Clostridium and Bacillus Binary Enterotoxins: Bad for the Bowels, and Eukaryotic Being



Abstract
:1. Introduction
2. Pathogenic Bacilli and Binary Enterotoxins: Some of the Basics

{kind=link}
{kind=link}
| Toxin Components | Gene Location | Protein Mr (kDa) |
|---|---|---|
| C. perfringens type E ι-toxin | 140 kb plasmid [13] | |
| Ia | 45 [13] | |
| Ib | 94 precursor [13] | |
| 81 activated [13] | ||
| C. perfringens type A BEC | 54.5 kb plasmid [10] | |
| BECa | 47 [10] | |
| BECb | 80 [10] | |
| C. spiroforme CST | chromosome [14] | |
| Sa | 44 [11,14] | |
| Sb | 92 precursor [11,14] | |
| 76 activated [11,14] | ||
| C. difficile CDT | chromosome [8] | |
| CDTa | 48 [8] | |
| CDTb | 99 precursor [8] | |
| 75 activated [8] | ||
| C. botulinum types C and D C2 | chromosome [15] or 107 kb plasmid [16] | |
| C2I | 49 [17] | |
| C2II | 81 precursor [15] | |
| 60 activated [18] | ||
| B. cereus VIP | chromosome [19] | |
| VIP2 | 52 [20] | |
| VIP1 | 100 precursor [20] | |
| 80 activated [20] |
2.1. Clostridium perfringens: ι-Toxin and Binary Enterotoxin (BEC)
2.2. Clostridium spiroforme Toxin (CST)
2.3. Clostridium difficile Toxin (CDT)
2.4. Clostridium botulinum C2 Toxin
2.5. Bacillus cereus Vegetative Insecticidal Proteins (VIPs)
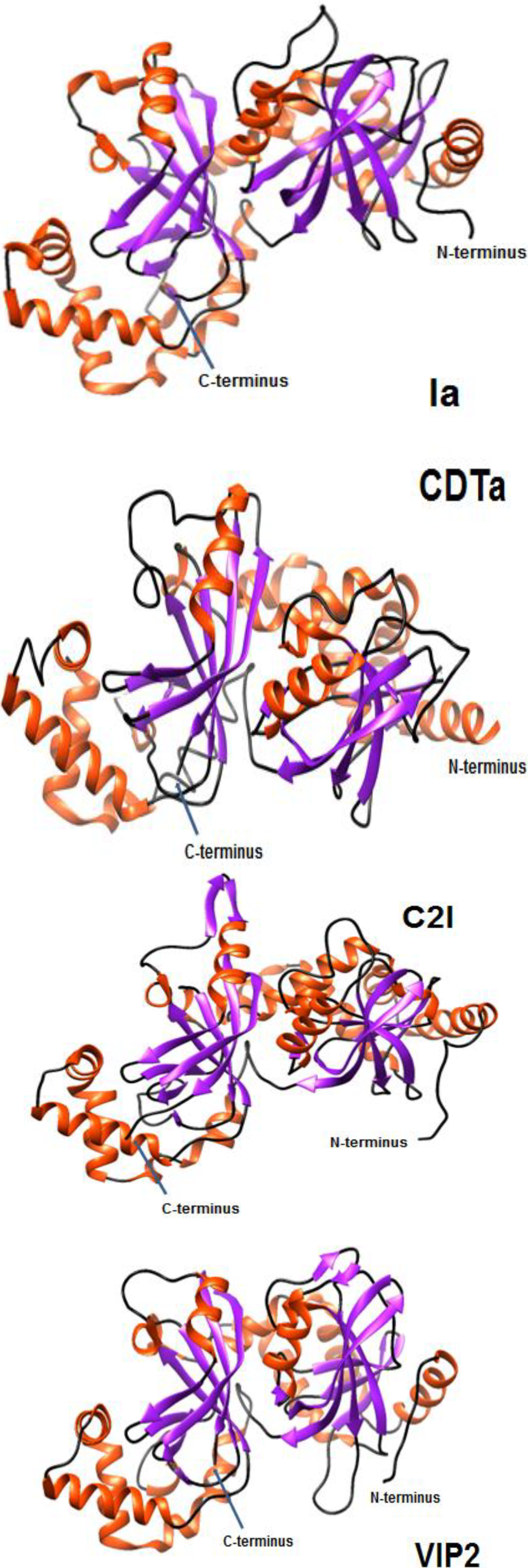
3. Protein Structure and Function

3.1. C. botulinum C2II and C2I
3.2. C. perfringens Ib and Ia
3.3. C. difficile CDTa and CDTb
4. Intoxication Process
4.1. Toxin Binding to Cell
4.2. A Docking to B and Internalization

4.3. ADP-Ribosylation: Destruction of the Actin Cytoskeleton, Intoxicated Cell, and Perhaps the Host
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Borriello, S.P.; Carman, R.J. Association of iota-like toxin and Clostridium spiroforme with both spontaneous and antibiotic-associated diarrhea and colitis in rabbits. J. Clin. Microbiol. 1983, 17, 414–418. [Google Scholar]
- McDonel, J.L. Toxins of Clostridium perfringens types A, B, C, D and E. In Pharmacology of Bacterial Toxins; Dorner, F., Drews, J., Eds.; Pergamon Press: New York, NY, USA, 1986; pp. 477–517. [Google Scholar]
- McKillip, J.L. Prevalence and expression of enterotoxins in Bacillus cereus and other Bacillus spp., a literature review. Antonie Van Leeuwenhoek 2000, 77, 393–399. [Google Scholar]
- Songer, J.G. Clostridial enteric diseases of domestic animals. Clin. Microbiol. Rev. 1996, 9, 216–234. [Google Scholar]
- Stoddart, B.; Wilcox, M.H. Clostridium difficile. Curr. Opin. Infect. Dis. 2002, 15, 513–518. [Google Scholar] [CrossRef]
- Aktories, K.; Schwan, C.; Papatheodorou, P.; Lang, A.E. Bidirectional attack on the actin cytoskeleton. Bacterial protein toxins causing polymerization or depolymerization of actin. Toxicon 2012, 60, 572–581. [Google Scholar]
- Ohishi, I.; Iwasaki, M.; Sakaguchi, G. Purification and characterization of two components of botulinum C2 toxin. Infect. Immun. 1980, 30, 668–673. [Google Scholar]
- Perelle, S.; Gibert, M.; Bourlioux, P.; Corthier, G.; Popoff, M.R. Production of a complete binary toxin (actin-specific ADP-ribosyltransferase) by Clostridium difficile CD196. Infect. Immun. 1997, 65, 1402–1407. [Google Scholar]
- Stiles, B.G.; Wilkins, T.D. Purification and characterization of Clostridium perfringens iota toxin: Dependence on two nonlinked proteins for biological activity. Infect. Immun. 1986, 54, 683–688. [Google Scholar]
- Yonogi, S.; Matsuda, S.; Kawai, T.; Yoda, T.; Harada, T.; Kumeda, Y.; Gotoh, K.; Hiyoshi, H.; Nakamura, S.; Kodama, T.; et al. BEC, a novel enterotoxin of Clostridium perfringens found in human clinical isolates from acute gastroenteritis outbreaks. Infect. Immun. 2014, 82, 2390–2399. [Google Scholar] [CrossRef]
- Popoff, M.R.; Milward, F.W.; Bancillon, B.; Boquet, P. Purification of the Clostridium spiroforme binary toxin and activity of the toxin on HEp-2 cells. Infect. Immun. 1989, 57, 2462–2469. [Google Scholar]
- Han, S.; Craig, J.A.; Putnam, C.D.; Carozzi, N.B.; Tainer, J.A. Evolution and mechanism from structures of an ADP-ribosylating toxin and NAD complex. Nature Struct. Biol. 1999, 6, 932–936. [Google Scholar] [CrossRef]
- Perelle, S.; Gibert, M.; Boquet, P.; Popoff, M.R. Characterization of Clostridium perfringens iota toxin genes and expression in Escherichia coli. Infect. Immun. 1993, 61, 5147–5156. [Google Scholar]
- Gibert, M.; Perelle, S.; Daube, G.; Popoff, M.R. Clostridium spiroforme toxin genes are related to C. perfringens iota toxin genes but have a different genomic localization. Syst. Appl. Microbiol. 1997, 20, 337–347. [Google Scholar]
- Kimura, K.; Kubota, T.; Ohishi, I.; Isogai, E.; Isogai, H.; Fujii, N. The gene for component-II of botulinum C2 toxin. Vet. Microbiol. 1998, 62, 27–34. [Google Scholar] [CrossRef]
- Sakaguchi, Y.; Hayashi, T.; Yamamoto, Y.; Nakayama, K.; Zhang, K.; Ma, S.; Arimitsu, H.; Oguma, K. Molecular analysis of an extrachromosomal element containing the C2 toxin gene discovered in Clostridium botulinum type C. J. Bacteriol. 2009, 191, 3282–3291. [Google Scholar] [CrossRef]
- Fujii, N.; Kubota, T.; Shirakawa, S.; Kimura, K.; Ohishi, I.; Moriishi, K.; Isogai, E.; Isogai, H. Characterization of component-I gene of botulinum C2 toxin and PCR detection of its gene in clostridial species. Biochem. Biophys. Res. Commun. 1996, 220, 353–359. [Google Scholar] [CrossRef]
- Blöcker, D.; Barth, H.; Maier, E.; Benz, R.; Barbieri, J.T.; Aktories, K. The C-terminus of component C2II of Clostridium botulinum C2 toxin is essential for receptor binding. Infect. Immun. 2000, 68, 4566–4573. [Google Scholar]
- Yu, X.; Liu, T.; Liang, X.; Tang, C.; Zhu, J.; Wang, S.; Li, S.; Deng, Q.; Wang, L.; Zheng, A.; Li, P. Rapid detection of vip1-type genes from Bacillus cereus and characterization of a novel vip binary toxin gene. FEMS Microbiol. Lett. 2011, 325, 30–36. [Google Scholar] [CrossRef]
- Warren, G.; Koziel, M.; Mullins, M.A.; Nye; Carr, B.; Desai, N.; Kostichka, K.; Duck, N.; Estruch, J.J. Novel pesticidal proteins and strains. World Intellectual Property Organization. Patent WO96/10083, 1996. [Google Scholar]
- Gibert, M.; Petit, L.; Raffestin, S.; Okabe, A.; Popoff, M.R. Clostridium perfringens iota toxin requires activation of both binding and enzymatic components for cytopathic activity. Infect. Immun. 2000, 68, 3848–3853. [Google Scholar] [CrossRef]
