Ochratoxin A Defective Aspergillus carbonarius Mutants as Potential Biocontrol Agents




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
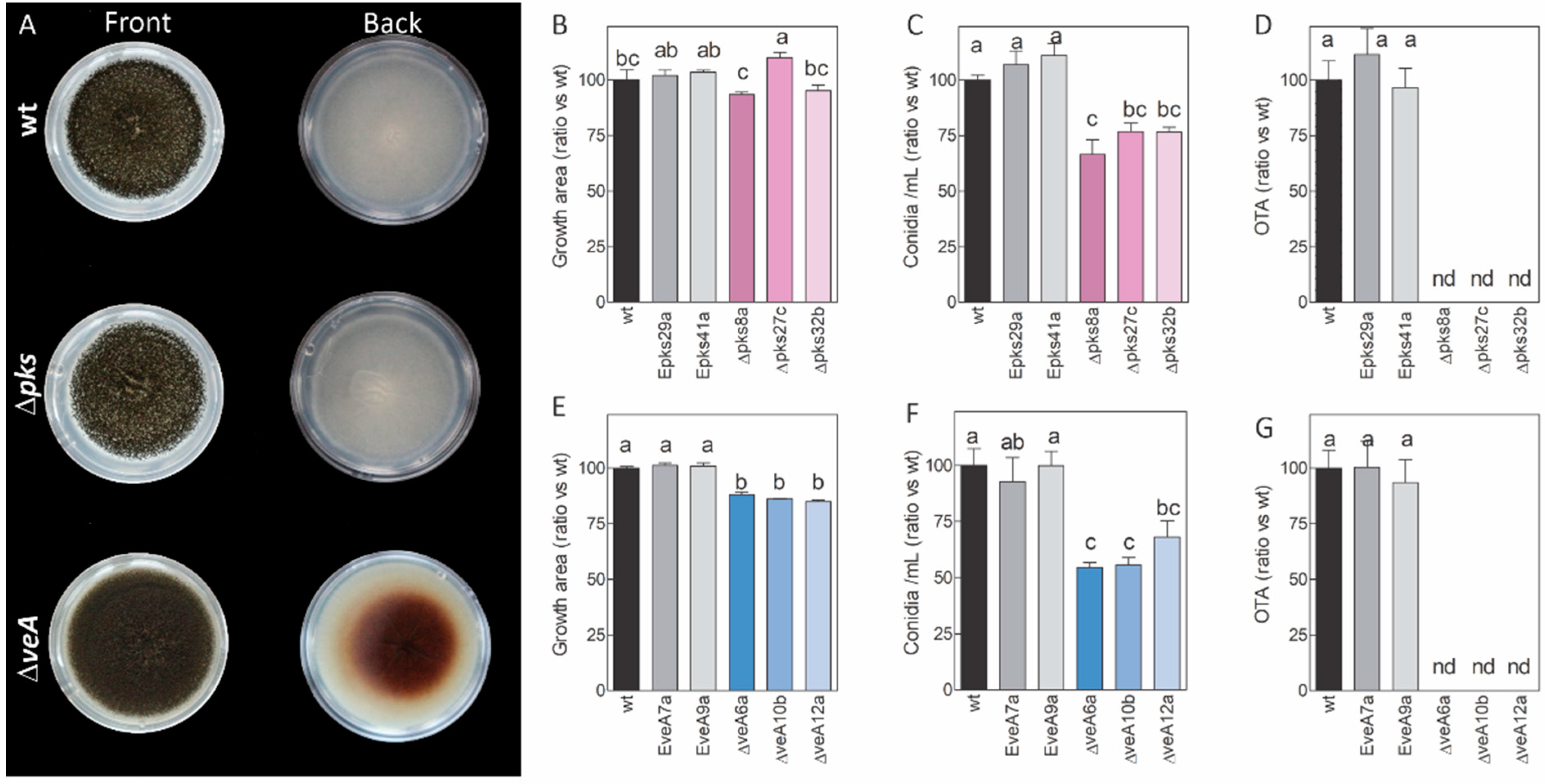
2.1. Construction and Characterization of Knockout Mutants
2.2. Different Stresses Did Not Affect the Growth of the Knockout Mutants
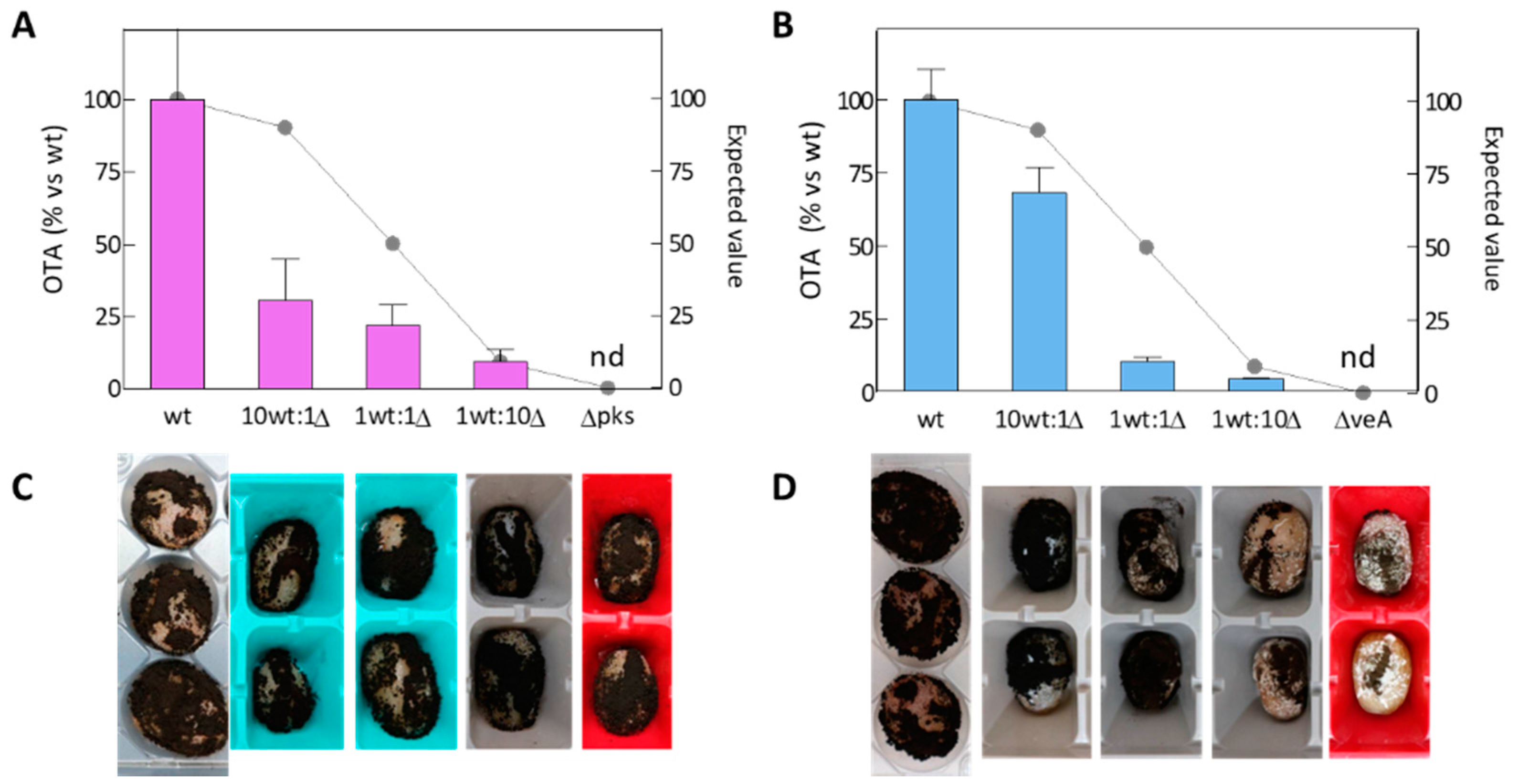
2.3. Competitiveness of Mutants Δpks and ΔveA during In Vitro Growth
2.4. Competitiveness of the Δpks and ΔveA Mutants during Grape Berry Infection
3. Discussion
4. Materials and Methods
4.1. Fungal Strains and Culture Conditions
4.2. Construction and Verification of the A. carbonarius Knockout Mutants
4.3. Characterization of the Knockout Mutants: Mycelial Growth, Conidiation and Growth under Stress Conditions
4.4. In Vitro Competition Assays
4.5. Artificial Inoculation of Grape Berries
4.6. OTA Quantification
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [Green Version]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide Contamination of Food-Crops with Mycotoxins: Validity of the Widely Cited ‘FAO Estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef]
- Liu, Y.; Yamdeu, J.H.G.; Gong, Y.Y.; Orfila, C. A Review of Postharvest Approaches to Reduce Fungal and Mycotoxin Contamination of Foods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1521–1560. [Google Scholar] [CrossRef]
- EPA. US Environmental Protection Agency Office of Pesticide Programs. In Biopesticide Registration Action Document: Aspergillus flavus AF36; PC Code 006456; EPA: Washington, DC, USA, 2003. [Google Scholar]
- EPA. US Environmental Protection Agency Office of Pesticide Programs. In Biopesticide Registration Action Document Aspergillus flavus (NRRL 21882) (PC Code 006500); EPA: Washington, DC, USA, 2004. [Google Scholar]
- Dorner, J.; Lamb, M. Development and Commercial Use of Afla-Guard®, an Aflatoxin Biocontrol Agent. Mycotoxin Res. 2006, 22, 33–38. [Google Scholar] [CrossRef]
