Microcystin Contamination in Irrigation Water and Health Risk
by
 , , , and
, , , and
Mohammed Haida
1 ,
,
Fatima El Khalloufi
2,
Richard Mugani
1 ,
,
Yasser Essadki
1,
Alexandre Campos
3,
Vitor Vasconcelos
3,4,* and
Brahim Oudra
1 1
Water, Biodiversity and Climate Change Laboratory, Faculty of Sciences Semlalia, Cadi Ayyad University, Marrakesh 40000, Morocco
2
Natural Resources Engineering and Environmental Impacts Team, Multidisciplinary Research and Innovation Laboratory, Polydisciplinary Faculty of Khouribga, Sultan Moulay Slimane University of Beni Mellal, B.P, 45, Khouribga 25000, Morocco
3
Interdisciplinary Centre of Marine and Environmental Research (CIIMAR), Terminal de Cruzeiros do Porto de Leixões, Av. General Norton de Matos, s/n, 4450-208 Porto, Portugal
4
Department of Biology, Faculty of Sciences, University of Porto, Rua do Campo Alegre, 4169-007 Porto, Portugal
*
Author to whom correspondence should be addressed.
Toxins 2024, 16(4), 196; https://doi.org/10.3390/toxins16040196
Submission received: 4 February 2024
/
Revised: 8 April 2024
/
Accepted: 10 April 2024
/
Published: 19 April 2024
Abstract
:Microcystins (MCs), natural hepatotoxic compounds produced by cyanobacteria, pose significant risks to water quality, ecosystem stability, and the well-being of animals, plants, and humans when present in elevated concentrations. The escalating contamination of irrigation water with MCs presents a growing threat to terrestrial plants. The customary practice of irrigating crops from local water sources, including lakes and ponds hosting cyanobacterial blooms, serves as a primary conduit for transferring these toxins. Due to their high chemical stability and low molecular weight, MCs have the potential to accumulate in various parts of plants, thereby increasing health hazards for consumers of agricultural products, which serve as the foundation of the Earth’s food chain. MCs can bioaccumulate, migrate, potentially biodegrade, and pose health hazards to humans within terrestrial food systems. This study highlights that MCs from irrigation water reservoirs can bioaccumulate and come into contact with plants, transferring into the food chain. Additionally, it investigates the natural mechanisms that organisms employ for conjugation and the microbial processes involved in MC degradation. To gain a comprehensive understanding of the role of MCs in the terrestrial food chain and to elucidate the specific health risks associated with consuming crops irrigated with water contaminated with these toxins, further research is necessary.
Keywords:
microcystins; irrigation water; bioaccumulation; transfer; conjugation; health risk; phytotoxicity; biodegradationKey Contribution: MCs present in surface waters used for irrigation can migrate to plants and accumulate in their edible parts, and then be transmitted to animals and humans through consumption, posing a health risk. Living organisms have developed detoxification methods based on glutathione. Additionally, the degradation of MCs by microorganisms could be an effective and environmentally friendly strategy to reduce MCs.
1. Introduction
Cyanobacteria proliferation has evolved as a direct result of climate change. This concerning trend is characterized by rising global temperatures and an increase in nutrient influx, mainly nitrogen and phosphorus, from sources such as agricultural runoff and wastewater treatment plants [1]. These cyanobacteria strains are potential toxins producers known as cyanotoxins. MCs are considered the most prevalent cyanotoxins in water bodies and have reached levels that exceed the limits set by the WHO (World Health Organization), posing a severe threat to worldwide public health [2,3].
In many underdeveloped and developing countries, water contaminated MCs is collected in lakes or artificial ponds and then used to irrigate all types of crops, including hydroponic crops, which are widespread due to the increase in human population and demand for agri-food products [4,5]. It should be noted that the presence of MCs in irrigation water is likely to induce a range of phytotoxic effects, including reduced seed germination, altered plant growth and development, reduced photosynthetic performance, and disruption of hormonal balance. These toxins also cause alterations in plant metabolism, such as increased lipid peroxidation, reduced protein content, and oxidative stress. As a result, yield losses attributable to MC exposure can range from 50% to 82%, depending on the specific crop and duration of exposure [6,7,8,9].
Irrigation water contaminated with MCs infiltrates and penetrates the root barriers of agricultural plants, and bioaccumulates in roots, stems, leaves, and edible crops, eventually entering the human body via consumption [10,11,12], with daily intakes above the recommended value of 0.04 μg of MC/kg of body weight/day [13].
The transfer of MCs through aquatic food chains has been the subject of several studies on a large number of organisms [14]. However, few studies have been conducted on MC transmission through terrestrial food chains. Little information is available about cases of MC transfer through the ingestion of contaminated food, involving bats, spiders, birds, and rats [15,16].
The bioaccumulation and transfer of MCs in the terrestrial food chain trigger adverse effects on animal and human health [17]. After entering the bloodstream, MCs cause severe damage to organs, such as the liver, kidneys, intestines, heart, brain, lungs, and gonads. MCs selectively attack these organs, causing long-term damage [18,19,20]. Although no case of human death has been recorded, exposure to MCs has been associated with the death of some animals [21,22,23,24,25].
Furthermore, MCs exhibit significant resistance to enzymatic hydrolysis [26]. Nevertheless, plant cells contaminated with MCs can undergo a natural detoxification process. In this process, MCs are converted into more hydrophilic compounds, which are then conjugated by enzymes such as glutathione S-transferase (GST) and glutathione (GSH). These compounds are then compartmentalized [27,28,29]. Studies have demonstrated that converted conjugates have substantially less toxicity than untransformed conjugates, even though the detoxification process does not completely remove MCs from plants [30].
Therefore, it is believed that the biological process of microorganism biodegradation in irrigation water offers an economical and effective means of purifying MC-contaminated water bodies [31]. First attempts have been made to isolate a strain of Sphingomonas from surface waters capable of using MC-LR as the only source of carbon and nitrogen for its growth. The results of these experiments showed that the degradation of 1 mg L−1 of MC–LR generally began 2 to 8 days after its addition to surface water samples [32]. The main objectives of this work were to determine the fate of MCs in irrigation water, investigate the phytotoxic effects of MCs on plants in hydroponic cultures, assess the bioaccumulation and transfer of MCs in a terrestrial food chain, determine the role of detoxification enzymes and bacterial strains capable of degrading MCs in the elimination of this toxin, and assess the risks associated with exposure to MCs for human and animal health. To achieve the objectives of this review, reference documents were collected using online databases such as Scopus, ScienceDirect, PubMed, and Web of Science, and journals have been carefully read and analyzed. Reference documents were collected using main keywords such as microcystins in irrigation water, bioaccumulation of microcystins in plants, transfer of microcystins in the food chain, conjugation of microcystins, health risk of microcystines, microcystine phytotoxicity, and microcystine biodegradation. The years of publication for reference elements were mainly between 1981 and 2023. More than 500 references were retrieved, and 147 articles were used as references in the manuscript.
2. Microcystins
The potential for cyanobacteria to produce harmful toxins to humans and animals makes their worldwide abundance a serious cause for concern [3]. Many cyanobacteria species produce secondary metabolites such as neurotoxins, dermatoxins, and hepatotoxins [33]. MCs have a cyclic heptapeptide structure: cyclo-(D-Ala1-X2-D-isoMeAsp3-Y4-Adda5-D-isoGlu6-Mdha7), where X and Y stand for the very L-amino acid pair [34] (Figure 1). Among these analogs, MC-LR (L: Leucine, R: Arginine) is the most hazardous and prevalent one, garnering significant interest in studies and reports [13].
The detrimental effects of MCs are believed to be caused by the Adda residue (3-amino-9-methoxy-2,6,8-trimethyl-10-phenyldeca-4,6-dienoic acid) [35]. Cyanobacteria capable of making MCs include mainly Microcystis, Anabaena, Nostoc, Planktothrix, and Oscillatoria [36,37,38,39]. The enormous impact of MCs is due to their capacity to cause acute hepatotoxicosis in both humans and animals, making them very harmful cyanotoxins with severe health and ecological concerns [40].
Figure 1.
Structure of the hepatotoxic cyclic peptides, MCs. The numbers 1–7 represent seven amino acid residues. X and Z in positions two and four are highly variable L-amino acids [41].
Figure 1.
Structure of the hepatotoxic cyclic peptides, MCs. The numbers 1–7 represent seven amino acid residues. X and Z in positions two and four are highly variable L-amino acids [41].

3. Emergence and Persistence of MCs in Irrigation Water
The synthesis and release of cyanotoxins are influenced by several factors. Physicochemical factors (phosphorus, nitrogen, iron, pH, temperature, and light) promote the growth of cyanobacteria and consequently the excessive release of MCs. Biological factors, where the production of MCs increases in the presence of competitors and predators, as well as genetic factors, because variation in the amount of MCs synthesized results from changes in the transcription of genes that encode this toxin, also have an impact [42,43]. Some investigations reported the presence of cyanobacteria and their MCs in irrigation water sources such as groundwater, natural reservoirs, lakes, and rivers [43,44,45,46,47].
Several studies found harmful cyanobacteria and important MC concentrations in irrigation water (Table 1). Quantities of up to 2100 µg/L of extracellular MCs and 3240 µg/g of intracellular MCs can be produced by these species (Table 1). These levels are high in irrigation water and can constitute a health risk, because they exceed the threshold set by WHO recommendations, namely, 1 µg/L of MCs in the lifetime of water and 12 µg/L in the short term of water in drinking water, and 24 µg/L in recreational water, in the absence of regulations on irrigation water quality [48]. Furthermore, several studies showed that both cyanobacteria and their MCs pose a severe problem in irrigation water because of the substantial risk to human health related to their presence in these waters; these water sources may be unsuitable for use as plant irrigation [49]. No study has specifically collected data on the occurrence of MCs in terms of concentrations, variants, and species in irrigation water.
4. Phytotoxicity of MC on Crops in Hydroponic Systems
MCs are reported to have several potent impacts on plant growth, physiology, and the yield and quality of fruits (Table 2). Studies have shown that the exposure of hydroponically grown plants to MCs can produce a significant inhibition of root growth of Oryza sativa and Fragaria vulgaris, and a reduction in plant weight, length, and surface area. We can say that in this type of culture, the roots are in direct contact with the toxin in the absence of soil, which constitutes a barrier against MCs [63,64]. Medicago sativa germination, total fresh weight, root, and shoot length were all reduced after 14-day exposure to 5–1000 µg/L of MCs [65]. According to [66], the leaf weight of Spinacia oleracea decreases considerably after 21 days of MC exposure (50 and 100 µg/L). Numerous investigations have demonstrated that Oryza sativa roots, stems, and leaves exhibit increased biomass, surface area, and dry weight when exposed to low concentrations of MCs [67,68,69]. Exposure to increased concentrations of MC for 7 days had negative effects on the growth of rice plants, reflected in a decrease in the dry weight of roots, stems, and leaves, in addition to the inhibition of photosynthesis [70]. The proportion of fruit set, the number of grains per panicle, and grain weight per panicle all decreased [67]. On the other hand, the early exposure of rice seedlings to MCs (100–1000 µg/L for 7 days) reduced the number of filled grains per panicle, the germination rate, panicle weight, and the contents of soluble proteins, sugars, and starch in the grains [67] (Table 2).
