Some Examples of Bacterial Toxins as Tools


Abstract
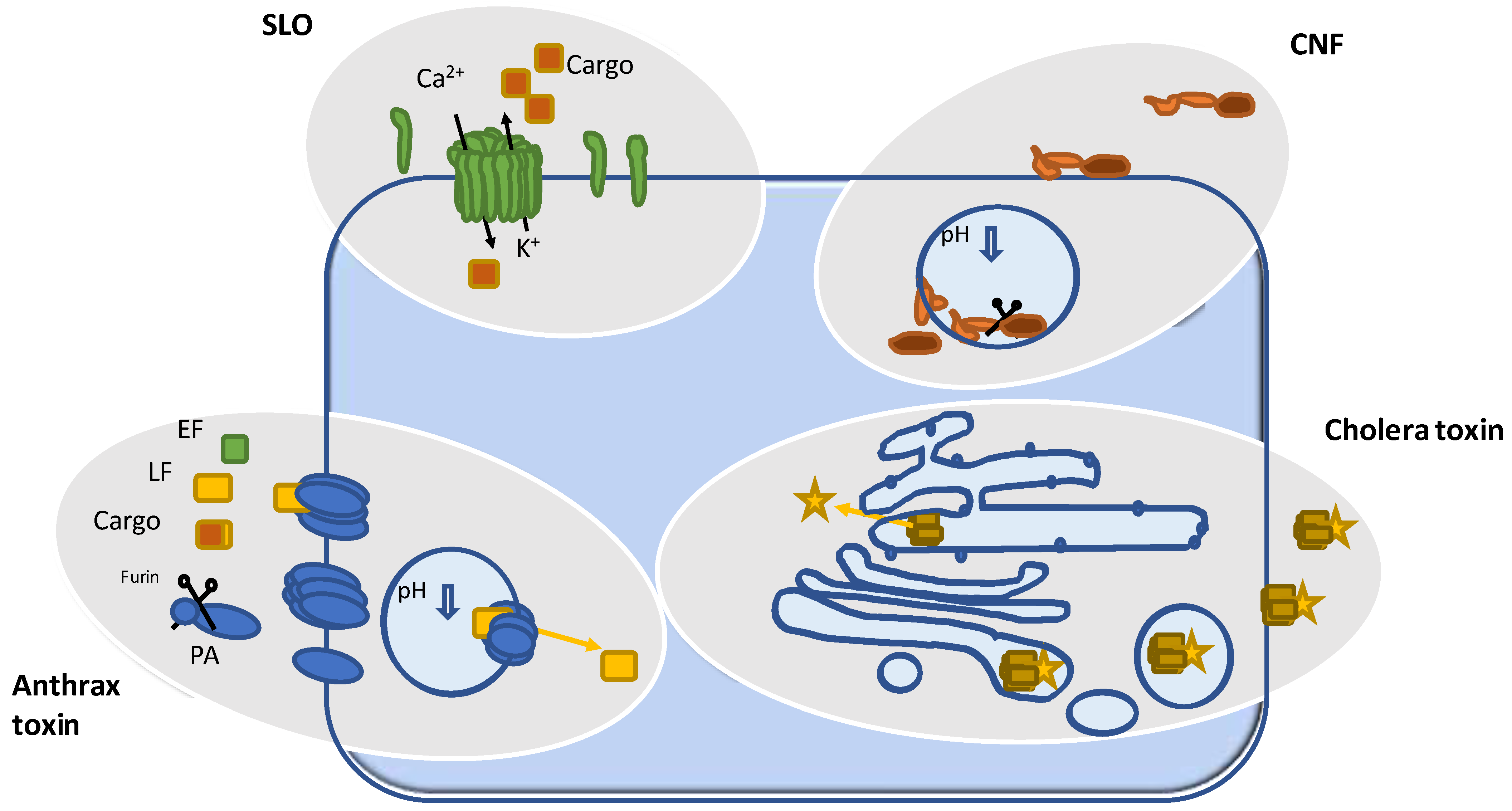
1. Pore Forming Bacterial Toxins
2. Toxins Forming Pores for Delivery of an Enzyme
3. The Benefit of Understanding the Enzymatic Activity of Bacterial Toxins
3.1. ADP Ribosyltransferases
3.2. The Deamidating Toxins CNF, PMT, and DNT
3.3. Toxins with Other Enzymatic Activities
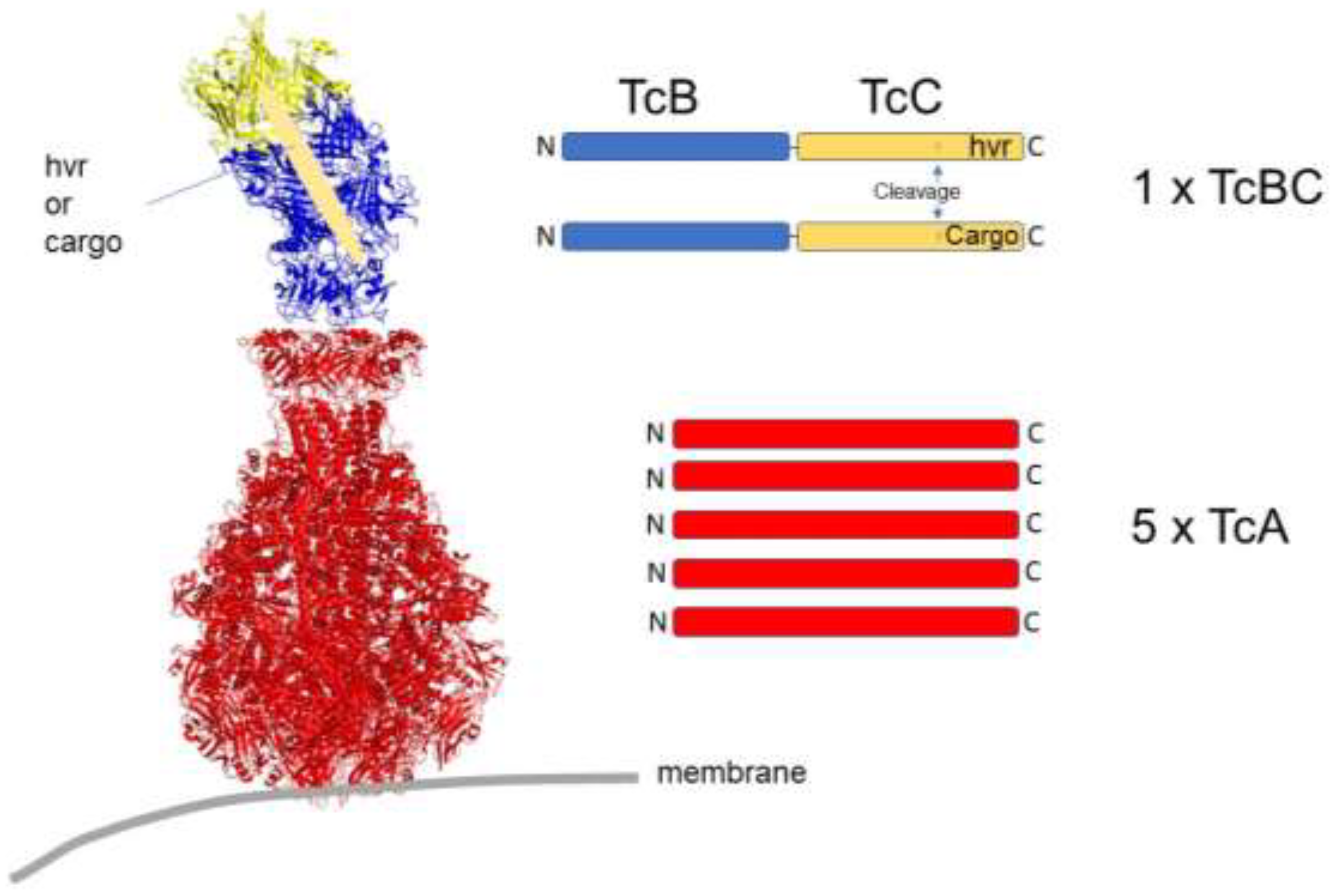
4. Secreted Injection Systems
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhakdi, S.; Valeva, A.; Walev, I.; Weller, U.; Palmer, M. Bacterial Toxins—Tools in Cell Biology and Pharmacology; Aktories, K., Ed.; Chapman & Hall: London, UK, 1997; pp. 241–257. [Google Scholar]
- Peraro, M.D.; van der Goot, F.G. Pore-forming toxins: Ancient, but never really out of fashion. Nat. Rev. Microbiol. 2016, 14, 77–92. [Google Scholar] [CrossRef]
- Madden, J.C.; Ruiz, N.; Caparon, M. Cytolysin-Mediated Translocation (CMT): A Functional Equivalent of Type III Secretion in Gram-Positive Bacteria. Cell 2001, 104, 143–152. [Google Scholar] [CrossRef]
- Gilbert, R.J. Cholesterol-dependent cytolysins. Adv. Exp. Med. Biol. 2010, 677, 56–66. [Google Scholar]
- Feil, S.C.; Ascher, D.B.; Kuiper, M.J.; Tweten, R.K.; Parker, M.W. Structural Studies of Streptococcus pyogenes Streptolysin O Provide Insights into the Early Steps of Membrane Penetration. J. Mol. Biol. 2014, 426, 785–792. [Google Scholar] [CrossRef]
- Ahnert-Hilger, G.; Pahner, I.; Höltje, M. Handbook of Experimental Pharmacology; Aktories, K., Just, I., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 557–575, Ch. 24. [Google Scholar]
- Verma, P.; Gandhi, S.; Lata, K.; Chattopadhyay, K. Pore-forming toxins in infection and immunity. Biochem. Soc. Trans. 2021, 49, 455–465. [Google Scholar] [CrossRef]
- Benz, R.; Hardie, K.R.; Hughes, C. Pore formation in artificial membranes by the secreted hemolysins of Proteus vulgaris and Morganella morganii. Eur. J. Biol. Inorg. Chem. 1994, 220, 339–347. [Google Scholar] [CrossRef]
- Benz, R.; Schmid, A.; E Hancock, R. Ion selectivity of gram-negative bacterial porins. J. Bacteriol. 1985, 162, 722–727. [Google Scholar] [CrossRef]
- Tweten, R.K.; Parker, M.W.; Johnson, A.E. The cholesterol-dependent cytolysins. Curr. Top. Microbiol. Immunol. 2001, 257, 15–33. [Google Scholar]
- Das, A.; Goldstein, J.L.; Anderson, D.D.; Brown, M.S.; Radhakrishnan, A. Use of mutant 125 I-perfringolysin O to probe transport and organization of cholesterol in membranes of animal cells. Proc. Natl. Acad. Sci. USA 2013, 110, 10580–10585. [Google Scholar] [CrossRef]
- Orlandi, P.; Curran, P.; Fishman, P. Brefeldin A blocks the response of cultured cells to cholera toxin. Implications for intracellular trafficking in toxin action. J. Biol. Chem. 1993, 268, 12010–12016. [Google Scholar] [CrossRef]
- Bergan, J.; Lingelem, A.B.D.; Simm, R.; Skotland, T.; Sandvig, K. Shiga toxins. Toxicon 2012, 60, 1085–1107. [Google Scholar] [CrossRef]
- Beauregard, K.E.; Wimer-Mackin, S.; Collier, R.J.; Lencer, W.I. Anthrax toxin entry into polarized epithelial cells. Infect. Immun. 1999, 67, 3026–3030. [Google Scholar] [CrossRef]
- Bradley, K.A.; Mogridge, J.; Mourez, M.; Collier, R.J.; Young, J.A.T. Identification of the cellular receptor for anthrax toxin. Nature 2001, 414, 225–229. [Google Scholar] [CrossRef]
