Comparative Evaluation of the Capacity of Commercial and Autochthonous Saccharomyces cerevisiae Strains to Remove Ochratoxin A from Natural and Synthetic Grape Juices

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
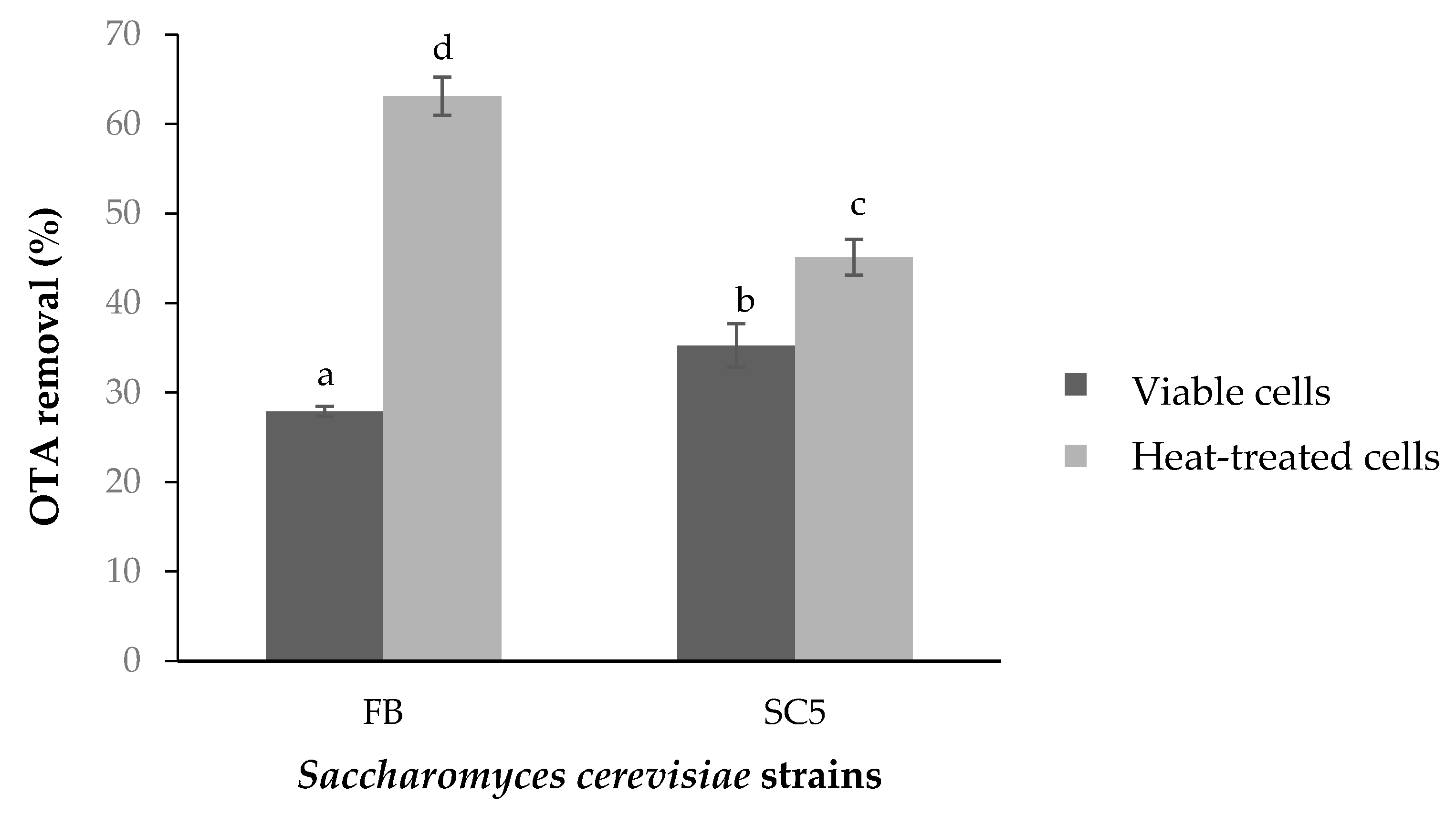
2. Results
2.1. Decontamination of OTA from Synthetic Grape Juice
2.2. Decontamination of OTA from Natural Grape Juice
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals
5.2. Yeast Strains and Culture Conditions
5.3. Preparation of Cell Yeasts
5.4. A. Carbonarius Strain and Preparation of Spore Suspension
5.5. Grape Juices Preparation
5.6. Evaluation of OTA Reduction from Grape Juice
5.7. OTA Extraction and HPLC Analysis
5.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, B.; Li, L.; Geng, H.; Zhang, C.; Wang, G.; Yang, S.; Gao, S.; Zhao, Y.; Xing, F. Inhibitory effect of allyl and benzyl isothiocyanates on ochratoxin a producing fungi in grape and maize. Food Microbiol. 2021, 100, 103865. [Google Scholar] [CrossRef] [PubMed]
- JECFA (Joint FAO/WHO Expert Committee on Food Additives). Safety Evaluation of Certain Mycotoxins in Food; WHO Food Additives Series, No. 59; World Health Organization: Geneva, Switzerland, 2008. [Google Scholar]
- Zimmerli, B.; Dick, R. Ochratoxin A in table wine and grape-juice: Occurrence and risk assessment. Food Addit. Contam. 1996, 13, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Battilani, P.; Pietri, A.; Bertuzzi, T.; Languasco, L.; Giorni, P.; Kozakiewicz, Z. Occurrence of ochratoxin A-producing fungi in grapes grown in Italy. J. Food Prot. 2003, 66, 633–636. [Google Scholar] [CrossRef]
- Bellí, N.; Pardo, E.; Marín, S.; Farré, G.; Ramos, A.J.; Sanchis, V. Occurrence of ochratoxin A and toxigenic potential of fungal isolates from Spanish grapes. J. Sci. Food Agric. 2004, 84, 541–546. [Google Scholar] [CrossRef]
- Dammak, I.; Lasram, S.; Hamdi, Z.; Ben Moussa, O.; Mkadmini Hammi, K.; Trigui, I.; Houissa, H.; Mliki, A.; Hassouna, M. In Vitro Antifungal and Anti-Ochratoxigenic Activities of Aloe vera gel against Aspergillus Carbonarius isolated from grapes. Ind. Crops Prod. 2018, 123, 416–423. [Google Scholar] [CrossRef]
