The Dynamics of Microcystis Genotypes and Microcystin Production and Associations with Environmental Factors during Blooms in Lake Chaohu, China


Abstract
:1. Introduction
2. Results
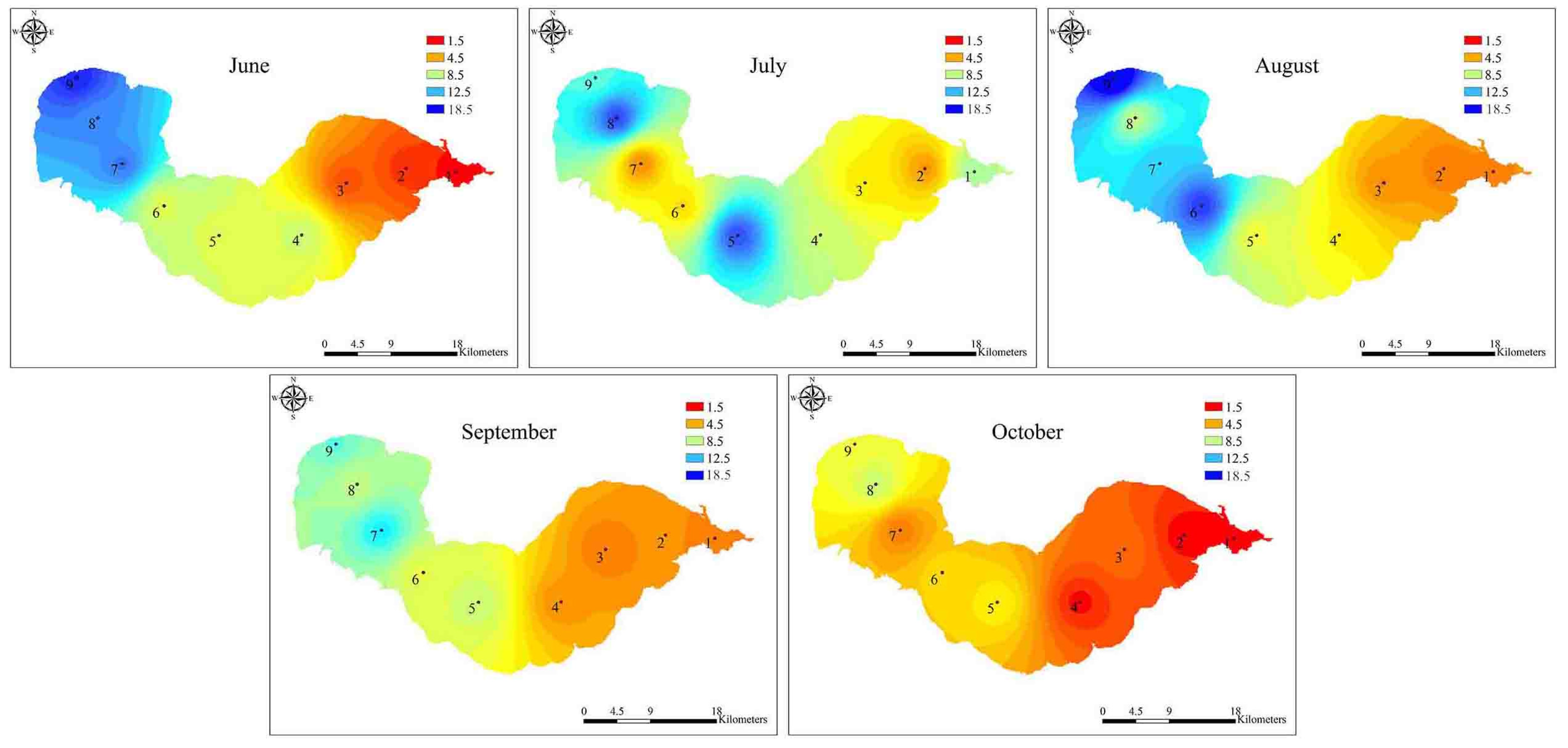
2.1. Variations in Environmental Factors

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Eastern Lake | Central Lake | Western Lake |
|---|---|---|---|
| TN (mg L−1) | 0.92 (0.56–1.48) | 1.44 (0.70–0.25) | 2.84 (1.45–5.09) |
| NH4+–N (mg L−1) | 0.15 (0.04–0.42) | 0.17 (0.04–0.30) | 0.56 (0.09–3.55) |
| NO3−–N (mg L−1) | 0.12 (0.07–0.25) | 0.13 (0.06–0.29) | 0.36 (0.08–0.70) |
| NO2−–N (mg L−1) | 0.01 (0.00–0.02) | 0.01 (0.00–0.06) | 0.08 (0.00–0.37) |
| TP (mg L−1) | 0.05 (0.03–0.09) | 0.11 (0.03–0.23) | 0.20 (0.07–0.39) |
| PO43−–P (mg L−1) | 0.00 (0.00–0.01) | 0.02 (0.00–0.05) | 0.06 (0.01–0.26) |
| DIC (mg L−1) | 14.55 (13.40–16.22) | 13.92 (10.59–17.71) | 12.60 (8.74–15.06) |
| DOC (mg L−1) | 4.64 (3.40–5.67) | 4.78 (4.01–5.52) | 5.40 (4.26–7.46) |
| Chl-a (μg L−1) | 14.42 (5.70–62.28) | 30.19 (7.80–65.43) | 37.56 (10.72–89.40) |
| Water temperature (°C) | 25.74 (19.18–30.75) | 25.30 (18.44–30.41) | 24.98 (18.43–30.04) |
| pH | 7.41 (4.73–9.12) | 7.76 (5.51–9.36) | 7.83 (6.12–9.40) |
| DO (mg L−1) | 9.37 (4.47–14.14) | 8.64 (4.79–10.35) | 8.34 (4.59–14.97) |
| Secchi depth (m) | 52.33 (28.00–85.00) | 35.73 (17.00–90.00) | 28.54 (15.00–45.00) |
| Water depth (m) | 3.70 (2.30–4.50) | 3.81 (1.90–4.50) | 3.21 (1.90–4.00) |
| Conductivity (μS cm−1) | 315.07 (280.00–370.00) | 289.07 (213.00–361.00) | 315.10 (223.00–392.00) |

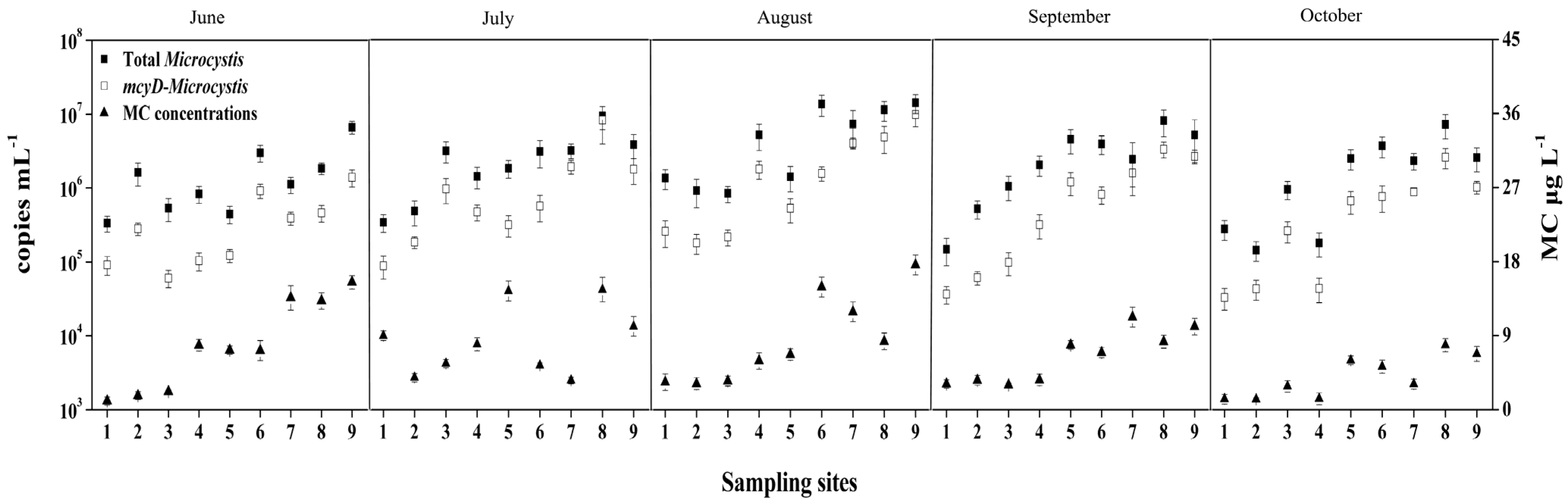
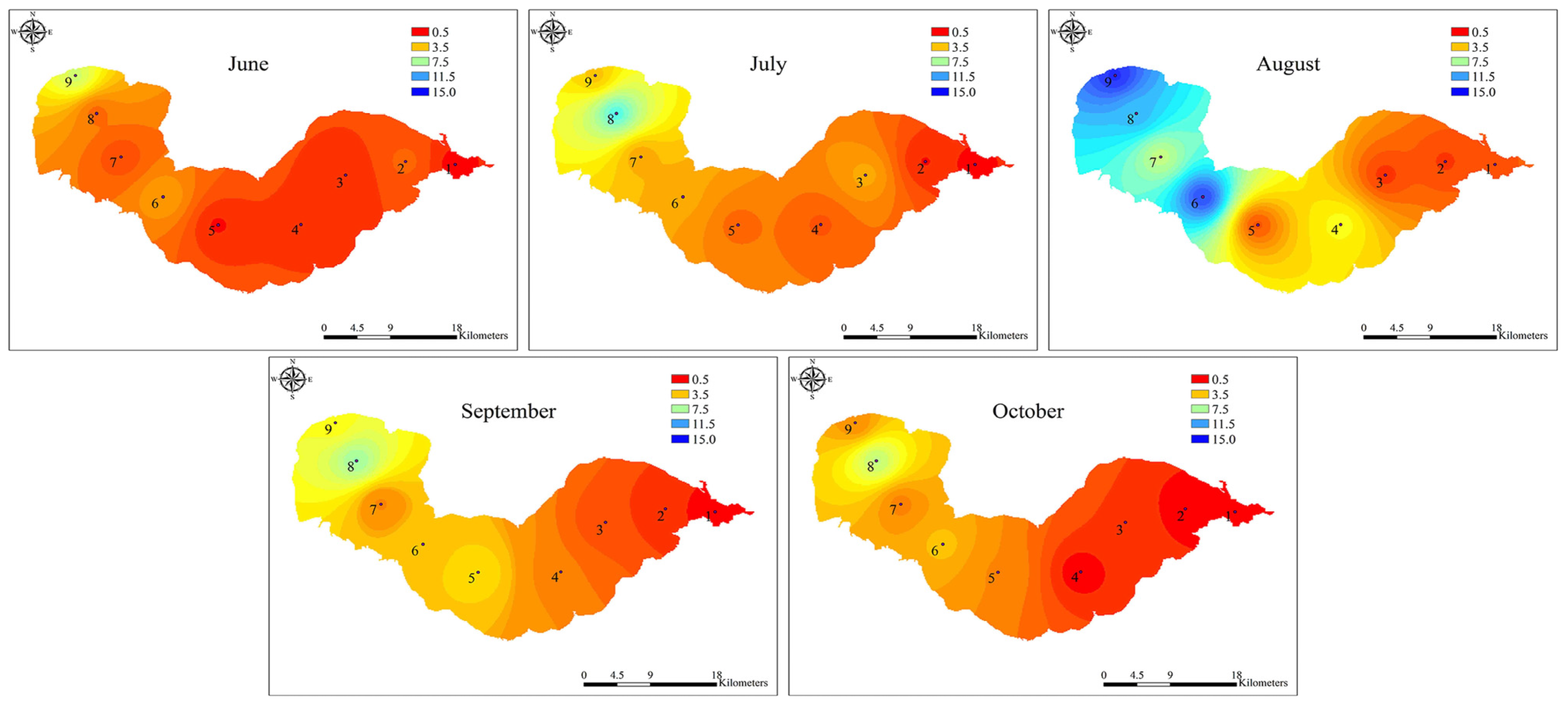
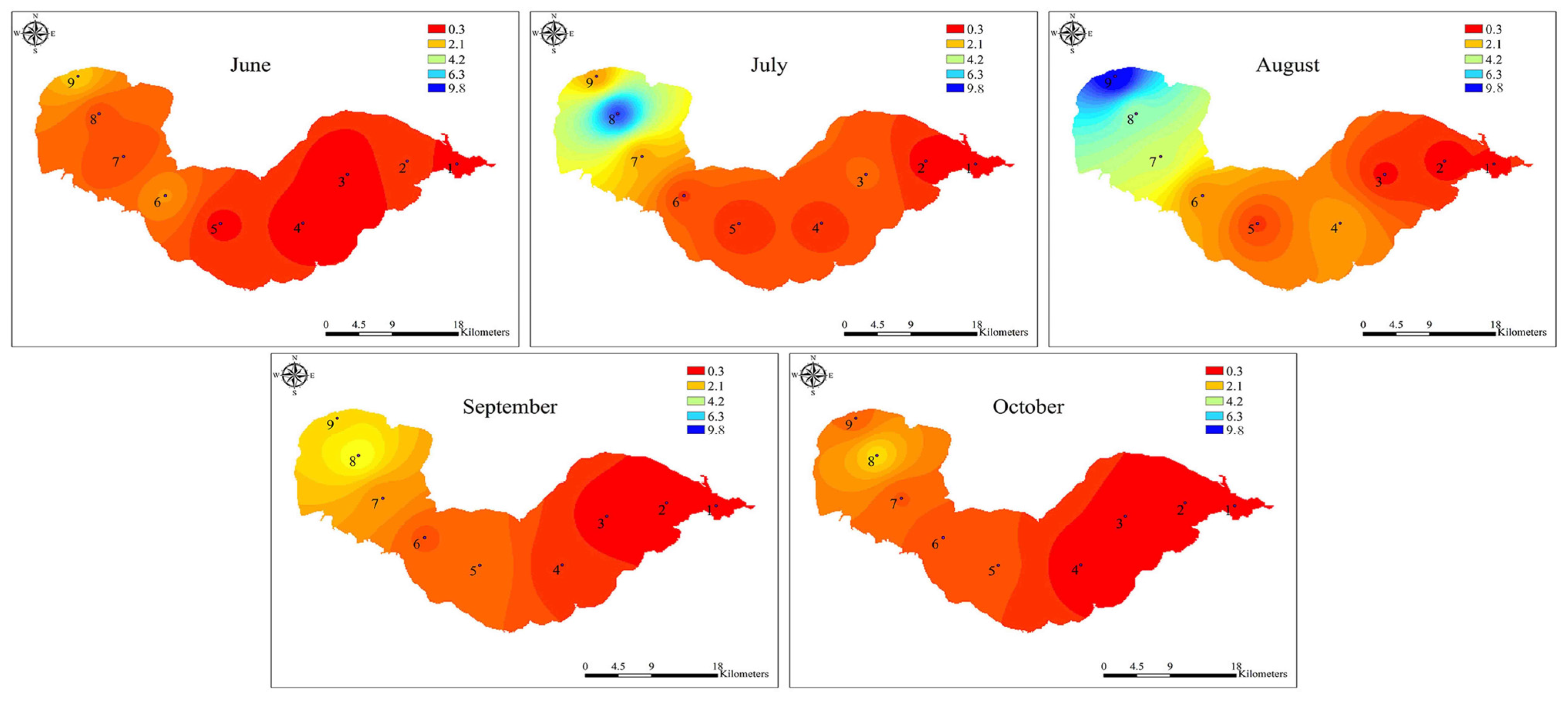
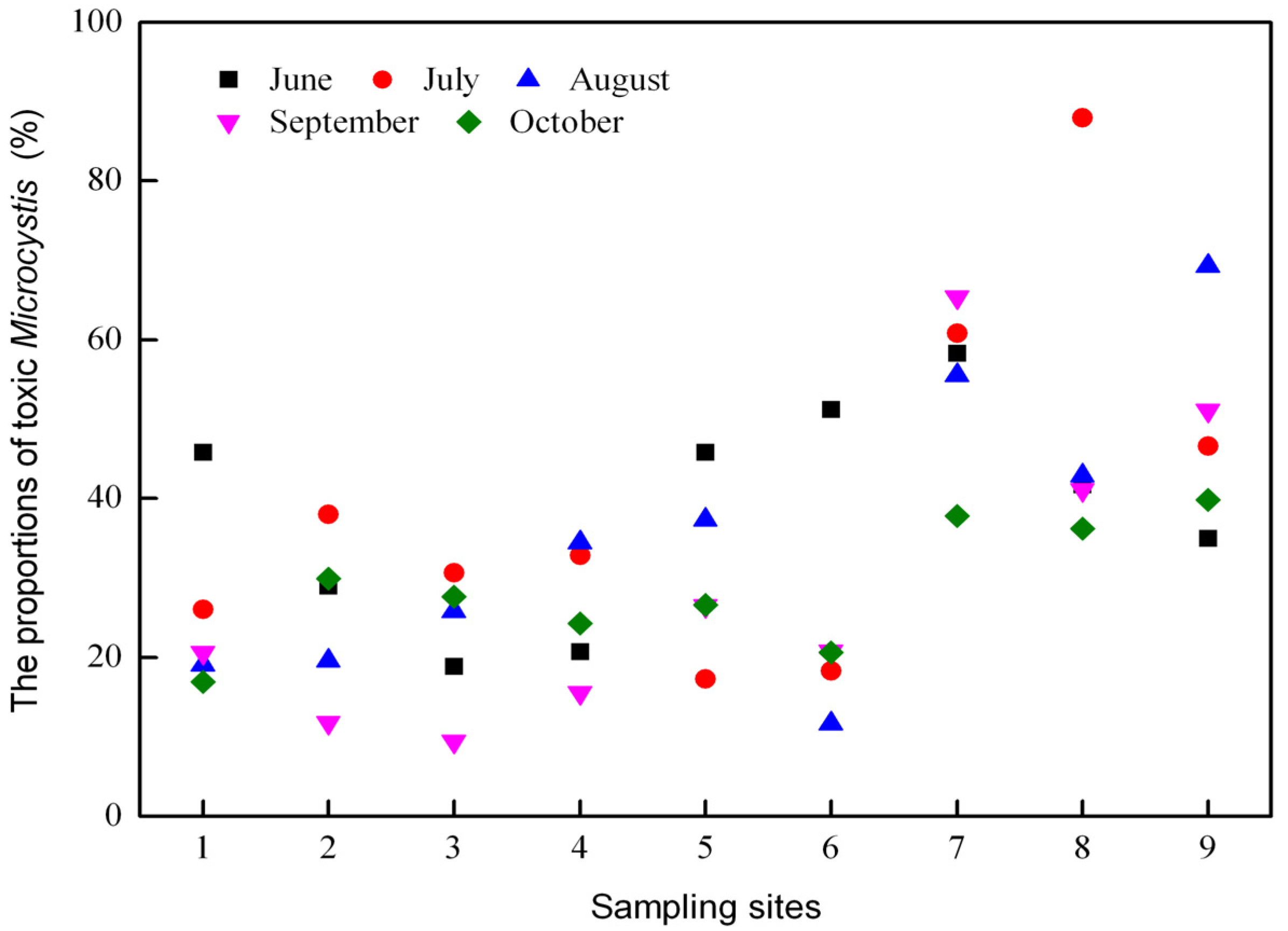
2.2. Dynamics of Microcystis Genotypes and MC Concentrations in Water Samples





2.3. Relationships of Environmental Variables with MC Concentrations, Microcystis 16S rDNA and mcyD Genotypes Abundance
| Variable | Microcystis 16S rDNA | Toxic Microcystis mcyD | MC | |||
|---|---|---|---|---|---|---|
| r | p | r | p | r | p | |
| Toxic Microcystis mcyD | 0.941 | 0.000 | - | - | - | - |
| MC | 0.670 | 0.000 | 0.690 | 0.000 | - | - |
| % toxic proportion | 0.310 | 0.039 | 0.595 | 0.000 | 0.394 | 0.007 |
| TN | 0.532 | 0.000 | 0.598 | 0.000 | 0.625 | 0.000 |
| NH4+–N | 0.298 | 0.046 | 0.351 | 0.018 | 0.368 | 0.013 |
| NO3−–N | 0.325 | 0.029 | 0.457 | 0.002 | 0.289 | 0.054 |
| NO2−–N | 0.273 | 0.069 | 0.364 | 0.014 | 0.381 | 0.010 |
| TP | 0.599 | 0.000 | 0.608 | 0.000 | 0.675 | 0.000 |
| PO43−–P | 0.381 | 0.010 | 0.396 | 0.007 | 0.379 | 0.010 |
| DIC | −0.467 | 0.001 | −0.359 | 0.015 | −0.374 | 0.011 |
| DOC | 0.395 | 0.007 | 0.432 | 0.003 | 0.554 | 0.000 |
| Chl-a | 0.562 | 0.000 | 0.553 | 0.000 | 0.656 | 0.000 |
| Temperature | 0.302 | 0.038 | 0.334 | 0.026 | 0.324 | 0.022 |
| pH | 0.263 | 0.008 | 0.314 | 0.036 | 0.423 | 0.004 |
