
Veal Calves Produce Less Antibodies against C. Perfringens Alpha Toxin Compared to Beef Calves

Abstract
:1. Introduction
2. Results and Discussion
2.1. Study 1: Antibody Dynamics against Alpha Toxin and Perfringolysin in Veal and Beef Calves
2.1.1. Alpha Toxin Antibody Dynamics


{kind=link}
{kind=link}
{kind=link}
| Alpha toxin blocking ELISA | Mean % Inhibition of OD ± Standard Deviation | Seroprevalence ± Standard Deviation | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Age (in weeks) | Age (in weeks) | ||||||||
| Populations | n | 2 | 8 | 14 | 26 | 2 | 8 | 14 | 26 |
| Veal total | 443 | a43 ± 21 | b35 ± 17 | c30 ± 15 | c30 ± 15 | 46 ± 13 | 29 ± 14 | 17 ± 13 | 16 ± 9 |
| Veal cross | 167 | a45 ± 22 | b40 ± 18 | c35 ± 15 | c33 ± 15 | 49 ± 18 | 45 ± 11 | 28 ± 12 | 20 ± 12 |
| Veal HF | 121 | a40 ± 21 | b29 ± 12 | b23 ± 10 | b29 ± 17 | 43 ± 10 | 17 ± 4 | 6 ± 10 | 17 ± 8 |
| Veal BB | 155 | a42 * ± 20 | b34 * ± 17 | c29 * ± 24 | c26 * ± 13 | 45 ± 13 | 26 ± 10 | 16 ± 4 | 9 ± 6 |
| Beef BB | 85 | a51 * ± 23 | a52 * ± 16 | a55 * ± 13 | a53 * ± 12 | 66 ± 40 | 76 ± 28 | 91 ± 8 | 85 ± 14 |
| All calves | 528 | a44 ± 21 | b37 ± 18 | b32 ± 16 | b33 ± 17 | 44 ± 22 | 37 ± 18 | 32 ± 16 | 33 ± 17 |
2.1.2. Perfringolysin Antibody Dynamics

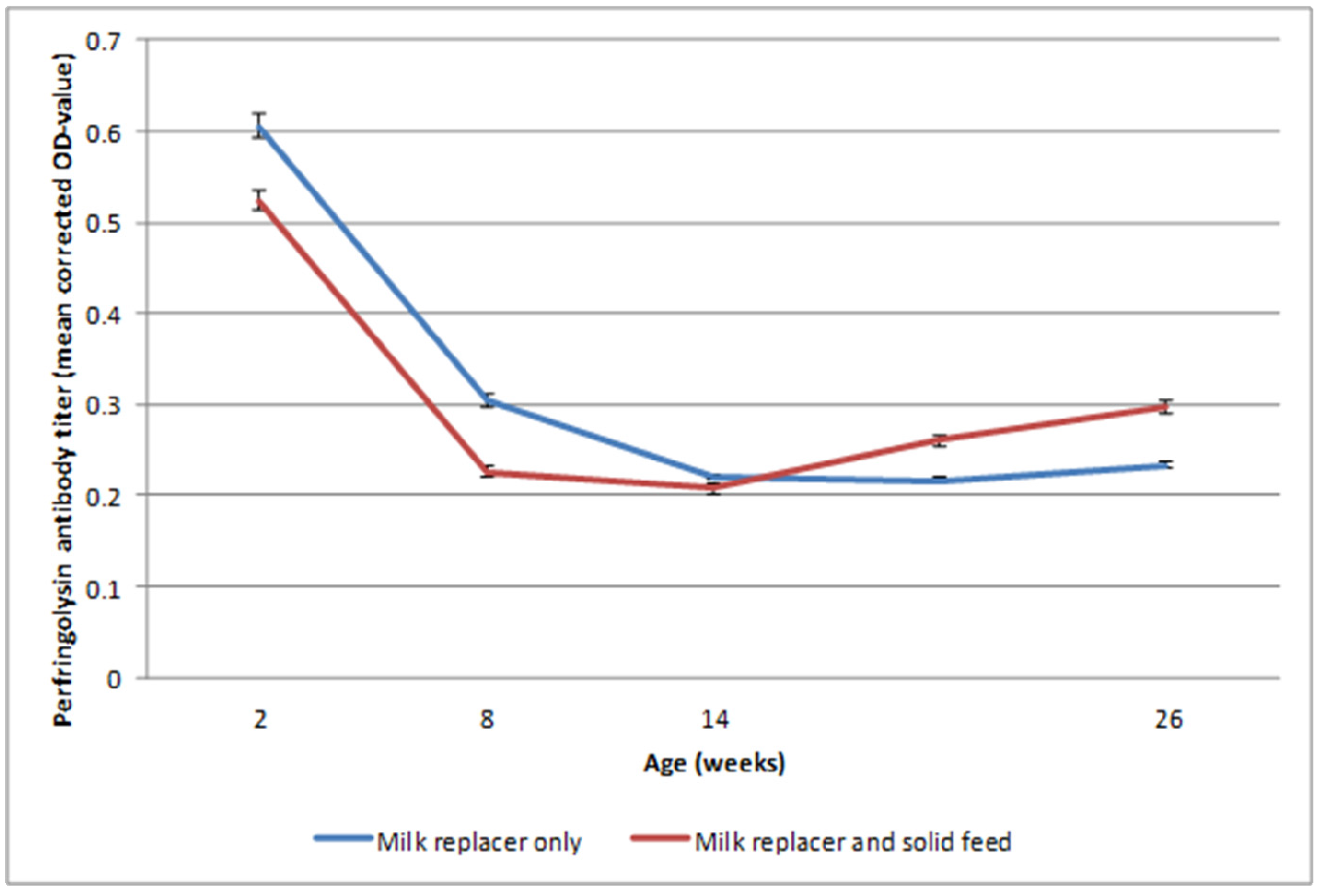
2.2. Study 2: Effect of Solid Feed Provision on Alpha Toxin and Perfringolysin Antibody Dynamics
2.2.1. Mortality, Incidence of Enterotoxaemia and Average Weight Gain
2.2.2. Alpha Toxin and Perfringolysin Antibody Dynamics
| Alpha Toxin Serology (% Inhibition of OD ± Standard Deviation) | ||||
|---|---|---|---|---|
| Group | Age in Weeks | |||
| 2 | 8 | 14 | 26 | |
| Group 1 | 72 ± 14 | 67 ± 11 | 67 ± 8 | 70 ± 5 |
| Group 2 | 64 ± 13 | 66 ± 8 | 68 ± 6 | 67 ± 5 |
| Total | 68 ± 14 | 66 ± 10 | 68 ± 7 | 69 ± 5 |

2.3. Discussion
3. Experimental Section
3.1. Study 1: Dynamics of Antibodies against Alpha Toxin and Perfringolysin in Veal and Beef Calves
3.1.1. Animals
| Herd | Number of Calves in Study | Production System | Breed | Floor | Feed | |||
|---|---|---|---|---|---|---|---|---|
| Milk Replacer | ||||||||
| % NP | % SMP | Weaning | Solid Feed | |||||
| 1 | 25 | Veal | HF | Slatted floor | 95 | 5 | No | <200 g/d |
| 2 | 25 | Veal | HF | Slatted floor | 95 | 5 | No | <200 g/d |
| 3 | 25 | Veal | HFXBB | Slatted floor | 95 | 5 | No | <200 g/d |
| 4 | 25 | Veal | BB | Slatted floor | 30 | 70 | No | <200 g/d |
| 5 | 25 | Veal | HFXBB | Slatted floor | 95 | 5 | No | <200 g/d |
| 6 | 25 | Veal | BB | Slatted floor | 30 | 70 | No | <200 g/d |
