Evaluation of the Therapeutic Effects of Protocatechuic Aldehyde in Diabetic Nephropathy

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
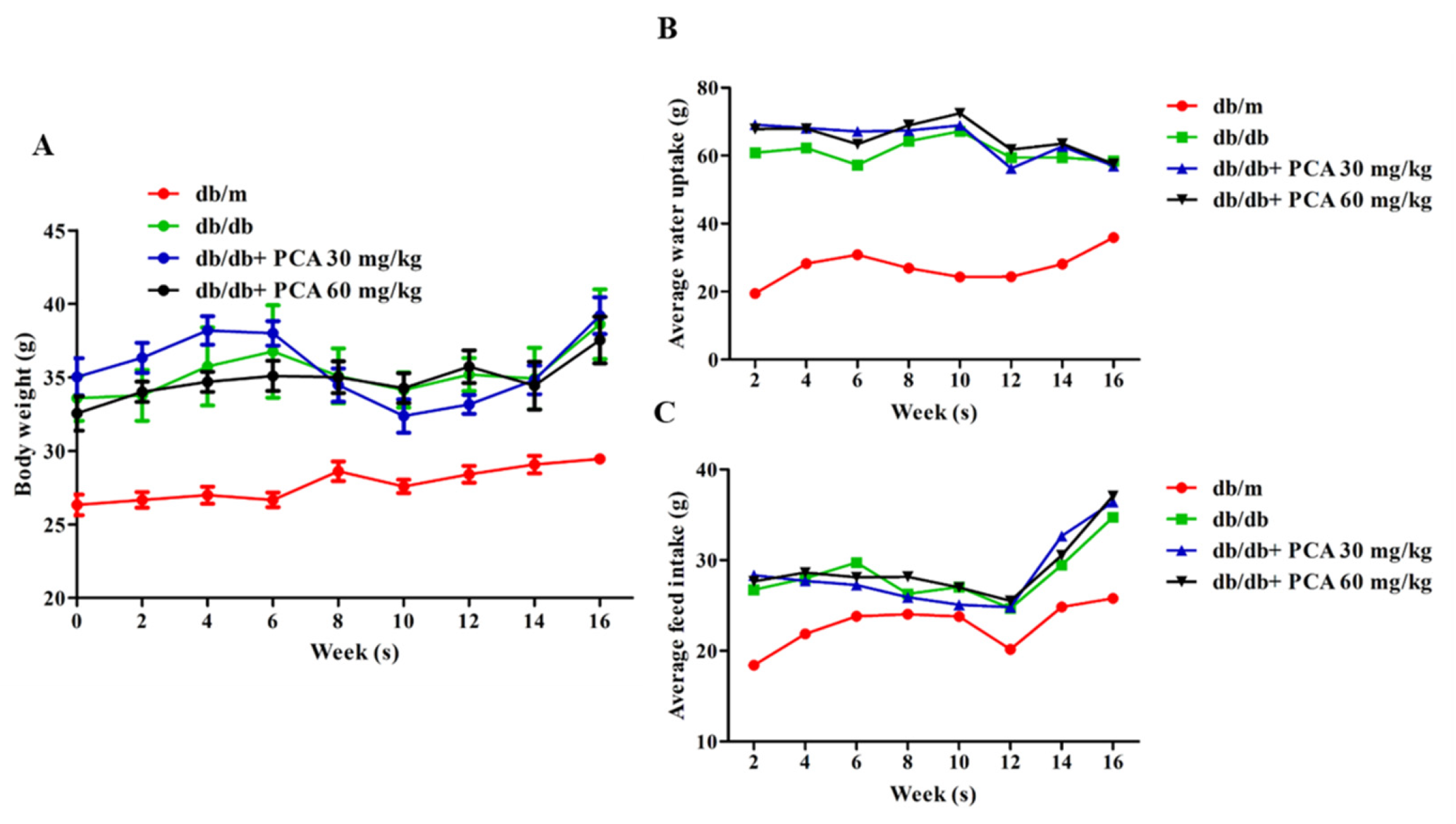
2.1. Effect of PCA Treatment on Body Weight, Food Intake, and Water Intake
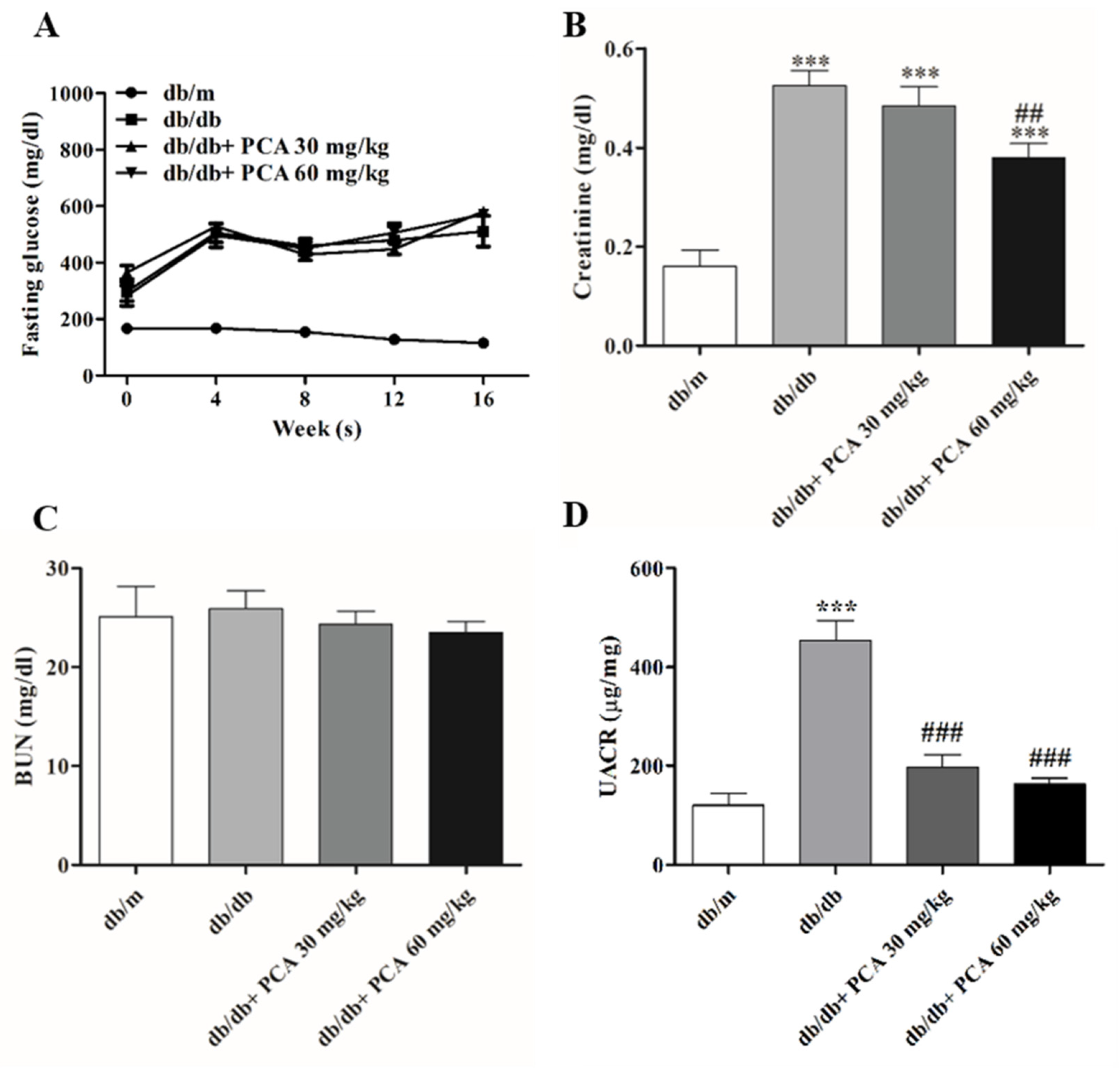
2.2. PCA Improves Renal Functions in Diabetic Mice
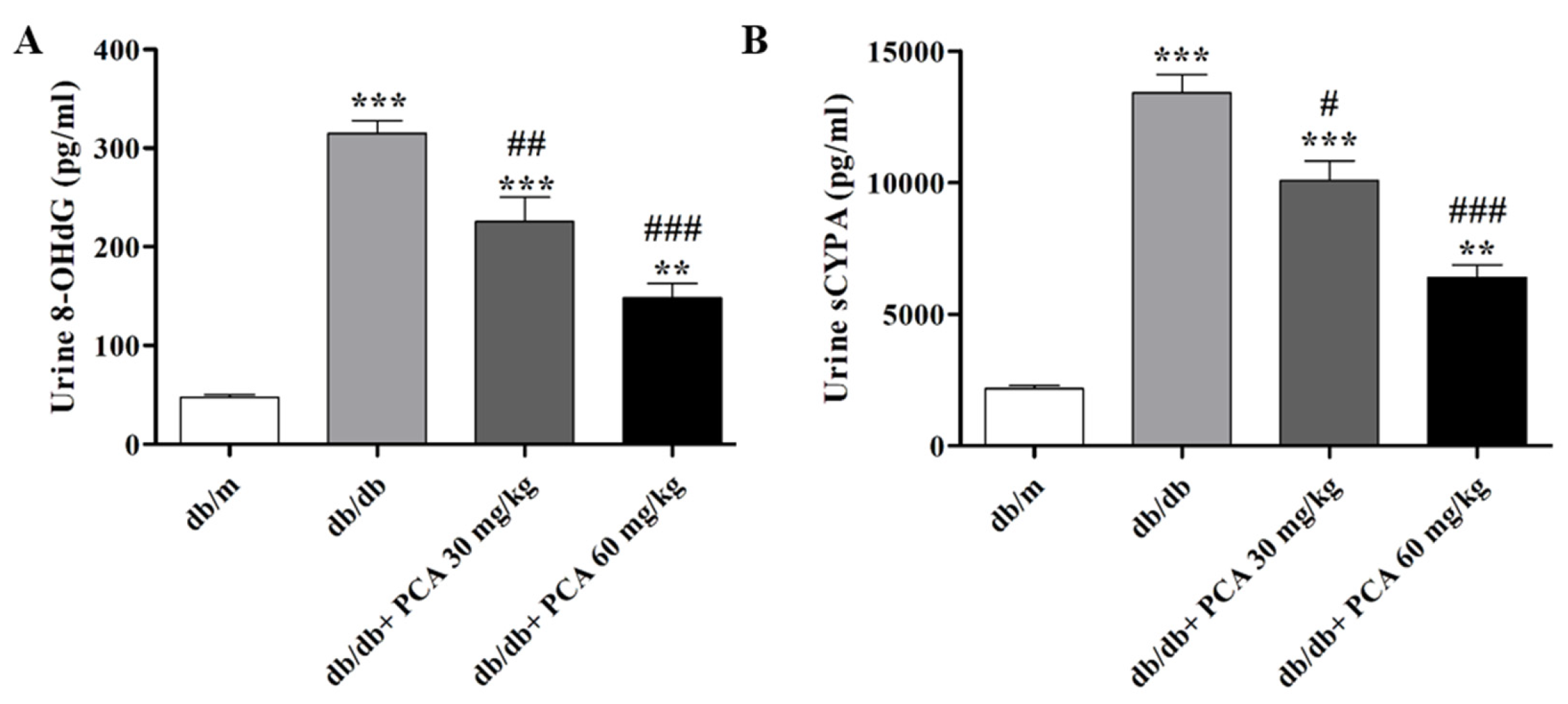
2.3. PCA Suppresses sCypA and 8-OHdG in Diabetic Mice Urine
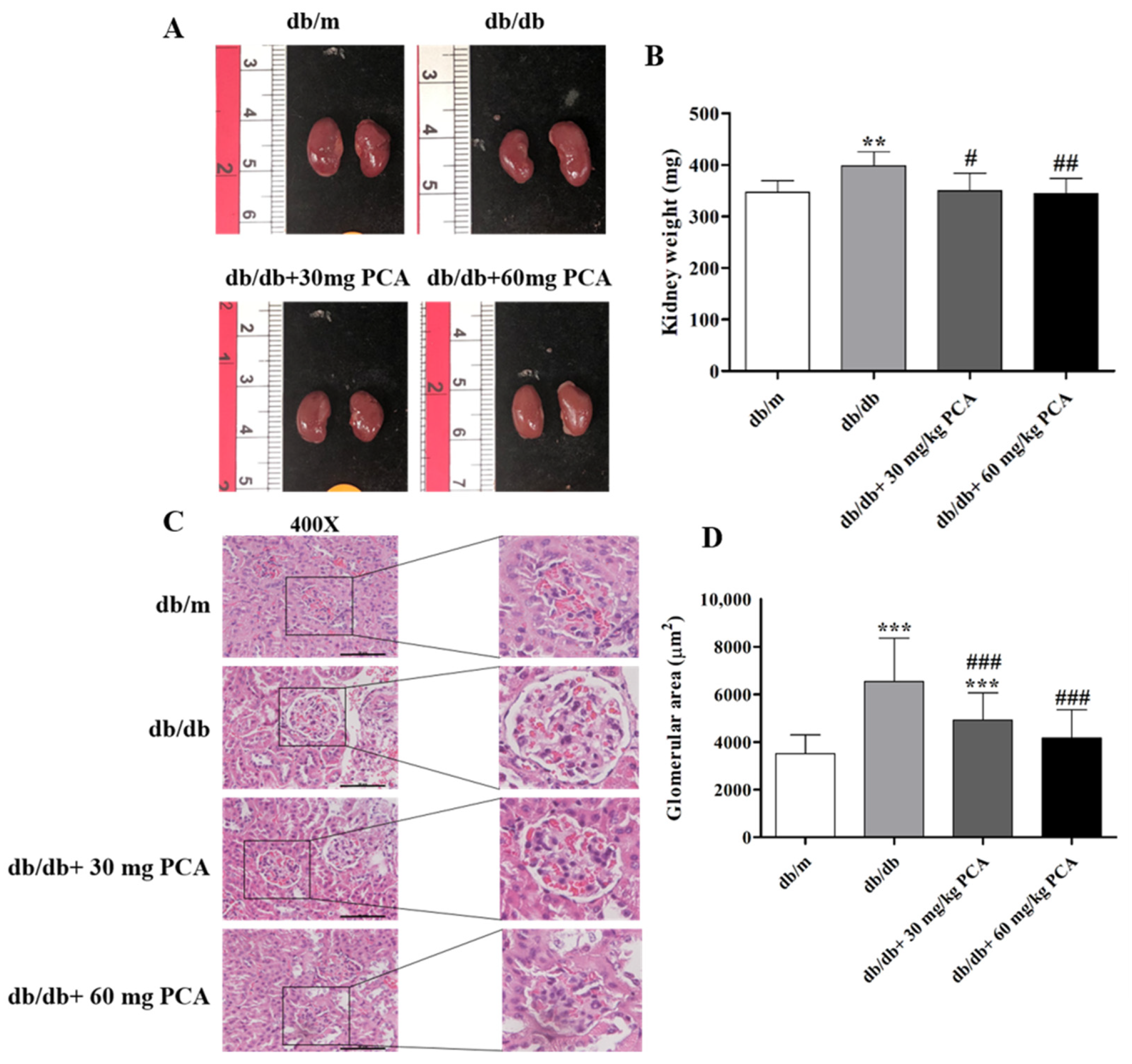
2.4. PCA Reduces Kidney Weight and Total Glomerular Area in db/db Mouse Kidneys
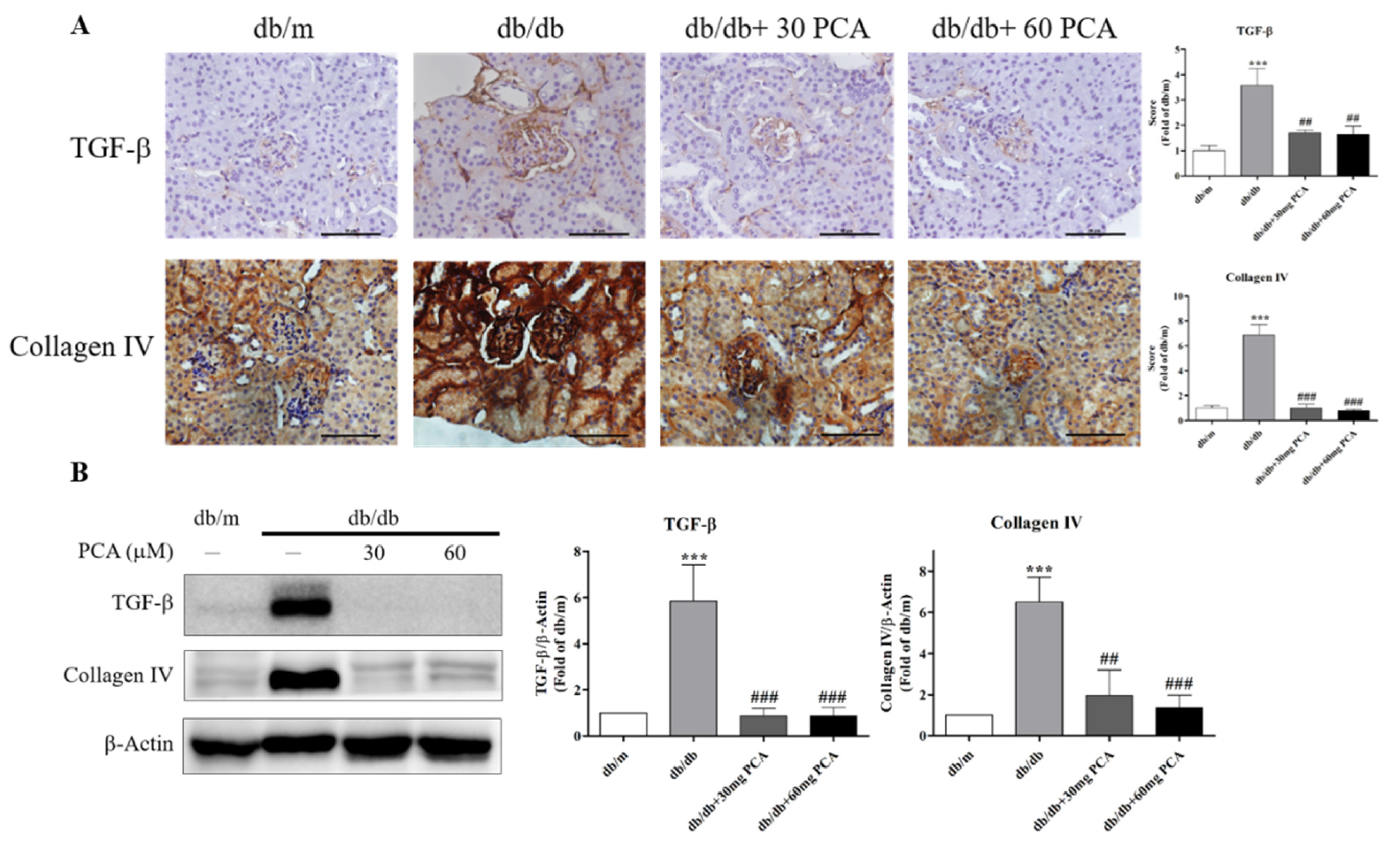
2.5. PCA Inhibits Renal Fibrosis in db/db Mouse Kidneys
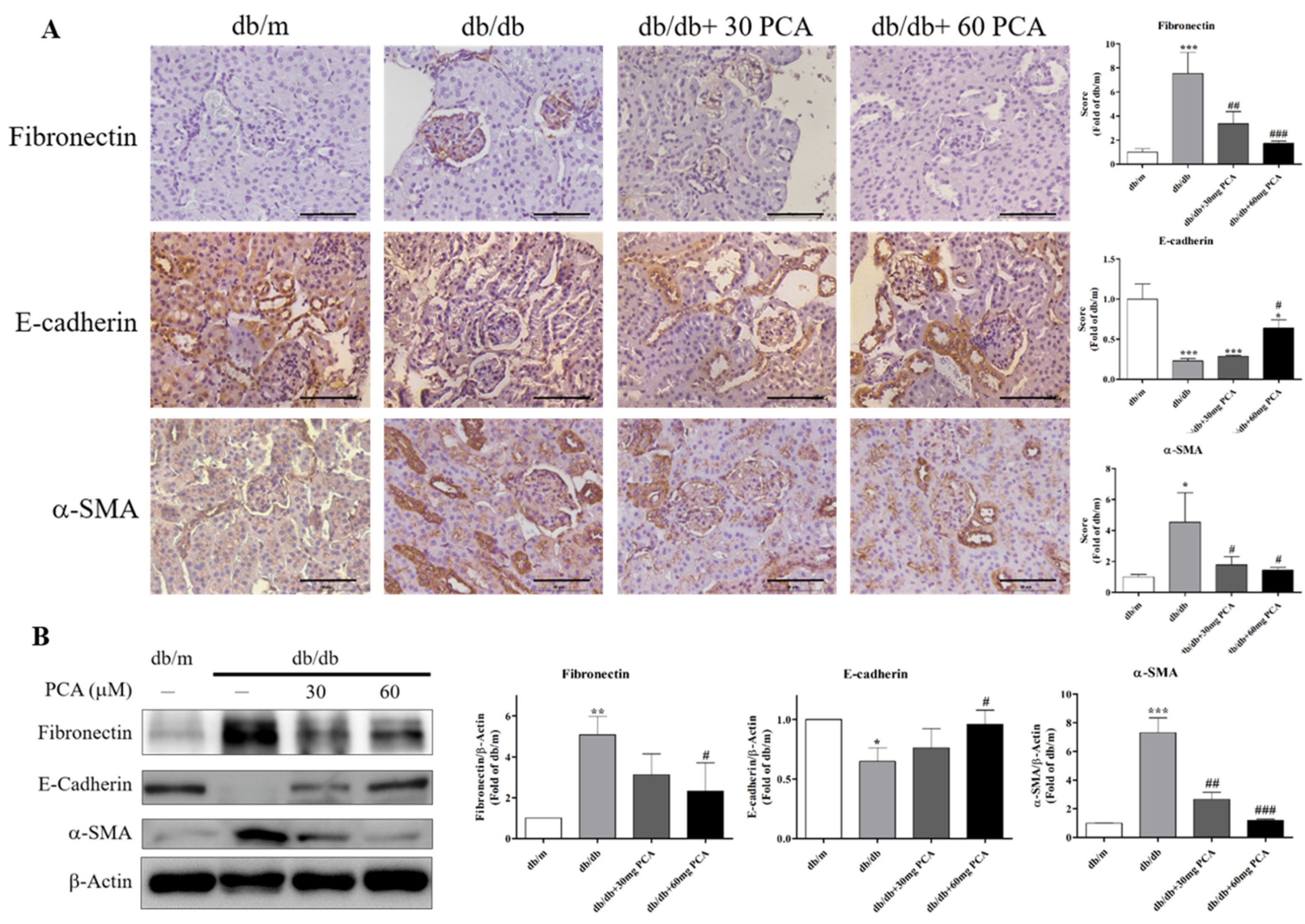
2.6. PCA Inhibits EMT in db/db Mouse Kidneys
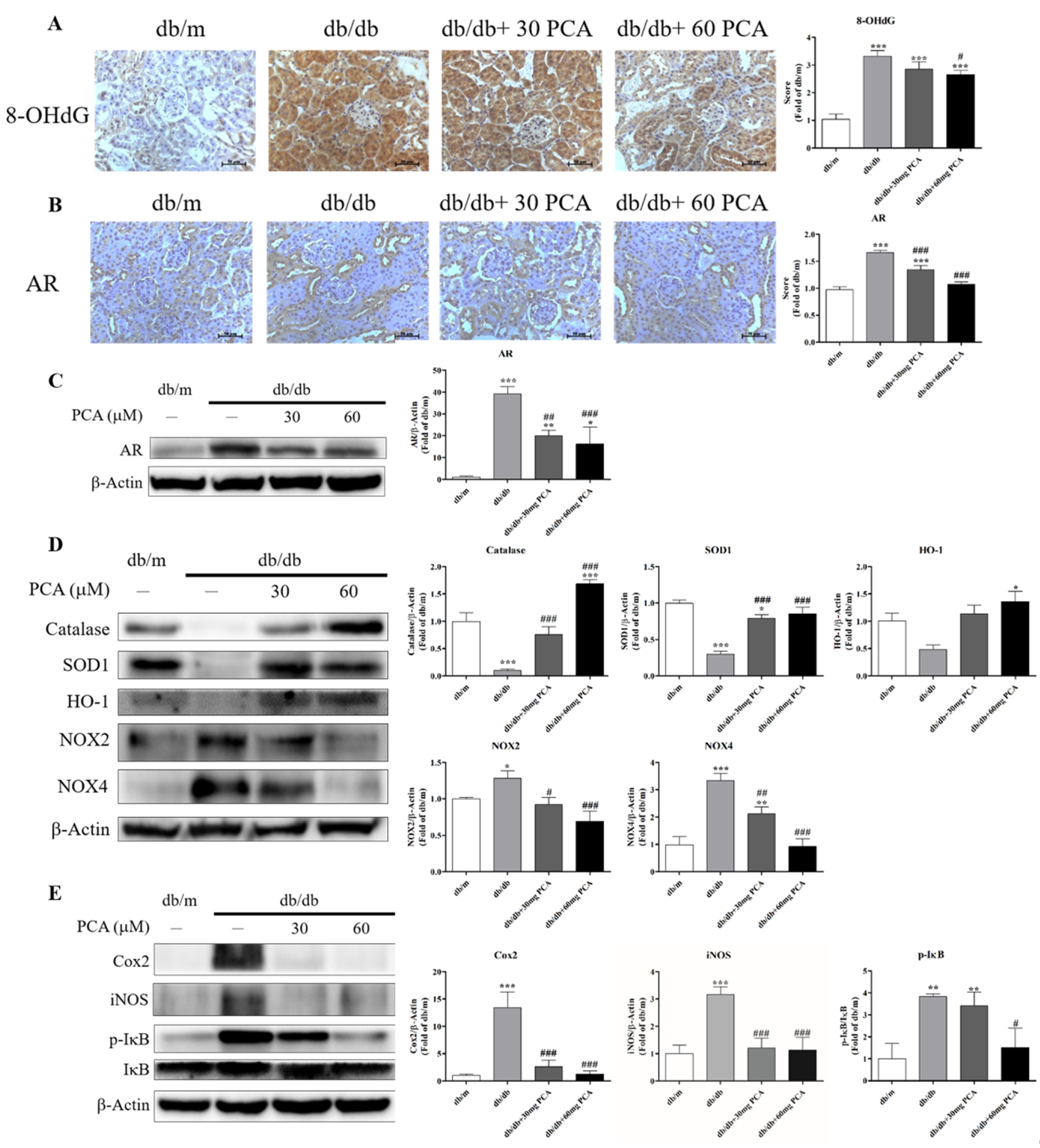
2.7. PCA Inhibits Expression of AR, Oxidative Stress and Inflammation in db/db Mice Kidneys
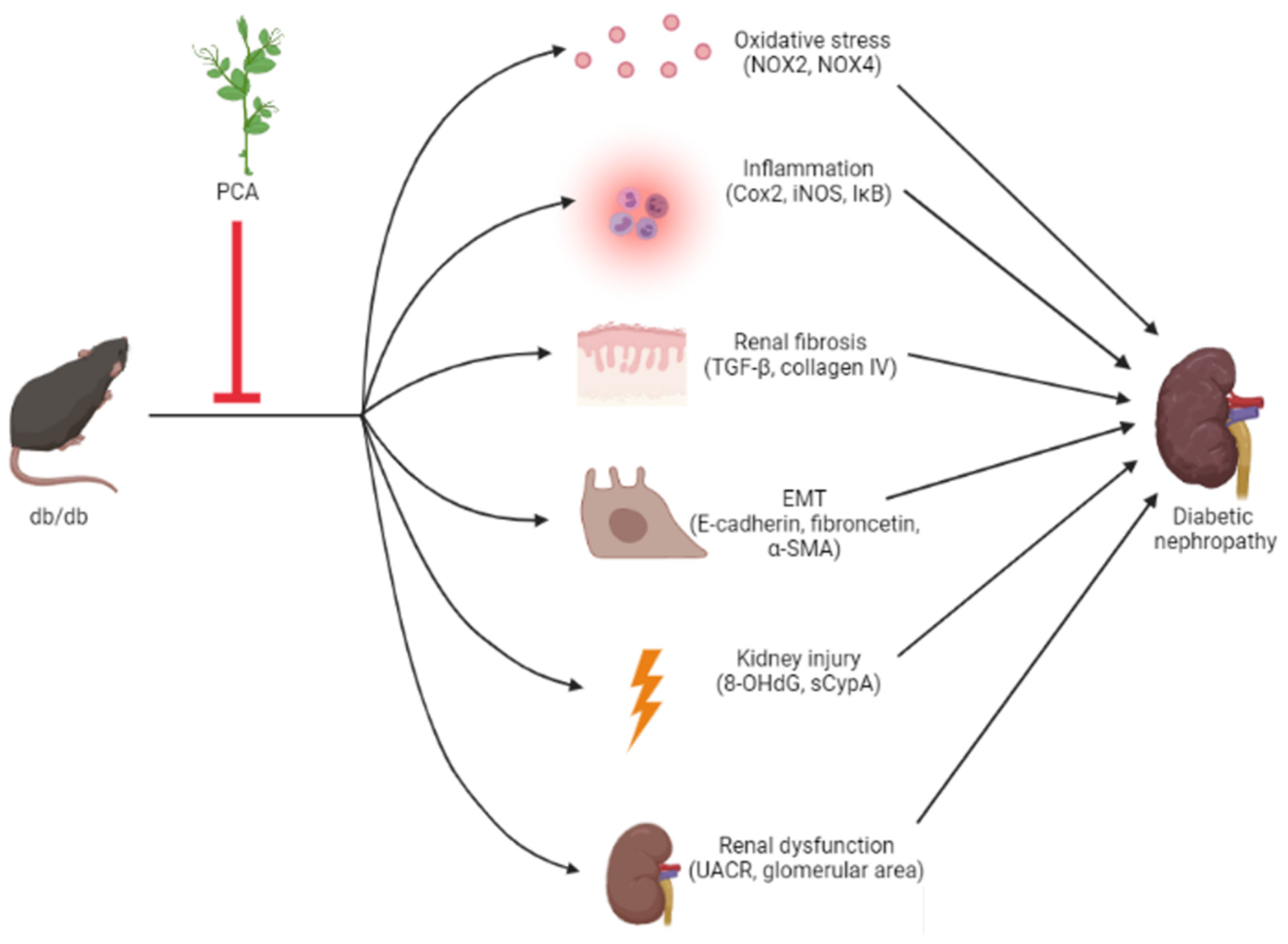
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals and Treatment (Type 2 Diabetic Animal Model)
5.2. Measurement of Plasma Glucose Levels
5.3. Renal Function Evaluation
5.4. 24 h Urinary Secreted Cyclophilin A and 8-Hydroxy-2′-Deoxyguanosine Detection
5.5. Protein Extraction and Western Blot Analysis
5.6. Histological Analysis
5.7. Immunohistochemical Staining
5.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boor, P.; Floege, J. The renal (myo-)fibroblast: A heterogeneous group of cells. Nephrol. Dial. Transplant. 2012, 27, 3027–3036. [Google Scholar] [CrossRef] [Green Version]
- Chronic Kidney Disease Prognosis Consortium; Matsushita, K.; van der Velde, M.; Astor, B.C.; Woodward, M.; Levey, A.S.; de Jong, P.E.; Coresh, J.; Gansevoort, R.T. Association of estimated glomerular filtration rate and albuminuria with all-cause and cardiovascular mortality in general population cohorts: A collaborative meta-analysis. Lancet 2010, 375, 2073–2081. [Google Scholar] [CrossRef] [Green Version]