- Perelle, S.; Scalzo, S.; Kochi, S.; Mock, M.; Popoff, M.R. Immunological and functional comparison between Clostridium perfringens iota toxin, C. spiroforme toxin, and anthrax toxins. FEMS Microbiol. Lett. 1997, 146, 117–121. [Google Scholar]
- Popoff, M.R. Molecular biology of actin-ADP-ribosylating toxins. In Handbook of Experimental Pharmacology, Bacterial Protein Toxins; Aktories, K., Just, I., Eds.; Springer-Verlag: Berlin, Germany, 2000; Volume 145, pp. 275–306. [Google Scholar]
- Nakamura, S.; Serikawa, T.; Yamakawa, K.; Nishida, S.; Kozaki, S.; Sakaguchi, G. Sporulation and C2 toxin production by Clostridium botulinum type C strains producing no C1 toxin. Microbiol. Immunol. 1978, 22, 591–596. [Google Scholar] [CrossRef]
- Oakley, C.; Warrack, G. Routine typing of Clostridium welchii. J. Hyg. Camb. 1953, 51, 102–107. [Google Scholar] [CrossRef]
- VPI Anaerobe Laboratory Manual; Holdeman, L.V.; Cato, E.P.; Moore, W.E.C. (Eds.) Southern Printing Co: Blacksburg, VA, USA, 1977; pp. 131–133.
- Walker, P.; Batty, I.; Egerton, J. The typing of C. perfringens and the veterinary background. Papua New Guinea Med. J. 1979, 22, 50–56. [Google Scholar]
- Sarker, M.R.; Singh, U.; McClane, B.A. An update on Clostridium perfringens enterotoxin. J. Nat. Toxins. 2000, 9, 251–266. [Google Scholar]
- Niilo, L. Measurement of biological activities of purified and crude enterotoxin of Clostridium perfringens. Infect. Immun. 1975, 12, 440–442. [Google Scholar]
- Daube, G.; Simon, P.; Limbourg, B.; Manteca, C.; Mainil, J.; Kaeckenbeeck, A. Hybridization of 2659 Clostridium perfringens isolates with gene probes for seven toxins (α, β, ɛ, ɩ, θ, µ and enterotoxin) and for sialidase. Am. J. Vet. Res. 1996, 57, 496–501. [Google Scholar]
- Fach, P.; Popoff, M.R. Detection of enterotoxigenic Clostridium perfringens in food and fecal samples with a duplex PCR and the slide agglutination test. Appl. Environ. Microbiol. 1997, 63, 4232–4236. [Google Scholar]
- Meer, R.R.; Songer, J.G. Multiplex polymerase chain reaction assay for genotyping Clostridium perfringens. Am. J. Vet. Res. 1997, 58, 702–705. [Google Scholar]
- Uzal, F.A.; Plumb, J.J.; Blackall, L.L.; Kelly, W.R. PCR detection of Clostridium perfringens producing different toxins in faeces of goats. Lett. Appl. Microbiol. 1997, 25, 339–344. [Google Scholar] [CrossRef]
- Bosworth, T. On a new type of toxin produced by Clostridium welchii. J. Comp. Path. 1943, 53, 245–255. [Google Scholar]
- Stiles, B.G.; Wilkins, T.D. Clostridium perfringens iota toxin: Synergism between two proteins. Toxicon 1986, 24, 767–773. [Google Scholar] [CrossRef]
- Sakurai, J.; Kobayashi, K. Lethal and dermonecrotic activities of Clostridium perfringens iota toxin: Biological activities induced by cooperation of two nonlinked components. Microbiol. Immunol. 1995, 39, 249–253. [Google Scholar]
- Simpson, L.L.; Stiles, B.G.; Zepeda, H.H.; Wilkins, T.D. Molecular basis for the pathological actions of Clostridium perfringens iota toxin. Infect. Immun. 1987, 55, 118–122. [Google Scholar]
- Schering, B.; Barmann, M.; Chhatwal, G.S.; Geipel, U.; Aktories, K. ADP-ribosylation of skeletal muscle and non-muscle actin by Clostridium perfringens iota toxin. Eur. J. Biochem. 1988, 171, 225–229. [Google Scholar] [CrossRef]
- Vandekerckhove, J.; Schering, B.; Bärmann, M.; Aktories, K. Clostridium perfringens iota toxin ADP-ribosylates skeletal muscle actin in Arg-177. FEBS Lett. 1987, 225, 48–52. [Google Scholar] [CrossRef]
- Blöcker, D.; Behelke, J.; Aktories, K.; Barth, H. Cellular uptake of the binary Clostridium perfringens iota toxin. Infect. Immun. 2001, 69, 2980–2987. [Google Scholar]
- Richard, J.F.; Mainguy, G.; Gibert, M.; Marvaud, J.C.; Stiles, B.G.; Popoff, M.R. Transcytosis of iota toxin across polarized CaCo-2 cells. Mol. Microbiol. 2002, 43, 907–917. [Google Scholar] [CrossRef]
- Stiles, B.G.; Hale, M.L.; Marvaud, J.C.; Popoff, M.R. Clostridium perfringens iota toxin: Binding studies and characterization of cell surface receptor by fluorescence-activated cytometry. Infect. Immun. 2000, 68, 3475–3484. [Google Scholar] [CrossRef]
- Papatheodorou, P.; Carette, J.E.; Bell, G.W.; Schwan, C.; Guttenberg, G.; Brummelkamp, T.R.; Aktories, K. Lipolysis-stimulated lipoprotein receptor (LSR) is the host receptor for the binary toxin Clostridium difficile transferase (CDT). Proc. Natl. Acad. Sci. USA 2011, 108, 16422–16427. [Google Scholar] [CrossRef]
- Wigelsworth, D.J.; Ruthel, G.; Schnell, L.; Herrlich, P.; Blonder, J.; Veenstra, T.D.; Carman, R.J.; Wilkins, T.D.; van Tran Nhieu, G.; Pauillac, S.; et al. CD44 promotes intoxication by the clostridial iota-family toxins. PLoS One 2012, 7, e51356. [Google Scholar] [CrossRef]
- Billington, S.J.; Wieckowski, E.U.; Sarker, M.R.; Bueschel, D.; Songer, J.G.; McClane, B.A. Clostridium perfringens type E animal enteritis isolates with highly conserved, silent enterotoxin gene sequences. Infect. Immun. 1998, 66, 4531–4536. [Google Scholar]
- Hart, B.; Hooper, P. Enterotoxaemia of calves due to Clostridium welchii type E. Aust. Vet. J. 1967, 43, 360–363. [Google Scholar] [CrossRef]
- Ross, H.E.; Warren, M.E.; Barnes, J.M. Clostridium welchii iota toxin: Its activation by trypsin. J. Gen. Microbiol. 1949, 3, 148–152. [Google Scholar]
- Knapp, O.; Benz, R.; Gibert, M.; Marvaud, J.C.; Popoff, M.R. Interaction of Clostridium perfringens iota toxin with lipid bilayer membranes: Demonstration of channel formation by the activated binding component Ib and channel block by the enzyme component Ia. J. Biol. Chem. 2002, 277, 6143–6152. [Google Scholar] [CrossRef]
- Nagahama, M.; Nagayasu, K.; Kobayashi, K.; Sakurai, J. Binding component of Clostridium perfringens iota toxin induces endocytosis in Vero cells. Infect. Immun. 2002, 70, 1909–1914. [Google Scholar] [CrossRef]
- Stiles, B.G.; Hale, M.L.; Marvaud, J.C.; Popoff, M.R. Clostridium perfringens iota toxin: Characterization of the cell-associated iota b complex. Biochem. J. 2002, 367, 801–808. [Google Scholar] [CrossRef]
- Simpson, L.L. The binary toxin produced by Clostridium botulinum enters cells by receptor-mediated endocytosis to exert its pharmacologic effects. J. Pharmacol. Exp. Ther. 1989, 251, 1223–1228. [Google Scholar]