| 7 | 25 | Veal | HF | Slatted floor | 95 | 5 | No | <200 g/d |
| 8 | 26 | Veal | HF | Slatted floor | 95 | 5 | No | <200 g/d |
| 9 | 25 | Veal | HF | Slatted floor | 95 | 5 | No | <200 g/d |
| 10 | 24 | Veal | BB | Slatted floor | 30 | 70 | No | <200 g/d |
| 11 | 30 | Veal | BB | Slatted floor | 30 | 70 | No | <200 g/d |
| 12 | 39 | Veal | HFXBB | Slatted floor | 95 | 5 | No | <200 g/d |
| 13 | 38 | Veal | BB | Slatted floor | 30 | 70 | No | <200 g/d |
| 14 | 41 | Veal | HFXBB | Slatted floor | 95 | 5 | No | <200 g/d |
| 15 | 39 | Veal | HFXBB | Slatted floor | 95 | 5 | No | <200 g/d |
| 16 | 21 | Beef | BB | Straw bedded | 0 | 100 | Yes | Ad lib |
| 17 | 20 | Beef | BB | Straw bedded | 0 | 100 | Yes | Ad lib |
| 18 | 27 | Beef | BB | Straw bedded | 0 | 100 | Yes | Ad lib |
| 19 | 23 | Beef | BB | Straw bedded | 0 | 100 | Yes | Ad lib |
3.1.2. Sampling
3.1.3. Antibody Determination
Alpha Toxin Antibody ELISA
Expression and Purification of Perfringolysin
Perfringolysin Antibody ELISA
3.1.4. Statistics
3.2. Study 2: Effect of Solid Feed Intake on Alpha Toxin and Perfringolysin Antibody Dynamics
3.2.1. Study Design
3.2.2. Animals, Housing and Feeding
3.2.3. Laboratory Analysis
3.2.4. Statistics
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Valgaeren, B.; Pardon, B.; Goossens, E.; Verherstraeten, S.; Schauvliege, S.; Timbermont, L.; Ducatelle, R.; Deprez, P.; Van Immerseel, F. Lesion Development in a New Intestinal Loop Model Indicates the Involvement of a Shared Clostridium perfringens Virulence Factor in Haemorrhagic Enteritis in Calves. J. Comp. Pathol. 2013, 149, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Verherstraeten, S.; Goossens, E.; Valgaeren, B.R.; Pardon, B.; Timbermont, L.; Vermeulen, K.; Schauvliege, S.; Haesebrouck, F.; Ducatelle, R.; Deprez, P.; et al. The synergistic necrohemorrhagic action of Clostridium perfringens perfringolysin and alpha toxin in the bovine intestine and against bovine endothelial cells. Vet. Res. 2013, 44, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manteca, C.; Daube, G.; Mainil, J. Study of the role of Clostridium perfringens in bovine enterotoxaemia. Bull. Mem. Med. Bel. Belg. 2001, 154, 319–325. [Google Scholar]
- Pardon, B.; De Bleecker, K.; Hostens, M.; Callens, J.; Dewulf, J.; Deprez, P. Longitudinal study on morbidity and mortality in white veal calves in Belgium. BMC Vet. Res. 2012, 8, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griner, L.A.; Bracken, F.K. Clostridium perfringens (type C) in acute hemorrhagic enteritis of calves. J. Am. Vet. Med. Assoc. 1953, 122, 99–102. [Google Scholar] [PubMed]