Concerning oxidative stress, exposure to MCs upstream increased the activity of oxidative enzymes such as superoxide dismutase (SOD), catalase (CAT), peroxidase (POD), polyphenoloxydase (PPO), and ascorbate peroxidase (APX) in Fragaria vulgaris plants [62].
Several studies have determined that exposure to M. aeruginosa extracts containing MCs increases the content of oxidative stress bioindicators such as H2O2, O2, and MDA in Oryza sativa L. seedlings [70]. Contact of MCs and plants in hydroponic systems enhances the bioavailability and transport of MCs to various plant organs and induces yield loss. The existence of MCs in hydroponic systems is a fundamental problem that must be carefully addressed to reduce any potential negative effects on plants’ health and yield, since hydroponic systems are gaining more attention, as an alternative to conventional agricultural systems.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 2.
The impact of MCs on the morphological and physiological parameters of various agricultural species in hydroponic systems.
Table 2.
The impact of MCs on the morphological and physiological parameters of various agricultural species in hydroponic systems.
| Plant Species | Toxin | Growth Stage | Exposure Time (Days) | Applied Concentration (μg/L) | Phytotoxicity on Plant | References |
|---|---|---|---|---|---|---|
| Spinacia oleracea | MC-LR | Plants | 21 | 50 | (-) leaf FW | [66] |
| Cucumis sativus | MCt | Seedlings | 7 | 5 | (+) H2O2, O2-, MDA | [71] |
| Seedlings/flowering /fruiting | 7 | 1–1000 | (-) stem diameter, plant height, leaf area, root DW, leaf number, shoot DW, yield | [72] | ||
| Lactuca sativa | MC-LR (P) | Plants | 10 | 1–100 | (-) leaf biomass, root biomass | [73] |
| Seedlings | 14 | 5–1000 | (-) root length, total FW, shoot length | [74] | ||
| Vicia faba | MCt | Seedlings | 48 | 10–100 | (-) shoot DW, root DW, nodule number, and DW | [75] |
| Oryza sativa | MCt | Seedlings | 7 | 5–500 | (-) plant height, shoot DW, root length, root DW; (+) membrane permeability | [76] |
| Oryza sativa | MCt | Seedlings | 7 | 1-3000 | (+) root biomass, (-) stem biomass, leaf biomass, grains per panicle, grain weight per panicle, root biomass, setting percentage | [68] |
| Oryza sativa | MCt | Seedlings | 7 | 1–100 | (+) root surface area, shoot height; (-) shoot height, root surface area | [67] |
| Oryza sativa | MCt | Seedlings | 7 | 1–3000 | (-) stem dry weight, leaf DW, net photosynthetic rate, root DW | [77] |
| Oryza sativa | MCt | Seedlings | 7 | 5–10 | (+) H2O2, O2-, MDA; (-) RGR | [71] |
| Oryza sativa | MCt | Seedlings | 7 | 10–1000 | (-) root surface area, plant height, filled grains per panicle, panicle weight, seed setting rate, soluble protein, sugar, and starch in the grain | [78] |
| Oryza sativa | MCt | Booting | 7 | 10–1000 | ||
| Oryza sativa | MCt | Filling | 7 | 10–1000 | ||
| Oryza sativa | MCt | Seedlings | 21 | 10 | (+) Phosphorus content in shoots and roots, root and shoot DW; (-) root DW | [69] |
| Oryza sativa | MCt | Seedlings | 30 | 5–500 | (-) root length, root surface area, root dry weight, surface area and volume, root volume, lateral root number, crown root number | [71] |
(-): increase; (+): decrease; P: pure MC; DW: dry weight; FW: fresh weight; MCt: total MCs; MDA: malondialdehyde; RGR: relative growth rate; H2O2: hydrogen peroxide; O2-: superoxide anion.
5. Bioaccumulation of MCs in Tissues of Plants in Hydroponic Systems
To date, information on the bioaccumulation of MCs by plants in hydroponic crops irrigated with water contaminated with cyanobacterial cells and toxins remains limited. This raises concerns about the mechanisms of bioaccumulation and the health consequences associated with this phenomenon. Nevertheless, studies have shown that MCs can accumulate in terrestrial plants, especially food crops, making them vulnerable to toxin bioaccumulation [79,80]. In hydroponic crops, found that Lactuca sativa exposed to concentrations of 5 to 2000 µg/L of MC-LR for 14 days absorbed the toxin at a high rate (700–1400 µg MC-LR/kg FW) [74], surpassing the WHO recommended daily dose of 0.04 µg kg−1 body weight [81] (Table 3). Similarly, irrigation of the Oryza sativa plant in three hydroponic cultures with water containing different concentrations of MCs showed bioaccumulation of the order of 30 µg/kg in leaves [78], 112,000 µg/kg in leaves [65], and 275 µg/kg in roots [77] (Table 3). Similarly, [72] showed that MCs bioaccumulated in Cucumis sativus L. fruits following exposure to concentrations ranging from 1 to 1000 μg/L, and in Oryza sativa roots, stem, and leaves following exposure to concentrations ranging from 1 to 3000 μg/L. Indeed, during the process of bioaccumulation, the levels of MCs can potentially decrease to the point of falling below the detection threshold. This phenomenon could be related to the gradual chemical changes that MCs undergo, either as a result of their binding to intrinsic biomolecules or because of detoxification by conjugation with GSH, catalyzed by GST [82]. Alternatively, the diminished detectability of MCs could be attributed to their increased dilution, owing to factors like larger fruit size, greater fruit numbers on plants, and water content during fruit growth and ripening [83]. Plants tend to be affected by several parameters that influence the accumulation of MCs, such as exposure duration (which is most noticeable when it is less than 15 days), concentration, the studied plants, and the particular organ (roots are frequently showing higher absorption rates) (Table 3).
6. Transfer and Fate of MCs in the Terrestrial Food Chain
A crucial route for MC transfer to plants in hydroponic systems is the use of cyanobacterial cell- and MC-contaminated water sources [80]. Under natural circumstances, MCs retain their chemical stability due to their cyclic heptapeptide structure. They are resistant to chemical destruction in extremely acidic and alkaline settings [84], elevated temperatures [85], and photolysis when exposed to natural sunshine [86]. These properties increase MCs’ persistence and availability to plants in irrigation water [87,88]. Moreover, the decomposition of cyanobacterial cells and MCs, introduced into the hydroponic systems, is delayed, leading to the prolonged release of MCs within the substrate and consequently a long-term persistence [89]. In this crop system, roots come into direct contact with MCs, where their degradation is relatively low, enhancing the transfer rate [90].
Several studies have been conducted in recent decades on the interactions of MCs with a variety of aquatic vertebrate and invertebrate organisms [91,92,93,94,95]. MC bioaccumulation was investigated in insect chironomid larvae that meet contaminated water surfaces [96]. This study suggests that invertebrates may be essential to the MC translocation to environments with higher trophic levels. A large amount of MCs, however, is delivered to the spleens of ducks, moles, turtles, aquatic birds, and newly emerging aquatic insects. These species may act as carriers, allowing MCs to move from water to terrestrial food chains [97,98,99]. Likewise, exposure through the consumption of imagos and subimagos of Hexagenia limbata showed a transfer of MCs to brown bats (Myotis lucifugus) [16]. In addition, significant concentrations of MCs were detected in spiders and birds consuming emergent aquatic insects in riparian areas [15]. As a result, land animals may consume meals based on plants irrigated with these MCs or come into direct contact with MC-contaminated water. It has been suggested that MC transmission via ingestion is one of the most serious threats to the terrestrial food chain. There have been few studies on MC transmission from aquatic ecosystems to terrestrial food chains. This scarcity highlights the need for additional research to properly understand the related potential health and ecological implications of MC exposure in terrestrial ecosystems.
7. Effects of MCs on Domestic/Wild/Aquatic Animals and Human Health Risks
The presence of toxic cyanobacterial blooms endangers the survival and well-being of several aquatic organisms exposed to MCs, including fish, turtles, ducks, and other species [93]. Studies have demonstrated that MCs can cause a variety of abnormalities and health problems in various fish species, such as coiled bodies and tails, enlarged livers, hepatic hemorrhage, reduced yolk absorption, vacuolar dystrophy, cardiac problems, and disturbances to both organ functions and developmental processes [100,101], in addition to an increase in glutathione –S- transferase (GST) activity in freshwater snails [102,103]. At the cell level, MCs disrupt protein phosphatase activities. The MeAsp residues first prevent PP1 and PP2A from functioning. The cysteine and serine residues of PP1 and PP2A are then covalently bound by the Mdha group of MCs, which permanently inhibits these phosphatases and causes liver damage [104]. MCs have exhibited a noteworthy effect on animals, and damaged the hypothalamic–pituitary system in men by altering mRNA expression and blood hormone levels [105]. Furthermore, studies have shown that MCs can promote tumors, cause acute lung inflammation in rats, alter the structure and function of the testes, induce normocytic anemia, disturb the bone marrow and the immune system, and cause cardiotoxicity in animals [106,107,108,109]. In addition, multiple investigations have shown deadly MC poisonings in domestic and wildland animals [110,111], such as cattle [112], cows [20], sheep [113], heifers [21], dogs [23,114], white rhinos [115], and wild deer [20], zebras, blue wildebeests, and impalas [24].
Microcystin-contaminated irrigation water is a common route of MC transfer to agricultural land. As a result, MCs can accumulate within food crops and induce negative effects on human health [16]. After entering the bloodstream, MCs can cause serious organ damage via several biological processes. MCs selectively disturb organs like the liver, kidney, gut, heart, and gonads [19]. Furthermore, MCs might cause an imbalance in cellular oxidation and increase MDA levels in cells, leading to lipid and peroxidation and compromising cell defense ability. This process causes abnormal cell apoptosis, disruption, mitochondrial dysfunction and cytoskeleton endoplasmic reticulum, chronic poisoning, DNA damage, and the emergence of symptoms like nausea, dizziness, vomiting, and other side effects [116,117]. MCs can migrate to vital biological organs such as the heart, brain, and lungs, causing long-term damage in humans [17,18]. Furthermore, MCs have been proven to pass the blood–brain barrier, resulting in unforeseen long-term side effects in humans [118,119].