- Minami, J.; Katayama, S.; Matsushita, O.; Matsushita, C.; Okabe, A. Lambda-toxin of Clostridium perfringens activates the precursor of epsilon-toxin by releasing its N- and C-terminal peptides. Microbiol. Immunol. 1997, 41, 527–535. [Google Scholar] [CrossRef]
- Sandvig, K. The Shiga toxins: Properties and action on cells. In The Comprehensive Sourcebook of Bacterial Protein Toxins, 3rd ed.; Alouf, J.E., Popoff, M.R., Eds.; Academic Press: Amsterdam, The Netherlands, 2006; Chapter 17; pp. 310–322. [Google Scholar]
- Garred, O.; Dubinina, E.; Polessakaya, A.; Olsnes, S.; Koslov, J.; Sandvig, K. Role of the disulfide bond in Shiga toxin A-chain for toxin entry into cells. J. Biol. Chem. 1997, 272, 11414–11419. [Google Scholar] [CrossRef]
- Garred, O.; van Deurs, B.; Sandvig, K. Furin-induced cleavage and activation of Shiga toxin. J. Biol. Chem. 1995, 270, 10817–10821. [Google Scholar]
- Hirst, T.R.; D’Souza, J.M. Vibrio cholerae and Escherichia coli thermolabile enterotoxin. In The Comprehensive Sourcebook of Bacterial Protein Toxins, 3rd ed.; Alouf, J.E., Popoff, M.R., Eds.; Academic Press: Amsterdam, The Netherlands, 2006; Chapter 15; pp. 270–290. [Google Scholar]
- Majoul, I.; Ferrari, D.; Soling, H.D. Reduction of protein disulfide bonds in an oxidizing environment—the disulfide bridge of cholera toxin A-subunit is reduced in the endoplasmic reticulum. FEBS Lett. 1997, 401, 104–108. [Google Scholar] [CrossRef]
- Nagahama, M.; Umezaki, M.; Oda, M.; Kobayashi, K.; Tone, S.; Suda, T.; Ishidoh, K.; Sakurai, J. Clostridium perfringens iota-toxin b induces rapid cell necrosis. Infect. Immun. 2011, 79, 4353–4360. [Google Scholar] [CrossRef]
- Hale, M.L.; Marvaud, J.C.; Popoff, M.R.; Stiles, B.G. Detergent-resistant membrane microdomains facilitate Ib oligomer formation and biological activity of Clostridium perfringens iota toxin. Infect. Immun. 2004, 72, 2186–2193. [Google Scholar] [CrossRef]
- Nagahama, M.; Yamaguchi, A.; Hagiyama, T.; Ohkubo, N.; Kobayashi, K.; Sakurai, J. Binding and internalization of Clostridium perfringens iota toxin in lipid rafts. Infect. Immun. 2004, 72, 3267–3275. [Google Scholar] [CrossRef]
- Borriello, S.P.; Davies, H.A.; Carman, R.J. Cellular morphology of Clostridium spiroforme. Vet. Microbiol. 1986, 11, 191–195. [Google Scholar] [CrossRef]
- Carman, R.J.; Borriello, S.P. Observations on an association between Clostridium spiroforme and Clostridium perfringens type E iota enterotoxaemia in rabbits. Eur. J. Chemother. Antibiot. 1982, 2, 143–144. [Google Scholar]
- Carman, R.J.; Borriello, S.P. Infectious nature of Clostridium spiroforme-mediated rabbit enterotoxaemia. Vet. Microbiol. 1984, 9, 497–502. [Google Scholar] [CrossRef]
- Carman, R.J.; Evans, R.H. Experimental and spontaneous clostridial enteropathies of laboratory and free living lagomorphs. Lab. Anim. Sci. 1984, 34, 443–452. [Google Scholar]
- Carman, R.J.; Perelle, S.; Popoff, M.R. Binary toxins from Clostridium spiroforme and Clostridium perfringens. In The Clostridia: Molecular Biology and Pathogenesis; Rood, J., McClane, B.A., Titball, R., Eds.; Academic Press: New York, NY, USA, 1997; Chapter 20; pp. 359–367. [Google Scholar]
- Carman, R.J.; Wilkins, T.D. In vitro susceptibility of rabbit strains of Clostridium spiroforme to antimicrobial agents. Vet. Microbiol. 1991, 28, 391–397. [Google Scholar] [CrossRef]
- Peeters, J.E.; Geeroms, R.; Carman, R.J.; Wilkins, T.D. Significance of Clostridium spiroforme in the enteritis-complex of commercial rabbits. Vet. Microbiol. 1986, 12, 25–31. [Google Scholar]
- Yonushonis, W.P.; Roy, M.J.; Carman, R.J.; Sims, R.E. Diagnosis of spontaneous Clostridium spiroforme iota enterotoxemia in a barrier rabbit breeding colony. Lab. Anim. Sci. 1987, 37, 69–71. [Google Scholar]
- Kaneuchi, C.; Miyazato, T.; Shinjo, T.; Mitsuoka, T. Taxonomic study of helically coiled, sporeforming anaerobes isolated from the intestines of humans and other animals: Clostridium cocleatum sp. nov. and Clostridium spiroforme sp. nov. Int. J. Syst. Bacteriol. 1979, 29, 1–12. [Google Scholar] [CrossRef]
- Popoff, M.R.; Boquet, P. Clostridium spiroforme toxin is a binary toxin which ADP-ribosylates cellular actin. Biochem. Biophys. Res. Commun. 1988, 152, 1361–1368. [Google Scholar] [CrossRef]
- Butt, M.T.; Papendick, R.E.; Carbone, L.G.; Quimby, F.W. A cytotoxicity assay for Clostridium spiroforme enterotoxin in cecal fluid of rabbits. Lab. Anim. Sci. 1994, 44, 52–54. [Google Scholar]
- Eaton, P.; Fernie, D.S. Enterotoxaemia involving Clostridium perfringens iota toxin in a hysterectomy-derived rabbit colony. Lab. Anim. 1980, 14, 347–351. [Google Scholar] [CrossRef]
- Katz, L.; Lamont, J.T.; Trier, J.S.; Sonnenblick, E.B.; Rothman, S.W.; Broitman, S.A.; Rieth, S. Experimental clindamycin associated colitis in rabbits. Evidence for toxin-mediated mucosal damage. Gastroenterology 1978, 74, 246–252. [Google Scholar]
- Lamont, J.T.; Sonnenblick, E.B.; Rothman, S. Role of clostridial toxin in the pathogenesis of clindamycin colitis in rabbits. Gastroenterology 1979, 76, 356–361. [Google Scholar]
- Drigo, I.; Bacchin, C.; Cocchi, M.; Bano, L.; Agnoletti, F. Development of PCR protocols for specific identification of Clostridium spiroforme and detection of sas and sbs genes. Vet. Microbiol. 2008, 13, 414–418. [Google Scholar] [CrossRef]
- Ellis, T.M.; Gregory, A.R.; Logue, G.D. Evaluation of a toxoid for protection of rabbits against enterotoxaemia experimentally induced by trypsin-activated supernatant of Clostridium spiroforme. Vet. Microbiol. 1991, 28, 93–102. [Google Scholar] [CrossRef]
- Agnoletti, F.; Ferro, T.; Guolo, A.; Marcon, B.; Cocchi, M.; Drigo, I.; Mazzolini, E.; Bano, L. A survey of Clostridium spiroforme antimicrobial susceptibility in rabbit breeding. Vet. Microbiol. 2009, 136, 188–191. [Google Scholar] [CrossRef]
- Hall, I.C.; O’Toole, E. Intestinal flora in new-born infants with a description of a new pathogenic anaerobe, Bacillus difficilis. Am. J. Dis. Child. 1935, 49, 390–402. [Google Scholar] [CrossRef]
- Jump, R.L. Clostridium difficile infection in older adults. Aging Health 2013, 9, 403–414. [Google Scholar] [CrossRef]