| DO | −0.051 | 0.741 | −0.038 | 0.803 | 0.024 | 0.875 |
| Secchi depth | −0.365 | 0.014 | −0.356 | 0.016 | −0.398 | 0.007 |
| Water depth | −0.156 | 0.308 | −0.264 | 0.008 | −0.384 | 0.009 |
| Conductivity | −0.126 | 0.409 | −0.002 | 0.987 | −0.006 | 0.970 |
2.4. Stepwise Multiple Regressions Determining Significant Environmental Variables Correlated with MC Concentrations and Abundance of Microcystis 16S rDNA and mcyD Genotypes (Stepwise Multiple Regression Analyses Were Performed to Determine the Key Environmental Variables that Explained the Abundances of MC and Total and Toxic Microcystis in Lake Chaohu (Table 3))
| Model and Variable | R2 | Coefficient | p-Value |
|---|---|---|---|
| Total Microcystis 16S rDNA (R2-adj = 0.53, p < 0.001) | |||
| Constant | - | 5.872 | 0.001 |
| TP | 0.359 | 11.441 | 0.000 |
| DIC | 0.480 | −2.724 | 0.006 |
| Temp | 0.555 | 1.979 | 0.012 |
| Toxic Microcystis mcyD (R2-adj = 0.59, p < 0.001) | |||
| Constant | - | 5.033 | 0.014 |
| TP | 0.370 | 10.765 | 0.000 |
| Temp | 0.516 | 2.667 | 0.003 |
| DIC | 0.561 | −3.490 | 0.004 |
| NO3−–N | 0.625 | 3.558 | 0.013 |
| MC (R2-adj = 0.72, p < 0.001) | |||
| Constant | - | −0.162 | 0.835 |
| TP | 0.391 | 7.605 | 0.000 |
| Temp | 0.625 | 1.892 | 0.000 |
| DIC | 0.735 | −1.816 | 0.000 |
3. Discussion
4. Experimental Section
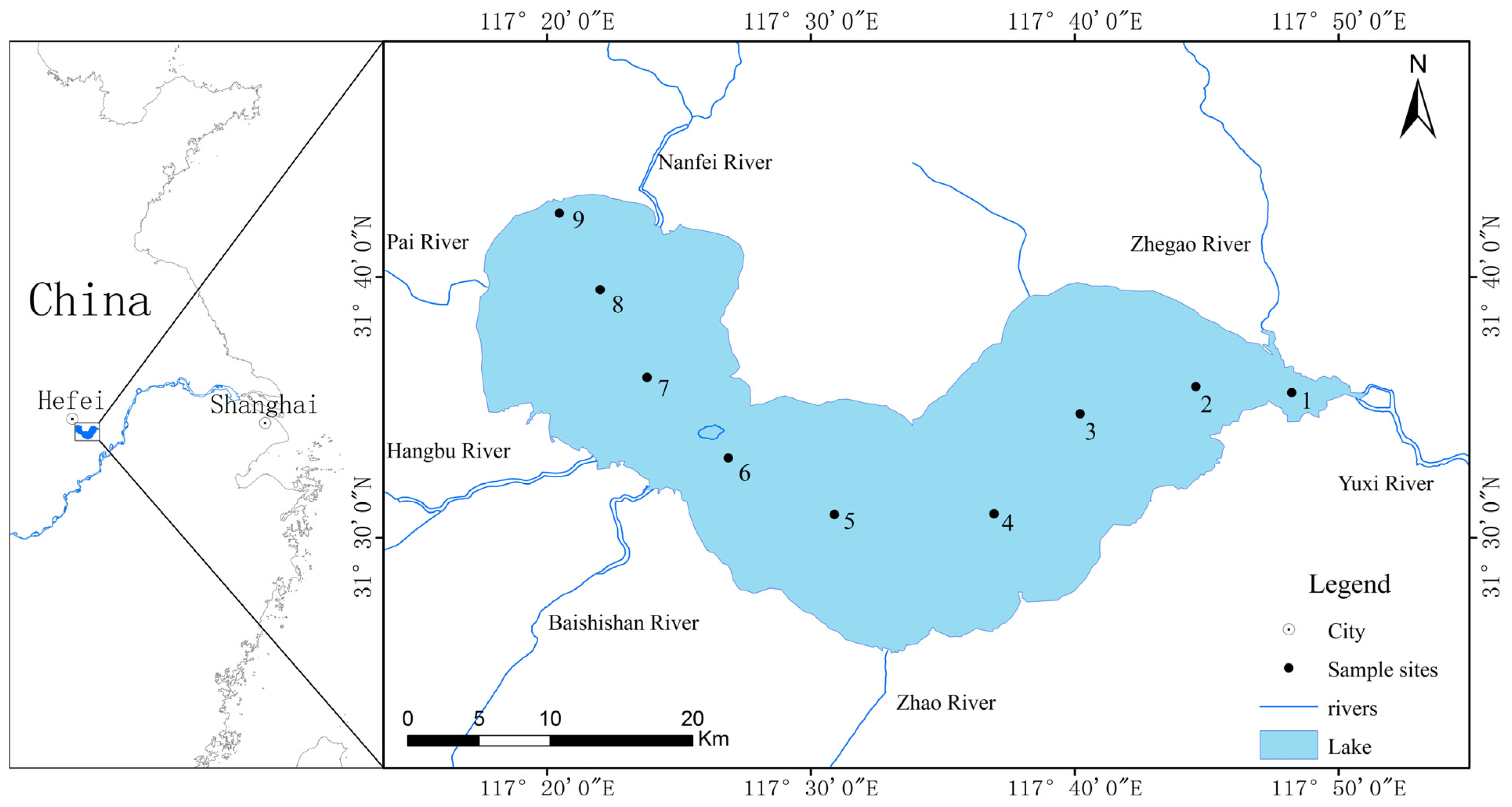
4.1. Sampling Sites and Collection Methods
4.2. Environmental Variables
4.3. Microcystin (MC) Analysis
4.4. DNA Extraction and Quantitative Real-Time PCR
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Env. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef]
- Ouellette, A.J.A.; Wilhelm, S.W. Toxic cyanobacteria: The evolving molecular toolbox. Front. Ecol. Environ. 2003, 1, 359–366. [Google Scholar] [CrossRef]
- Ye, W.; Liu, X.; Tan, J.; Li, D.; Yang, H. Diversity and dynamics of microcystin—Producing cyanobacteria in China’s third largest lake, lake taihu. Harmful Algae 2009, 8, 637–644. [Google Scholar] [CrossRef]
- Falconer, I.R.; Beresford, A.M.; Runnegar, M.T. Evidence of liver damage by toxin from a bloom of the blue-green alga, Microcystis aeruginosa. Med. J. Aust. 1983, 1, 511–514. [Google Scholar] [PubMed]
- Carmichael, W.W. Cyanobacteria secondary metabolites—The cyanotoxins. J. Appl. Bacteriol. 1992, 72, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Welker, M.; von Döhren, H. Cyanobacterial peptides–nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; World Health Organization, St Edmundsbury Press: Bury St Edmunds, Suffolk, UK, 1999. [Google Scholar]
- Bartram, J.; Chorus, I. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Carmichael, W.W.; Falconer, I.R. Diseases related to freshwater blue-green algal toxins, and control measures. In Algal Toxins in Seafood and Drinking Water; Falconer, I.R., Ed.; Academic Press: New York, NY, USA, 1993; pp. 187–209. [Google Scholar]