- Lasram, S.; Bellí, N.; Chebil, S.; Nahla, Z.; Ahmed, M.; Sanchis, V.; Ghorbel, A. occurrence of ochratoxigenic fungi and ochratoxin A in grapes from a Tunisian Vineyard. Int. J. Food Microbiol. 2007, 114, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Lasram, S.; Oueslati, S.; Mliki, A.; Ghorbel, A.; Silar, P.; Chebil, S. Ochratoxin A and ochratoxigenic black Aspergillus species in Tunisian grapes cultivated in different geographic areas. Food Control 2012, 25, 75–80. [Google Scholar] [CrossRef]
- European Commission. The 2005 Projections of Age-Related Expenditure (2004-50) for the EU-25 Member States: Underlying Assumptions and Projection Methodologies 230; European Commission: Brussels, Belgium, 2006. [Google Scholar]
- Dammak, I.; Hamdi, Z.; Kammoun El Euch, S.; Zemni, H.; Mliki, A.; Hassouna, M.; Lasram, S. Evaluation of antifungal and anti-ochratoxigenic activities of Salvia Officinalis, Lavandula Dentata and Laurus Nobilis essential oils and a major monoterpene constituent 1,8-cineole against Aspergillus Carbonarius. Ind. Crops Prod. 2019, 128, 85–93. [Google Scholar] [CrossRef]
- Park, D.L.; Njapau, H.; Boutrif, E. Minimizing risks posed by mycotoxins utilizing the HACCP concept. Food Nutr. Agric. 1999, 22, 49–54. [Google Scholar]
- Anli, E.; Bayram, M. Ochratoxin A in wines. Food Rev. Int. 2009, 25, 214–232. [Google Scholar] [CrossRef]
- Amézqueta, S.; González-Peñas, E.; Murillo-Arbizu, M.; López de Cerain, A. Ochratoxin A decontamination: A review. Food Control 2009, 20, 326–333. [Google Scholar] [CrossRef]
- Shetty, P.H.; Jespersen, L. Saccharomyces cerevisiae and lactic acid bacteria as potential mycotoxin decontaminating agents. Trends Food Sci. Technol. 2006, 17, 48–55. [Google Scholar] [CrossRef]
