
Doses of Immunogen Contribute to Specificity Spectrums of Antibodies against Aflatoxin



 ,
, 
Abstract
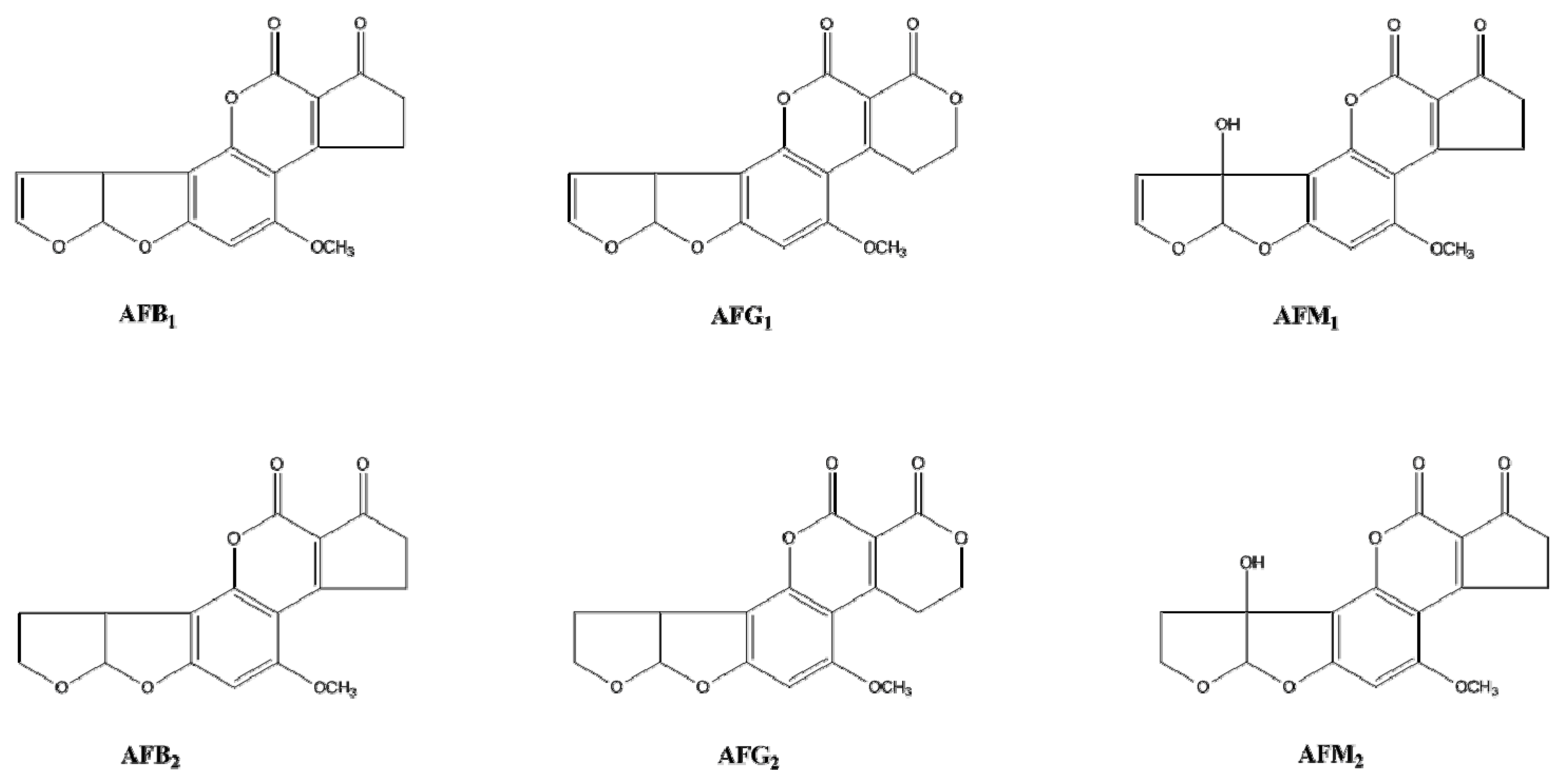
:1. Introduction
2. Results and Discussion
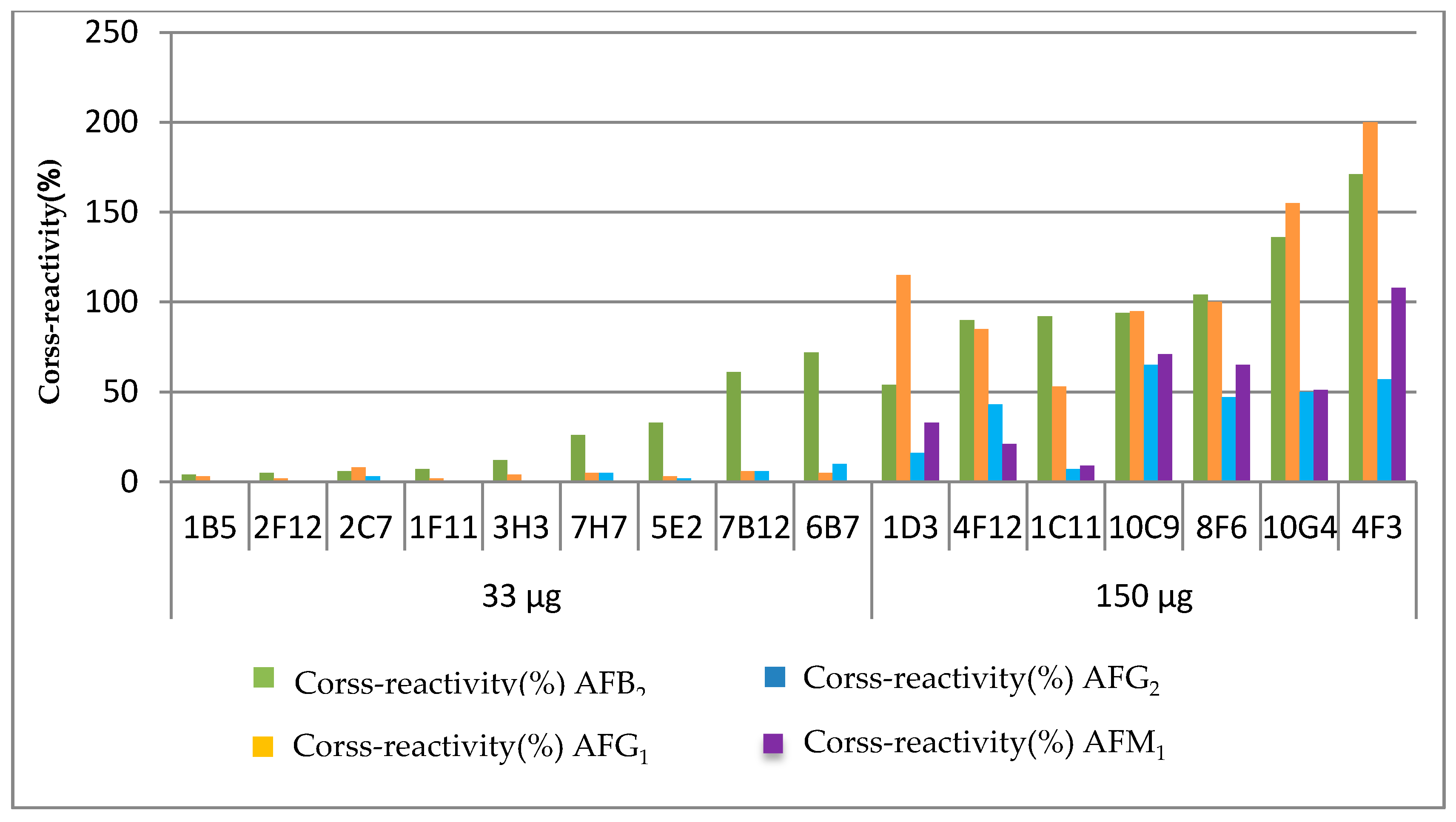
2.1. The Influence on the Dose of the Immunogen against AFB1
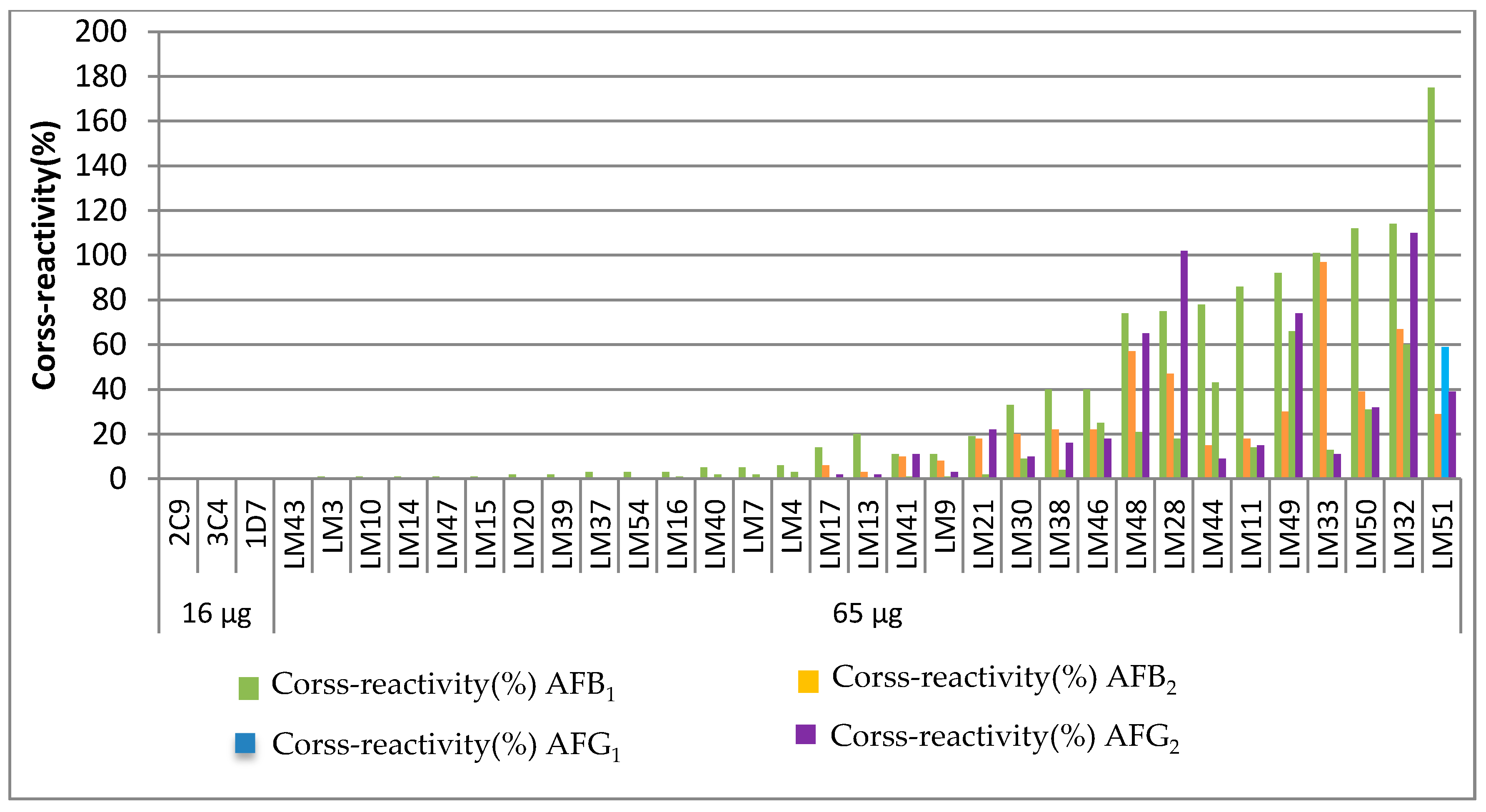
2.2. The Influence on the Dose of the Immunogen against AFM1
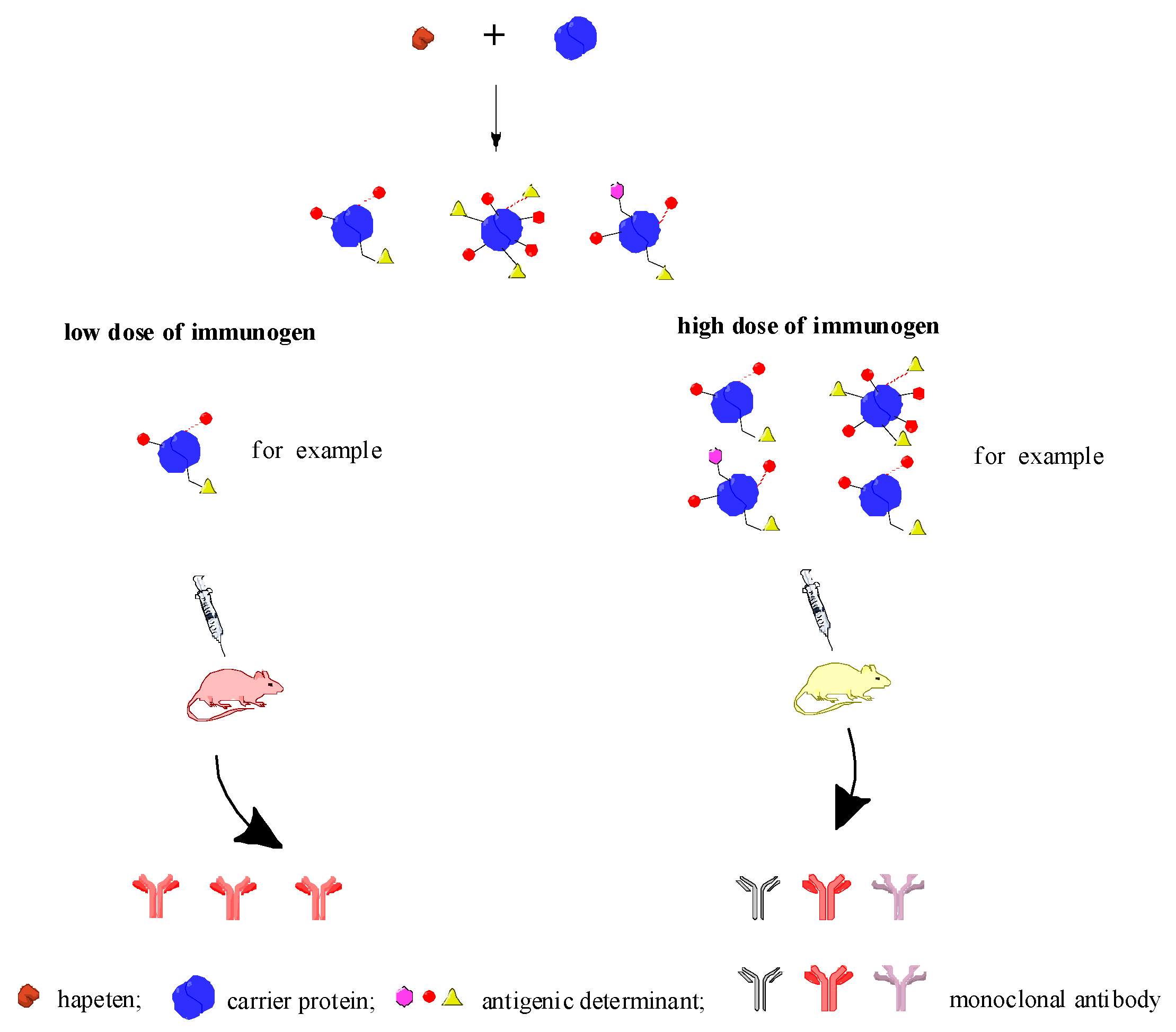
2.3. Discussion
3. Conclusions
4. Experimental Section
4.1. Chemicals and Instruments
4.2. Immunization
4.3. Production of mAbs
4.4. Evaluation of Antibody Sensitivity and Cross-Reactivity
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, S.T.; Gui, W.J.; Guo, Y.R.; Zhu, G.N. Preparation of a multihapten antigen and broad specificity polyclonal antibodies for a multiple pesticide immunoassay. Anal. Chim. Acta 2007, 587, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhu, Y.; Ding, S.; He, F.; Beier, R.C.; Li, J.; Jiang, H.; Feng, C.; Wan, Y.; Zhang, S.; et al. Development of a monoclonal antibody-based broad-specificity ELISA for fluoroquinolone antibiotics in foods and molecular modeling studies of cross-reactive compounds. Anal. Chem. 2007, 79, 4471–4483. [Google Scholar] [CrossRef] [PubMed]
- Chu, F.S.; Steinert, B.W.; Gaur, P.K. Production and characterization of antibody against aflatoxin G1. J. Food Saf. 1985, 7, 161–170. [Google Scholar] [CrossRef]