- Escuyer, V.; Collier, R.J. Anthrax protective antigen interacts with a specific receptor on the surface of CHO-K1 cells. Infect. Immun. 1991, 59, 3381–3386. [Google Scholar] [CrossRef]
- Collier, R.J.; Young, J.A. Anthrax toxin. Annu. Rev. Cell Dev. Biol. 2003, 19, 45–70. [Google Scholar] [CrossRef]
- Ezzell, J.W.; Abshire, T.G. Serum protease cleavage of Bacillus anthracis protective antigen. J. Gen. Microbiol. 1992, 138, 543–549. [Google Scholar] [CrossRef]
- Bachran, C.; Leppla, S.H. Tumor Targeting and Drug Delivery by Anthrax Toxin. Toxins 2016, 8, 197. [Google Scholar] [CrossRef]
- Mechaly, A.; McCluskey, A.J.; Collier, R.J. Changing the receptor specificity of anthrax toxin. mBio 2012, 3, 10–128. [Google Scholar] [CrossRef]
- McCluskey, A.J.; Olive, A.J.; Starnbach, M.N.; Collier, R.J. Targeting HER2-positive cancer cells with receptor-redirected anthrax protective antigen. Mol. Oncol. 2013, 7, 440–451. [Google Scholar] [CrossRef]
- Zahaf, N.-I.; Lang, A.E.; Kaiser, L.; Fichter, C.D.; Lassmann, S.; McCluskey, A.; Augspach, A.; Aktories, K.; Schmidt, G. Targeted delivery of an ADP-ribosylating bacterial toxin into cancer cells. Sci. Rep. 2017, 7, 41252. [Google Scholar] [CrossRef]
- Abrami, L.; Bischofberger, M.; Kunz, B.; Groux, R.; van der Goot, F.G. Endocytosis of the anthrax toxin is mediated by clathrin, actin and unconventional adaptors. PLoS Pathog. 2010, 6, e1000792. [Google Scholar] [CrossRef]
- Liu, W.; Nestorovich, E.M. Anthrax toxin channel: What we know based on over 30 years of research. Biochim. Biophys. Acta (BBA) Biomembr. 2021, 1863, 183715. [Google Scholar] [CrossRef]
- Collier, R.J. Membrane translocation by anthrax toxin. Mol. Asp. Med. 2009, 30, 413–422. [Google Scholar] [CrossRef]
- Friebe, S.; Van der Goot, F.G.; Bürgi, J. The Ins and Outs of Anthrax Toxin. Toxins 2016, 8, 69. [Google Scholar] [CrossRef]
- Eckhardt, M.; Barth, H.; Blöcker, D.; Aktories, K. Binding of Clostridium botulinum C2 toxin to asparagine-linked complex and hybrid carbohydrates. J. Biol. Chem. 2000, 275, 2328–2334. [Google Scholar] [CrossRef]
- Barth, H.; Blöcker, D.; Behlke, J.; Bergsma-Schutter, W.; Brisson, A.; Benz, R.; Aktories, K. Cellular uptake of Clostridium botulinum C2 toxin requires oligomerization and acidification. J. Biol. Chem. 2000, 275, 18704–18711. [Google Scholar] [CrossRef]
- Heber, S.; Borho, J.; Stadler, N.; Wondany, F.; König, I.; Michaelis, J.; Papatheodorou, P.; Barth, H.; Fellermann, M. The Clostridium botulinum C2 Toxin Subunit C2IIa Delivers Enzymes with Positively Charged N-Termini into the Cytosol of Target Cells. Toxins 2023, 15, 390. [Google Scholar] [CrossRef]
- Barth, H.; Stiles, B.G. Binary actin-ADP-ribosylating toxins and their use as molecular trojan horses for drug delivery into eukaryotic cells. Curr. Med. Chem. 2008, 15, 459–469. [Google Scholar] [CrossRef]
- Voskoboinik, I.; Whisstock, J.C.; Trapani, J.A. Perforin and granzymes: Function, dysfunction and human pathology. Nat. Rev. Immunol. 2015, 15, 388–400. [Google Scholar] [CrossRef]
- Olsnes, S.; Klingenberg, O.; Munoz, R.; Falnes, P.O.; Wiedlocha, A. Bacterial Toxins: Tools in Cell Biology and Pharmacology; Aktories, K., Ed.; Chapman & Hall: London, UK, 1997; pp. 273–294. [Google Scholar]