- Piotrowska, M. The adsorption of ochratoxin a by Lactobacillus species. Toxins 2014, 6, 2826–2839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunez, Y.P.; Pueyo, E.; Carrascosa, A.V.; Martínez-Rodríguez, A.J. Effects of aging and heat treatment on whole yeast cells and yeast cell walls and on adsorption of ochratoxin a in a wine model system. J. Food Prot. 2008, 71, 1496–1499. [Google Scholar] [CrossRef] [PubMed]
- Caridi, A.; Galvano, F.; Tafuri, A.; Ritieni, A. Ochratoxin A removal during winemaking. Enzym. Microb. Technol. 2006, 40, 122–126. [Google Scholar] [CrossRef]
- Angioni, A.; Caboni, P.; Garau, A.; Farris, A.; Orro, D.; Budroni, M.; Cabras, P. In vitro interaction between ochratoxin A and different strains of Saccharomyces cerevisiae and Kloeckera apiculata. J. Agric. Food Chem. 2007, 55, 2043–2048. [Google Scholar] [CrossRef]
- Moruno, E.G.; Sanlorenzo, C.; Boccaccino, B.; di Stefano, R. Treatment with yeast to reduce the concentration of ochratoxin A in red wine. Am. J. Enol. Vitic. 2005, 56, 73–76. [Google Scholar]
- Fiori, S.; Urgeghe, P.P.; Hammami, W.; Razzu, S.; Jaoua, S.; Migheli, Q. Biocontrol activity of four non- and low-fermenting yeast strains against Aspergillus carbonarius and their ability to remove ochratoxin A from grape juice. Int. J. Food Microbiol. 2014, 189, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Ringot, D.; Lerzy, B.; Bonhoure, J.P.; Auclair, E.; Oriol, E.; Larondelle, Y. Effect of temperature on in vitro ochratoxin A biosorption onto yeast cell wall derivatives. Process Biochem. 2005, 40, 3008–3016. [Google Scholar] [CrossRef]
- Ringot, D.; Lerzy, B.; Chaplain, K.; Bonhoure, J.P.; Auclair, E.; Larondelle, Y. In vitro biosorption of ochratoxin A on the yeast industry by-products: Comparison of isotherm models. Bioresour. Technol. 2007, 98, 1812–1821. [Google Scholar] [CrossRef]
- Nguyen, T.D. Protection de la Levure Saccharomyces cerevisiae Par un Système Biopolymérique Multicouche: Effet sur Son Activité Métabolique en Réponse Aux Conditions de l ’Environnement. Thèse de Doctorat, Université de Bourgogne, Dijon, France. 2016. Available online: https://tel.archives-ouvertes.fr/tel-01510068 (accessed on 19 April 2017).