- Locatelli, F.; Nissenson, A.R.; Barrett, B.J.; Walker, R.G.; Wheeler, D.C.; Eckardt, K.U.; Lameire, N.H.; Eknoyan, G. Clinical practice guidelines for anemia in chronic kidney disease: Problems and solutions. A position statement from Kidney Disease: Improving Global Outcomes (KDIGO). Kidney Int. 2008, 74, 1237–1240. [Google Scholar] [CrossRef] [PubMed]
- Naylor, K.L.; Prior, J.; Garg, A.X.; Berger, C.; Langsetmo, L.; Adachi, J.D.; Goltzman, D.; Kovacs, C.S.; Josse, R.G.; Leslie, W.D. Trabecular Bone Score and Incident Fragility Fracture Risk in Adults with Reduced Kidney Function. Clin. J. Am. Soc. Nephrol. 2016, 11, 2032–2040. [Google Scholar] [CrossRef] [PubMed]
- Ritter, C.S.; Slatopolsky, E. Phosphate Toxicity in CKD: The Killer among Us. Clin. J. Am. Soc. Nephrol. 2016, 11, 1088–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merono, O.; Cladellas, M.; Ribas-Barquet, N.; Poveda, P.; Recasens, L.; Bazan, V.; Garcia-Garcia, C.; Ivern, C.; Enjuanes, C.; Orient, S.; et al. Iron Deficiency Is a Determinant of Functional Capacity and Health-related Quality of Life 30 Days After an Acute Coronary Syndrome. Rev. Esp. Cardiol. 2017, 70, 363–370. [Google Scholar] [CrossRef]
- McClung, J.P.; Murray-Kolb, L.E. Iron nutrition and premenopausal women: Effects of poor iron status on physical and neuropsychological performance. Annu. Rev. Nutr. 2013, 33, 271–288. [Google Scholar] [CrossRef] [PubMed]
- Lim, A. Diabetic nephropathy—Complications and treatment. Int. J. Nephrol. Renovasc. Dis. 2014, 7, 361–381. [Google Scholar] [CrossRef] [Green Version]
- Schieppati, A.; Remuzzi, G. Chronic renal diseases as a public health problem: Epidemiology, social, and economic implications. Kidney Int. Suppl. 2005, 68, S7–S10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuta, T.; Saito, T.; Ootaka, T.; Soma, J.; Obara, K.; Abe, K.; Yoshinaga, K. The role of macrophages in diabetic glomerulosclerosis. Am. J. Kidney Dis. 1993, 21, 480–485. [Google Scholar] [CrossRef]