- Fleming, L.E.; Rivero, C.; Burns, J.; Williams, C.; Bean, J.A.; Shea, K.A.; Stinn, J. Blue green algal (cyanobacterial) toxins, surface drinking water, and liver cancer in florida. Harmful Algae 2002, 1, 157–168. [Google Scholar] [CrossRef]
- Paerl, H. Nutrient and other environmental controls of harmful cyanobacterial blooms along the freshwater–marine continuum. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Springer: New York, NY, USA, 2008; pp. 217–237. [Google Scholar]
- Carmichael, W.W. A status Report of Planktonic Cyanobacteria (Blue-Green Algae) and Their Toxins; EPA/600/R-92/079; United States Environmental Protection Agency: Cincinnati, OH, USA, 1992. [Google Scholar]
- Kardinaal, W.E.A.; Janse, I.; Kamst-van Agterveld, M.; Meima, M.; Snoek, J.; Mur, L.R.; Huisman, J.; Zwart, G.; Visser, P.M. Microcystis genotype succession in relation to microcystin concentrations in freshwater lakes. Aquat. Microb. Ecol. 2007, 48, 1–12. [Google Scholar] [CrossRef]
- Ohtake, A.; Shirai, M.; Aida, T.; Mori, N.; Harada, K.; Matsuura, K.; Suzuki, M.; Nakano, M. Toxicity of microcystis species isolated from natural blooms and purification of the toxin. Appl. Environ. Microbiol. 1989, 55, 3202–3207. [Google Scholar] [PubMed]
- Welker, M.; von Döhren, H.; Täuscher, H.; Steinberg, C.E.; Erhard, M. Toxic microcystis in shallow lake müggelsee (germany)—Dynamics, distribution, diversity. Arch. Hydrobiol. 2003, 157, 227–248. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.W.; Berry, D.L.; Boyer, G.L.; Gobler, C.J. The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae 2009, 8, 715–725. [Google Scholar] [CrossRef]
- Kardinaal, W.E.A.; Visser, P.M. Dynamics of cyanobacterial toxins. In Harmful Cyanobacteria Aquatic Ecology Series; Springer: Dordrecht, The Netherlands, 2005; pp. 41–64. [Google Scholar]
- Yoshida, M.; Yoshida, T.; Takashima, Y.; Hosoda, N.; Hiroishi, S. Dynamics of microcystin-producing and non-microcystin-producing microcystis populations is correlated with nitrate concentration in a japanese lake. FEMS Microbiol. Lett. 2007, 266, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Tillett, D.; Parker, D.L.; Neilan, B.A. Detection of toxigenicity by a probe for the microcystin synthetase a gene (mcya) of the cyanobacterial genus microcystis: Comparison of toxicities with 16s rrna and phycocyanin operon (phycocyanin intergenic spacer) phylogenies. Appl. Environ. Microbiol. 2001, 67, 2810–2818. [Google Scholar] [CrossRef] [PubMed]
- Vaitomaa, J.; Rantala, A.; Halinen, K.; Rouhiainen, L.; Tallberg, P.; Mokelke, L.; Sivonen, K. Quantitative real-time PCR for determination of microcystin synthetase e copy numbers for microcystis and anabaena in lakes. Appl. Environ. Microbiol. 2003, 69, 7289–7297. [Google Scholar] [CrossRef] [PubMed]