MC exposure causes glycogen loss, liver cancer, and hepatocyte necrosis [120]. Extensive liver injury can impair bile pigment production, eventually leading to internal bleeding or death from hemorrhage. Hemorrhagic shock has the potential to be fatal to the liver [121]. Similarly, MC exposure alters microRNA expression in liver cells, resulting in severe liver damage and an increase in the formation of liver tumors [122]. Chronic exposure has deleterious consequences, as evidenced by a decrease in human kidney cell viability and the induction of genes involved in apoptosis [123,124]. Cyanobacterial blooms and their associated toxins are widespread in irrigation water sources around the world. Several studies showed their ability to affect biological processes by disturbing organisms’ normal physiology and biochemistry. Despite this, research on the toxicity of MCs and the severe health threats they bring to humans and terrestrial species is still minimal. As a result, more research and tests are needed to fill these gaps and completely assess the MC’s associated impacts on the trophic chain.
8. Conceptual Diagram of MC Fate in a Terrestrial Food Chain
Eutrophication is a process related to an excessive enrichment of waterbodies with minerals and nutrients, often because of land runoff, leading to an excessive growth of toxic microalgae. This phenomenon can lead to the release of MCs produced by different cyanobacteria species. The use of contaminated water for irrigation purposes leads to possible MC bioaccumulation in various organisms of the terrestrial food web, notably plants (direct contact with MCs) and other terrestrial animals and humans (indirect transfer of MCs). Research has shown that MCs can accumulate in various organs of terrestrial organisms, leading to deleterious impacts on animal and human health. The bioaccumulation of MCs in the food web has been rarely studied in various locations, highlighting the potential risks associated with MC transfer in the food chain [125,126,127,128] (Figure 2).
9. Depuration through the Conjugation of MCs
The ADDA group on MC-LR is important for the binding of the toxin to its target enzyme, protein phosphatases 1 and 2A [75]. Moreover, the Mdha group in MCs can subsequently covalently bind to cysteine in a protein phosphatase enzyme [19]. MCs permanently block the active site and destroy the functionality of the protein phosphatase enzyme. In response to exposure to MCs and their negative effects, several organisms have developed detoxification processes for MCs. However, several enzymes are involved in the process of eliminating toxic products [129,130].
Studies on the detoxification mechanism of MCs in plant tissues are limited. After the absorption of MCs by the plant, a certain quantity will be conjugated by nonenzymatic glutathione (GSH). A second quantity will be enzymatically conjugated into GSH via the glutathione S-transferase system. Part of the remaining MC-LR binds to phosphatases and possibly other cellular proteins. Finally, the remaining MC-LR could be taken by chloroplasts, in which three main pathways have been postulated, a non-enzymatic pathway by binding to the GSH, enzymatic conjugation to GSH via the GST system, and reactions with proteins or structures of the photosynthetic apparatus of the chloroplast. To remove conjugates from glutathione, plants transfer these conjugates into the vacuole through multi-drug resistance proteins (MRPs), which are part of the ABC family of conveyors, for the temporary storage and subsequent processing of GSH conjugates [75].
In animals, when the toxin arrives in the cell, oxygen-containing groups are attached to the toxins, and phase I enzymes, notably cytochrome P-450, initiate oxidative reactions. Phase II enzymes, such as glutathione-S-transferase (GST), intervene to modify the toxin and form glutathione conjugates. Glutathione (GSH) is commonly used in phase II biotransformations. The formation of the glutathione conjugate by the phase II enzyme, glutathione-S-transferase (GST), is one of the most common types of toxin modification [129]. This reaction occurs between a nucleophilic center of the toxin (Mdha group) and the sulfhydryl group of reduced glutathione (GSH) [129]. When a toxin conjugates with GSH, the enzyme γ-glutamyl transferase cleaves the γ-glutamic acid group of the GSH molecule to produce the intermediate γ-glutamylcysteine [129,131]. The glycine of this γ-glutamylcysteine intermediate is subsequently cleaved by a dipeptidase to create a cysteine-conjugated product, which is ultimately oxidized to yield the mercapturic acid metabolite. While other metabolic pathway conjugates may be excreted from cells, this mercapturic acid derivative is easily eliminated in the urine [129] (Figure 3).
10. Approaching the Bacterial Enzymatic Biodegradation of MCs
The quality of irrigation water must be assessed to prevent short- or long-term potential negative effects of MCs. To protect household, agricultural, and recreational applications, water sources and supply need to be monitored for the possible presence of hazardous cyanobacteria and MCs. When treating toxin-contaminated water, the physical and chemical properties of the toxin, its nature, and the development and proliferation patterns of cyanobacteria must all be taken into account [133]. There have also been attempts to remove MCs from irrigation-grade water using treatment techniques such as ozonation and chlorination [134]. Several studies stated that standard water treatment techniques can be costly to maintain, ineffective in getting rid of or degrading MCs, and potentially hazardous by their byproducts. Therefore, it is imperative to investigate a treatment strategy that is economical and efficient while also not endangering the environment or producing unfavorable byproducts after treatment. Several studies focused on using bacteria as a biological treatment approach to break down and eliminate MCs from water. MC breakdown has been seen in a variety of different taxa, including Sphingomonas, Arthrobacter, Acinetobacter, Novosphingobium, Bacillus, Paucibacter, Pseudomonas, Stenotrophomonas, and Sphingopyxis [11,135,136]. The rate of degradation varies amongst different strains; Bacillus sp. AMRI-03, for instance, took five days to fully break down MC-RR [137]. Within 72 h, Arthrobacter spp. F10, F7, C6, R1, R4, R6, and R9 were able to eradicate MC-LR [138]. In 24 days, MC-LR was eradicated by Pseudomonas aeruginosa DMXS and Novosphingobium sp. KKU-25s [139]. Furthermore, MC-LR, MC-LF, and MC-RR were eliminated by Stenotrophomonas maltophilia 4B4 in 10, 12, and 14 days, respectively [140]. The Sphingopyxis bacterium YF1 is the fastest; it was reported to completely degrade MC-LR within 120 min [141]. Based on the existing evidence, bacterial strains globally widespread in several habitats possess a capacity for chemical compound breakdown and may play a major role in natural processes [142]. Furthermore, it was shown that a mixture of different bacterial strains may break down MC-LR. For example, the complete degradation of MC-LR was demonstrated by a combination of ten isolates (Aeromonas sp., Acinetobacter sp., Novosphingobium sp., Pseudomonas sp., Ochrobactrum sp., Sphingomonas sp., Rhodococcus sp., Sphingopyxis sp., Stenotrophomonas sp., and Steroidobacter sp.), and another combination of seven isolates (Hyphomicrobium aestuarii, Acinetobacter sp., Rhizobium sp., Pseudoxanthomonas sp., Sphingomonas sp., Sphingobium sp., and Steroidobacter sp.) [142]. A naturally occurring group of bacteria collected from the mucilage of M. aeruginosa colonies during their flowering stage, comprising Agrobacterium sp., Brevundimonas sp., Bosea sp., Rasbo sp., Hyphomicrobium sp., Rhizobium sp., Roseomonas sp., Rhodococcus sp., Mesorhizobium sp., Nitrosococcus sp., Sphingomonas sp., and Sandaracinobacter sp., effectively decomposed MC-LR [142,143].
Several studies focusing on genotypic analysis have identified specific gene clusters, including mlrA, mlrB, mlrC, and mlrD, in certain bacterial groups. These gene clusters encode enzymes such as MlrA, MlrB, MlrC, and MlrD, which are involved in the degradation of MCs [141,142].
The degradation process begins with the cleavage of the Adda-Arg peptide bond of MC-LR by the enzyme MlrA, which is encoded by the mlrA gene and produces linearized MC-LR. The linearized MC-LR is subsequently converted into a tetrapeptide by the enzyme MlrB, which is generated by the mlrB gene. This process involves hydrolyzing the Ala-Leu link. Furthermore, the tetrapeptide is broken down into smaller peptides and amino acids by the enzyme MlrC, which is expressed by the mlrC gene. It is believed that the mlrD gene facilitates the transmission of MCs and their breakdown products [144].
As a result, breakdown routes may differ between MC genotypes and bacterial strains [144]. In addition to well-known MC-LR degradation products (tetrapeptide, adda, and linearized MC-LR), studies have identified eight other degradation intermediaries: three tripeptides (ADA-Glu-MDHA, GLU-MDHA-ALA, and LEU-MEASP-ARG), three dipeptides (GLU-MDHA, MDHA-ALA, and MESP-ARG), and two amino acids (Leu and ARG) for the MC-LR degradation pathway use of Sphingopyxis sp. [141,145]. These biodegradation pathways are important for developing strategies to degrade MCs in contaminated environments (Figure 4).
11. Conclusions and Research Requirements
In recent years, research into cyanobacteria and their toxins has developed very rapidly around the world. However, it is clear from the data synthesized and presented above that the distribution of MCs in irrigation water, their transfer and bioaccumulation in hydroponically grown plants and animals, their phytotoxicity and effects on animals and humans, and the ways in which MCs are eliminated are still poorly understood. This research aims to synthesize existing knowledge on the fate and effects of MCs that contaminate the irrigation water of hydroponic crops and to propose methods for eliminating these toxic substances in order to mitigate the associated risks. Many studies have shown that MCs released into irrigation water can migrate to plant roots, then to leaves, to accumulate in edible parts like fruit. This bioaccumulation of MCs can negatively affect plant morphological and physiological parameters and fruit yield and quality. The assessment of the phytotoxicity and bioaccumulation of MCs in agricultural plants, particularly in hydroponic crops, highlighting the potential risks to plants associated with the presence of MCs in irrigation water, requires further and more detailed research. In addition, MCs can be transferred and bioaccumulated in various animal and human tissues, leading to adverse effects in animals and humans. This shows that MCs can enter the terrestrial food chain through the ingestion of edible plant parts or drinking water. However, the transfer of MCs to terrestrial animals through the direct consumption of edible agricultural products, particularly those grown hydroponically, has not been considered a major concern, despite its potential impact on animal health and the food chain in general. To this end, the efforts of researchers concerned with MCs are focusing on approaches to eliminating MCs, either by eliminating cyanobacteria and MCs from water or by investigating MCs’ detoxification mechanisms in animal and plant cells affected by MCs.
This comprehensive review addresses many critical aspects of MCs, including their distribution in irrigation water, their transfer and bioaccumulation in hydroponic food plants, their transfer to terrestrial animals, and the impact they have on flora and fauna, including humans; this research highlights the ubiquitous nature of these toxins in terrestrial ecosystems. Understanding these multifaceted interactions provides a holistic perspective essential to the development of effective strategies to mitigate the negative consequences of MCs on hydroponic agricultural products, animal welfare, and human well-being. Such an integrated approach is essential to promote informed decision-making and develop sustainable solutions to preserve the quality of irrigation water and hydroponic crops, and to protect ecosystems and human health from the threats posed by MCs.