- Nguyen, D.; Ping, F.; Mu, W.; Hill, P.; Atkins, R.C.; Chadban, S.J. Macrophage accumulation in human progressive diabetic nephropathy. Nephrology (Carlton) 2006, 11, 226–231. [Google Scholar] [CrossRef]
- Yonemoto, S.; Machiguchi, T.; Nomura, K.; Minakata, T.; Nanno, M.; Yoshida, H. Correlations of tissue macrophages and cytoskeletal protein expression with renal fibrosis in patients with diabetes mellitus. Clin. Exp. Nephrol. 2006, 10, 186–192. [Google Scholar] [CrossRef]
- Pelletier, K.; Bonnefoy, A.; Chapdelaine, H.; Pichette, V.; Lejars, M.; Madore, F.; Brachemi, S.; Troyanov, S. Clinical Value of Complement Activation Biomarkers in Overt Diabetic Nephropathy. Kidney Int. Rep. 2019, 4, 797–805. [Google Scholar] [CrossRef] [Green Version]
- Nosadini, R.; Velussi, M.; Brocco, E.; Bruseghin, M.; Abaterusso, C.; Saller, A.; Dalla Vestra, M.; Carraro, A.; Bortoloso, E.; Sambataro, M.; et al. Course of renal function in type 2 diabetic patients with abnormalities of albumin excretion rate. Diabetes 2000, 49, 476–484. [Google Scholar] [CrossRef] [Green Version]
- Verhave, J.C.; Bouchard, J.; Goupil, R.; Pichette, V.; Brachemi, S.; Madore, F.; Troyanov, S. Clinical value of inflammatory urinary biomarkers in overt diabetic nephropathy: A prospective study. Diabetes Res. Clin. Pract. 2013, 101, 333–340. [Google Scholar] [CrossRef]
- Nadkarni, G.N.; Rao, V.; Ismail-Beigi, F.; Fonseca, V.A.; Shah, S.V.; Simonson, M.S.; Cantley, L.; Devarajan, P.; Parikh, C.R.; Coca, S.G. Association of Urinary Biomarkers of Inflammation, Injury, and Fibrosis with Renal Function Decline: The ACCORD Trial. Clin. J. Am. Soc. Nephrol. 2016, 11, 1343–1352. [Google Scholar] [CrossRef] [Green Version]