- Mauro, A.; Garcia-Cela, E.; Pietri, A.; Cotty, P.J.; Battilani, P. Biological Control Products for Aflatoxin Prevention in Italy: Commercial Field Evaluation of Atoxigenic Aspergillus flavus Active Ingredients. Toxins 2018, 10, E30. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, K.C.; Cotty, P.J. An Isolate of Aspergillus flavus Used to Reduce Aflatoxin Contamination in Cottonseed Has a Defective Polyketide Synthase Gene. Appl. Microbiol. Biotechnol. 2004, 65, 473–478. [Google Scholar] [CrossRef]
- Kolawole, O.; Meneely, J.; Petchkongkaew, A.; Elliott, C. A Review of Mycotoxin Biosynthetic Pathways: Associated Genes and Their Expressions under the Influence of Climatic Factors. Fungal Biol. Rev. 2021, 37, 8–26. [Google Scholar] [CrossRef]
- Perrone, G.; Ferrara, M.; Medina, A.; Pascale, M.; Magan, N. Toxigenic Fungi and Mycotoxins in a Climate Change Scenario: Ecology, Genomics, Distribution, Prediction and Prevention of the Risk. Microorganisms 2020, 8, 1496. [Google Scholar] [CrossRef]
- Gil-Serna, J.; García-Díaz, M.; González-Jaén, M.T.; Vázquez, C.; Patiño, B. Description of an Orthologous Cluster of Ochratoxin A Biosynthetic Genes in Aspergillus and Penicillium Species. A Comparative Analysis. Int. J. Food Microbiol. 2018, 268, 35–43. [Google Scholar] [CrossRef]
- Ferrara, M.; Gallo, A.; Perrone, G.; Magistà, D.; Baker, S.E. Comparative Genomic Analysis of Ochratoxin A Biosynthetic Cluster in Producing Fungi: New Evidence of a Cyclase Gene Involvement. Front. Microbiol. 2020, 11, 581309. [Google Scholar] [CrossRef]
- Castellá, G.; Bragulat, M.R.; Puig, L.; Sanseverino, W.; Cabañes, F.J. Genomic Diversity in Ochratoxigenic and Non Ochratoxigenic Strains of Aspergillus carbonarius. Sci. Rep. 2018, 8, 5439. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, L.; Wu, F.; Liu, F.; Wang, Q.; Zhang, X.; Selvaraj, J.N.; Zhao, Y.; Xing, F.; Yin, W.-B.; et al. A Consensus Ochratoxin A Biosynthetic Pathway: Insights from the Genome Sequence of Aspergillus ochraceus and a Comparative Genomic Analysis. Appl. Environ. Microbiol. 2018, 84, e01009-18. [Google Scholar] [CrossRef] [Green Version]
- Gallo, A.; Knox, B.P.; Bruno, K.S.; Solfrizzo, M.; Baker, S.E.; Perrone, G. Identification and Characterization of the Polyketide Synthase Involved in Ochratoxin A Biosynthesis in Aspergillus carbonarius. Int. J. Food Microbiol. 2014, 179, 10–17. [Google Scholar] [CrossRef]