- Rinta-Kanto, J.M.; Ouellette, A.J.A.; Boyer, G.L.; Twiss, M.R.; Bridgeman, T.B.; Wilhelm, S. Quantification of toxic microcystis spp. During the 2003 and 2004 blooms in western lake erie using quantitative real-time pcr. Environ. Sci. Technol. 2005, 39, 4198–4205. [Google Scholar] [CrossRef]
- Pimentel, J.S.M.; Giani, A. Estimating toxic cyanobacteria in a brazilian reservoir by quantitative real-time pcr, based on the microcystin synthetase d gene. J. Appl. Phycol. 2013, 25, 1545–1554. [Google Scholar] [CrossRef]
- Rinta-Kanto, J.M.; Konopko, E.A.; DeBruyn, J.M.; Bourbonniere, R.A.; Boyer, G.L.; Wilhelm, S.W. Lake Erie Microcystis: Relationship between microcystin production, dynamics of genotypes and environmental parameters in a large lake. Harmful Algae 2009, 8, 665–673. [Google Scholar] [CrossRef]
- Conradie, K.R.; Barnard, S. The dynamics of toxic microcystis strains and microcystin production in two hypertrofic south african reservoirs. Harmful Algae 2012, 20, 1–10. [Google Scholar] [CrossRef]
- Falconer, I.R. Is there a human health hazard from microcystins in the drinking water supply? Acta Hydroch. Hydrob. 2005, 33, 64–71. [Google Scholar] [CrossRef]
- Krüger, T.; Wiegand, C.; Kun, L.; Luckas, B.; Pflugmacher, S. More and more toxins around-analysis of cyanobacterial strains isolated from lake chao (Anhui province, China). Toxicon 2010, 56, 1520–1524. [Google Scholar] [CrossRef] [PubMed]
- Krüger, T.; Hölzel, N.; Luckas, B. Influence of cultivation parameters on growth and microcystin production of Microcystis aeruginosa (cyanophyceae) isolated from Lake Chao (China). Microbial. Ecol. 2012, 63, 199–209. [Google Scholar] [CrossRef]
- Jiang, Y.J.; He, W.; Liu, W.X.; Qin, N.; Ouyang, H.L.; Wang, Q.M.; Kong, X.Z.; He, Q.S.; Yang, C.; Yang, B. The seasonal and spatial variations of phytoplankton community and their correlation with environmental factors in a large eutrophic chinese lake (Lake Chaohu). Ecol. Indic. 2014, 40, 58–67. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, S.; Zhong, L.; Jin, X.; Sun, S. Seasonal variation characteristics of algae biomass in Chaohu Lake. J. Environ. Sci. 2010, 31, 2056–2062. [Google Scholar]
- Yang, H.; Xie, P.; Xu, J.; Zheng, L.; Deng, D.; Zhou, Q.; Wu, S. Seasonal variation of microcystin concentration in Lake Chaohu, a shallow subtropical lake in the people’s republic of China. Bull. Environ. Contam. Toxicol. 2006, 77, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Zurawell, R.W.; Chen, H.R.; Burke, J.M.; Prepas, E.E. Hepatotoxic cyanobacteria: A review of the biological importance of microcystins in freshwater environments. J. Toxicol. Environ. Health B 2005, 8, 1–37. [Google Scholar] [CrossRef]
- Kardinaal, W.E.A.; Tonk, L.; Janse, I.; Hol, S.; Slot, P.; Huisman, J.; Visser, P.M. Competition for light between toxic and nontoxic strains of the harmful cyanobacterium Microcystis. Appl. Environ. Microbiol. 2007, 73, 2939–2946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falconer, I.R. An overview of problems caused by toxic blue-green algae (cyanobacteria) in drinking and recreational water. Environ. Toxicol. 1999, 14, 5–12. [Google Scholar] [CrossRef]