- Niilo, L.; Harris, W.N.; Jones, G.A. Clostridium perfringens type C in hemorrhagic enterotoxaemia in neonatal calves in Alberta. Can. Vet. J. 1974, 15, 224–226. [Google Scholar] [PubMed]
- Manteca, C.; Daube, G. Etude de l’entérotoxémie bovine en Belgique I-Introduction et context bibliographique. Ann. Med. Vet. 1994, 138, 155–164. [Google Scholar]
- Pardon, B.; Catry, B.; Boone, R.; Theys, H.; De Bleecker, K.; Dewulf, J.; Deprez, P. Characteristics and challenges of the modern Belgian veal industry. Vlaams Diergeneeskd. Tijdschr. 2014, 83, 155. [Google Scholar]
- Schofield, F.W. Enterotoxemia (sudden death) in calves due to Clostridium welchii. J. Am. Vet. Med. Assoc. 1955, 126, 192–194. [Google Scholar] [PubMed]
- Griesemer, R.A.; Krill, W.R. Enterotoxemia in beef calves—30 years’ observation. J. Am. Vet. Med. Assoc. 1962, 140, 154–158. [Google Scholar] [PubMed]
- Uzal, F.A.; Kelly, W.R. Enterotoxaemia in goats. Vet. Res. Commun. 1996, 20, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Berghaus, R.D.; McCluskey, B.J.; Callan, R.J. Risk factors associated with hemorrhagic bowel syndrome in dairy cattle. J. Am. Vet. Med. Assoc. 2005, 226, 1700–1706. [Google Scholar] [CrossRef] [PubMed]
- Timbermont, L.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Necrotic enteritis in broilers: An updated review on the pathogenesis. Avian Pathol. 2011, 40, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Veschi, J.L.; Bruzzone, O.A.; Losada-Eaton, D.M.; Dutra, I.S.; Fernandez-Miyakawa, M.E. Naturally acquired antibodies against Clostridium perfringens epsilon toxin in goats. Vet. Immunol. Immunopathol. 2008, 125, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Troxel, T.R.; Burke, G.L.; Wallace, W.T.; Keaton, L.W.; McPeake, S.R.; Smith, D.; Nicholson, I. Clostridial vaccination efficacy on stimulating and maintaining an immune response in beef cows and calves. J. Anim. Sci. 2001, 75, 19–25. [Google Scholar]
- Heier, B.T.; Lovland, A.; Soleim, K.B.; Kaldhusdal, M.; Jarp, J. A field study of naturally occurring specific antibodies against Clostridium perfringens alpha toxin in Norwegian broiler flocks. Avian Dis. 2001, 45, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Dastur, F.D.; Awatramani, V.P.; Dixit, S.K.; D’Sa, J.A.; Cooverji, N.D.; Anand, M.P. Response to single dose of tetanus vaccine in subjects with naturally acquired tetanus antitoxin. Lancet 1981, 2, 219–222. [Google Scholar] [CrossRef]