Author Contributions
Conceptualization, M.H., F.E.K., B.O., R.M. and Y.E.; data acquisition and formal analysis, M.H.; drafting—preparation of original version, M.H.; drafting—revision and editing, M.H., R.M., Y.E., F.E.K., V.V. and B.O.; supervision, B.O. and F.E.K.; project administration A.C.; acquisition of funds, A.C., V.V. and B.O. All authors have read and agreed to the published version of the manuscript.
Funding
This project received funding from the European Union’s Horizon 2020 research and innovation program under the Marie Skłodowska-Curie grant agreement No. 823860. https://cordis.europa.eu/project/id/823860/fr or https://www.toxicrop.com/toxicrop-flyer/ and co-supported by the strategical funding from FCT UIDB/04423/2020 and UIDP/04423/2020.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Madamwar, D.; Incharoensakdi, A. Bloom dynamics of cyanobacteria and their toxins: Environmental health impacts and mitigation strategies. Front. Microbiol. 2015, 6, 1254. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Otten, T.G. Duelling ‘CyanoHABs’: Unravelling the environmental drivers controlling dominance and succession among diazotrophic and non-N2-fixing harmful cyanobacteria. Environ. Microbiol. 2016, 18, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Khatri, N.; Tyagi, S. Influences of natural and anthropogenic factors on surface and groundwater quality in rural and urban areas. Front. Life Sci. 2015, 8, 23–39. [Google Scholar] [CrossRef]
- Lamastra, L.; Balderacchi, M.; Trevisan, M. Inclusion of emerging organic contaminants in groundwater monitoring plans. MethodsX 2016, 3, 459–476. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Steinman, A.D.; Wan, X.; Xie, L. Bioaccumulation of microcystin congeners in soil-plant system and human health risk assessment: A field study from Lake Taihu region of China. Environ. Pollut. 2018, 240, 44–50. [Google Scholar] [CrossRef] [PubMed]
- El Khalloufi, F.; Oufdou, K.; Lahrouni, M.; El Ghazali, I.; Saqrane, S.; Vasconcelos, V.; Oudra, B. Allelopatic effects of cyanobacteria extracts containing microcystins on Medicago sativa-Rhizobia symbiosis. Ecotoxicol. Environ. Saf. 2011, 74, 431–438. [Google Scholar] [CrossRef]
- Saqrane, S.; Ghazali, I.E.; Oudra, B.; Bouarab, L.; Vasconcelos, V. Effects of cyanobacteria producing microcystins on seed germination and seedling growth of several agricultural plants. J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes 2008, 43, 443–451. [Google Scholar] [CrossRef]
- do Carmo Bittencourt-Oliveira, M.; Cordeiro-Araújo, M.K.; Chia, M.A.; de Toledo Arruda-Neto, J.D.; de Oliveira, Ê.T.; dos Santos, F. Lettuce irrigated with contaminated water: Photosynthetic effects, antioxidative response and bioaccumulation of microcystin congeners. Ecotoxicol. Environ. Saf. 2016, 128, 83–90. [Google Scholar] [CrossRef]
- Lee, S.; Jiang, X.; Manubolu, M.; Riedl, K.; Ludsin, S.A.; Martin, J.F.; Lee, J. Fresh produce and their soils accumulate cyanotoxins from irrigation water: Implications for public health and food security. Food Res. Int. 2017, 102, 234–245. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, Q.; Ding, Q.; Yin, L.; Pu, Y. A novel and native microcystin-degrading bacterium of Sphingopyxis sp. isolated from lake Taihu. Int. J. Environ. Res. Public Health 2017, 14, 1187. [Google Scholar] [CrossRef]
- WHO. WHO Report on the Global Tobacco Epidemic, 2011: Warning about the Dangers of Tobacco; World Health Organization: Geneva, Switzerland, 2011; ISBN 9244564262. [Google Scholar]
- Poste, A.E.; Ozersky, T. Invasive dreissenid mussels and round gobies: A benthic pathway for the trophic transfer of microcystin. Environ. Toxicol. Chem. 2013, 32, 2159–2164. [Google Scholar] [CrossRef]
- Moy, N.J.; Dodson, J.; Tassone, S.J.; Bukaveckas, P.A.; Bulluck, L.P. Biotransport of algal toxins to riparian food webs. Environ. Sci. Technol. 2016, 50, 10007–10014. [Google Scholar] [CrossRef]
- Woller-Skar, M.M.; Jones, D.N.; Luttenton, M.R.; Russell, A.L. Microcystin detected in little brown bats (Myotis lucifugus). Am. Midl. Nat. 2015, 174, 331–334. [Google Scholar] [CrossRef]
- Mutoti, M.I.; Edokpayi, J.; Mutileni, N.; Durowoju, O.; Munyai, F.L. Cyanotoxins in groundwater; occurrence, potential sources, health impacts and knowledge gap for public health. Toxicon 2023, 226, 107077. [Google Scholar] [CrossRef]
- Menezes, C.; Churro, C.; Dias, E. Risk levels of toxic cyanobacteria in Portuguese recreational freshwaters. Toxins 2017, 9, 327. [Google Scholar] [CrossRef]
- Welten, R.D.; Meneely, J.P.; Chevallier, O.P.; Kosek, V.; Greer, B.; Hajšlová, J.; Elliott, C.T. Oral microcystin-LR does not cause hepatotoxicity in pigs: Is the risk of microcystin-LR overestimated? Expo. Health 2020, 12, 775–792. [Google Scholar] [CrossRef]
- Liu, Y.; Qi, C.-L.; Li, D.-W.; Li, H.-Y.; Li, R.-M.; Yang, W.-D. Microcystin-LR exposure interfered maintenance of colonic microenvironmental homeostasis in rat. Food Chem. Toxicol. 2023, 173, 113611. [Google Scholar] [CrossRef]
- Odriozola, E.; Ballabene, N.; Salamanco, A. Poisoning in cattle caused by blue-green algae. Rev. Argent. Microbiol. 1984, 16, 219–224. [Google Scholar]
- Puschner, B.; Galey, F.D.; Johnson, B.; Dickie, C.W.; Vondy, M.; Francis, T.; Holstege, D.M. Blue-green algae toxicosis in cattle. J. Am. Vet. Med. Assoc. 1998, 213, 1605–1607. [Google Scholar] [CrossRef]
- van der Merwe, D.; Sebbag, L.; Nietfeld, J.C.; Aubel, M.T.; Foss, A.; Carney, E. Investigation of a Microcystis aeruginosa cyanobacterial freshwater harmful algal bloom associated with acute microcystin toxicosis in a dog. J. Vet. Diagn. Investig. 2012, 24, 679–687. [Google Scholar] [CrossRef]
- Van De Merwe, K.; Van Dijk, H.; Zon, R. Eye movements as an indicator of situation awareness in a flight simulator experiment. Int. J. Aviat. Psychol. 2012, 22, 78–95. [Google Scholar] [CrossRef]
- Masango, C.A. Indigenous traditional knowledge protection: Prospects in South Africa’s intellectual property framework? South Afr. J. Libr. Inf. Sci. 2010, 76, 74–80. [Google Scholar]
- Alonso-Andicoberry, C.; García-Viliada, L.; Lopez-Rodas, V.; Costas, E. Catastrophic mortality of flamingos in a Spanish national park caused by cyanobacteria. Vet. Rec. 2002, 151, 706. [Google Scholar]
- Kim, M.S.; Lee, C. Ozonation of microcystins: Kinetics and toxicity decrease. Environ. Sci. Technol. 2019, 53, 6427–6435. [Google Scholar] [CrossRef]
- Sandermann Jr, H. Plant metabolism of xenobiotics. Trends Biochem. Sci. 1992, 17, 82–84. [Google Scholar] [CrossRef]
- Mishra, B.S.; Singh, M.; Aggrawal, P.; Laxmi, A. Glucose and auxin signaling interaction in controlling Arabidopsis thaliana seedlings root growth and development. PLoS ONE 2009, 4, e4502. [Google Scholar] [CrossRef]
- Von Hoffmann, V. Goûter le Monde. Une Histoire Culturelle du Goût à L’époque Moderne; Peter Lang: Bruxelles, Belgium, 2013; ISBN 2875741160. [Google Scholar]
- Kondo, N. Uptake, Metabolism, and Detoxification of Sulfur. In Air Pollution and Plant Biotechnology; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; p. 179. [Google Scholar]
- Manage, P.M.; Edwards, C.; Singh, B.K.; Lawton, L.A. Isolation and identification of novel microcystin-degrading bacteria. Appl. Environ. Microbiol. 2009, 75, 6924–6928. [Google Scholar] [CrossRef]
- Jones, G.J.; Orr, P.T. Release and degradation of microcystin following algicide treatment of a Microcystis aeruginosa bloom in a recreational lake, as determined by HPLC and protein phosphatase inhibition assay. Water Res. 1994, 28, 871–876. [Google Scholar] [CrossRef]
- Kultschar, B.; Llewellyn, C. Secondary Metabolites in Cyanobacteria. In Secondary Metabolites—Sources and Applications; IntechOpen: London, UK, 2018; p. 64. [Google Scholar]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Kaufononga, S.A.F.; Cary, S.C.; Hamilton, D.P. High levels of structural diversity observed in microcystins from Microcystis CAWBG11 and characterization of six new microcystin congeners. Mar. Drugs 2014, 12, 5372–5395. [Google Scholar] [CrossRef]
- Jaeg, J.-P. Microcystines: Intoxication des animaux domestiques et sécurité des aliments d’origine animale. Rev. Med. Vet. 2007, 1, 46–58. [Google Scholar]
- Botes, D.P.; Kruger, H.; Viljoen, C.C. Isolation and characterization of four toxins from the blue-green alga, Microcystis aeruginosa. Toxicon 1982, 20, 945–954. [Google Scholar] [CrossRef]
- Sivonen, K. Effects of light, temperature, nitrate, orthophosphate, and bacteria on growth of and hepatotoxin production by Oscillatoria agardhii strains. Appl. Environ. Microbiol. 1990, 56, 2658–2666. [Google Scholar] [CrossRef]