- Bueno, D.J.; Sar Casale, C.H.; Pizzolitto, R.P.; Salvano, M.A.; Oliver, G. Physical adsorption of aflatoxin B 1 by lactic acid bacteria and Saccharomyces cerevisiae: A theoretical model. J. Food Prot. 2007, 70, 2148–2154. [Google Scholar] [CrossRef] [PubMed]
- Pizzolitto, R.P.; Armando, M.R.; Salvano, M.A.; Dalcero, A.M.; Rosa, C.A. Evaluation of Saccharomyces cerevisiae as an Antiaflatoxicogenic agent in broiler feedstuffs. Poult. Sci. 2013, 92, 1655–1663. [Google Scholar] [CrossRef] [PubMed]
- Bejaoui, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Ochratoxin A removal in synthetic and natural grape juices by selected oenological Saccharomyces strains. J. Appl. Microbiol. 2004, 97, 1038–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petruzzi, L.; Bevilacqua, A.; Baiano, A.; Beneduce, L.; Corbo, M.R.; Sinigaglia, M. Study of Saccharomyces cerevisiae W13 as a functional starter for the removal of ochratoxin A. Food Control 2014, 35, 373–377. [Google Scholar] [CrossRef]
- El-Nezami, H.; Polychronaki, N.; Salminen, S.; Mykkänen, H. Binding Rather than Metabolism May Explain the Interaction of Two Food-Grade Lactobacillus Strains with Zearalenone and Its Derivative ά-Zearalenol. Appl. Environ. Microbiol. 2002, 68, 3545–3549. [Google Scholar] [CrossRef] [Green Version]
- Armando, M.R.; Pizzolitto, R.P.; Dogi, C.A.; Cristofolini, A.; Merkis, C.; Poloni, V.; Dalcero, A.M.; Cavaglieri, L.R. Adsorption of ochratoxin A and zearalenone by potential probiotic Saccharomyces cerevisiae strains and its relation with cell wall thickness. J. Appl. Microbiol. 2012, 113, 256–264. [Google Scholar] [CrossRef]
- El-Sharkawy, S.H.; Selim, M.I.; Afifi, M.S.; Halaweish, F.T. Microbial transformation of zearalenone to a zearalenone sulfate. Appl. Environ. Microbiol. 1991, 57, 549–552. [Google Scholar] [CrossRef] [Green Version]
- Shetty, P.H.; Hald, B.; Jespersen, L. Surface binding of aflatoxin B1 by Saccharomyces cerevisiae strains with potential decontaminating abilities in indigenous fermented foods. Int. J. Food Microbiol. 2007, 113, 41–46. [Google Scholar] [CrossRef]
- Rahaie, S.; Emam-Djomeh, Z.; Razavi, S.H.; Mazaheri, M. Immobilized Saccharomyces cerevisiae as a potential aflatoxin decontaminating agent in pistachio nuts. Braz. J. Microbiol. 2010, 41, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Huwig, A.; Freimund, S.; Käppeli, O.; Dutler, H. Mycotoxin detoxication of animal feed by different adsorbents. Toxicol. Lett. 2001, 122, 179–188. [Google Scholar] [CrossRef]
- Yiannikouris, A.; André, G.; Poughon, L.; François, J.; Dussap, C.G.; Jeminet, G.; Bertin, G.; Jouany, J.P. Chemical and conformational study of the interactions involved in mycotoxin complexation with β-D-Glucans. Biomacromolecules 2006, 7, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, M.; Nowak, A.; Czyzowska, A. Removal of ochratoxin A by wine Saccharomyces cerevisiae strains. Eur. Food Res. Technol. 2013, 236, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Quintela, S.; Villarán, M.C.; López de Armentia, I.; Elejalde, E. Ochratoxin A removal in wine: A review. Food Control 2013, 30, 439–445. [Google Scholar] [CrossRef]