- Camilla, R.; Brachemi, S.; Pichette, V.; Cartier, P.; Laforest-Renald, A.; MacRae, T.; Madore, F.; Troyanov, S. Urinary monocyte chemotactic protein 1: Marker of renal function decline in diabetic and nondiabetic proteinuric renal disease. J. Nephrol. 2011, 24, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Garud, M.S.; Kulkarni, Y.A. Hyperglycemia to nephropathy via transforming growth factor beta. Curr. Diabetes Rev. 2014, 10, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Nakerakanti, S.; Trojanowska, M. The Role of TGF-beta Receptors in Fibrosis. Open Rheumatol. J. 2012, 6, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Zhang, Y.; Luo, Y.; Wang, Z.; Bi, S.; Song, D.; Dai, Y.; Wang, T.; Qiu, L.; Wen, L.; et al. Aldose reductase regulates miR-200a-3p/141-3p to coordinate Keap1-Nrf2, Tgfbeta1/2, and Zeb1/2 signaling in renal mesangial cells and the renal cortex of diabetic mice. Free Radic. Biol. Med. 2014, 67, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Iso, K.; Tada, H.; Kuboki, K.; Inokuchi, T. Long-term effect of epalrestat, an aldose reductase inhibitor, on the development of incipient diabetic nephropathy in Type 2 diabetic patients. J. Diabetes Complicat. 2001, 15, 241–244. [Google Scholar] [CrossRef]
- Ikeda, T.; Iwata, K.; Tanaka, Y. Long-term effect of epalrestat on cardiac autonomic neuropathy in subjects with non-insulin dependent diabetes mellitus. Diabetes Res. Clin. Pract. 1999, 43, 193–198. [Google Scholar] [CrossRef]
- He, J.; Gao, H.X.; Yang, N.; Zhu, X.D.; Sun, R.B.; Xie, Y.; Zeng, C.H.; Zhang, J.W.; Wang, J.K.; Ding, F.; et al. The aldose reductase inhibitor epalrestat exerts nephritic protection on diabetic nephropathy in db/db mice through metabolic modulation. Acta Pharmacol. Sin. 2019, 40, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Kim, N.H.; Lee, S.W.; Lee, Y.M.; Jang, D.S.; Kim, J.S. Effect of protocatechualdehyde on receptor for advanced glycation end products and TGF-beta1 expression in human lens epithelial cells cultured under diabetic conditions and on lens opacity in streptozotocin-diabetic rats. Eur. J. Pharmacol. 