- Auger, A.; Park, M.; Nitschke, F.; Minassian, L.M.; Beilhartz, G.L.; Minassian, B.A.; Melnyk, R.A. Efficient Delivery of Structurally Diverse Protein Cargo into Mammalian Cells by a Bacterial Toxin. Mol. Pharm. 2015, 12, 2962–2971. [Google Scholar] [CrossRef]
- Aktories, K.; Jakobs, K.H.; Robbel, L. Further contributions to an improved pertussis vaccine: I. Molecular mechanisms of pertussis toxin. Behring Inst. Res. Commun. 1984, 76, 106–109. [Google Scholar]
- Merritt, E.A.; Hol WG, J. AB5 toxins. Curr. Opin. Struct. Biol. 1995, 5, 165–171. [Google Scholar] [CrossRef]
- Gierschik, P.; Jakobs, K.H. Handbook of Experimental Pharmacology; Herken, H., Hucho, F., Eds.; Springer: Berlin/Heidelberg, Germany, 1992; Volume 102, pp. 807–839, Ch. 23. [Google Scholar]
- Aktories, K.; Braun, U.; Rösener, S.; Just, I.; Hall, A. The rho gene product expressed in E. Coli is a substrate of botulinum ADP-ribosyltransferase C3. Biochem. Biophys. Res. Commun. 1989, 158, 209–213. [Google Scholar] [CrossRef]
- Auer, M.; Schweigreiter, R.; Hausott, B.; Thongrong, S.; Höltje, M.; Just, I.; Bandtlow, C.; Klimaschewski, L. Rho-independent stimulation of axon outgrowth and activation of the ERK and Akt signaling pathways by C3 transferase in sensory neurons. Front. Cell. Neurosci. 2012, 6, 43. [Google Scholar] [CrossRef]
- Jeon, C.; Moon, M.; Kim, J.; Kim, H.; Kim, J.; Li, Y.; Jin, J.; Kim, P.; Kim, H.; Meier, K.E.; et al. Control of neurite outgrowth by RhoA inactivation. J. Neurochem. 2012, 120, 684–698. [Google Scholar] [CrossRef]
- Höltje, M.; Djalali, S.; Hofmann, F.; Münster-Wandowski, A.; Hendrix, S.; Boato, F.; Dreger, S.C.; Große, G.; Henneberger, C.; Grantyn, R.; et al. A 29-amino acid fragment of Clostridium botulinum C3 protein enhances neuronal outgrowth, connectivity, and reinnervation. FASEB J. 2008, 23, 1115–1126. [Google Scholar] [CrossRef]
- Rohrbeck, A.; Schröder, A.; Hagemann, S.; Pich, A.; Höltje, M.; Ahnert-Hilger, G.; Just, I. Vimentin mediates uptake of C3 exoenzyme. PLoS ONE 2014, 9, e101071. [Google Scholar] [CrossRef]
- Chung, D.W.; Collier, R. The mechanism of ADP-ribosylation of elongation factor 2 catalyzed by fragment a from diphtheria toxin. Biochim. Biophys. Acta (BBA) Enzym. 1977, 483, 248–257. [Google Scholar] [CrossRef]
- Teruya, S.; Hiramatsu, Y.; Nakamura, K.; Fukui-Miyazaki, A.; Tsukamoto, K.; Shinoda, N.; Motooka, D.; Nakamura, S.; Ishigaki, K.; Shinzawa, N.; et al. Bordetella Dermonecrotic Toxin Is a Neurotropic Virulence Factor That Uses CaV3.1 as the Cell Surface Receptor. mBio 2020, 11, e03146-19. [Google Scholar] [CrossRef]
- Schoellkopf, J.; Mueller, T.; Hippchen, L.; Mueller, T.; Reuten, R.; Backofen, R.; Orth, J.; Schmidt, G. Genome wide CRISPR screen for Pasteurella multocida toxin (PMT) binding proteins reveals LDL Receptor Related Protein 1 (LRP1) as crucial cellular receptor. PLoS Pathog. 2022, 18, e1010781. [Google Scholar] [CrossRef]
- Xu, Y.; Viswanatha, R.; Sitsel, O.; Roderer, D.; Zhao, H.; Ashwood, C.; Voelcker, C.; Tian, S.; Raunser, S.; Perrimon, N.; et al. CRISPR screens in Drosophila cells identify Vsg as a Tc toxin receptor. Nature 2022, 610, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, A.C.; Szwed, E.; Spears, C.D.; Thaper, S.; Dang, L.H.; Dang, N.H. Denileukin Diftitox (Ontak) as Maintenance Therapy for Peripheral T-Cell Lymphomas: Three Cases with Sustained Remission. Case Rep. Oncol. Med. 2015, 2015, 123756. [Google Scholar] [CrossRef] [PubMed]