- Joannis-Cassan, C.; Tozlovanu, M.; Hadjeba-Medjdoub, K.; Ballet, N.; Pfohl-Leszkowicz, A. Binding of zearalenone, aflatoxin B 1, and ochratoxin a by yeast-based products: A method for quantification of adsorption performance. J. Food Prot. 2011, 74, 1175–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González Pereyra, M.L.; Dogi, C.; Torres Lisa, A.; Wittouck, P.; Ortíz, M.; Escobar, F.; Bagnis, G.; Yaciuk, R.; Poloni, L.; Torres, A.; et al. Genotoxicity and cytotoxicity evaluation of probiotic Saccharomyces cerevisiae RC016: A 60-day subchronic oral toxicity study in rats. J. Appl. Microbiol. 2014, 117, 824–833. [Google Scholar] [CrossRef]
- Yiannikouris, A.; Franç Ois, J.; Poughon, L.; Dussap, C.-G.; Bertin, G.; Jeminet, G.; Jouany, J.-P. Adsorption of zearalenone by B-D-Glucans in the Saccharomyces cerevisiae cell wall. J. Food Prot. 2004, 67, 1195–1200. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Altschup, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Abarca, M.L.; Accensi, F.; Cano, J.; Cabañes, F.J. Taxonomy and significance of Black Aspergilli. Antonie Van Leeuwenhoek 2004, 86, 33–49. [Google Scholar] [CrossRef]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]



{kind=link}
{kind=link}
| Saccharomyces cerevisiae Strains | Concentration of OTA (µg/L) (±SD) | |
|---|---|---|
| Viable Cells | Heat-Treated Cells | |
| Control | 9.81 ± 0.12 a | 9.81 ± 0.12 a |
| Levulin FB | 7.11 ± 0.06 b | 3.63 ± 0.21 e |
| SC5 | 6.38 ± 0.23 c | 5.41 ± 0.19 d |
| Saccharomyces cerevisiae Strains | Concentration of OTA (µg/L) (±SD) | |
|---|---|---|
| Viable Cells | Heat-Treated Cells | |
| Control | 6.64 ± 0.19 a | 6.64 ± 0.19 a |
| Levulin FB | 5.21 ± 0.11 b | 3.36 ± 0.09 d |
| SC5 | 4.46 ± 0.02 b | 4.02 ± 0.03 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dammak, I.; Alsaiari, N.S.; Fhoula, I.; Amari, A.; Hamdi, Z.; Hassouna, M.; Ben Rebah, F.; Mechichi, T.; Lasram, S. Comparative Evaluation of the Capacity of Commercial and Autochthonous Saccharomyces cerevisiae Strains to Remove Ochratoxin A from Natural and Synthetic Grape Juices. Toxins 2022, 14, 465. https://doi.org/10.3390/toxins14070465
Dammak I, Alsaiari NS, Fhoula I, Amari A, Hamdi Z, Hassouna M, Ben Rebah F, Mechichi T, Lasram S. Comparative Evaluation of the Capacity of Commercial and Autochthonous Saccharomyces cerevisiae Strains to Remove Ochratoxin A from Natural and Synthetic Grape Juices. Toxins. 2022; 14(7):465. https://doi.org/10.3390/toxins14070465
Chicago/Turabian StyleDammak, Islem, Norah Salem Alsaiari, Imene Fhoula, Abdelfattah Amari, Zohra Hamdi, Mnasser Hassouna, Faouzi Ben Rebah, Tahar Mechichi, and Salma Lasram. 2022. "Comparative Evaluation of the Capacity of Commercial and Autochthonous Saccharomyces cerevisiae Strains to Remove Ochratoxin A from Natural and Synthetic Grape Juices" Toxins 14, no. 7: 465. https://doi.org/10.3390/toxins14070465
APA StyleDammak, I., Alsaiari, N. S., Fhoula, I., Amari, A., Hamdi, Z., Hassouna, M., Ben Rebah, F., Mechichi, T., & Lasram, S. (2022). Comparative Evaluation of the Capacity of Commercial and Autochthonous Saccharomyces cerevisiae Strains to Remove Ochratoxin A from Natural and Synthetic Grape Juices. Toxins, 14(7), 465. https://doi.org/10.3390/toxins14070465