- He, M.; Sebaihia, M.; Lawley, T.D.; Stabler, R.A.; Dawson, L.F.; Martin, M.J.; Holt, K.E.; Seth-Smith, H.M.; Quail, M.A.; Rance, R.; et al. Evolutinary dynamics of Clostridium difficile over short and long time scales. Proc. Natl. Acad. Sci. USA 2010, 107, 7527–7532. [Google Scholar] [CrossRef]
- Loo, V.G.; Bourgault, A.-M.; Poirier, L.; Lamothe, F.; Michaud, S.; Turgeon, N.; Toye, B.; Beaudoin, A.; Frost, E.H.; Gilca, R.; et al. Host and pathogen factors for Clostridium difficile infection and colonization. N. Eng. J. Med. 2011, 365, 1693–1703. [Google Scholar] [CrossRef]
- Voelker, R. Increased Clostridium difficile virulence demands new treatment approach. JAMA 2010, 303, 2017–2019. [Google Scholar] [CrossRef]
- Tschudin-Sutter, S.; Widmer, A.F.; Perl, T.M. Clostridium difficile: Novel insights on an incessantly challenging disease. Curr. Opin. Infect. Dis. 2012, 25, 405–411. [Google Scholar]
- Seekatz, A.M.; Aas, J.; Gessert, C.E.; Rubin, T.A.; Saman, D.M.; Bakken, J.S.; Young, V.B. Recovery of the gut microbiome following fecal microbiota transplantation. mBio 2014, 5, e00893–e00914. [Google Scholar] [CrossRef]
- Shah, N.; Shaaban, H.; Spira, R.; Slim, J.; Boghossian, J. Intravenous immunoglobulin in the treatment of severe Clostridium difficile colitis. J. Glob. Infect. Dis. 2014, 6, 82–85. [Google Scholar] [CrossRef]
- Gerding, D.N.; Johnson, S.; Rupnik, M.; Aktories, K. Clostridium difficile binary toxin CDT. Mechanism, epidemiology, and potential clinical importance. Gut Microbes 2014, 5, 1–13. [Google Scholar]
- Elliott, B.; Reed, R.; Chang, B.J.; Riley, T.V. Bacteremia with a large clostridial toxin-negative, binary toxin-positive strain of Clostridium difficile. Anaerobe 2009, 15, 249–251. [Google Scholar] [CrossRef]
- Geric, B.; Carman, R.J.; Rupnik, M.; Genheimer, C.W.; Sambol, S.P.; Lyerly, D.M.; Gerding, D.N.; Johnson, S. Binary toxin-producing, large clostridial toxin-negative Clostridium difficile strains are enterotoxic but do not cause disease in hamsters. J. Infect. Dis. 2006, 193, 1143–1150. [Google Scholar] [CrossRef]
- Barbut, F.; Gariazzo, B.; Bonne, L.; Lalande, V.; Burghoffer, B.; Luiuz, R.; Petit, J.C. Clinical features of Clostridium difficile-associated infections and molecular characterization of strains: Results of a retrospective study, 2000–2004. Infect. Cont. Hosp. Epidemiol. 2007, 28, 131–139. [Google Scholar] [CrossRef]
- Bacci, S.; Molbak, K.; Kjeldsen, M.K.; Olsen, K.E. Binary toxin and death after Clostridium difficile infection. Emerg. Infect. Dis. 2011, 17, 976–982. [Google Scholar] [CrossRef]
- Keessen, E.C.; Gaastra, W.; Lipman, L.J. Clostridium difficile infection in humans and animals, differences and similarities. Vet. Microbiol. 2011, 153, 205–217. [Google Scholar] [CrossRef]
- Knight, D.R.; Riley, T.V. Prevalence of gastrointestinal Clostridium difficile carriage in Australian sheep and lambs. Appl. Environ. Microbiol. 2013, 79, 5689–5692. [Google Scholar] [CrossRef]
- Hensgens, M.P.; Keessen, E.C.; Squire, M.M.; Riley, T.V.; Koene, M.G.; de Boer, E.; Lipman, L.J.; Kuijper, E.J. European Society of Clinical Microbiology and Infectious Diseases Study Group for Clostridium difficile. Clostridium difficile infection in the community: A zoonotic disease? Clin. Microbiol. Infect. 2012, 18, 635–645. [Google Scholar] [CrossRef]
- Gould, L.H.; Limbago, B. Clostridium difficile in food and domestic animals: A new food-borne pathogen? Clin. Infect. Dis. 2010, 51, 577–582. [Google Scholar] [CrossRef]
- Metcalf, D.S.; Costa, M.C.; Dew, W.M.; Weese, J.S. Clostridium difficile in vegetables, Canada. Lett. Appl. Microbiol. 2010, 51, 600–602. [Google Scholar] [CrossRef]
- Harvey, R.B.; Norman, K.N.; Andrews, K.; Norby, B.; Hume, M.E.; Scanlan, C.M.; Hardin, M.D.; Scott, H.M. Clostridium difficile in retail meat and processing plants in Texas. J. Vet. Diagn. Invest. 2014, 23, 807–811. [Google Scholar] [CrossRef]
- Romano, V.; Pasquale, V.; Krovacek, K.; Mauri, F.; Demarta, A.; Dumontet, S. Toxigenic Clostridium difficile PCR ribotypes from wastewater treatment plants in southern Switzerland. Appl. Environ. Microbiol. 2012, 78, 6643–6646. [Google Scholar] [CrossRef]
- Xu, C.; Weese, J.S.; Flemming, C.; Odumeru, J.; Warriner, K. Fate of Clostridium difficile during wastewater treatment and incidence in Southern Ontario watersheds. J. Appl. Microbiol. 2014, 117, 891–904. [Google Scholar] [CrossRef]
- Carman, R.J.; Stevens, A.L.; Lyerly, M.W.; Hiltonsmith, M.F.; Stiles, B.G.; Wilkins, T.D. Clostridium difficile binary toxin (CDT) and diarrhea. Anaerobe 2011, 17, 161–165. [Google Scholar] [CrossRef]
- Popoff, M.R.; Rubin, E.J.; Gill, D.M.; Boquet, P. Actin-specific ADP-ribosyltransferase produced by a Clostridium difficile strain. Infect. Immun. 1988, 56, 2299–2306. [Google Scholar]
- Gülike, I.; Pfeifer, G.; Liese, J.; Fritz, M.; Hofmann, F.; Aktories, K.; Barth, H. Characterization of the enzymatic component of the ADP-ribosyltransferase toxin CDTa from Clostridium difficile. Infect. Immun. 2001, 69, 6004–6011. [Google Scholar]
- Braun, M.; Herholz, C.; Straub, R.; Choisat, B.; Frey, J.; Nicolet, J.; Kuhnert, P. Detection of the ADP-ribosyltransferase toxin gene (cdtA) and its activity in Clostridium difficile isolates from equidae. FEMS Microbiol. Lett. 2000, 184, 29–33. [Google Scholar] [CrossRef]
- Geric, B.; Johnson, S.; Gerding, D.N.; Grabnar, M.; Rupnik, M. Frequency of binary toxin genes among Clostridium difficile strains that do not produce large clostridial toxins. J. Clin. Microbiol. 2003, 41, 5227–5232. [Google Scholar] [CrossRef]
- Stubbs, S.; Rupnik, M.; Gibert, M.; Brazier, J.; Duerden, B.; Popoff, M. Production of actin-specific ADP-ribosyltransferase (binary toxin) by strains of Clostridium difficile. FEMS Microbiol. Lett. 2000, 186, 307–312. [Google Scholar] [CrossRef]
- Paltansing, S.; van den Berg, R.J.; Guseinova, R.A.; Visser, C.E.; van der Vorm, R.R.; Kuijper, E.J. Characteristics and incidence of Clostridium difficile-associated disease in The Netherlands, 2005. Eur. Soc. Clin. Microbiol. Infect. Dis. 2007, 13, 1058–1064. [Google Scholar] [CrossRef]