- Beziat, G.; Ysebaert, L. Tagraxofusp for the Treatment of Blastic Plasmacytoid Dendritic Cell Neoplasm (BPDCN): A Brief Report on Emerging Data. OncoTargets Ther. 2020, 13, 5199–5205. [Google Scholar] [CrossRef]
- FitzGerald, D.J. Targeted diphtheria toxin to treat BPDCN. Blood 2014, 124, 310–312. [Google Scholar] [CrossRef]
- Zhang, T.; Dong, S.; Zhai, Y.; Naatz, L.; Zhou, Z.; Chen, M. Diphtheria toxin-derived, anti-PD-1 immunotoxin, a potent and practical tool to selectively deplete PD-1+ cells. Protein Sci. 2023, 32, e4741. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Wang, P.; Dong, S.; Zhou, Z.; Cao, Y.; Yagita, H.; He, X.; Zheng, S.G.; Fisher, S.J.; Fujinami, R.S.; et al. Depletion of PD-1-positive cells ameliorates autoimmune disease. Nat. Biomed. Eng. 2019, 3, 292–305. [Google Scholar] [CrossRef] [PubMed]
- Iglewski, B.H.; Liu, P.V.; Kabat, D. Mechanism of action of Pseudomonas aeruginosa exotoxin Aiadenosine diphosphate-ribosylation of mammalian elongation factor 2 in vitro and in vivo. Infect. Immun. 1977, 15, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, C.; Schmidt, G. CNF and DNT. Rev. Physiol. Biochem. Pharmacol. 2004, 152, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, G.; Sehr, P.; Wilm, M.; Selzer, J.; Mann, M.; Aktories, K. Gln 63 of Rho is deamidated by Escherichia coli cytotoxic necrotizing factor-1. Nature 1997, 387, 725–729. [Google Scholar] [CrossRef]
- Flatau, G.; Lemichez, E.; Gauthier, M.; Chardin, P.; Paris, S.; Fiorentini, C.; Boquet, P. Toxin-induced activation of the G protein p21 Rho by deamidation of glutamine. Nature 1997, 387, 729–733. [Google Scholar] [CrossRef]
- Wilson, B.A.; Ho, M. Pasteurella multocida toxin as a tool for studying G(q) signal transduction. Rev. Physiol Biochem. Pharmacol. 2004, 152, 93–109. [Google Scholar] [PubMed]
- Kubatzky, K.F. Pasteurella multocida toxin—lessons learned from a mitogenic toxin. Front. Immunol. 2022, 13, 1058905. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.A.; Ho, M. Pasteurella multocida Toxin Interaction with Host Cells: Entry and Cellular Effects. Curr. Top. Microbiol. Immunol. 2012, 361, 93–111. [Google Scholar] [CrossRef]
- Brothers, M.C.; Ho, M.; Maharjan, R.; Clemons, N.C.; Bannai, Y.; Waites, M.A.; Faulkner, M.J.; Kuhlenschmidt, T.B.; Kuhlenschmidt, M.S.; Blanke, S.R.; et al. Membrane interaction of Pasteurella multocida toxin involves sphingomyelin. FEBS J. 2011, 278, 4633–4648. [Google Scholar] [CrossRef] [PubMed]
- Kowarschik, S.; Schöllkopf, J.; Müller, T.; Tian, S.; Knerr, J.; Bakker, H.; Rein, S.; Dong, M.; Weber, S.; Grosse, R.; et al. Yersinia pseudotuberculosis cytotoxic necrotizing factor interacts with glycosaminoglycans. FASEB J. 2021, 35, e21647. [Google Scholar] [CrossRef]
- Umata, T.; Moriyama, Y.; Futai, M.; Mekada, E. The cytotoxic action of diphtheria toxin and its degradation in intact Vero cells are inhibited by bafilomycin A1, a specific inhibitor of vacuolar-type H(+)-ATPase. J. Biol. Chem. 1990, 265, 21940–21945. [Google Scholar] [CrossRef] [PubMed]