2007, 569, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Shim, S.H.; Kim, J.S.; Shin, K.H.; Kang, S.S. Aldose reductase inhibitors from the fruiting bodies of Ganoderma applanatum. Biol. Pharm. Bull. 2005, 28, 1103–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byun, J.W.; Hwang, S.; Kang, C.W.; Kim, J.H.; Chae, M.K.; Yoon, J.S.; Lee, E.J. Therapeutic Effect of Protocatechuic Aldehyde in an In Vitro Model of Graves’ Orbitopathy. Investig. Ophthalmol. Vis. Sci. 2016, 57, 4055–4062. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Li, J.; Tan, R.; He, X.; Lin, X.; Zhong, X.; Fan, J.; Wang, L. Protocatechualdehyde attenuates obstructive nephropathy through inhibiting lncRNA9884 induced inflammation. Phytother. Res. 2021, 35, 1521–1533. [Google Scholar] [CrossRef]
- Wan, Y.J.; Guo, Q.; Liu, D.; Jiang, Y.; Zeng, K.W.; Tu, P.F. Protocatechualdehyde reduces myocardial fibrosis by directly targeting conformational dynamics of collagen. Eur. J. Pharmacol. 2019, 855, 183–191. [Google Scholar] [CrossRef]
- Zhang, L.; Ji, Y.; Kang, Z.; Lv, C.; Jiang, W. Protocatechuic aldehyde ameliorates experimental pulmonary fibrosis by modulating HMGB1/RAGE pathway. Toxicol. Appl. Pharmacol. 2015, 283, 50–56. [Google Scholar] [CrossRef]
- Tsai, S.F.; Su, C.W.; Wu, M.J.; Chen, C.H.; Fu, C.P.; Liu, C.S.; Hsieh, M. Urinary Cyclophilin A as a New Marker for Diabetic Nephropathy: A Cross-Sectional Analysis of Diabetes Mellitus. Medicine 2015, 94, e1802. [Google Scholar] [CrossRef]
- Park, I.S.; Kiyomoto, H.; Abboud, S.L.; Abboud, H.E. Expression of transforming growth factor-beta and type IV collagen in early streptozotocin-induced diabetes. Diabetes 1997, 46, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Border, W.A.; Huang, Y.; Noble, N.A. TGF-beta isoforms in renal fibrogenesis. Kidney Int. 2003, 64, 844–856. [Google Scholar] [CrossRef] [Green Version]