- Rafila, A.; Indra, A.; Popescu, G.A.; Wewalka, G.; Allerberger, F.; Benea, S.; Badicut, I.; Aschbacher, R.; Huhulescu, S. Occurrence of Clostridium difficile infections due to PCR ribotype 027 in Bucharest, Romania. J. Infect. Dev. Ctries. 2014, 8, 694–698. [Google Scholar] [CrossRef]
- Spigaglia, P.; Mastrantonio, P. Comparative analysis of Clostridium difficile clinical isolates belonging to different genetic lineages and time periods. J. Med. Microbiol. 2004, 53, 1129–1136. [Google Scholar] [CrossRef]
- Devriese, P.P. On the discovery of Clostridium botulinum. J. Hist. Neurosci. 1999, 8, 43–50. [Google Scholar] [CrossRef]
- Simpson, L.L. The origin, structure, and pharmacological activity of botulinum toxin. Pharmacol. Rev. 1981, 33, 155–187. [Google Scholar]
- Ermert, L.; Bruckner, H.; Walmrath, D.; Grimminger, F.; Aktories, K.; Suttorp, N.; Duncker, H.R.; Seeger, W. Role of endothelial cytoskeleton in high-permeability edema due to botulinum C2 toxin in perfused rabbit lungs. Am. J. Physiol. 1995, 268, 753–761. [Google Scholar]
- Iwasaki, M.; Ohishi, I.; Sakaguchi, G. Evidence that botulinum C2 toxin has two dissimilar components. Infect. Immun. 1980, 29, 390–394. [Google Scholar]
- Jensen, W.I.; Duncan, R.M. The susceptibility of the mallard duck (Anas platyrhynchos) to Clostridium botulinum C2 toxin. Jpn. J. Med. Sci. Biol. 1980, 33, 81–86. [Google Scholar] [CrossRef]
- Kurazono, H.; Hosokawa, M.; Matsuda, H.; Sakaguchi, G. Fluid accumulation in the ligated intestinal loop and histopathological changes of the intestinal mucosa caused by Clostridium botulinum C2 toxin in the pheasant and chicken. Res. Vet. Sci. 1987, 42, 349–353. [Google Scholar]
- Ohishi, I. Response of mouse intestinal loop to botulinum C2 toxin: Enterotoxic activity induced by cooperation of nonlinked protein components. Infect. Immun. 1983, 40, 691–695. [Google Scholar]
- Ohishi, I. Lethal and vascular permeability activities of botulinum C2 toxin induced by separate injections of the two toxin components. Infect. Immun. 1983, 40, 336–339. [Google Scholar]
- Ohishi, I.; Iwasaki, M.; Sakaguchi, G. Vascular permeability activity of botulinum C2 toxin elicited by cooperation of two dissimilar protein components. Infect. Immun. 1980, 31, 890–895. [Google Scholar]
- Ohishi, I.; Sakaguchi, G. Oral toxicities of Clostridium botulinum type C and D toxins of different molecular sizes. Infect. Immun. 1980, 28, 303–309. [Google Scholar]
- Simpson, L.L. A comparison of the pharmacological properties of Clostridium botulinum type C1 and C2 toxins. J. Pharmacol. Exp. Ther. 1982, 223, 695–701. [Google Scholar]
- Yamakawa, K.; Nishida, S.; Nakamura, S. C2 toxicity in extract of Clostridium botulinum type C spores. Infect. Immun. 1983, 41, 858–860. [Google Scholar]
- Frieben, W.R.; Duncan, C.L. Homology between enterotoxin protein and spore structural protein in Clostridium perfringens type A. Eur. J. Biochem. 1973, 39, 393–401. [Google Scholar]
- Barth, H.; Blöcker, D.; Behlke, J.; Bergsma-Schutter, W.; Brisson, A.; Benz, R.; Aktories, K. Cellular uptake of Clostridium botulinum C2 toxin requires oligomerization and acidification. J. Biol. Chem. 2000, 275, 18704–18711. [Google Scholar] [CrossRef]
- Schmid, A.; Benz, R.; Just, I.; Aktories, K. Interaction of Clostridium botulinum C2 toxin with lipid bilayer membranes: Formation of cation-selective channels and inhibition of channel function by chloroquine and peptides. J. Biol. Chem. 1994, 269, 16706–16711. [Google Scholar]
- Miyake, M.; Ohishi, I. Response of tissue-cultured cynomolgus monkey kidney cells to botulinum C2 toxin. Microb. Pathog. 1987, 3, 279–286. [Google Scholar] [CrossRef]
- Ohishi, I.; Yanagimoto, A. Visualizations of binding and internalization of two nonlinked protein components of botulinum C2 toxin in tissue culture cells. Infect. Immun. 1992, 60, 4648–4655. [Google Scholar]
- Ohishi, I.; Hama, Y. Purification and characterization of heterologous component IIs of botulinum C2 toxin. Microbiol. Immunol. 1992, 36, 221–229. [Google Scholar] [CrossRef]
- Ohishi, I.; Okada, Y. Heterogeneities of two components of C2 toxin produced by Clostridium botulinum types C and D. J. Gen. Microbiol. 1986, 132, 125–131. [Google Scholar]
- Simpson, L.L. Molecular basis for the pharmacological actions of Clostridium botulinum type C2 toxin. J. Pharmacol. Exp. Ther. 1984, 230, 665–669. [Google Scholar]
- Aktories, K.; Bärmann, M.; Ohishi, I.; Tsuyama, S.; Jakobs, K.H.; Habermann, E. Botulinum C2 toxin ADP-ribosylates actin. Nature 1986, 322, 390–392. [Google Scholar] [CrossRef]
- Ohishi, I.; Tsuyama, S. ADP-ribosylation of nonmuscle actin with component I of C2 toxin. Biochem. Biophys. Res. Commun. 1986, 136, 802–806. [Google Scholar] [CrossRef]
- Aktories, K.; Ankenbauer, T.; Schering, B.; Jakobs, K.H. ADP-ribosylation of platelet actin by botulinum C2 toxin. Eur. J. Biochem. 1986, 161, 155–162. [Google Scholar] [CrossRef]
- Jucovic, M.; Walters, F.S.; Warren, G.W.; Palekar, N.V.; Chen, J.S. From enzyme to zymogen: Engineering Vip2, an ADP-ribosyltransferase from Bacillus cereus, for conditional toxicity. Protein Eng. Des. Select. 2008, 21, 631–638. [Google Scholar] [CrossRef]
- Fang, J.; Xu, X.; Wang, P.; Zhao, J.Z.; Shelton, A.M.; Cheng, J.; Feng, M.G.; Shen, Z. Characterization of chimeric Bacillus thuringiensis Vip3 toxins. Appl. Env. Microbiol. 2007, 73, 956–961. [Google Scholar] [CrossRef]
- Michelet, N.; Granum, P.E.; Mahillon, J. Bacillus cereus enterotoxins, bi- and tricomponent cytolysins, and other hemolysins. In The Comprehensive Sourcebook of Bacterial Protein Toxins, 3rd ed.; Alouf, J.E., Popoff, M.R., Eds.; Academic Press: Amsterdam, The Netherlands, 2006; Chapter 46; pp. 779–790. [Google Scholar]
- Margulis, L.; Jorgensen, J.Z.; Dolan, S.; Kolchinsky, R.; Rainey, F.A.; Lo, S.C. The Arthromitus stage of Bacillus cereus: Intestinal symbionts of animals. Proc. Natl. Acad. Sci. USA 1998, 95, 1236–1241. [Google Scholar]
- Schleberger, C.; Hochmann, H.; Barth, H.; Aktories, K.; Schulz, G.E. Structure and action of the binary C2 toxin from Clostridium botulinum. J. Mol. Biol. 2006, 364, 705–715. [Google Scholar] [CrossRef]
- Tsuge, H.; Nagahama, M.; Nishimura, H.; Hisatsune, J.; Sakaguchi, Y.; Itogawa, Y.; Katunuma, N; Sakurai, J. Crystal structure and site-directed mutagenesis of enzymatic components from Clostridium perfringens iota-toxin. J. Mol. Biol. 2003, 325, 471–483. [Google Scholar]