- Xu, F.L.; Tao, S.; Dawson, R.; Xu, Z. The distributions and effects of nutrients in the sediments of a shallow eutrophic chinese lake. Hydrobiologia 2003, 492, 85–93. [Google Scholar] [CrossRef]
- Cai, Y.F. Comparative Study of Composition and Dynamics of Cyanobacteria and Their Driving Factors in Lake Taihu and Lake Chaohu; Nanjing Institute of Geography and Limnology Chinese Academy of Sciences: Nanjing, China, 2012. [Google Scholar]
- Kurmayer, R.; Kutzenberger, T. Application of real-time PCR for quantification of microcystin genotypes in a population of the toxic cyanobacterium Microcystis sp. Appl. Environ. Microbiol. 2003, 69, 6723–6730. [Google Scholar] [CrossRef]
- Hotto, A.M.; Satchwell, M.F.; Berry, D.L.; Gobler, C.J.; Boyer, G.L. Spatial and temporal diversity of microcystins and microcystin-producing genotypes in oneida lake, ny. Harmful Algae 2008, 7, 671–681. [Google Scholar] [CrossRef]
- Yu, T.; Xie, P.; Dai, M.; Liang, G.D. Determinations of MC-LR and (Dha7) MC-LR concentrations and physicochemical properties by liquid chromatography-tandem mass spectrometry. Bull. Environ. Contam. Toxicol. 2009, 83, 757–760. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Song, L.R.; Ou, D.Y.; Gan, N.Q. Chronic toxicity and responses of several important enzymes in Daphnia magna on exposure to sublethal microcystin-LR. Environ. Toxicol. 2005, 20, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xie, P. Tissue distributions and seasonal dynamics of the hepatotoxic microcystins-LR and-RR in two freshwater shrimps, Palaemon modestus and Macrobrachium nipponensis, from a large shallow, eutrophic lake of the subtropical China. Toxicon 2005, 45, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.Q.; Xie, P.; Guo, L.G.; Li, L.; Miyabara, Y.; Park, H.D. Organ distribution and bioaccumulation of microcystins in freshwater fish at different trophic levels from the eutrophic lake Chaohu, China. Environ. Toxicol. 2005, 20, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xie, P. Accumulation of hepatotoxic microcystins in freshwater mussels, aquatic insect larvae and oligochaetes in a large, shallow eutrophic lake (Lake Chaohu) of subtropical China. Fresenius Environ. Bull. 2008, 17, 849–854. [Google Scholar]
- Chen, J.; Xie, P.; Li, L.; Xu, J. First identification of the hepatotoxic microcystins in the serum of a chronically exposed human population together with indication of hepatocellular damage. Toxicol. Sci. 2009, 108, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Giani, A.; Bird, D.; Prairie, Y.; Lawrence, J. Empirical study of cyanobacterial toxicity along a trophic gradient of lakes. Can. J. Fish. Aquat. Sci. 2005, 62, 2100–2109. [Google Scholar] [CrossRef]