- Maor, U.; Barda, O.; Sadhasivam, S.; Bi, Y.; Levin, E.; Zakin, V.; Prusky, D.B.; Sionov, E. Functional Roles of LaeA, Polyketide Synthase, and Glucose Oxidase in the Regulation of Ochratoxin A Biosynthesis and Virulence in Aspergillus carbonarius. Mol. Plant Pathol. 2021, 22, 117–129. [Google Scholar] [CrossRef]
- Gallo, A.; Bruno, K.S.; Solfrizzo, M.; Perrone, G.; Mulè, G.; Visconti, A.; Baker, S.E. New Insight into the Ochratoxin A Biosynthetic Pathway through Deletion of a Nonribosomal Peptide Synthetase Gene in Aspergillus carbonarius. Appl. Environ. Microbiol. 2012, 78, 8208–8218. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, M.; Perrone, G.; Gambacorta, L.; Epifani, F.; Solfrizzo, M.; Gallo, A. Identification of a Halogenase Involved in the Biosynthesis of Ochratoxin A in Aspergillus carbonarius. Appl. Environ. Microbiol. 2016, 82, 5631–5641. [Google Scholar] [CrossRef] [Green Version]
- Gerin, D.; Garrapa, F.; Ballester, A.-R.; González-Candelas, L.; De Miccolis Angelini, R.M.; Faretra, F.; Pollastro, S. Functional Role of Aspergillus carbonarius AcOTAbZIP Gene, a BZIP Transcription Factor within the OTA Gene Cluster. Toxins 2021, 13, 111. [Google Scholar] [CrossRef]
- Ferrara, M.; Gallo, A.; Cervini, C.; Gambacorta, L.; Solfrizzo, M.; Baker, S.E.; Perrone, G. Evidence of the Involvement of a Cyclase Gene in the Biosynthesis of Ochratoxin A in Aspergillus carbonarius. Toxins 2021, 13, 892. [Google Scholar] [CrossRef]
- Crespo-Sempere, A.; Marín, S.; Sanchis, V.; Ramos, A.J. VeA and LaeA Transcriptional Factors Regulate Ochratoxin A Biosynthesis in Aspergillus carbonarius. Int. J. Food Microbiol. 2013, 166, 479–486. [Google Scholar] [CrossRef]
- Cervini, C.; Verheecke-Vaessen, C.; Ferrara, M.; García-Cela, E.; Magistà, D.; Medina, A.; Gallo, A.; Magan, N.; Perrone, G. Interacting Climate Change Factors (CO2 and Temperature Cycles) Effects on Growth, Secondary Metabolite Gene Expression and Phenotypic Ochratoxin A Production by Aspergillus carbonarius Strains on a Grape-Based Matrix. Fungal Biol. 2021, 125, 115–122. [Google Scholar] [CrossRef]