- Sivonen, K.; Namikoshi, M.; Evans, W.R.; Carmichael, W.W.; Sun, F.; Rouhiainen, L.; Luukkainen, R.; Rinehart, K.L. Isolation and characterization of a variety of microcystins from seven strains of the cyanobacterial genus Anabaena. Appl. Environ. Microbiol. 1992, 58, 2495–2500. [Google Scholar] [CrossRef]
- Luukkainen, R.; Sivonen, K.; Namikoshi, M.; Färdig, M.; Rinehart, K.L.; Niemelä, S.I. Isolation and identification of eight microcystins from thirteen Oscillatoria agardhii strains and structure of a new microcystin. Appl. Environ. Microbiol. 1993, 59, 2204–2209. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Boyer, G.L. Health impacts from cyanobacteria harmful algae blooms: Implications for the North American Great Lakes. Harmful Algae 2016, 54, 194–212. [Google Scholar] [CrossRef]
- Cao, L.; Massey, I.Y.; Feng, H.; Yang, F. A Review of Cardiovascular To xicity of Microcystins. Toxins 2019, 11, 507. [Google Scholar] [CrossRef]
- Dai, R.; Wang, P.; Jia, P.; Zhang, Y.; Chu, X.; Wang, Y. A review on factors affecting microcystins production by algae in aquatic environments. World J. Microbiol. Biotechnol. 2016, 32, 1–7. [Google Scholar] [CrossRef]
- Liu, J.; Chen, L.; Zhang, X. Current research scenario for biological effect of exogenous factors on microcystin synthesis. Environ. Sci. Pollut. Res. 2022, 29, 26190–26201. [Google Scholar] [CrossRef]
- Spoof, L.; Jaakkola, S.; Važić, T.; Häggqvist, K.; Kirkkala, T.; Ventelä, A.-M.; Kirkkala, T.; Svirčev, Z.; Meriluoto, J. Elimination of cyanobacteria and microcystins in irrigation water—Effects of hydrogen peroxide treatment. Environ. Sci. Pollut. Res. 2020, 27, 8638–8652. [Google Scholar] [CrossRef]
- El Ghazali, I.; Saqrane, S.; Saker, M.; Youness, O.; Oudra, B.; Vasconcelos, V.; Del Campo, F.F. Caractérisation biochimique et moléculaire d’efflorescences à cyanobactéries toxiques dans le réservoir Lalla Takerkoust (Maroc). Rev. Sci. L’eau 2011, 24, 117–128. [Google Scholar] [CrossRef]
- Rushford, C.A.; North, R.L.; Miller, G.L. Detection of cyanotoxins in irrigation water and potential impact on putting green health. Int. Turfgrass Soc. Res. J. 2022, 14, 994–996. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; El-Sharouny, H.M.; Ali, W.S.M. Microcystin production in benthic mats of cyanobacteria in the Nile River and irrigation canals, Egypt. Toxicon 2006, 47, 584–590. [Google Scholar] [CrossRef]
- WHO. WHO Cyanobacterial Toxins: Microcystins; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Liu, W.; Chen, C.; Chen, L.; Wang, L.; Li, J.; Chen, Y.; Jin, J.; Kawan, A.; Zhang, X. Sex-dependent effects of microcystin-LR on hypothalamic-pituitary-gonad axis and gametogenesis of adult zebrafish. Sci. Rep. 2016, 6, 22819. [Google Scholar] [CrossRef]
- Crush, J.R.; Briggs, L.R.; Sprosen, J.M.; Nichols, S.N. Effect of irrigation with lake water containing microcystins on microcystin content and growth of ryegrass, clover, rape, and lettuce. Environ. Toxicol. Int. J. 2008, 23, 246–252. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Al Shehri, A.M. Microcystin-producing blooms of Anabaenopsis arnoldi in a potable mountain lake in Saudi Arabia. FEMS Microbiol. Ecol. 2009, 69, 98–105. [Google Scholar] [CrossRef]
- Levizou, E.; Papadimitriou, T.; Papavasileiou, E.; Papadimitriou, N.; Kormas, K.A. Root vegetables bioaccumulate microcystins-LR in a developmental stage-dependent manner under realistic exposure scenario: The case of carrot and radish. Agric. Water Manag. 2020, 240, 106274. [Google Scholar] [CrossRef]
- Xiang, L.; Li, Y.-W.; Liu, B.-L.; Zhao, H.-M.; Li, H.; Cai, Q.-Y.; Mo, C.-H.; Wong, M.-H.; Li, Q.X. High ecological and human health risks from microcystins in vegetable fields in southern China. Environ. Int. 2019, 133, 105142. [Google Scholar] [CrossRef]
- Tsoumalakou, E.; Papadimitriou, T.; Berillis, P.; Kormas, K.A.; Levizou, E. Spray irrigation with microcystins-rich water affects plant performance from the microscopic to the functional level and food safety of spinach (Spinacia oleracea L.). Sci. Total Environ. 2021, 789, 147948. [Google Scholar] [CrossRef]
- Cao, H.; Han, L. Hourly remote sensing monitoring of harmful algal blooms (HABs) in Taihu Lake based on GOCI images. Environ. Sci. Pollut. Res. 2021, 28, 35958–35970. [Google Scholar] [CrossRef]
- Trifirò, G.; Barbaro, E.; Gambaro, A.; Vita, V.; Clausi, M.T.; Franchino, C.; Palumbo, M.P.; Floridi, F.; De Pace, R. Quantitative determination by screening ELISA and HPLC-MS/MS of microcystins LR, LY, LA, YR, RR, LF, LW, and nodularin in the water of Occhito lake and crops. Anal. Bioanal. Chem. 2016, 408, 7699–7708. [Google Scholar] [CrossRef]
- Petrou, M.; Karas, P.A.; Vasileiadis, S.; Zafiriadis, I.; Papadimitriou, T.; Levizou, E.; Kormas, K.; Karpouzas, D.G. Irrigation of radish (Raphanus sativus L.) with microcystin-enriched water holds low risk for plants and their associated rhizopheric and epiphytic microbiome. Environ. Pollut. 2020, 266, 115208. [Google Scholar] [CrossRef]
- Bakr, A.; Alzain, M.N.; Alzamel, N.M.; Loutfy, N. Accumulation of microcystin from Oscillatoria limnetica Lemmermann and Microcystis aeruginosa (Kützing) in two leafy green vegetable crop plants Lactuca sativa L. and Eruca sativa. Plants 2022, 11, 1733. [Google Scholar] [CrossRef]
- Wijewickrama, M.M.; Manage, P.M. Accumulation of Microcystin-LR in grains of two rice varieties (Oryza sativa L.) and a leafy vegetable, Ipomoea aquatica. Toxins 2019, 11, 432. [Google Scholar] [CrossRef]
- Douma, M.; Loudiki, M.; Oudra, B.; Mouhri, K.; Ouahid, Y.; del Campo, F.F. Taxonomic diversity and toxicological assessment of Cyanobacteria in Moroccan inland waters. Rev. Sci. L’eau 2009, 22, 435–449. [Google Scholar] [CrossRef]
- Oudra, B.; Loudiki, M.; Vasconcelos, V.; Sabour, B.; Sbiyyaa, B.; Oufdou, K.; Mezrioui, N. Detection and quantification of microcystins from cyanobacteria strains isolated from reservoirs and ponds in Morocco. Environ. Toxicol. Int. J. 2002, 17, 32–39. [Google Scholar] [CrossRef]
- Aboal, M.; Puig, M. Intracellular and dissolved microcystin in reservoirs of the river Segura basin, Murcia, SE Spain. Toxicon 2005, 45, 509–518. [Google Scholar] [CrossRef]
- Haida, M.; El Khalloufi, F.; Mugani, R.; Redouane, E.M.; Campos, A.; Vasconcelos, V.; Oudra, B. Effects of Irrigation with Microcystin-Containing Water on Growth, Physiology, and Antioxidant Defense in Strawberry Fragaria vulgaris under Hydroponic Culture. Toxins 2022, 14, 198. [Google Scholar] [CrossRef]
- Cao, X.; Gu, Y.; Fang, Y.; Johnson, D.; Chen, C.; Chen, J.; Tian, H.; Huang, Y. Self-assembled BiVO4 mesocrystals for efficient photocatalytic decontamination of microcystin-LR. Environ. Chem. Lett. 2022, 20, 1595–1601. [Google Scholar] [CrossRef]
- Cao, Q.; Rediske, R.R.; Yao, L.; Xie, L. Effect of microcystins on root growth, oxidative response, and exudation of rice (Oryza sativa). Ecotoxicol. Environ. Saf. 2018, 149, 143–149. [Google Scholar] [CrossRef]
- Llana-Ruiz-Cabello, M.; Jos, A.; Cameán, A.; Oliveira, F.; Barreiro, A.; Machado, J.; Azevedo, J.; Pinto, E.; Almeida, A.; Campos, A. Analysis of the Use of Cylindrospermopsin and/or Microcystin-Contaminated Water in the Growth, Mineral Content, and Contamination of Spinacia oleracea and Lactuca sativa. Toxins 2019, 11, 624. [Google Scholar] [CrossRef]
- Liang, C.; Liu, H. Response of hormone in rice seedlings to irrigation contaminated with cyanobacterial extract containing microcystins. Chemosphere 2020, 256, 127157. [Google Scholar] [CrossRef]
- Liang, Y.; Chen, X.; Lu, X.; Jin, S.; Min, Y.; Yang, J. Combined effects of microcystin and nitrite on the growth, lipid peroxidation, and antioxidant responses of the freshwater rotifer Brachionus calycif lorus. Aquat. Toxicol. 2017, 192, 78–88. [Google Scholar] [CrossRef]
- Kuang, X.; Gu, J.-D.; Tie, B.; Yao, B.; Shao, J. Interactive effects of cadmium and Microcystis aeruginosa (cyanobacterium) on the growth, antioxidative responses and accumulation of cadmium and microcystins in rice seedlings. Ecotoxicology 2016, 25, 1588–1599. [Google Scholar] [CrossRef]
- Liang, Y.; Lu, X.; Min, Y.; Liu, L.; Yang, J. Interactive effects of microcystin and ammonia on the reproductive performance and phenotypic traits of the rotifer Brachionus calyciflorus. Ecotoxicol. Environ. Saf. 2018, 147, 413–422. [Google Scholar] [CrossRef]
- Gu, Y.; Liang, C. Responses of antioxidative enzymes and gene expression in Oryza sativa L and Cucumis sativus L seedlings to microcystins stress. Ecotoxicol. Environ. Saf. 2020, 193, 110351. [Google Scholar] [CrossRef]
- Zhu, J.; Ren, X.; Liu, H.; Liang, C. Effect of irrigation with microcystins-contaminated water on growth and fruit quality of Cucumis sativus L. and the health risk. Agric. Water Manag. 2018, 204, 91–99. [Google Scholar] [CrossRef]