- Giunti, S.; Barit, D.; Cooper, M.E. Mechanisms of diabetic nephropathy: Role of hypertension. Hypertension 2006, 48, 519–526. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Das, A.; Sen, S. Extracellular matrix density promotes EMT by weakening cell-cell adhesions. Mol. Biosyst. 2014, 10, 838–850. [Google Scholar] [CrossRef]
- Davidson, S.M. Endothelial mitochondria and heart disease. Cardiovasc. Res. 2010, 88, 58–66. [Google Scholar] [CrossRef]
- Ji, B.; Yuan, K.; Li, J.; Ku, B.J.; Leung, P.S.; He, W. Protocatechualdehyde restores endothelial dysfunction in streptozotocin-induced diabetic rats. Ann. Transl. Med. 2021, 9, 711. [Google Scholar] [CrossRef]
- Kong, B.S.; Cho, Y.H.; Lee, E.J. G protein-coupled estrogen receptor-1 is involved in the protective effect of protocatechuic aldehyde against endothelial dysfunction. PLoS ONE 2014, 9, e113242. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, H.; Shikata, K.; Hirata, K.; Akiyama, K.; Matsuda, M.; Kushiro, M.; Shikata, Y.; Miyatake, N.; Miyasaka, M.; Makino, H. Increased expression of intercellular adhesion molecule-1 (ICAM-1) in diabetic rat glomeruli: Glomerular hyperfiltration is a potential mechanism of ICAM-1 upregulation. Diabetes 1997, 46, 2075–2081. [Google Scholar] [CrossRef]
- Sango, K.; Suzuki, T.; Yanagisawa, H.; Takaku, S.; Hirooka, H.; Tamura, M.; Watabe, K. High glucose-induced activation of the polyol pathway and changes of gene expression profiles in immortalized adult mouse Schwann cells IMS32. J. Neurochem. 2006, 98, 446–458. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Mizuno, K.; Yashima, S.; Watanabe, K.; Taniko, K.; Yabe-Nishimura, C. Characterization of polyol pathway in Schwann cells isolated from adult rat sciatic nerves. J. Neurosci. Res. 1999, 57, 495–503. [Google Scholar] [CrossRef]
- Ferraz, M.; Ishii-Iwamoto, E.L.; Batista, M.R.; Brunaldi, K.; Bazotte, R.B. Sorbitol accumulation in rats kept on diabetic condition for short and prolonged periods. Zhongguo Yao Li Xue Bao 1997, 18, 309–311. [Google Scholar] [PubMed]