- Assaf, C.E.H.; Snini, S.P.; Tadrist, S.; Bailly, S.; Naylies, C.; Oswald, S.P.; Lorber, S.; Puel, O. Impact of VeA on the Development, Aggressiveness, Dissemination and Secondary Metabolism of Penicillium expansum. Mol. Plant Pathol. 2018, 19, 1971–1983. [Google Scholar] [CrossRef] [Green Version]
- Wiemann, P.; Brown, D.W.; Kleigrewe, K.; Bok, J.W.; Keller, N.P.; Humpf, H.U.; Tudzynski, B. FfVel1 and FfLae1, Components of a Velvet-like Complex in Fusarium fujikuroi, Affect Differentiation, Secondary Metabolism and Virulence. Mol. Microbiol. 2010, 77, 972–994. [Google Scholar] [CrossRef] [Green Version]
- Frandsen, R.J.; Andersson, J.A.; Kristensen, M.B.; Giese, H. Efficient Four Fragment Cloning for the Construction of Vectors for Targeted Gene Replacement in Filamentous Fungi. BMC Mol. Biol. 2008, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Dorner, J.W. Biological Control of Aflatoxin Contamination in Corn Using a Nontoxigenic Strain of Aspergillus flavus. J. Food Prot. 2009, 72, 801–804. [Google Scholar] [CrossRef]
- Cotty, P.J.; Bayman, P. Competitive Exclusion of a Toxigenic Strain of Aspergillus flavus by an Atoxigenic Strain. Phytopathology 1993, 83, 1283. [Google Scholar] [CrossRef]
- Lewis, M.H.; Carbone, I.; Luis, J.M.; Payne, G.A.; Bowen, K.L.; Hagan, A.K.; Kemerait, R.; Heiniger, R.; Ojiambo, P.S. Biocontrol Strains Differentially Shift the Genetic Structure of Indigenous Soil Populations of Aspergillus flavus. Front. Microbiol. 2019, 10, 1738. [Google Scholar] [CrossRef] [Green Version]
- Castellá, G.; Bragulat, M.R.; Cigliano, R.A.; Cabañes, F.J. Transcriptome Analysis of Non-Ochratoxigenic Aspergillus carbonarius Strains and Interactions between Some Black Aspergilli Species. Int. J. Food Microbiol. 2020, 317, 108498. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.; Wang, H.; Wang, Y.; Ge, Y.; Ren, X.; Ren, C.; Wang, Y.; Ren, X.; Liu, Y.; Liu, Z. The Role of the VeA Gene in Adjusting Developmental Balance and Environmental Stress Response in Aspergillus cristatus. Fungal Biol. 2018, 122, 952–964. [Google Scholar] [CrossRef]
- Bayram, Ö.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.-J.; Keller, N.P.; Yu, J.-H.; et al. VelB/VeA/LaeA Complex Coordinates Light Signal with Fungal Development and Secondary Metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef]