- Albrecht, R. Development of antibacterial agents of the nalidixic acid type. Drug Res. 1977, 21, 91–104. [Google Scholar]
- Brimfield, A.A.; Lenz, D.E.; Graham, C.; Hunter, K.W. Mouse monoclonal antibodies against paraoxon: Potential reagents for immunoassay with constant immunochemical characteristics. J. Agric. Food Chem. 1985, 33, 1237–1242. [Google Scholar] [CrossRef]
- Bucknall, S.; Silverlight, N.J.; Coldham, L.; Thorne, R. Antibodies to the quinolones and fluoroquinolones for the development of generic and specific immunoassays for detection of these residues in animal products. Food Addit. Contam. 2003, 20, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.L.; Shen, Y.D.; Zheng, W.X.; Beier, R.C.; Xie, G.M.; Dong, J.X.; Yang, J.Y.; Wang, H.; Lei, H.T.; She, Z.G.; et al. Broad-specificity immunoassay for O,O-diethyl organophosphorus pesticides: Application of molecular modeling to improve assay sensitivity and study antibody recognition. Anal. Chem. 2010, 82, 9314–9321. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Ihara, Y.; Nakata, E.; Miyazawa, M.; Sasaki, M.; Kodaira, T.; Nakazawa, H. Development of enrofloxacin ELISA using a monoclonal antibody tolerating an organic solvent with broad cross-reactivity to other new quinolones. Food Agric. Immunol. 2007, 18, 179–187. [Google Scholar] [CrossRef]