- Kinoshita, J.H. Cataracts in galactosemia. The Jonas S. Friedenwald Memorial Lecture. Invest. Ophthalmol. 1965, 4, 786–799. [Google Scholar] [PubMed]
- Kinoshita, J.H. Mechanisms initiating cataract formation. Proctor Lecture. Invest. Ophthalmol. 1974, 13, 713–724. [Google Scholar]
- Greene, D.A.; Lattimer, S.A.; Sima, A.A. Sorbitol, phosphoinositides, and sodium-potassium-ATPase in the pathogenesis of diabetic complications. N. Engl. J. Med. 1987, 316, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, J.H.; Nishimura, C. The involvement of aldose reductase in diabetic complications. Diabetes Metab. Rev. 1988, 4, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Larkins, R.G.; Dunlop, M.E. The link between hyperglycaemia and diabetic nephropathy. Diabetologia 1992, 35, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.X.; Zhang, Y.D.; Shen, J.P.; Wu, H.W.; Zhu, X.; Li, L.M.; Qiu, J.; Jiang, S.J.; Zheng, X.G. Protective effects of bendazac lysine on early experimental diabetic nephropathy in rats. Acta. Pharmacol. Sin. 2005, 26, 721–728. [Google Scholar] [CrossRef] [Green Version]
- Oates, P.J. Aldose reductase inhibitors and diabetic kidney disease. Curr. Opin. Investig. Drugs 2010, 11, 402–417. [Google Scholar]
- Kasajima, H.; Yamagishi, S.; Sugai, S.; Yagihashi, N.; Yagihashi, S. Enhanced in situ expression of aldose reductase in peripheral nerve and renal glomeruli in diabetic patients. Virchows Arch. 2001, 439, 46–54. [Google Scholar] [CrossRef]
- Corder, C.N.; Braughler, J.M.; Culp, P.A. Quantitative histochemistry of the sorbitol pathway in glomeruli and small arteries of human diabetic kidney. Folia Histochem. Cytochem. 1979, 17, 137–145. [Google Scholar]
- Sands, J.M.; Terada, Y.; Bernard, L.M.; Knepper, M.A. Aldose reductase activities in microdissected rat renal tubule segments. Am. J. Physiol. 1989, 256, F563–F569. [Google Scholar] [CrossRef]