- Freitas, M.; Campos, A.; Azevedo, J.; Barreiro, A.; Planchon, S.; Renaut, J.; Vasconcelos, V. Lettuce (Lactuca sativa L.) leaf-proteome profiles after exposure to cylindrospermopsin and a microcystin-LR/cylindrospermopsin mixture: A concentration-dependent response. Phytochemistry 2015, 110, 91–103. [Google Scholar] [CrossRef]
- Cao, Q.; Steinman, A.D.; Wan, X.; Xie, L. Combined toxicity of microcystin-LR and copper on lettuce (Lactuca sativa L.). Chemosphere 2018, 206, 474–482. [Google Scholar] [CrossRef]
- Lahrouni, M.; Oufdou, K.; Khalloufi, F.E.; Pajuelo, E.; Oudra, B. Impact of cyanobacterial toxins (microcystins) on growth and root development of in vitro Vicia faba cultures. Int. J. Innov. Appl. Stud. 2015, 12, 2028–9324. [Google Scholar]
- Cao, Q.; Steinman, A.D.; Yao, L.; Xie, L. Increment of root membrane permeability caused by microcystins result in more elements uptake in rice (Oryza sativa). Ecotoxicol. Environ. Saf. 2017, 145, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Wang, W. Response and recovery of rice (Oryza sativa) seedlings to irrigation with microcystin-contaminated water. Environ. Earth Sci. 2015, 73, 4573–4580. [Google Scholar] [CrossRef]
- Liang, C.; Ma, X.; Liu, H. Effect of microcystins at different rice growth stages on its yield, quality, and safety. Environ. Sci. Pollut.Res. 2020, 28, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mitrovic, S.M.; Allis, O.; Furey, A.; James, K.J. Bioaccumulation and harmful effects of microcystin-LR in the aquatic plants Lemna minor and Wolffia arrhiza and the filamentous alga Chladophora fracta. Ecotoxicol. Environ. Saf. 2005, 61, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Corbel, S.; Bouaïcha, N.; Mougin, C. Dynamics of the toxic cyanobacterial microcystin-leucine-arginine peptide in agricultural soil. Environ. Chem. Lett. 2014, 12, 535–541. [Google Scholar] [CrossRef]
- WHO. Guidelines for drinking-water quality. WHO Chron. 2011, 38, 104–108. [Google Scholar]
- Pflugmacher, S.; Wiegand, C.; Beattie, K.A.; Krause, E.; Steinberg, C.E.W.; Codd, G.A. Uptake, effects, and metabolism of cyanobacterial toxins in the emergent reed plant Phragmites australis (cav.) trin. ex steud. Environ. Toxicol. Chem. Int. J. 2001, 20, 846–852. [Google Scholar] [CrossRef]
- Gutiérrez-Praena, D.; Campos, A.; Azevedo, J.; Neves, J.; Freitas, M.; Guzmán-Guillén, R.; Cameán, A.M.; Renaut, J.; Vasconcelos, V. Exposure of Lycopersicon esculentum to microcystin-LR: Effects in the leaf proteome and toxin translocation from water to leaves and fruits. Toxins 2014, 6, 1837–1854. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Tsuji, K.; Watanabe, M.F.; Kondo, F. Stability of microcystins from cyanobacteria—III. Effect of pH and temperature. Phycologia 1996, 35, 83–88. [Google Scholar] [CrossRef]
- Zhang, X.-X.; Zhang, Z.; Fu, Z.; Wang, T.; Qin, W.; Xu, L.; Cheng, S.; Yang, L. Stimulation effect of microcystin-LR on matrix metalloproteinase-2/-9 expression in mouse liver. Toxicol. Lett. 2010, 199, 377–382. [Google Scholar] [CrossRef]
- Tsuji, K.; Watanuki, T.; Kondo, F.; Watanabe, M.F.; Suzuki, S.; Nakazawa, H.; Suzuki, M.; Uchida, H.; Harada, K.-I. Stability of microcystins from cyanobacteria—II. Effect of UV light on decomposition and isomerization. Toxicon 1995, 33, 1619–1631. [Google Scholar] [CrossRef]
- Eynard, F.; Mez, K.; Walther, J.-L. Risk of cyanobacterial toxins in Riga waters (Latvia). Water Res. 2000, 34, 2979–2988. [Google Scholar] [CrossRef]
- Corbel, S.; Mougin, C.; Bouaïcha, N. Cyanobacterial toxins: Modes of actions, fate in aquatic and soil ecosystems, phytotoxicity and bioaccumulation in agricultural crops. Chemosphere 2014, 96, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xie, P.; Zhang, D.; Ke, Z.; Yang, H. In situ studies on the bioaccumulation of microcystins in the phytoplanktivorous silver carp (Hypophthalmichthys molitrix) stocked in Lake Taihu with dense toxic Microcystis blooms. Aquaculture 2006, 261, 1026–1038. [Google Scholar] [CrossRef]
- Haida, M.; Tamegart, L.; Mugani, R.; Essadki, Y.; Azevedo, J.; Araújo, M.J.; Campos, A.; Vasconcelos, V.; Gamrani, H.; Oudra, B. Tracing the fate of microcystins from irrigation water to food chains: Studies with Fragaria vulgaris and Meriones shawi. Toxicon 2023, 236, 107345. [Google Scholar] [CrossRef] [PubMed]
- Pires, L.M.D.; Karlsson, K.M.; Meriluoto, J.A.O.; Kardinaal, E.; Visser, P.M.; Siewertsen, K.; Van Donk, E.; Ibelings, B.W. Assimilation and depuration of microcystin–LR by the zebra mussel, Dreissena polymorpha. Aquat. Toxicol. 2004, 69, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Park, H.-D. Determination of microcystins in fish tissues using HPLC with a rapid and efficient solid phase extraction. Aquaculture 2007, 271, 530–536. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.d.S.; Kozlowsky-Suzuki, B. Cyanotoxins: Bioaccumulation and effects on aquatic animals. Mar. Drugs 2011, 9, 2729–2772. [Google Scholar] [CrossRef]
- Zhang, D.; Deng, X.; Xie, P.; Chen, J.; Guo, L. Risk assessment of microcystins in silver carp (Hypophthalmichthys molitrix) from eight eutrophic lakes in China. Food Chem. 2013, 140, 17–21. [Google Scholar] [CrossRef]
- Lance, E.; Petit, A.; Sanchez, W.; Paty, C.; Gerard, C.; Bormans, M. Evidence of trophic transfer of microcystins from the gastropod Lymnaea stagnalis to the fish Gasterosteus aculeatus. Harmful Algae 2014, 31, 9–17. [Google Scholar] [CrossRef]
- Toporowska, M.; Pawlik-Skowronska, B.; Kalinowska, R. Accumulation and effects of cyanobacterial microcystins and anatoxin-a on benthic larvae of Chironomus spp. (Diptera: Chironomidae). Eur. J. Entomol. 2014, 111, 83. [Google Scholar] [CrossRef]
- Orr, P.T.; Jones, G.J.; Hunter, R.A.; Berger, K. Exposure of beef cattle to sub-clinical doses of Microcystis aeruginosa: Toxin bioaccumulation, physiological effects and human health risk assessment. Toxicon 2003, 41, 613–620. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, D.; Xie, P.; Wang, Q.; Ma, Z. Simultaneous determination of microcystin contaminations in various vertebrates (fish, turtle, duck and water bird) from a large eutrophic Chinese lake, Lake Taihu, with toxic Microcystis blooms. Sci. Total Environ. 2009, 407, 3317–3322. [Google Scholar] [CrossRef]
- Woller-Skar, M.M.; Russell, A.L.; Gaskill, J.A.; Luttenton, M.R. Microcystin in multiple life stages of Hexagenia limbata, with implications for toxin transfer. J. Great Lakes Res. 2020, 46, 666–671. [Google Scholar] [CrossRef]
- Malbrouck, C.; Kestemont, P. Effects of microcystins on fish. Environ. Toxicol. Chem. Int. J. 2006, 25, 72–86. [Google Scholar] [CrossRef] [PubMed]
- Nasri, H.; El Herry, S.; Bouaïcha, N. First reported case of turtle deaths during a toxic Microcystis spp. bloom in Lake Oubeira, Algeria. Ecotoxicol. Environ. Saf. 2008, 71, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, K.; Gomes, A.; Calado, L.; Yasui, G.; Assis, D.; Henry, T.; Fonseca, A.; Pinto, E. Toxicity of cyanopeptides from two microcystis strains on larval development of Astyanax altiparanae. Toxins 2019, 11, 220. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Paerl, H.W.; Zhu, G.; Wu, T.; Li, W.; Shi, K.; Zhao, L.; Zhang, Y.; Qin, B.; Caruso, A.M. The role of tropical cyclones in stimulating cyanobacterial (Microcystis spp.) blooms in hypertrophic Lake Taihu, China. Harmful Algae 2014, 39, 310–321. [Google Scholar] [CrossRef]
- Bouaïcha, N.; Miles, C.O.; Beach, D.G.; Labidi, Z.; Djabri, A.; Benayache, N.Y.; Nguyen-Quang, T. Structural diversity, characterization and toxicology of microcystins. Toxins 2019, 11, 714. [Google Scholar] [CrossRef]
- Wang, X.; Ying, F.; Chen, Y.; Han, X. Microcystin (-LR) affects hormones level of male mice by damaging hypothalamic-pituitary system. Toxicon 2012, 59, 205–214. [Google Scholar] [CrossRef]
- Nishiwaki-Matsushima, R.; Ohta, T.; Nishiwaki, S.; Suganuma, M.; Kohyama, K.; Ishikawa, T.; Carmichael, W.W.; Fujiki, H. Liver tumor promotion by the cyanobacterial cyclic peptide toxin microcystin-LR. J. Cancer Res. Clin. Oncol. 1992, 118, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, Z.; Zhou, N.; Shen, J.; Ye, M. Degradation and detoxification of microcystin-LR in drinking water by sequential use of UV and ozone. J. Environ. Sci. 2010, 22, 1897–1902. [Google Scholar] [CrossRef]
- Zhang, H.-Z.; Zhang, F.-Q.; Li, C.-F.; Yi, D.; Fu, X.-L.; Cui, L.-X. A cyanobacterial toxin, microcystin-LR, induces apoptosis of sertoli cells by changing the expression levels of apoptosis-related proteins. Tohoku J. Exp. Med. 2011, 224, 235–242. [Google Scholar] [CrossRef]