- Anklam, E.; Stroka, J.; Boenke, A. Acceptance of analytical methods for implementation of EU legislation with a focus on mycotoxins. Food Control 2002, 13, 173–183. [Google Scholar] [CrossRef]
- Ma, F.; Wu, R.; Li, P.W.; Yu, L. Analytical approaches for measuring pesticides, mycotoxins and heavy metals in vegetable oils: A review. Eur. J. Lipid Sci. Technol. 2016, 118, 339–352. [Google Scholar] [CrossRef]
- Zhang, D.H.; Li, P.W.; Zhang, Q.; Yang, Y.; Zhang, W.; Guan, D.; Ding, X.X. Extract-free immunochromatographic assay for on-site tests of aflatoxin M1 in milk. Anal. Methods 2012, 4, 3307–3313. [Google Scholar] [CrossRef]
- Xu, Z.L.; Xie, G.M.; Li, Y.H.; Wang, B.F.; Lei, H.L.; Wang, H.; Shen, Y.D.; Sun, Y.M. Production and characterization of a broad-specificity polyclonal antibody for O,O-diethyl organophosphorus pesticides and a quantitative structure-activity relationship study of antibody recognition. Anal. Chim. Acta 2009, 647, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Anfossi, L.; Calderara, M.; Baggiani, C.; Giovannoli, C.; Arletti, E.; Giraudi, G. Development and application of solvent-free extraction for the detection of aflatoxin M1 in dairy products by enzyme immunoassay. J. Agric. Food Chem. 2008, 56, 1852–1857. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Kong, D.; Sui, J.; Jiang, T.; Li, J.; Ma, L.; Lin, H. Broad-specific antibodies for a generic immunoassay of quinolone: Development of a molecular model for selection of haptens based on molecular field-overlapping. Anal. Chem. 2009, 81, 3246–3251. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.P.; Chen, J.H.; Zhang, C.J.; Huang, X.A.; Sun, Y.M.; Xu, Z.L.; Lei, H.T. Broad-Specificity chemiluminescence enzyme immunoassay for (fluoro)quinolones: Hapten design and molecular modeling study of antibody recognition. Anal. Chem. 2016, 88, 3909–3916. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Liu, B.; Liu, E.M.; Sheng, W.; Zhang, Y.; Crossan, A.; Kennedy, I.; Wang, S. Immunoassay for phenylurea herbicides: Application of molecular modeling and quantitative structure-activity relationship analysis on an antigen-antibody interaction study. Anal. Chem. 2011, 83, 4767–4774. [Google Scholar] [CrossRef] [PubMed]
- Aly, S.E. Distrubution of aflatoxins in product and by-products during glucose production from contaminated corn. Nahrung 2002, 46, 341–344. [Google Scholar] [CrossRef]
- Gacem, M.A.; El-Hadj-Khelil, A.O. Toxicology, biosynthesis, bio-control of aflatoxin and new methods of detection. Asian Pac. J. Trop. Biomed. 2016, 9, 808–814. [Google Scholar] [CrossRef]
- Horn, P.R.; Mao, Y.; Head-Gordon, M. Defining the contributions of permanent electrostatics, Pauli repulsion, and dispersion in density functional theory calculations of intermolecular interaction energies. J. Chem. Phys. 2016, 144, 114107. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.M.; Pennington, J.E.; Kubler, A.M.; Conscience, J.F. A simple, single-step technique for selecting and cloning hybridomas for the production of monoclonal antibodies. J. Immunol. Methods 1982, 50, 161–171. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Li, P.W.; Zhang, W.; Zhang, Q.; Ding, X.X.; Chen, X.M. Production and Characterization of Monoclonal Antibodies against Aflatoxin G1. Hybridoma 2009, 28, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Holtzapple, C.K.; Buckley, S.; Stanker, L.H. Production and characterization of monoclonal antibodies against sarafloxacin and cross-reactivity studies of related fluoroquinolones. J. Agric. Food Chem. 1997, 45, 1984–1990. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.L.; Pan, Y.H. Preparation of a generic monoclonal antibody and development of a highly sensitive indirect competitive ELISA for the detection of phenothiazines in animal feed. Food Chem. 2017, 221, 1004–1013. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dose of Immunogen | mAb | IC50 for AFB1 (ng mL−1) | Cross-Reactivity (%) | ||||
|---|---|---|---|---|---|---|---|
| (µg per Balb/c Mice) | AFB1 | AFB2 | AFG1 | AFG2 | AFM1 | ||
| 33 | 1B5 | 0.012 | 100 | 4 | 3 | <0.1 | <0.1 |
| 33 | 2F12 | 0.010 | 100 | 5 | 2 | 0.2 | <0.1 |
| 33 | 2C7 | 0.020 | 100 | 6 | 8 | 3 | <0.1 |
| 33 | 1F11 | 0.052 | 100 | 7 | 2 | <0.1 | <0.1 |
| 33 | 3H3 | 0.023 | 100 | 12 | 4 | <1 | <0.1 |