- Graham, J.L.; Jones, J.R.; Jones, S.B.; Downing, J.A.; Clevenger, T.E. Environmental factors influencing microcystin distribution and concentration in the midwestern united states. Water Res. 2004, 38, 4395–4404. [Google Scholar] [CrossRef] [PubMed]
- Joung, S.H.; Oh, H.M.; Ko, S.R.; Ahn, C.Y. Correlations between environmental factors and toxic and non-toxic Microcystis dynamics during bloom in daechung reservoir, korea. Harmful Algae 2011, 10, 188–193. [Google Scholar] [CrossRef]
- Boopathi, T.; Ki, J.S. Impact of environmental factors on the regulation of cyanotoxin production. Toxins 2014, 6, 1951–1978. [Google Scholar] [CrossRef] [PubMed]
- Vézie, C.; Rapala, J.; Vaitomaa, J.; Seitsonen, J.; Sivonen, K. Effect of nitrogen and phosphorus on growth of toxic and nontoxic Microcystis strains and on intracellular microcystin concentrations. Microb. Ecol. 2002, 43, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Tillett, D.; Dittmann, E.; Erhard, M.; von Döhren, H.; Börner, T.; Neilan, B.A. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: An integrated peptide–polyketide synthetase system. Chem.Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Hesse, K.; Kohl, J. Effects of light and nutrient supply on growth and microcystin concentration of different strains of Microcystis aeruginosa. In Cyanotoxins: Occurrence, Effects, Controlling Factors; Chorus, I., Ed.; Springer: Heidelberg/Berlin, Germany, 2001. [Google Scholar]
- Orr, P.T.; Jones, G.J. Relationship between microcystin production and cell division rates in nitrogen-limited Microcystis aeruginosa cultures. Limnol. Oceanogr. 1998, 43, 1604–1614. [Google Scholar] [CrossRef]
- Gobler, C.; Davis, T.W.; Coyne, K.; Boyer, G. Interactive influences of nutrient loading, zooplankton grazing, and microcystin synthetase gene expression on cyanobacterial bloom dynamics in a eutrophic new york lake. Harmful Algae 2007, 6, 119–133. [Google Scholar] [CrossRef]
- Davis, T.W.; Harke, M.J.; Marcoval, M.; Goleski, J.; Orano-Dawson, C.; Berry, D.L.; Gobler, C.J. Effects of nitrogenous compounds and phosphorus on the growth of toxic and non-toxic strains of Microcystis during cyanobacterial blooms. Aquat. Microb. Ecol. 2010, 61, 149–162. [Google Scholar] [CrossRef]
- Te, S.H.; Gin, K.Y.H. The dynamics of cyanobacteria and microcystin production in a tropical reservoir of singapore. Harmful Algae 2011, 10, 319–329. [Google Scholar] [CrossRef]
- Otten, T.G.; Xu, H.; Qin, B.; Zhu, G.; Paerl, H. Spatiotemporal patterns and ecophysiology of toxigenic microcystis blooms in lake taihu, China: Implications for water quality management. Environ. Sci. Technol. 2012, 46, 3480–3488. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A.; Geider, R.J. Temperature and algal growth. New Phytol. 1988, 110, 441–461. [Google Scholar] [CrossRef]