- Yuan, G.; Xie, P.; Zhang, X.; Tang, R.; Gao, Y.; Li, D.; Li, L. In vivo studies on the immunotoxic effects of microcystins on rabbit. Environ. Toxicol. 2012, 27, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Briand, J.-F.; Jacquet, S.; Bernard, C.; Humbert, J.-F. Health hazards for terrestrial vertebrates from toxic cyanobacteria in surface water ecosystems. Vet. Res. 2003, 34, 361–377. [Google Scholar] [CrossRef] [PubMed]
- Stewart, I.; Seawright, A.A.; Shaw, G.R. Cyanobacterial poisoning in livestock, wild mammals and birds—An overview. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Springer: Berlin/Heidelberg, Germany, 2008; pp. 613–637. [Google Scholar]
- Mez, K.; Beattie, K.A.; Codd, G.A.; Hanselmann, K.; Hauser, B.; Naegeli, H.; Preisig, H.R. Identification of a microcystin in benthic cyanobacteria linked to cattle deaths on alpine pastures in Switzerland. Eur. J. Phycol. 1997, 32, 111–117. [Google Scholar] [CrossRef]
- Carbis, C.R.; Waldron, D.L.; Mitchell, G.F.; Anderson, J.W.; McCauley, I. Recovery of hepatic function and latent mortalities in sheep exposed to the blue-green alga Microcystis aeruginosa. Vet. Rec. 1995, 137, 12–15. [Google Scholar] [CrossRef]
- Wood, S.A.; Rueckert, A.; Hamilton, D.P.; Cary, S.C.; Dietrich, D.R. Switching toxin production on and off: Intermittent microcystin synthesis in a Microcystis bloom. Environ. Microbiol. Rep. 2011, 3, 118–124. [Google Scholar] [CrossRef]
- Soll, M.P.; Williams, M.C. Mortality of white rhinoceros (Ceratotherium simum) suspected to be associated with the blue-green alga Microcystis aeruginosa. J. S. Afr. Vet. Assoc. 1985, 56, 49–51. [Google Scholar]
- Cao, L.; Huang, F.; Massey, I.Y.; Wen, C.; Zheng, S.; Xu, S.; Yang, F. Effects of microcystin-LR on the microstructure and inflammation-related factors of jejunum in mice. Toxins 2019, 11, 482. [Google Scholar] [CrossRef]
- Xu, S.; Yi, X.; Liu, W.; Zhang, C.; Massey, I.Y.; Yang, F.; Tian, L. A review of nephrotoxicity of microcystins. Toxins 2020, 12, 693. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W.J.; Altheimer, S.; Cattori, V.; Meier, P.J.; Dietrich, D.R.; Hagenbuch, B. Organic anion transporting polypeptides expressed in liver and brain mediate uptake of microcystin. Toxicol. Appl. Pharmacol. 2005, 203, 257–263. [Google Scholar] [CrossRef]
- Massey, I.Y.; Yang, F.; Ding, Z.; Yang, S.; Guo, J.; Al-Osman, M.; Kamegni, R.B.; Zeng, W. Exposure routes and health effects of microcystins on animals and humans: A mini-review. Toxicon 2018, 151, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Drobac, D.; Tokodi, N.; Lujić, J.; Marinović, Z.; Subakov-Simić, G.; Dulić, T.; Važić, T.; Nybom, S.; Meriluoto, J.; Codd, G.A. Cyanobacteria and cyanotoxins in fishponds and their effects on fish tissue. Harmful Algae 2016, 55, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Badar, M.; Batool, F.; Khan, S.S.; Khokhar, I.; Qamar, M.K.; Yasir, C. Effects of microcystins toxins contaminated drinking water on hepatic problems in animals (cows and buffalos) and toxins removal chemical method. Buffalo Bull. 2017, 36, 43–56. [Google Scholar]
- Yang, F.; Guo, J.; Huang, F.; Massey, I.Y.; Huang, R.; Li, Y.; Wen, C.; Ding, P.; Zeng, W.; Liang, G. Removal of microcystin-LR by a novel native effective bacterial community designated as YFMCD4 isolated from Lake Taihu. Toxins 2018, 10, 363. [Google Scholar] [CrossRef] [PubMed]
- Piyathilaka, M.; Pathmalal, M.M.; Tennekoon, K.H.; De Silva, B.; Samarakoon, S.R.; Chanthirika, S. Microcystin-LR-induced cytotoxicity and apoptosis in human embryonic kidney and human kidney adenocarcinoma cell lines. Microbiology 2015, 161, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Kubickova, B.; Babica, P.; Hilscherová, K.; Šindlerová, L. Effects of cyanobacterial toxins on the human gastrointestinal tract and the mucosal innate immune system. Environ. Sci. Eur. 2019, 31, 1–27. [Google Scholar] [CrossRef]
- Xie, L.; Xie, P.; Guo, L.; Li, L.; Miyabara, Y.; Park, H. Organ distribution and bioaccumulation of microcystins in freshwater fish at different trophic levels from the eutrophic Lake Chaohu, China. Environ. Toxicol. Int. J. 2005, 20, 293–300. [Google Scholar] [CrossRef]
- Giblin, S.M.; Larson, J.H.; King, J.D. Environmental drivers of cyanobacterial abundance and cyanotoxin production in backwaters of the Upper Mississippi River. River Res. Appl. 2022, 38, 1115–1128. [Google Scholar] [CrossRef]
- Zamora-Barrios, C.A.; Nandini, S.; Sarma, S.S.S. Bioaccumulation of microcystins in seston, zooplankton and fish: A case study in Lake Zumpango, Mexico. Environ. Pollut. 2019, 249, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Wang, Y.; Zhang, K.; Ding, Y.; Zhang, W.; Wu, M.; Xiao, B.; Gu, P. Transmission of microcystins in natural systems and resource processes: A review of potential risks to humans health. Toxins 2023, 15, 448. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, N.P.E. Role of metabolism in chemecal toxicity. In Cytochromes P450 Metabolic and Toxicological Aspects; CRC Press: Boca Raton, FL, USA, 1996; Volume 29. [Google Scholar]
- Manahan, S.E. Toxicological Chemistry and Biochemistry; CRC Press: Boca Raton, FL, USA, 2002; ISBN 1420032127. [Google Scholar]
- Lilja, H.; Jeppsson, J.-O.; Kristensson, H. Evaluation of serum gamma-glutamyltransferase by electrofocusing, and variations in isoform patterns. Clin. Chem. 1983, 29, 1034–1037. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.R.; Wilhelm, S.W.; Boyer, G.L. The fate of microcystins in the environment and challenges for monitoring. Toxins 2014, 6, 3354–3387. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K.; Jones, G. Cyanobacterial toxins. Toxic Cyanobacteria. Water A Guide Their Health Conseq. Monit. Manag. 1999, 1, 43–112. [Google Scholar]
- Guo, Y.C.; Lee, A.K.; Yates, R.S.; Liang, S.; Rochelle, P.A. Analysis of microcystins in drinking water by ELISA and LC/MS/MS. J. Am. Water Work. Assoc. 2017, 109, 13–25. [Google Scholar] [CrossRef]
- Saito, T.; Okano, K.; Park, H.-D.; Itayama, T.; Inamori, Y.; Neilan, B.A.; Burns, B.P.; Sugiura, N. Detection and sequencing of the microcystin LR-degrading gene, mlrA, from new bacteria isolated from Japanese lakes. FEMS Microbiol. Lett. 2003, 229, 271–276. [Google Scholar] [CrossRef]
- Bourne, D.G.; Jones, G.J.; Blakeley, R.L.; Jones, A.; Negri, A.P.; Riddles, P. Enzymatic pathway for the bacterial degradation of the cyanobacterial cyclic peptide toxin microcystin LR. Appl. Environ. Microbiol. 1996, 62, 4086–4094. [Google Scholar] [CrossRef] [PubMed]
- Alamri, S.A. Biodegradation of microcystin by a new Bacillus sp. isolated from a Saudi freshwater lake. Afr. J. Biotechnol. 2010, 9, 6552–6559. [Google Scholar]
- Lawton, L.A.; Welgamage, A.; Manage, P.M.; Edwards, C. Novel bacterial strains for the removal of microcystins from drinking water. Water Sci. Technol. 2011, 63, 1137–1142. [Google Scholar] [CrossRef]
- Lemes, G.A.F.; Kist, L.W.; Bogo, M.R.; Yunes, J.S. Biodegradation of [D-Leu1] microcystin-LR by a bacterium isolated from sediment of Patos Lagoon estuary, Brazil. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Idroos, F.S.; De Silva, B.; Manage, P.M. Biodegradation of microcystin analogues by Stenotrophomonas maltophilia isolated from Beira Lake Sri Lanka. J. Natl. Sci. Found. Sri Lanka 2017, 45, 91–99. [Google Scholar] [CrossRef]
- Yang, F.; Huang, F.; Feng, H.; Wei, J.; Massey, I.Y.; Liang, G.; Zhang, F.; Yin, L.; Kacew, S.; Zhang, X. A complete route for biodegradation of potentially carcinogenic cyanotoxin microcystin-LR in a novel indigenous bacterium. Water Res. 2020, 174, 115638. [Google Scholar] [CrossRef]
- Massey, I.Y.; Yang, F. A mini review on microcystins and bacterial degradation. Toxins 2020, 12, 268. [Google Scholar] [CrossRef] [PubMed]
- Briand, E.; Humbert, J.; Tambosco, K.; Bormans, M.; Gerwick, W.H. Role of bacteria in the production and degradation of Microcystis cyanopeptides. Microbiologyopen 2016, 5, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Bourne, D.G.; Riddles, P.; Jones, G.J.; Smith, W.; Blakeley, R.L. Characterisation of a gene cluster involved in bacterial degradation of the cyanobacterial toxin microcystin LR. Environ. Toxicol. 2001, 16, 523–534. [Google Scholar] [CrossRef]
- Ding, Q.; Liu, K.; Xu, K.; Sun, R.; Zhang, J.; Yin, L.; Pu, Y. Further understanding of degradation pathways of microcystin-LR by an indigenous Sphingopyxis sp. in environmentally relevant pollution concentrations. Toxins 2018, 10, 536. [Google Scholar] [CrossRef]
Figure 2.
Diagram showing the bioaccumulation, transfer, and effect of MCs in the terrestrial ecosystem.
Figure 2.
Diagram showing the bioaccumulation, transfer, and effect of MCs in the terrestrial ecosystem.

Figure 3.
Glutathione metabolic pathway for microcystin-LR [132].
Figure 3.
Glutathione metabolic pathway for microcystin-LR [132].

