is an essential regulator of cell development, pathogenicity, DON biosynthesis, and resistance to the fungicide phenamacril in Fusarium asiaticum. Eur. J. Plant Pathol. 2017, 148, 709–722. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Yu, J.J.; Zhang, Y.N.; Zhang, X.; Cheng, C.J.; Wang, J.X.; Hollomon, D.W.; Fan, P.S.; Zhou, M.G. Effect of carbendazim resistance on trichothecene production and aggressiveness of Fusarium graminearum. Mol. Plant-Microbe Interact. 2009, 22, 1143–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, G.F.; Chen, Y.; Xu, J.R.; Kistler, H.C.; Ma, Z.H. The fungal myosin I is essential for Fusarium toxisome formation. PLoS Path. 2018, 14, e1006827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Chen, W.; Shao, W.; Wang, J.; Lv, C.; Ma, H.; Chen, C. Molecular, biological and physiological characterizations of resistance to phenamacril in Fusarium graminearum. Plant Pathol. 2017, 66, 1404–1412. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, M.H.; Liu, W.D.; Xiang, J.; Li, J.M.; Li, Z.; Liu, X.P.; Huang, M.Z.; Liu, A.P.; Zheng, X.L. Synthesis and fungicidal activities of perfluoropropan-2-yl-based novel quinoline derivatives. Heterocycl. Commun. 2019, 25, 91–97. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, W.X.; Zhang, A.F.; Gu, C.Y.; Zhou, M.G.; Gao, T.C. Activity of the fungicide JS399-19 against Fusarium head blight of wheat and the risk of resistance. Agric. Sci. China 2011, 10, 1906–1913. [Google Scholar] [CrossRef]
- Liu, S.M.; Liu, J.L.; Fu, L.Y.; Chen, J.P.; Wang, S.; Jiang, J.; Che, Z.P.; Tian, Y.E.; Chen, G.Q. Baseline sensitivity of Fusarium graminearum from wheat fields in Henan, China, to metconazole and analysis of cross resistance with carbendazim and phenamacril. J. Phytopathol. 2020, 168, 156–161. [Google Scholar] [CrossRef]
- Duan, Y.B.; Lu, F.; Zhou, Z.H.; Zhao, H.H.; Zhang, J.; Mao, Y.S.; Li, M.X.; Wang, J.X.; Zhou, M.G. Quinone outside inhibitors affect DON biosynthesis, mitochondrial structure and toxisome formation in Fusarium graminearum. J. Hazard. Mater. 2020, 398, 122908. [Google Scholar] [CrossRef] [PubMed]
- Proctor, R.H.; Hohn, T.M.; Mccormick, S.P. Reduced virulence of Gibberella zeae caused by disruption of a trichothecene toxin biosynthetic gene. Mol. Plant-Microbe Interact. 1995, 8, 593–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goswami, R.S.; Kistler, H.C. Pathogenicity and in planta mycotoxin accumulation among members of the Fusarium graminearum species complex on wheat and rice. Phytopathology 2005, 95, 1397–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.J.; Zhu, Y.Y.; Li, Y.J.; Duan, Y.B.; Zhang, R.S.; Zhou, M.G. Beta 1 tubulin rather than beta 2 tubulin is the preferred binding target for carbendazim in Fusarium graminearum. Phytopathology 2016, 106, 978–985. [Google Scholar] [CrossRef] [Green Version]
- Segmuller, N.; Ellendorf, U.; Tudzynski, B.; Tudzynski, P. BcSAK1, a stress-activated mitogen-activated protein kinase, is involved in vegetative differentiation and pathogenicity in Botrytis cinerea. Eukaryot. Cell 2007, 6, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boenisch, M.J.; Broz, K.L.; Purvine, S.O.; Chrisler, W.B.; Nicora, C.D.; Connolly, L.R.; Freitag, M.; Baker, S.E.; Kistler, H.C. Structural reorganization of the fungal endoplasmic reticulum upon induction of mycotoxin biosynthesis. Sci. Rep. 2017, 7, 44296. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungicide | Concentration (μg/mL) | Protective Activity | Curative Activity | ||
|---|---|---|---|---|---|
| Lesion Length a (cm) | Control Efficacy a (%) | Lesion Length a (cm) | Control Efficacy a (%) | ||
| Quinofumelin | 20 | 0.93 ± 0.24 a | 57.69 d | 1.04 ± 0.22 b | 53.58 c |
| 40 | 0.48 ± 0.18 b | 78.19 c | 0.56 ± 0.11 c | 75.16 b | |
| 80 | 0.42 ± 0.18 b | 81.05 c | 0.16 ± 0.07 d | 92.93 a | |
| Carbendazim | 100 | 0.16 ± 0.09 c | 92.44 b | 1.35 ± 0.24 a | 40.00 d |
| Phenamacril | 50 | 0.05 ± 0.01 d | 97.58 a | 0.09 ± 0.02 d | 95.98 a |
| Primer | Sequence (5’-3’) | Use |
|---|---|---|
| FGSG_03537-qF | GGCTTCCCTCCAAACAAT | RT-qPCR for the expression of Tri5 |
| FGSG_03537-qR | TGGGAAAGTGCTCGTTGA | |
| GAPDH-qF | CTTACTGCCTCCACCAACTG | RT-qPCR for the internal control |
| GAPDH-qR | TGACGTTGGAAGGAGCGAAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiu, Q.; Bi, L.; Xu, H.; Li, T.; Zhou, Z.; Li, Z.; Wang, J.; Duan, Y.; Zhou, M. Antifungal Activity of Quinofumelin against Fusarium graminearum and Its Inhibitory Effect on DON Biosynthesis. Toxins 2021, 13, 348. https://doi.org/10.3390/toxins13050348
Xiu Q, Bi L, Xu H, Li T, Zhou Z, Li Z, Wang J, Duan Y, Zhou M. Antifungal Activity of Quinofumelin against Fusarium graminearum and Its Inhibitory Effect on DON Biosynthesis. Toxins. 2021; 13(5):348. https://doi.org/10.3390/toxins13050348
Chicago/Turabian StyleXiu, Qian, Lianyu Bi, Haorong Xu, Tao Li, Zehua Zhou, Zhongke Li, Jianxin Wang, Yabing Duan, and Mingguo Zhou. 2021. "Antifungal Activity of Quinofumelin against Fusarium graminearum and Its Inhibitory Effect on DON Biosynthesis" Toxins 13, no. 5: 348. https://doi.org/10.3390/toxins13050348
APA StyleXiu, Q., Bi, L., Xu, H., Li, T., Zhou, Z., Li, Z., Wang, J., Duan, Y., & Zhou, M. (2021). Antifungal Activity of Quinofumelin against Fusarium graminearum and Its Inhibitory Effect on DON Biosynthesis. Toxins, 13(5), 348. https://doi.org/10.3390/toxins13050348