- Knust, Z.; Blumenthal, B.; Aktories, K.; Schmidt, G. Cleavage of Escherichia coli cytotoxic necrotizing factor 1 is required for full biologic activity. Infect. Immun. 2009, 77, 1835–1841. [Google Scholar] [CrossRef]
- Bergmann, S.; Jehle, D.; Schwan, C.; Orth, J.H.C.; Aktories, K. Pasteurella multocida Toxin as a Transporter of Non-Cell-Permeating Proteins. Infect. Immun. 2013, 81, 2459–2467. [Google Scholar] [CrossRef]
- Mohamadzadeh, M. Microbial Toxins: Current Research and Future Trends. Expert Rev. Anti-Infect. Ther. 2009, 7, 695–696. [Google Scholar] [CrossRef][Green Version]
- Hildebrand, D.; Walker, P.; Dalpke, A.; Heeg, K.; Kubatzky, K.F. Pasteurella multocida Toxin-induced Pim-1 expression disrupts suppressor of cytokine signalling (SOCS)-1 activity. Cell. Microbiol. 2010, 12, 1732–1745. [Google Scholar] [CrossRef]
- Verma, A.K.; Aga, P.; Singh, S.K.; Singh, R. The stone man disease: Fibrodysplasia ossificans progressiva: Imaging revisited. BMJ Case Rep. 2012, 2012, bcr2012006422. [Google Scholar] [CrossRef] [PubMed]
- Strack, J.; Heni, H.; Gilsbach, R.; Hein, L.; Aktories, K.; Orth, J.H.C. Noncanonical G-protein-dependent modulation of osteoclast differentiation and bone resorption mediated by Pasteurella multocida toxin. mBio 2014, 5, e02190-14. [Google Scholar] [CrossRef] [PubMed]
- Boquet, P. The cytotoxic necrotizing factor 1 (CNF1) from Escherichia coli. Toxicon 2001, 39, 1673–1680. [Google Scholar] [CrossRef] [PubMed]
- Colarusso, A.; Maroccia, Z.; Parrilli, E.; Germinario, E.A.P.; Fortuna, A.; Loizzo, S.; Ricceri, L.; Tutino, M.L.; Fiorentini, C.; Fabbri, A. Cnf1 Variants Endowed with the Ability to Cross the Blood–Brain Barrier: A New Potential Therapeutic Strategy for Glioblastoma. Toxins 2020, 12, 291. [Google Scholar] [CrossRef] [PubMed]
- Pavone, F.; Luvisetto, S.; Marinelli, S.; Straface, E.; Fabbri, A.; Falzano, L.; Fiorentini, C.; Malorni, W. The Rac GTPase-activating bacterial protein toxin CNF1 induces analgesia up-regulating μ-opioid receptors. Pain 2009, 145, 219–229. [Google Scholar] [CrossRef]
- Vannini, E.; Mori, E.; Tantillo, E.; Schmidt, G.; Caleo, M.; Costa, M. CTX-CNF1 Recombinant Protein Selectively Targets Glioma Cells in Vivo. Toxins 2021, 13, 194. [Google Scholar] [CrossRef]
- Diana, G.; Valentini, G.; Travaglione, S.; Falzano, L.; Pieri, M.; Zona, C.; Meschini, S.; Fabbri, A.; Fiorentini, C. Enhancement of learning and memory after activation of cerebral Rho GTPases. Proc. Natl. Acad. Sci. USA 2007, 104, 636–641. [Google Scholar] [CrossRef]
- Malchiodi-Albedi, F.; Paradisi, S.; Di Nottia, M.; Simone, D.; Travaglione, S.; Falzano, L.; Guidotti, M.; Frank, C.; Cutarelli, A.; Fabbri, A.; et al. CNF1 Improves astrocytic ability to support neuronal growth and differentiation in vitro. PLoS ONE 2012, 7, e34115. [Google Scholar] [CrossRef]
- Loizzo, S.; Rimondini, R.; Travaglione, S.; Fabbri, A.; Guidotti, M.; Ferri, A.; Campana, G.; Fiorentini, C. CNF1 Increases brain energy level, counteracts neuroinflammatory markers and rescues cognitive deficits in a murine model of alzheimer’s disease. PLoS ONE 2013, 8, e65898. [Google Scholar] [CrossRef]