- Nakayama, T.; Kosugi, T.; Gersch, M.; Connor, T.; Sanchez-Lozada, L.G.; Lanaspa, M.A.; Roncal, C.; Perez-Pozo, S.E.; Johnson, R.J.; Nakagawa, T. Dietary fructose causes tubulointerstitial injury in the normal rat kidney. Am. J. Physiol. Renal. Physiol. 2010, 298, F712–F720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanaspa, M.A.; Ishimoto, T.; Cicerchi, C.; Tamura, Y.; Roncal-Jimenez, C.A.; Chen, W.; Tanabe, K.; Andres-Hernando, A.; Orlicky, D.J.; Finol, E.; et al. Endogenous fructose production and fructokinase activation mediate renal injury in diabetic nephropathy. J. Am. Soc. Nephrol. 2014, 25, 2526–2538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gersch, M.S.; Mu, W.; Cirillo, P.; Reungjui, S.; Zhang, L.; Roncal, C.; Sautin, Y.Y.; Johnson, R.J.; Nakagawa, T. Fructose, but not dextrose, accelerates the progression of chronic kidney disease. Am. J. Physiol. Renal. Physiol. 2007, 293, F1256–F1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, Y.-T.; Chung, M.-C.; Hsieh, C.-C.; Shieh, J.-J.; Wu, M.-J. Evaluation of the Therapeutic Effects of Protocatechuic Aldehyde in Diabetic Nephropathy. Toxins 2021, 13, 560. https://doi.org/10.3390/toxins13080560
Chang Y-T, Chung M-C, Hsieh C-C, Shieh J-J, Wu M-J. Evaluation of the Therapeutic Effects of Protocatechuic Aldehyde in Diabetic Nephropathy. Toxins. 2021; 13(8):560. https://doi.org/10.3390/toxins13080560
Chicago/Turabian StyleChang, Yu-Teng, Mu-Chi Chung, Chang-Chi Hsieh, Jeng-Jer Shieh, and Ming-Ju Wu. 2021. "Evaluation of the Therapeutic Effects of Protocatechuic Aldehyde in Diabetic Nephropathy" Toxins 13, no. 8: 560. https://doi.org/10.3390/toxins13080560
APA StyleChang, Y.-T., Chung, M.-C., Hsieh, C.-C., Shieh, J.-J., & Wu, M.-J. (2021). Evaluation of the Therapeutic Effects of Protocatechuic Aldehyde in Diabetic Nephropathy. Toxins, 13(8), 560. https://doi.org/10.3390/toxins13080560