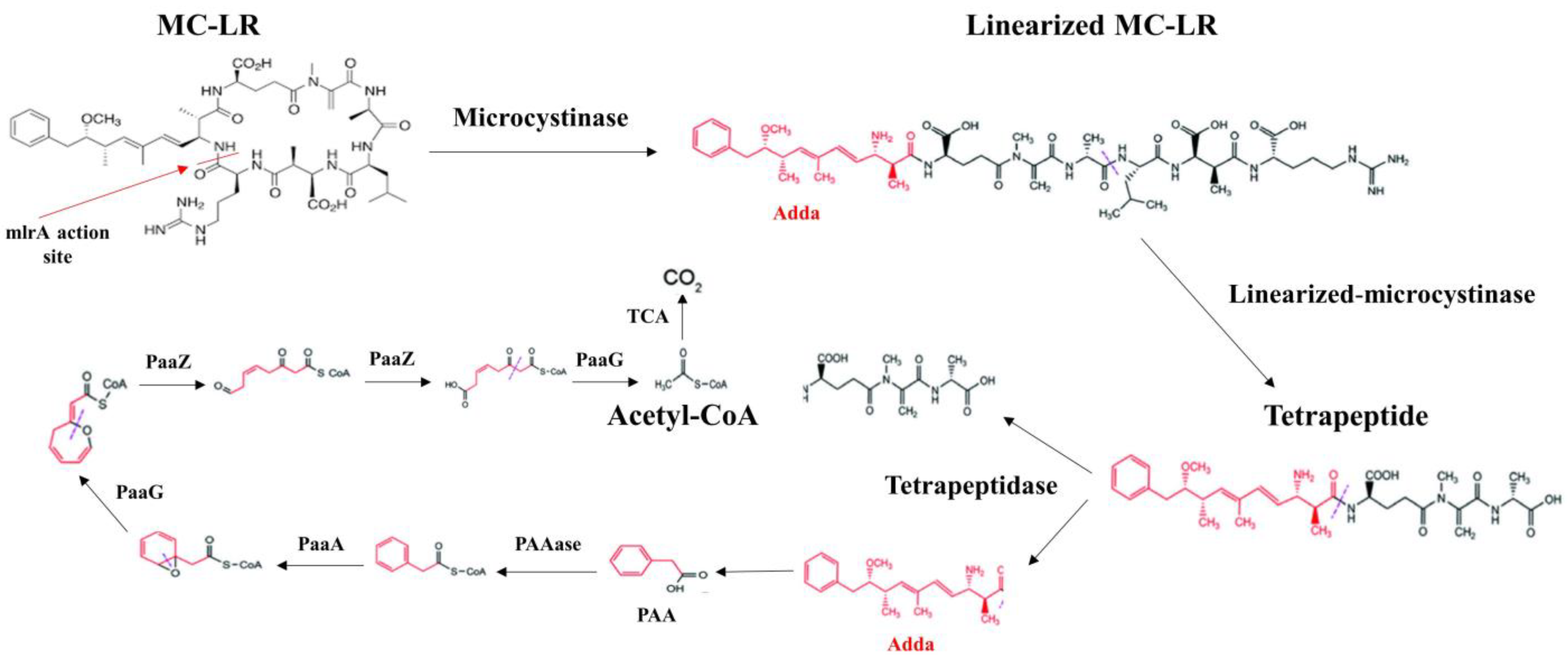
Figure 4.
Biodegradation pathway of MC-LR by Sphingopyxis sp. YF1. A potential connection was found between the mlrABCD gene cluster and the paa gene clusters involved in the degradation of phenylacetic acid (PAA). The process of degradation proceeded as follows: sequentially, MC-LR was broken down into linearized MC-LR, Adda, and tetrapeptide. After that, Adda was broken down into PAA, and PAAase was then changed into PAA-CoA. PAA-CoA was further broken down to acetyl coenzyme A (acetyl-CoA) by PaaA homologs PaaG and PaaZ. Ultimately, acetyl-CoA was entirely transformed into CO2 by the tricarboxylic acid (TCA) cycle. Based on available data, it is highly probable that PAA is the subsequent metabolic product resulting from Adda’s conversion to PAA [141].
Figure 4.
Biodegradation pathway of MC-LR by Sphingopyxis sp. YF1. A potential connection was found between the mlrABCD gene cluster and the paa gene clusters involved in the degradation of phenylacetic acid (PAA). The process of degradation proceeded as follows: sequentially, MC-LR was broken down into linearized MC-LR, Adda, and tetrapeptide. After that, Adda was broken down into PAA, and PAAase was then changed into PAA-CoA. PAA-CoA was further broken down to acetyl coenzyme A (acetyl-CoA) by PaaA homologs PaaG and PaaZ. Ultimately, acetyl-CoA was entirely transformed into CO2 by the tricarboxylic acid (TCA) cycle. Based on available data, it is highly probable that PAA is the subsequent metabolic product resulting from Adda’s conversion to PAA [141].

Table 1.
Occurrence of MC congeners in freshwater destined for irrigation around the world.
| Lakes/Reservoirs | Location | Cyanobacteria | Extracellular MCs (µg/L) | Intracellular MCs | Congeners | References |
|---|---|---|---|---|---|---|
| Hakanoa Lake | North Island, New Zealand | Microcystis aeruginosa; Anabaena cf. smithii | 2100 | ND | MC-RR, -LR, -FR, -WR, -LY, -AR, -LA, and YR | [50] |
| Köyliönjärvi Lake | southwest Finland | Microcystis spp. (M. wesenbergii, M. botrys and M. aeruginosa); Dolichospermumsp., and Aphanizomenon sp.; Planktothrix sp.; Aulacoseira ambigua and Cyclotella spp.; Planktolyngbya limnetica; | 0.12–0.28 | 0.97–2.4 µg/L | MC-RR, -LR, -YR, and -RR | [46] |
| Groundwater wells | Asir region, Saudi Arabia | Oscillatoria limnetica | 0.3–1.8 | 336 µg/g | MC-RR, -LR, and -YR | [51] |
| Karla Reservoir | Central Greece | Planktothrix cf. agardhii; Anabaena sp. | 1.5–33 | ND | MC-LR, and -RR | [52] |
| Dianchi Lake; Xingyun Lake | Yunnan province, China | ND | 0.117–46.7 | 0.062–514.6 µg/L | MC-RR, -LR, and -YR | [53] |
| Dashahe Reservoir | Jiangmen, Guangdong province, China | 0.016–3.1 | 0.594–450.7 µg/L | |||
| Karla Reservoir | Central Greece | ND | 3.8 | ND | MCt | [54] |
| Nile river | Egypt | Anabaena subcylindrica; Nostoc spongiaeforme; Plectonema boryanum; Phormidium corium; Aulacoseira ambigua and Cyclotella sp. | ND | 1.6–3.66 µg/L | MC-RR, and -YR | [47] |
| Lalla Takerkoust reservoir | Marrakech, Morocco | Microcystis aeruginosa Kütz | 60 | 3240 µg/g | MCt | [43] |
| Taihu Lake | Suzhou, China | Microcystis | 4.14 | 17.57 µg/L | MC-RR, -LR, and -YR | [55] |
| Occhito reservoir | Italy | ND | 0.18 µg/L | ND | MC-LR, -RR, -LA, -YR, -LY, -LF, and -LW | [56] |
| Karla Reservoir | Central Greece | Microcystis aeruginosa | 1.43–2.03 | ND | MC-LR, and -RR | [57] |
| Sources of irrigation water | Egypt | Oscillatoria limnetica and Microcystis aeruginosa | 45.04–600 | 58,000–87,000 µg/L | MC-LR, and -RR | [58] |
| Beira Lake | Sri Lanka | Microcystis aeruginosa | 180 | 340 µg/g | MC-LR | [59] |
| Irrigation heads; Irrigation intakes; Epilimnion of surface water sources | Missouri and Kansas, USA | ND | 8.53–8.65 | ND | ND | [46] |
| Mansour Eddahbi Reservoir | South of Ouarzazate city, Morocco | Microcystis aeruginosa Kütz; Pseudanabaena papillaterminata Kuk; and Oscillatoria sp. | ND | 64.4 μg/g | MC, -LR, -RR, -YR, -FR, and -WR | [60] |
| Lalla Takerkoust reservoir | Central regions of Morocco | Microcystis aeruginosa | ND | 2.2–944 µg/g | MC-LR | [61] |
| Reservoirs of the river Segura | Murcia, SE Spain | Aphanizomenon flos-aquae and Microcystis aeruginosa | 0.067–1.586 | ND | MC-RR, -LR, and -YR | [62] |
ND: Not determined; MCt: Total MCs.
Table 3.
A view of MC bioaccumulation in hydroponically irrigated plants.
| Plants Species | Applied Concentration (μg/L) | Organs | Concentration Accumulated (µg/kg) | Exposure Time (Days) | EDIAdu/EDIchi (μg/kg) | BFA of MCs | Reference |
|---|---|---|---|---|---|---|---|
| Lactuca sativa | 5–2000 MC-LR (P) | Leaves | 700–1400 MC-LR FW | 14 | 0.43–0.86/0.28–0.56 | 140/0.7 | [74] |
| Cucumis sativus L. | 1–1000 CE FW MCt | Fruit | 2.87–29.64 MCt FW | 7 | 0.001–0.018/0.001–0.011 | 0.29 | [72] |
| Oryza sativa | 5–500 CE FW MCt | Roots | ≈7000–35,000 MCt FW | 30 | - | 1400/70 | [76] |
| Stems | ≈10,000–38,000 MCt FW | - | 2000/76 | ||||
| Leaves | ≈20,000–112,000 MCt FW | - | 4000/224 | ||||
| Oryza sativa | 1–3000 CE FW MCt | Roots | ≈20–275 MCt FW | 7 | - | 20/0.091 | [78] |
| Stems | ≈18–30 MCt FW | - | 18/0.01 | ||||
| Leaves | ≈22–230 MCt FW | - | 22/0.076 | ||||
| Oryza sativa | 9.79 CE FW MCt | Roots | ≈1.22 MCt FW | 21 | - | 0.12 | [69] |
| Leaves | ≈0.98 MCt FW | - | 0.1 | ||||
| Leaves | 1.55–6.59 MCt FW | 0.001/0.0005 | 1.55–0.32 | ||||
| Fruit | 1.04 MCt FW | 0–0.052 |
Not determined (ND); Crude extract (CE); Pure toxin (P); Estimated daily intake (EDI); Estimated daily intake for adult (EDIAdu); Estimated daily intake for children (EDIChi); Bioaccumulation factor (BFA); Total MCs (MCt); Fresh weight (FW).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Haida, M.; El Khalloufi, F.; Mugani, R.; Essadki, Y.; Campos, A.; Vasconcelos, V.; Oudra, B. Microcystin Contamination in Irrigation Water and Health Risk. Toxins 2024, 16, 196. https://doi.org/10.3390/toxins16040196
AMA Style
Haida M, El Khalloufi F, Mugani R, Essadki Y, Campos A, Vasconcelos V, Oudra B. Microcystin Contamination in Irrigation Water and Health Risk. Toxins. 2024; 16(4):196. https://doi.org/10.3390/toxins16040196
Chicago/Turabian StyleHaida, Mohammed, Fatima El Khalloufi, Richard Mugani, Yasser Essadki, Alexandre Campos, Vitor Vasconcelos, and Brahim Oudra. 2024. "Microcystin Contamination in Irrigation Water and Health Risk" Toxins 16, no. 4: 196. https://doi.org/10.3390/toxins16040196
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.