- Vannini, E.; Olimpico, F.; Middei, S.; Ammassari-Teule, M.; de Graaf, E.L.; McDonnell, L.; Schmidt, G.; Fabbri, A.; Fiorentini, C.; Baroncelli, L.; et al. Electrophysiology of glioma: A Rho GTPase-activating protein reduces tumor growth and spares neuron structure and function. Neuro-Oncology 2016, 18, 1634–1643. [Google Scholar] [CrossRef]
- Urbinati, C.; Cosentino, L.; Germinario, E.A.P.; Valenti, D.; Vigli, D.; Ricceri, L.; Laviola, G.; Fiorentini, C.; Vacca, R.A.; Fabbri, A.; et al. Treatment with the Bacterial Toxin CNF1 Selectively Rescues Cognitive and Brain Mitochondrial Deficits in a Female Mouse Model of Rett Syndrome Carrying a MeCP2-Null Mutation. Int. J. Mol. Sci. 2021, 22, 6739. [Google Scholar] [CrossRef] [PubMed]
- Montecucco, C.; Schiavo, G. Tetanus and botulism neurotoxins: A new group of zinc proteases. Trends Biochem. Sci. 1993, 18, 324–327. [Google Scholar] [CrossRef]
- Schiavo, G.G.; Benfenati, F.; Poulain, B.; Rossetto, O.; de Laureto, P.P.; DasGupta, B.R.; Montecucco, C. Tetanus and botulinum-B neurotoxins block neurotransmitter release by proteolytic cleavage of synaptobrevin. Nature 1992, 359, 832–835. [Google Scholar] [CrossRef]
- Montecucco, C.; Papini, E.; Schiavo, G. Bacterial protein toxins penetrate cells via a four-step mechanism. FEBS Lett. 1994, 346, 92–98. [Google Scholar] [CrossRef]
- Montecucco, C.; Schiavo, G. Structure and function of tetanus and botulinum neurotoxins. Q. Rev. Biophys. 1995, 28, 423–472. [Google Scholar] [CrossRef]
- Montecucco, C. Protein toxins and membrane transport. Curr. Opin. Cell Biol. 1998, 10, 530–536. [Google Scholar] [CrossRef] [PubMed]
- McNew, J.A.; Parlati, F.; Fukuda, R.; Johnston, R.J.; Paz, K.; Paumet, F.; Söllner, T.H.; Rothman, J.E. Compartmental specificity of cellular membrane fusion encoded in SNARE proteins. Nature 2000, 407, 153–159. [Google Scholar] [CrossRef]
- Schiavo, G.; Matteoli, M.; Montecucco, C.; Kasai, H.; Takahashi, N.; Tokumaru, H.; Martin, J.H.; Engber, D.; Meng, Z.; Banerjee, A.; et al. Neurotoxins affecting neuroexocytosis. Physiol. Rev. 2000, 80, 717–766. [Google Scholar] [CrossRef]
- Carruthers, A.; Bruce, S.; Cox, S.E.; Kane, M.A.; Lee, E.; Gallagher, C.J. OnabotulinumtoxinA for Treatment of Moderate to Severe Crow’s Feet Lines: A Review. Aesthetic Surg. J. 2016, 36, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Benz, R.; Maier, E.; Ladant, D.; Ullmann, A.; Sebo, P. Adenylate cyclase toxin (CyaA) of Bordetella pertussis. Evidence for the formation of small ion-permeable channels and comparison with HlyA of Escherichia coli. J. Biol. Chem. 1994, 269, 27231–27239. [Google Scholar] [CrossRef] [PubMed]
- Vojtova-Vodolanova, J.; Basler, M.; Osicka, R.; Knapp, O.; Maier, E.; Cerny, J.; Benada, O.; Benz, R.; Sebo, P. Oligomerization is involved in pore formation by Bordetella adenylate cyclase toxin. FASEB J. 2009, 23, 2831–2843. [Google Scholar] [CrossRef] [PubMed]
- Veneziano, R.; Rossi, C.; Chenal, A.; Devoisselle, J.-M.; Ladant, D.; Chopineau, J. Bordetella pertussis adenylate cyclase toxin translocation across a tethered lipid bilayer. Proc. Natl. Acad. Sci. USA 2013, 110, 20473–20478. [Google Scholar] [CrossRef] [PubMed]
- Bumba, L.; Masin, J.; Fiser, R.; Sebo, P. Bordetella Adenylate Cyclase Toxin Mobilizes Its β2 Integrin Receptor into Lipid Rafts to Accomplish Translocation across Target Cell Membrane in Two Steps. PLoS Pathog. 2010, 6, e1000901. [Google Scholar] [CrossRef] [PubMed]