- Ehrengut, W.; Sarateanu, D.E.; AgRhagly, A.; Koumaré, B.; Simaga, S.Y.; Diallo, D. Naturally acquired tetanus antitoxin in the serum of children in Mali. Immun. Infekt. 1983, 11, 229–232. [Google Scholar] [PubMed]
- Veronesi, R.; Bizzini, B.; Focaccia, R.; Coscina, A.L.; Mazza, C.C.; Focaccia, M.T.; Carraro, F.; Honningman, M.N. Naturally acquired antibodies to tetanus toxin in humans and animals from the galapos islands. J. Infect. Dis. 1983, 147, 308–311. [Google Scholar] [CrossRef] [PubMed]
- Matzin, H.; Regev, S. Naturally acquired immunity to tetanus toxin in an isolated community. Infect. Immun. 1985, 48, 267–268. [Google Scholar]
- Haghighi, H.R.; Gong, J.; Gyles, C.L.; Hayes, M.A.; Zhou, H.; Sanei, B.; Chambers, J.R.; Sharif, S. Probiotics stimulate production of natural antibodies in chickens. Clin. Vaccine Immunol. 2006, 13, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.H.; Hendee, E.D.; Pappenheimer, A.M. Factors involved in the production of Clostridium Welchii alpha toxin. J. Exp. Med. 1947, 85, 701–713. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valgaeren, B.R.; Pardon, B.; Goossens, E.; Verherstraeten, S.; Roelandt, S.; Timbermont, L.; Vekens, N.V.D.; Stuyvaert, S.; Gille, L.; Van Driessche, L.; et al. Veal Calves Produce Less Antibodies against C. Perfringens Alpha Toxin Compared to Beef Calves. Toxins 2015, 7, 2586-2597. https://doi.org/10.3390/toxins7072586
Valgaeren BR, Pardon B, Goossens E, Verherstraeten S, Roelandt S, Timbermont L, Vekens NVD, Stuyvaert S, Gille L, Van Driessche L, et al. Veal Calves Produce Less Antibodies against C. Perfringens Alpha Toxin Compared to Beef Calves. Toxins. 2015; 7(7):2586-2597. https://doi.org/10.3390/toxins7072586
Chicago/Turabian StyleValgaeren, Bonnie R., Bart Pardon, Evy Goossens, Stefanie Verherstraeten, Sophie Roelandt, Leen Timbermont, Nicky Van Der Vekens, Sabrina Stuyvaert, Linde Gille, Laura Van Driessche, and et al. 2015. "Veal Calves Produce Less Antibodies against C. Perfringens Alpha Toxin Compared to Beef Calves" Toxins 7, no. 7: 2586-2597. https://doi.org/10.3390/toxins7072586
APA StyleValgaeren, B. R., Pardon, B., Goossens, E., Verherstraeten, S., Roelandt, S., Timbermont, L., Vekens, N. V. D., Stuyvaert, S., Gille, L., Van Driessche, L., Haesebrouck, F., Ducatelle, R., Van Immerseel, F., & Deprez, P. (2015). Veal Calves Produce Less Antibodies against C. Perfringens Alpha Toxin Compared to Beef Calves. Toxins, 7(7), 2586-2597. https://doi.org/10.3390/toxins7072586