- Sundriyal, A.; Roberts, A.K.; Shone, C.C.; Acharya, K.R. Structural basis for substrate recognition in the enzymatic component of ADP-ribosyltransferase toxin CDTa from Clostridium difficile. J. Biol. Chem. 2009, 284, 28713–28719. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Blöcker, D.; Bachmeyer, C.; Benz, R.; Aktories, K.; Barth, H. Channel formation by the binding component of Clostridium botulinum C2 toxin: Glutamate 307 of C2II affects channel properties in vitro and pH-dependent C2I translocation in vivo. Biochemistry 2003, 42, 5368–5377. [Google Scholar]
- Barth, H.; Roebling, R.; Fritz, M.; Aktories, K. The binary Clostridium botulinum C2 toxin as a protein delivery system: Identification of the minimal protein region necessary for interaction of toxin components. J. Biol. Chem. 2002, 277, 5074–5081. [Google Scholar] [CrossRef]
- Stein, P.E.; Boodhoo, A.; Armstrong, G.D.; Cockle, S.A.; Klein, M.H.; Read, R.J. The crystal structure of pertussis toxin. Structure 1994, 2, 45–57. [Google Scholar] [CrossRef]
- Choe, S.; Bennett, M.J.; Fujii, G.; Curmi, P.M.; Kantardjieff, K.A.; Collier, R.J.; Eisenberg, D. The crystal structure of diphtheria toxin. Nature 1992, 357, 216–222. [Google Scholar] [CrossRef]
- Sixma, T.K.; Pronk, S.E.; Kalk, K.H.; Wartna, E.S.; van Zanten, B.A.; Witholt, B.; Hol, W.G. Crystal structure of a cholera toxin-related heat-labile enterotoxin from E. coli. Nature 1991, 351, 371–377. [Google Scholar]
- Li, M.; Dyda, F.; Benhar, I.; Pastan, I.; Davies, D.R. Crystal structure of the catalytic domain of Pseudomonas exotoxin A complexed with a nicotinamide adenine dinucleotide analog: Implications for the activation process and for ADP ribosylation. Proc. Natl. Acad. Sci. USA 1995, 93, 6902–6906. [Google Scholar]
- Van Damme, J.; Jung, M.; Hofmann, F.; Just, I.; Vandekerckhove, J.; Aktories, K. Analysis of the catalytic site of the actin ADP-ribosylating Clostridium perfringens iota toxin. FEBS Lett. 1996, 380, 291–295. [Google Scholar]
- Barth, H.; Preiss, J.C.; Hofmann, F.; Aktories, K. Characterization of the catalytic site of the ADP-ribosyltransferase Clostridium botulinum C2 toxin by site-directed mutagenesis. J. Biol. Chem. 1998, 273, 29506–29511. [Google Scholar]
- Marvaud, J.C.; Smith, T.; Hale, M.L.; Popoff, M.R.; Smith, L.A.; Stiles, B.G. Clostridium perfringens iota toxin: Mapping of receptor binding and Ia docking domains on Ib. Infect. Immun. 2001, 69, 2435–2441. [Google Scholar] [CrossRef]
- Fritz, G.; Schroeder, P.; Aktories, K. Isolation and characterization of a Clostridium botulinum C2 toxin-resistant cell line: Evidence for possible involvement of the cellular C2II receptor in growth regulation. Infect. Immun. 1995, 63, 2334–2340. [Google Scholar]
- Stiles, B.G.; Blöcker, D.; Hale, M.L.; Guetthoff, M.A.; Barth, H. Clostridium botulinum C2 toxin: Binding studies with fluorescence-activated cytometry. Toxicon 2002, 40, 1135–1140. [Google Scholar] [CrossRef]
- Sakurai, J.; Nagahama, M.; Oda, M.; Tsuge, H.; Kobayashi, K. Clostridium perfringens iota toxin: Structure and function. Toxins 2009, 1, 208–228. [Google Scholar] [CrossRef]
- Kobayashi, K.; Nagahama, M.; Ohkubo, N.; Kojima, T.; Shirai, H.; Iwamoto, S.; Oda, M.; Sakurai, J. Role of Ca2+-binding motif in cytotoxicity induced by Clostridium perfringens iota toxin. Microb. Pathog. 2008, 44, 265–270. [Google Scholar] [CrossRef]
- Tsuge, H.; Nagahama, M.; Oda, M.; Iwamoto, S.; Utsunomiya, H.; Marquez, V.E.; Katunuma, N.; Nishizawa, M.; Sakurai, J. Structural basis of actin recognition and arginine ADP-ribosylation by Clostridium perfringens iota toxin. Proc. Natl. Acad. Sci. USA 2008, 105, 7399–7404. [Google Scholar] [CrossRef]
- Nagahama, M.; Kihara, A.; Miyawaki, T.; Mukai, M.; Sakaguchi, Y.; Ochi, S.; Sakurai, J. Characterization of the enzymatic component of Clostridium perfringens iota toxin. J. Bacteriol. 2000, 183, 2096–2103. [Google Scholar]
- Carroll, S.F.; Collier, R.J. NAD binding site of diphtheria toxin: Identification of a residue within the nicotinamide subsite by photochemical modification with NAD. Proc. Natl. Acad. Sci. USA 1984, 81, 3307–3311. [Google Scholar] [CrossRef]
- Jung, M.; Just, I.; van Damme, J.; Vandekerckhove, J.; Aktories, K. NAD-binding site of the C3-like ADP-ribosyltransferase from Clostridium limosum. J. Biol. Chem. 1993, 268, 23215–23218. [Google Scholar]
- Sakurai, J.; Nagahama, M.; Hisatsune, J.; Katunuma, N.; Tsuge, H. Clostridium perfringens iota-toxin, ADP-ribosyltransferase: Structure and mechanism of action. Adv. Enzyme Regul. 2003, 43, 361–377. [Google Scholar] [CrossRef]
- Takada, T.; Iida, K.; Moss, J. Conservation of a common motif in enzymes catalyzing ADP-ribose transfer. J. Biol. Chem. 1995, 270, 541–544. [Google Scholar] [CrossRef]
- Chowdhury, H.H.; Popoff, M.R.; Zorec, R. Actin cytoskeleton depolymerization with Clostridium spiroforme toxin enhances the secretory activity of rat melanotrophs. J. Physiol. 1999, 521, 389–395. [Google Scholar] [CrossRef]
- Marvaud, J.C.; Stiles, B.G.; Chenal, A.; Gillet, D.; Gibert, M.; Smith, L.A.; Popoff, M.R. Clostridium perfringens iota toxin. Mapping of the Ia domain involved in docking with Ib and cellular internalization. J. Biol. Chem. 2002, 277, 43659–43666. [Google Scholar]
- Papatheodorou, P.; Hornuss, D.; Nolke, T.; Hemmasi, S.; Castonguay, J.; Picchianti, M.; Aktories, K. Clostridium difficile binary toxin CDT induces clustering of the lipolysis-stimulated lipoprotein receptor into lipid rafts. mBio 2013, 4, e00244–13. [Google Scholar] [CrossRef]
- Fivaz, M.; Abrami, L.; Tsitrin, Y.; van der Goot, F.G. Not as simple as just punching a hole. Toxicon 2001, 39, 1637–1645. [Google Scholar] [CrossRef]
- Lafont, F.; van Tran Nhieu, G.; Hanada, K.; Sansonetti, P.; van der Goot, F.G. Initial steps of Shigella infection depend on the cholesterol/sphingolipid raft-mediated CD44-IpaB interaction. EMBO J. 2002, 21, 4449–4457. [Google Scholar] [CrossRef]
- Miyata, S.; Minami, J.; Tamai, E.; Matsushita, O.; Shimamota, S.; Okabe, A. Clostridium perfringens epsilon-toxin forms a heptameric pore within the detergent-insoluble microdomains of Madin-Darby canine kidney cells and rat synaptosomes. J. Biol. Chem. 2002, 277, 39463–39468. [Google Scholar] [CrossRef]