- Sebo, P.; Osicka, R.; Masin, J. Adenylate cyclase toxin-hemolysin relevance for pertussis vaccines. Expert Rev. Vaccines 2014, 13, 1215–1227. [Google Scholar] [CrossRef] [PubMed]
- Svedova, M.; Masin, J.; Fiser, R.; Cerny, O.; Tomala, J.; Freudenberg, M.; Tuckova, L.; Kovar, M.; Dadaglio, G.; Adkins, I.; et al. Pore-formation by adenylate cyclase toxoid activates dendritic cells to prime CD8+ and CD4+ T cells. Immunol. Cell Biol. 2016, 94, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, G.R. Type III secretion: A bacterial device for close combat with cells of their eukaryotic host. Philos. Trans. R. Soc. Lond. 2000, 355, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Daborn, P.J.; Waterfield, N.; Blight, M.A.; Ffrench-Constant, R.H. measuring virulence factor expression by the pathogenic bacterium Photorhabdus luminescens in culture and during insect infection. J. Bacteriol. 2001, 183, 5834–5839. [Google Scholar] [CrossRef] [PubMed]
- Meusch, D.; Gatsogiannis, C.; Efremov, R.G.; Lang, A.E.; Hofnagel, O.; Vetter, I.R.; Aktories, K.; Raunser, S. Mechanism of Tc toxin action revealed in molecular detail. Nature 2014, 508, 61–65. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.; Waterfield, M.D. DNA Sequences from TCD Genomic Region of Photorhabdus luminescens. U.S. Patent US 7,777,100 B2, 17 August 2010. [Google Scholar]
- Gatsogiannis, C.; Lang, A.E.; Meusch, D.; Pfaumann, V.; Hofnagel, O.; Benz, R.; Aktories, K.; Raunser, S. A syringe-like injection mechanism in Photorhabdus luminescens toxins. Nature 2013, 495, 520–523. [Google Scholar] [CrossRef]
- Ost, G.S.; Ng’Ang’A, P.N.; Lang, A.E.; Aktories, K. Photorhabdus luminescens Tc toxin is inhibited by the protease inhibitor MG132 and activated by protease cleavage resulting in increased binding to target cells. Cell. Microbiol. 2019, 21, e12978. [Google Scholar] [CrossRef]
- Gatsogiannis, C.; Merino, F.; Prumbaum, D.; Roderer, D.; Leidreiter, F.; Meusch, D.; Raunser, S. Membrane insertion of a Tc toxin in near-atomic detail. Nat. Struct. Mol. Biol. 2016, 23, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Ng’ang´a, P.N.; Ebner, J.K.; Plessner, M.; Aktories, K.; Schmidt, G. Engineering Photorhabdus luminescens toxin complex (PTC) into a recombinant injection nanomachine. Life Sci. Alliance 2019, 2, e201900485. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Li, N.; Wang, X.; Cheng, J.; Huang, Y.; Yang, Y.; Yang, J.; Cai, B.; Wang, Y.-P.; Jin, Q.; et al. Cryo-EM Structure and Assembly of an Extracellular Contractile Injection System. Cell 2019, 177, 370–383.e15. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Shen, J.; Cheng, J.; Wang, X.; Yang, J.; Li, N.; Gao, N.; Jin, Q. N-terminal signal peptides facilitate the engineering of PVC complex as a potent protein delivery system. Sci. Adv. 2022, 8, eabm2343. [Google Scholar] [CrossRef] [PubMed]
- Gerhard, R.; Tatge, H.; Genth, H.; Thum, T.; Borlak, J.; Fritz, G.; Just, I. Clostridium difficile toxin a induces expression of the stress-induced early gene product RhoB. J. Biol. Chem. 2005, 280, 1499–1505. [Google Scholar] [CrossRef] [PubMed]