- Bok, J.W.; Soukup, A.A.; Chadwick, E.; Chiang, Y.-M.; Wang, C.C.C.; Keller, N.P. VeA and MvlA Repression of the Cryptic Orsellinic Acid Gene Cluster in Aspergillus nidulans Involves Histone 3 Acetylation. Mol. Microbiol. 2013, 89, 963–974. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, H.; Wang, Y.; Liu, F.; Li, E.; Ma, J.; Yang, B.; Zhang, C.; Li, L.; Liu, Y. Requirement of LaeA, VeA, and VelB on Asexual Development, Ochratoxin A Biosynthesis, and Fungal Virulence in Aspergillus ochraceus. Front. Microbiol. 2019, 10, 2759. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Chen, H.; Sumarah, M.W.; Gao, Q.; Wang, D.; Zhang, Y. VeA Gene Acts as a Positive Regulator of Conidia Production, Ochratoxin a Biosynthesis, and Oxidative Stress Tolerance in Aspergillus niger. J. Agric. Food Chem. 2018, 66, 13199–13208. [Google Scholar] [CrossRef]
- Amaike, S.; Keller, N.P. Distinct Roles for VeA and LaeA in Development and Pathogenesis of Aspergillus flavus. Eukaryot. Cell 2009, 8, 1051–1060. [Google Scholar] [CrossRef] [Green Version]
- Myung, K.; Zitomer, N.C.; Duvall, M.; Glenn, A.E.; Riley, R.T.; Calvo, A.M. The Conserved Global Regulator VeA Is Necessary for Symptom Production and Mycotoxin Synthesis in Maize Seedlings by Fusarium verticillioides. Plant Pathol. 2012, 61, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Barda, O.; Maor, U.; Sadhasivam, S.; Bi, Y.; Zakin, V.; Prusky, D.; Sionov, E. The PH-Responsive Transcription Factor PacC Governs Pathogenicity and Ochratoxin A Biosynthesis in Aspergillus carbonarius. Front. Microbiol. 2020, 11, 210. [Google Scholar] [CrossRef]
- Baidya, S.; Duran, R.M.; Lohmar, J.M.; Harris-Coward, P.Y.; Cary, J.W.; Hong, S.-Y.; Roze, L.V.; Linz, J.E.; Calvo, A.M. VeA Is Associated with the Response to Oxidative Stress in the Aflatoxin Producer Aspergillus flavus. Eukaryot. Cell 2014, 13, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Dhingra, S.; Andes, D.; Calvo, A.M. VeA Regulates Conidiation, Gliotoxin Production and Protease Activity in the Opportunistic Human Pathogen Aspergillus fumigatus. Eukaryot Cell 2012, 11, 1531–1543. [Google Scholar] [CrossRef] [Green Version]