- Simons, K.; Ehehalt, R. Cholesterol, lipid rafts, and disease. J. Clin. Invest. 2002, 110, 597–603. [Google Scholar] [CrossRef]
- Nagahama, M.; Hagiyama, T.; Kojima, T.; Aoyanagi, K.; Takahashi, C.; Oda, M.; Sakaguchi, Y.; Oguma, K.; Sakurai, J. Binding and internalization of Clostridium botulinum C2 toxin. Infect. Immun. 2009, 77, 5139–5148. [Google Scholar] [CrossRef]
- Nagahama, M.; Umezaki, M.; Tashiro, R.; Oda, M.; Kobayashi, K.; Shibutani, M.; Takagishi, T.; Ishidoh, K.; Fukuda, M.; Sakurai, J. Intracellular trafficking of Clostridium perfringens iota-toxin b. Infect. Immun. 2012, 80, 3410–3416. [Google Scholar] [CrossRef]
- Ohishi, I.; Miyake, M. Binding of the two components of C2 toxin to epithelial cells and brush borders of mouse intestine. Infect. Immun. 1985, 48, 769–775. [Google Scholar]
- Sugii, S.; Kozaki, S. Hemagglutinating and binding properties of botulinum C2 toxin. Biochim. Biophys. Acta 1990, 1034, 176–179. [Google Scholar]
- Eckhardt, M.; Barth, H.; Blöcker, D.; Aktories, K. Binding of Clostridium botulinum C2 toxin to asparagine-linked complex and hybrid carbohydrates. J. Biol. Chem. 2000, 275, 2328–2334. [Google Scholar] [CrossRef]
- Bachmeyer, C.; Benz, R.; Barth, H.; Aktories, K.; Gibert, M.; Popoff, M. Interaction of Clostridium botulinum C2-toxin with lipid bilayer membranes and Vero cells: Inhibition of channel function by chloroquine and related compounds in vitro and intoxication in vivo. FASEB J. 2001, 15, 1658–1660. [Google Scholar]
- Bachmeyer, C.; Orlik, F.; Barth, H.; Aktories, K.; Benz, R. Mechanism of C2-toxin inhibition by fluphenazine and related compounds: Investigation of their binding kinetics to the C2II-channel using the current noise analysis. J. Mol. Biol. 2003, 333, 527–540. [Google Scholar] [CrossRef]
- Blöcker, D.; Pohlamnn, K.; Haug, G.; Bachmeyer, C.; Benz, R.; Aktories, K.; Barth, H. Clostridium botulinum C2 toxin: Low pH-induced pore formation is required for translocation of the enzyme component C2I into the cytosol of host cells. J. Biol. Chem. 2003, 278, 37360–37367. [Google Scholar]
- Barth, H.; Stiles, B.G. Binary actin-ADP-ribosylating toxins and their use as molecular Trojan horses for drug delivery into eukaryotic cells. Curr. Med. Chem. 2008, 15, 459–469. [Google Scholar] [CrossRef]
- Ohishi, I.; Miyake, M.; Ogura, H.; Nakamura, S. Cytopathic effect of botulinum C2 toxin on tissue-culture cells. FEMS Microbiol. Lett. 1984, 23, 281–284. [Google Scholar] [CrossRef]
- Papatheodorou, P.; Wilczek, C.; Nolke, T.; Guttenberg, G.; Hornuss, D.; Schwan, C.; Aktories, K. Identification of the cellular receptor of Clostridium spiroforme toxin. Infect. Immun. 2012, 80, 1418–1423. [Google Scholar] [CrossRef]
- Blonder, J.; Hale, M.L.; Chan, K.C.; Yu, L.R.; Lucas, D.A.; Conrads, T.P.; Zhou, M.; Popoff, M.R.; Issaq, H.J.; Stiles, B.G.; et al. Quantitative profiling of the detergent-resistant membrane proteome of iota-b toxin induced Vero cells. J. Prot. Res. 2005, 4, 523–531. [Google Scholar] [CrossRef]
- Fagan-Solis, K.; Reaves, D.K.; Rangel, M.C.; Popoff, M.R.; Stiles, B.G.; Fleming, J.M. Challenging the roles of CD44 and lipolysis stimulated lipoprotein receptor in conveying Clostridium perfringens iota toxin cytotoxicity in breast cancer. Mol. Cancer 2014, 13, 163–168. [Google Scholar]
- Sandvig, K.; van Deurs, B. Endocytosis, intracellular transport, and cytotoxic action of shiga toxin and ricin. Physiol. Rev. 1996, 76, 949–966. [Google Scholar]
- Orlandi, P.A.; Curran, P.K.; Fishman, P.H. Brefeldin A blocks the response of cultured cells to cholera toxin. Implications for intracellular trafficking in toxin action. J. Biol. Chem. 1993, 8, 12010–12016. [Google Scholar]
- Chardin, P.; McCormick, F. Brefeldin A: The advantage of being uncompetitive. Cell 1999, 97, 153–155. [Google Scholar] [CrossRef]
- Madshus, I.H.; Stenmark, H.; Sandvig, K.; Olsnes, S. Entry of diphtheria toxin-protein A chimeras into cells. J. Biol. Chem. 1991, 266, 17446–17453. [Google Scholar]
- Friedlander, A.M. Macrophages are sensitive to anthrax lethal toxin through an acid-dependent process. J. Biol. Chem. 1986, 261, 7123–7126. [Google Scholar]
- Gruenberg, J.; Howell, K.E. Membrane traffic in endocytosis: Insights from cell-free assays. Ann. Rev. Cell Biol. 1989, 5, 453–481. [Google Scholar] [CrossRef]
- Sakai, T.; Yamashina, S.; Ohnishi, S. Microtubule-disrupting drugs blocked delivery of endocytosed transferrin to the cytocenter, but did not affect return of transferrin to plasma membrane. J. Biochem. 1991, 109, 528–533. [Google Scholar]
- Menard, A.; Altendorf, K.D.; Berves, D.D.; Mock, M.; Montecucco, C. The vacuolar ATPase proton pump is required for the cytotoxicity of Bacillus anthracis lethal toxin. FEBS Lett. 1996, 386, 161–164. [Google Scholar] [CrossRef]
- Werner, G.; Hagenmaier, H.; Drautz, H.; Baumgartner, A.; Zahner, H. Metabolic products of microorganisms. 224. Bafilomycins, a new group of macrolide antibiotics. Production, isolation, chemical structure and biological activity. J. Antibiotics 1984, 37, 110–117. [Google Scholar]
- Lord, J.M.; Smith, D.C.; Roberts, L.M. Toxin entry: How bacterial proteins get into mammalian cells. Cell. Microbiol. 1999, 1, 85–91. [Google Scholar] [CrossRef]
- Haug, G.; Leemhuis, J.; Tiemann, D.; Meyer, D.K.; Aktories, K.; Barth, H. The host cell chaperone Hsp90 is essential for translocation of the binary Clostridium botulinum C2 toxin into the cytosol. J. Biol. Chem. 2003, 278, 32266–32274. [Google Scholar] [CrossRef]
- Kaiser, E.; Pust, S.; Kroll, C.; Barth, H. Cyclophilin A facilitates translocation of the Clostridium botulinum C2 toxin across membranes of acidified endosomes into the cytosol of mammalian cells. Cell. Microbiol. 2009, 11, 780–795. [Google Scholar] [CrossRef]
- Kaiser, E.; Kroll, C.; Ernst, K.; Schwan, C.; Popoff, M.; Fischer, G.; Buchner, J.; Aktories, K.; Barth, H. Membrane translocation of binary actin-ADP-ribosylating toxins from Clostridium difficile and Clostridium perfringens is facilitated by cyclophilin A and Hsp90. Infect. Immun. 2011, 79, 3913–3921. [Google Scholar] [CrossRef]