- Reynolds, C.S.; Usher, M.; Saunders, D.; Dobson, A.; Peet, R.; Adam, P.; Birks, H.; Gustafsson, L.; McNeely, J.; Paine, R. Ecology of Phytoplankton; Cambridge University Press: Cambridge, UK, 2006; Volume 535. [Google Scholar]
- Joehnk, K.D.; Huisman, J.E.F.; Sharples, J.; Sommeijer, B.; Visser, P.M.; Stroom, J.M. Summer heatwaves promote blooms of harmful cyanobacteria. Glob. Chang. Biol. 2008, 14, 495–512. [Google Scholar] [CrossRef]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Lee, S.J.; Jang, M.H.; Kim, H.S.; Yoon, B.D.; Oh, H.M. Variation of microcystin content of Microcystis aeruginosa relative to medium N: P ratio and growth stage. J. Appl. Microbiol. 2000, 89, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Yu, Y.; Yang, Z.; Kong, F.; Zhang, T.; Tang, S. The dynamics of toxic and nontoxic Microcystis during bloom in the large shallow lake, Lake taihu, China. Environ. Monit. Assess. 2014, 186, 3053–3062. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W. Nuisance phytoplankton blooms in coastal, estuarine, and inland waters. Limnol. Oceanogr. 1988, 33, 823–847. [Google Scholar] [CrossRef]
- Shapiro, J. The role of carbon dioxide in the initiation and maintenance of blue-green dominance in lakes. Freshw. Biol. 1997, 37, 307–323. [Google Scholar] [CrossRef]
- Raven, J.A.; Giordano, M.; Beardall, J.; Maberly, S.C. Algal evolution in relation to atmospheric CO2: Carboxylases, carbon-concentrating mechanisms and carbon oxidation cycles. Philos. Trans. R. Soc. B 2012, 367, 493–507. [Google Scholar] [CrossRef] [Green Version]
- Giordano, M.; Beardall, J.; Raven, J.A. Co 2 concentrating mechanisms in algae: Mechanisms, environmental modulation, and evolution. Annu. Rev. Plant Biol. 2005, 56, 99–131. [Google Scholar] [CrossRef] [PubMed]
- Okello, W.; Kurmayer, R. Seasonal development of cyanobacteria and microcystin production in ugandan freshwater lakes. Lakes Reserv. Res. Manag. 2011, 16, 123–135. [Google Scholar] [CrossRef]
- Nakano, S.I.; Murabe, A.; Tsujimura, S.; Hayakawa, K.; Nakajima, T.; Kumagai, M.; Jiao, C.; Kawabata, Z.I. Dominance of Microcystis with special reference to carbon availability in Lake water. Microbes Environ. 2003, 18, 38–42. [Google Scholar] [CrossRef]
- Qiu, B.; Gao, K. Effects of CO2 enrichment on the bloom-forming cyanobacterium Microcystis aeruginosa (cyanophyceae): Physiological responses and relationships with the availability of dissolved inorganic carbon. J. Phycol. 2002, 38, 721–729. [Google Scholar] [CrossRef]
- Jähnichen, S.; Ihle, T.; Petzoldt, T.; Benndorf, J. Impact of inorganic carbon availability on microcystin production by Microcystis aeruginosa PCC 7806. Appl. Eenviron. Microbiol. 2007, 73, 6994–7002. [Google Scholar] [CrossRef]
- Jähnichen, S.; Petzoldt, T.; Benndorf, J. Evidence for control of microcystin dynamics in Bautzen Reservoir (germany) by cyanobacterial population growth rates and dissolved inorganic carbon. Arch. Hydrobiol. 2001, 150, 177–196. [Google Scholar]