- Schamann, A.; Schmidt-Heydt, M.; Geisen, R. Analysis of the Competitiveness between a Non-Aflatoxigenic and an Aflatoxigenic Aspergillus flavus Strain on Maize Kernels by Droplet Digital PCR. Mycotoxin Res. 2022, 38, 27–36. [Google Scholar] [CrossRef]
- Garcia-Lopez, M.T.; Luo, Y.; Ortega-Beltran, A.; Jaime, R.; Moral, J.; Michailides, T.J. Quantification of the Aflatoxin Biocontrol Strain Aspergillus flavus AF36 in Soil and in Nuts and Leaves of Pistachio by Real-Time PCR. Plant Dis. 2021, 105, 1657–1665. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New Capabilities and Interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Crespo-Sempere, A.; López-Pérez, M.; Martínez-Culebras, P.V.; González-Candelas, L. Development of a Green Fluorescent Tagged Strain of Aspergillus carbonarius to Monitor Fungal Colonization in Grapes. Int. J. Food Microbiol. 2011, 148, 135–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerin, D.; González-Candelas, L.; Ballester, A.-R.; Pollastro, S.; De Miccolis Angelini, R.; Faretra, F. Functional Characterization of the Alb1 Orthologue Gene in the Ochratoxigenic Fungus Aspergillus carbonarius (AC49 Strain). Toxins 2018, 10, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Kahm, M.; Hasenbrink, G.; Lichtenberg-Fraté, H.; Ludwig, J.; Kschischo, M. Grofit: Fitting Biological Growth Curves with R. J. Stat. Softw. 2010, 33, 1–21. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Llobregat, B.; González-Candelas, L.; Ballester, A.-R. Ochratoxin A Defective Aspergillus carbonarius Mutants as Potential Biocontrol Agents. Toxins 2022, 14, 745. https://doi.org/10.3390/toxins14110745
Llobregat B, González-Candelas L, Ballester A-R. Ochratoxin A Defective Aspergillus carbonarius Mutants as Potential Biocontrol Agents. Toxins. 2022; 14(11):745. https://doi.org/10.3390/toxins14110745
Chicago/Turabian StyleLlobregat, Belén, Luis González-Candelas, and Ana-Rosa Ballester. 2022. "Ochratoxin A Defective Aspergillus carbonarius Mutants as Potential Biocontrol Agents" Toxins 14, no. 11: 745. https://doi.org/10.3390/toxins14110745