- Lang, A.E.; Ernst, K.; Lee, H.; Papatheodorou, P.; Schwan, C.; Barth, H.; Aktories, K. The chaperone Hsp90 and PPIases of the cyclophilin and FKBP families facilitate membrane translocation of Photorhabdus luminescens ADP-ribosyltransferases. Cell. Microbiol. 2014, 16, 490–503. [Google Scholar] [CrossRef]
- Kaiser, E.; Bohm, N.; Ernst, K.; Langer, S.; Schwan, C.; Aktories, K.; Popoff, M.; Fischer, G.; Barth, H. FK506-binding protein 51 interacts with Clostridium botulinum C2 toxin and FK506 inhibits membrane translocation of the toxin in mammalian cells. Cell. Microbiol. 2012, 14, 1193–1205. [Google Scholar] [CrossRef]
- Pratt, W.B.; Toft, D.O. Regulation of signaling protein function and trafficking by the hsp90/hsp70-based chaperone machinery. Exp. Biol. Med. 2003, 228, 111–133. [Google Scholar]
- Falnes, P.O.; Choe, S.; Madhus, I.H.; Wilson, B.A.; Olsnes, S. Inhibition of membrane translocation of diphtheria toxin A-fragment by internal disulfide bridges. J. Biol. Chem. 1994, 296, 8402–8407. [Google Scholar]
- Ratts, R.; Zeng, H.; Berg, E.A.; Blue, C.; McComb, M.E.; Costello, C.E.; vanderSpek, J.C.; Murphy, J.R. The cytosolic entry of diphtheria toxin catalytic domain requires a host cell cytosolic translocation factor complex. J. Cell Biol. 2003, 160, 1139–1150. [Google Scholar] [CrossRef]
- Kistner, A.; Habermann, E. Reductive cleavage of tetanus toxin and botulinum neurotoxin A by the thioredoxin system from brain. Evidence for two redox isomers of tetanus toxin. Naunyn Schmiedebergs Arch. Pharmacol. 1992, 345, 227–234. [Google Scholar]
- Reuner, K.H.; Presek, P.; Boschek, C.B.; Aktories, K. Botulinum C2 toxin ADP-ribosylates actin and disorganizes the microfilament network in intact cells. Eur. J. Cell Biol. 1987, 43, 134–140. [Google Scholar]
- Perelle, S.; Domenighini, M.; Popoff, M.R. Evidence that Arg-295, Glu-378, and Glu-380 are active-site residues of the ADP-ribosyltransferase activity of iota toxin. FEBS Lett. 1996, 395, 191–194. [Google Scholar] [CrossRef]
- Simpson, L.L.; Stiles, B.G.; Zepeda, H.; Wilkins, T.D. Production by Clostridium spiroforme of an iota-like toxin that possesses mono(ADP-ribosyl)transferase activity: Identification of a novel class of ADP-ribosyltransferases. Infect. Immun. 1989, 57, 255–261. [Google Scholar]
- Egelman, E.H. A tale of two polymers: New insights into helical filaments. Nat. Rev. Mol. Cell Biol. 2003, 4, 621–630. [Google Scholar] [CrossRef]
- Pollard, T.D.; Cooper, J.A. Actin, a central player in cell shape and movement. Science 2009, 326, 1208–1212. [Google Scholar] [CrossRef]
- Aktories, K.; Reuner, K.H.; Presek, P.; Barmann, M. Botulinum C2 toxin treatment increases the G-actin pool in intact chicken cells: A model for the cytopathic action of actin-ADP-ribosylating toxins. Toxicon 1989, 27, 989–993. [Google Scholar] [CrossRef]
- Just, I.; Wille, M.; Chaponnier, C.; Aktories, K. Gelsolin-actin complex is target for ADP-ribosylation by Clostridium botulinum C2 toxin in intact human neutrophils. Eur. J. Pharmacol. Mol. Pharmacol. 1993, 246, 293–297. [Google Scholar] [CrossRef]
- Just, I.; Hennessey, E.S.; Drummond, D.R.; Aktories, K.; Sparrow, J.C. ADP-ribosylation of Drosophila indirect-flight-muscle actin and arthrin by Clostridium botulinum C2 toxin and Clostridium perfringens iota toxin. Biochem. J. 1993, 291, 409–412. [Google Scholar]
- Vandekerckhove, J.; Schering, B.; Bärmann, M.; Aktories, K. Botulinum C2 toxin ADP-ribosylates cytoplasmic β/γ-actin in arginine 177. J. Biol. Chem. 1988, 263, 696–700. [Google Scholar]
- Mauss, S.; Chaponnier, C.; Just, I.; Aktories, K.; Gabbiani, G. ADP-ribosylation of actin isoforms by Clostridium botulinum C2 toxin and Clostridium perfringens iota toxin. Eur. J. Biochem. 1990, 194, 237–241. [Google Scholar]
- Prekeris, R.; Mayhew, M.W.; Cooper, J.B.; Terrian, D.M. Identification and localization of an actin-binding motif that is unique to the epsilon isoform of protein kinase C and participates in the regulation of synaptic function. J. Cell. Biol. 1996, 132, 77–90. [Google Scholar] [CrossRef]
- Wegner, A.; Aktories, K. ADP-ribosylated actin caps the barbed ends of actin filaments. J. Biol. Chem. 1988, 263, 13739–13742. [Google Scholar]
- Weigt, C.; Just, I.; Wegner, A.; Aktories, K. Nonmuscle actin ADP-ribosylated by botulinum C2 toxin caps actin filaments. FEBS Lett. 1989, 246, 181–184. [Google Scholar] [CrossRef]
- Al-Mohanna, F.A.; Ohishi, I.; Hallett, M.B. Botulinum C2 toxin potentiates activation of the neutrophil oxidase. Further evidence of a role for actin polymerization. FEBS Lett. 1987, 219, 40–44. [Google Scholar]
- Schwan, C.; Stecher, B.; Tzivelekidis, T.; van Ham, M.; Rohde, M.; Hardt, W.D.; Wehland, J.; Aktories, K. Clostridium difficile toxin CDT induces formation of microtubule-based protrusions and increases adherence of bacteria. PLoS Path. 2009, 5, e1000626. [Google Scholar] [CrossRef]
- Schwan, C.; Kruppke, A.S.; Nolke, T.; Schumacher, L.; Koch-Nolte, F.; Kudryashev, M.; Stahlberg, H.; Aktories, K. Clostridium difficile toxin CDT hijacks microtubule organization and reroutes vesicle traffic to increase pathogen adherence. Proc. Natl. Acad. Sci. USA 2014, 111, 2313–2318. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Stiles, B.G.; Pradhan, K.; Fleming, J.M.; Samy, R.P.; Barth, H.; Popoff, M.R. Clostridium and Bacillus Binary Enterotoxins: Bad for the Bowels, and Eukaryotic Being. Toxins 2014, 6, 2626-2656. https://doi.org/10.3390/toxins6092626
Stiles BG, Pradhan K, Fleming JM, Samy RP, Barth H, Popoff MR. Clostridium and Bacillus Binary Enterotoxins: Bad for the Bowels, and Eukaryotic Being. Toxins. 2014; 6(9):2626-2656. https://doi.org/10.3390/toxins6092626
Chicago/Turabian StyleStiles, Bradley G., Kisha Pradhan, Jodie M. Fleming, Ramar Perumal Samy, Holger Barth, and Michel R. Popoff. 2014. "Clostridium and Bacillus Binary Enterotoxins: Bad for the Bowels, and Eukaryotic Being" Toxins 6, no. 9: 2626-2656. https://doi.org/10.3390/toxins6092626
APA StyleStiles, B. G., Pradhan, K., Fleming, J. M., Samy, R. P., Barth, H., & Popoff, M. R. (2014). Clostridium and Bacillus Binary Enterotoxins: Bad for the Bowels, and Eukaryotic Being. Toxins, 6(9), 2626-2656. https://doi.org/10.3390/toxins6092626