- Poste, A.E.; Hecky, R.E.; Guildford, S.J. Phosphorus enrichment and carbon depletion contribute to high Microcystis biomass and microcystin concentrations in Ugandan lakes. Limnol. Oceanogr. 2013, 58, 1075–1088. [Google Scholar] [CrossRef]
- Van de Waal, D.B.; Verspagen, J.M.; Finke, J.F.; Vournazou, V.; Immers, A.K.; Kardinaal, W.E.; Tonk, L.; Becker, S.; van Donk, E.; Visser, P.M.; et al. Reversal in competitive dominance of a toxic versus non-toxic cyanobacterium in response to rising CO2. ISME J. 2011, 5, 1438–1450. [Google Scholar]
- Lorenzen, C.J. Determination of chlorophyll and pheopigments: Spectrophotometric equations. Limnol. Oceanogr. 1967, 12, 343–346. [Google Scholar] [CrossRef]
- Barco, M.; Lawton, L.A.; Rivera, J.; Caixach, J. Optimization of intracellular microcystin extraction for their subsequent analysis by high-performance liquid chromatography. J. Chromatogr. A 2005, 1074, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Tillett, D.; Neilan, B.A. Xanthogenate nucleic acid isolation from cultured and environmental cyanobacteria. J. Phycol. 2000, 36, 251–258. [Google Scholar] [CrossRef]
- Neilan, B.A.; Jacobs, D.; Blackall, L.L.; Hawkins, P.R.; Cox, P.T.; Goodman, A.E. Rrna sequences and evolutionary relationships among toxic and nontoxic cyanobacteria of the genus Microcystis. Int. J. Syst. Bacteriol. 1997, 47, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Kaebernick, M.; Neilan, B.A.; Börner, T.; Dittmann, E. Light and the transcriptional response of the microcystin biosynthesis gene cluster. Appl. Environ. Microbiol. 2000, 66, 3387–3392. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, L.; Kong, F.; Zhang, M.; Yang, Z.; Shi, X.; Du, M. The Dynamics of Microcystis Genotypes and Microcystin Production and Associations with Environmental Factors during Blooms in Lake Chaohu, China. Toxins 2014, 6, 3238-3257. https://doi.org/10.3390/toxins6123238
Yu L, Kong F, Zhang M, Yang Z, Shi X, Du M. The Dynamics of Microcystis Genotypes and Microcystin Production and Associations with Environmental Factors during Blooms in Lake Chaohu, China. Toxins. 2014; 6(12):3238-3257. https://doi.org/10.3390/toxins6123238
Chicago/Turabian StyleYu, Li, Fanxiang Kong, Min Zhang, Zhen Yang, Xiaoli Shi, and Mingyong Du. 2014. "The Dynamics of Microcystis Genotypes and Microcystin Production and Associations with Environmental Factors during Blooms in Lake Chaohu, China" Toxins 6, no. 12: 3238-3257. https://doi.org/10.3390/toxins6123238
APA StyleYu, L., Kong, F., Zhang, M., Yang, Z., Shi, X., & Du, M. (2014). The Dynamics of Microcystis Genotypes and Microcystin Production and Associations with Environmental Factors during Blooms in Lake Chaohu, China. Toxins, 6(12), 3238-3257. https://doi.org/10.3390/toxins6123238