- Barth, H.; Olenik, C.; Sehr, P.; Schmidt, G.; Aktories, K.; Meyer, D.K. Neosynthesis and activation of Rho by escherichia coli cytotoxic necrotizing factor (CNF1) reverse cytopathic effects of ADP-ribosylated Rho. J. Biol. Chem. 1999, 274, 27407–27414. [Google Scholar] [CrossRef]
- Pop, M.; Aktories, K.; Schmidt, G. Isotype-specific degradation of Rac activated by the cytotoxic necrotizing factor 1 (CNF1). J. Biol. Chem. 2004, 279, 35840–35848. [Google Scholar] [CrossRef]
- Bellows, S.; Jankovic, J. Immunogenicity Associated with Botulinum Toxin Treatment. Toxins 2019, 11, 491. [Google Scholar] [CrossRef]
- Carr, W.W.; Jain, N.; Sublett, J.W. Immunogenicity of Botulinum Toxin Formulations: Potential Therapeutic Implications. Adv. Ther. 2021, 38, 5046–5064. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Pore-forming toxins (selection) | ||||
| Toxin | Bacterium | Class | Receptor | |
| Aerolysin | Aeromonas spp. | β | CD52 | |
| Cytolysin A | Escherichia coli, Salmonella enterica | α | Cholesterol | |
| ε-Toxin | Clostridium perfringens | β | HAVCR1 | |
| Hemolysin α/γ | Staphylococcus aureus | β | Phosphatidylcholin/ADAM10 | |
| Hemolysin BL | Bacillus cereus | α | Cholesterol | |
| Listeriolysin | Listeria monocytogenes | β | Cholesterol | |
| Perfringolysin O | Clostridium perfringens | β | Cholesterol | |
| Streptolysin O | Streptococcus pyogenes | β | Cholesterol | |
| Toxins with enzymatic activity (selection) | ||||
| Toxin | Bacterium | Enzymatic Activity | Substrate | Protein Receptor |
| Anthrax toxin (EF) | Bacillus anthracis | Adenylylcyclase | ATP | TM8, CMG2 |
| Anthrax toxin (LF) | Bacillus anthracis | Protease | MAPKK, Nlrp1 | TM8, CMG2 |
| Botulinum neurotoxin | Clostridium botulinum | Protease | SNARE proteins | Synaptotagmin |
| C2 toxin | Clostridium botulinum | ADP-Ribosyltransferase | Actin | |
| C3 toxin | Clostridium botulinum | ADP-Ribosyltransferase | RhoA | Vimentin |
| Cholera toxin | Vibrio cholerae | ADP-Ribosyltransferase | Gα s | |
| Cytotoxic necrotizing factors 1,2,3 | Escherichia coli | Deamidase | Rho GTPases | |
| Cytotoxic necrotizing factor Y | Yersinia pseudotuberculosis | Deamidase | Rho GTPases | |
| CyaA | Bordetella pertussis | Adenylylcyclase | ATP | CD11b/CD18 |
| Dermonecrotic toxin | Bordetella bronchiseptica | Transglutaminase | Rho GTPases | Cav3.1 |
| Diphtheria toxin | Corynebacterium diphtheriae | ADP-Ribosyltransferase | Elongation-factor | |
| Pertussis toxin | Bordetella pertussis | ADP-Ribosyltransferase | Gα i | |
| Pasteurella multocida toxin | Pasteurella multocida | Deamidase | Gα subunits | LRP1 |
| Shiga toxin | Shigella dysenteriae Escherichia coli | Depurinase | RNA | |
| Toxin A | Clostridoides difficile | Glucosyltransferase | Rho GTPases | LRP1 |
| Toxin B | Clostridoides difficile | Glucosyltransferase | Rho GTPases | LRP1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmidt, G. Some Examples of Bacterial Toxins as Tools. Toxins 2024, 16, 202. https://doi.org/10.3390/toxins16050202
Schmidt G. Some Examples of Bacterial Toxins as Tools. Toxins. 2024; 16(5):202. https://doi.org/10.3390/toxins16050202
Chicago/Turabian StyleSchmidt, Gudula. 2024. "Some Examples of Bacterial Toxins as Tools" Toxins 16, no. 5: 202. https://doi.org/10.3390/toxins16050202
APA StyleSchmidt, G. (2024). Some Examples of Bacterial Toxins as Tools. Toxins, 16(5), 202. https://doi.org/10.3390/toxins16050202