| 33 | 7H7 | 0.052 | 100 | 26 | 5 | 5 | <0.1 |
| 33 | 5E2 | 0.013 | 100 | 33 | 3 | 2 | <0.1 |
| 33 | 7B12 | 0.012 | 100 | 61 | 6 | 6 | <0.1 |
| 33 | 6B7 | 0.027 | 100 | 72 | 5 | 10 | <0.1 |
| 150 | 1D3 | 0.44 | 100 | 54 | 115 | 16 | 33 |
| 150 | 4F12 | 0.086 | 100 | 90 | 85 | 43 | 21 |
| 150 | 1C11 | 0.001 | 100 | 92 | 53 | 7 | 9 |
| 150 | 10C9 | 2.09 | 100 | 94 | 95 | 65 | 71 |
| 150 | 8F6 | 1.70 | 100 | 104 | 100 | 47 | 65 |
| 150 | 10G4 | 0.73 | 100 | 136 | 155 | 50 | 51 |
| 150 | 4F3 | 0.29 | 100 | 171 | 200 | 57 | 108 |
| Dose of Immunogen (µg per Balb/c Mice) | mAb | IC50 for AFM1 (ng mL−1) | Cross-Reactivity (%) | ||||
|---|---|---|---|---|---|---|---|
| AFM1 | AFB1 | AFB2 | AFG1 | AFG2 | |||
| 16 | 2C9 | 0.067 | 100 | <1 | <1 | <1 | <1 |
| 16 | 3C4 | 0.043 | 100 | <1 | <1 | <1 | <1 |
| 16 | 1D7 | 0.058 | 100 | <1 | <1 | <1 | <1 |
| 65 | LM43 | 0.014 | 100 | <1 | <1 | <1 | <1 |
| 65 | LM3 | 0.029 | 100 | 1 | <1 | <1 | <1 |
| 65 | LM10 | 0.034 | 100 | 1 | <1 | <1 | <1 |
| 65 | LM14 | 0.017 | 100 | 1 | <1 | <1 | <1 |
| 65 | LM47 | 0.020 | 100 | 1 | <1 | <1 | <1 |
| 65 | LM15 | 0.006 | 100 | 1 | <1 | <1 | <1 |
| 65 | LM20 | 0.011 | 100 | 2 | <1 | <1 | <1 |
| 65 | LM39 | 0.069 | 100 | 2 | <1 | <1 | <1 |
| 65 | LM37 | 0.017 | 100 | 3 | <1 | <1 | <1 |
| 65 | LM54 | 0.029 | 100 | 3 | <1 | <1 | <1 |
| 65 | LM16 | 0.030 | 100 | 3 | <1 | 1 | <1 |
| 65 | LM40 | 0.029 | 100 | 5 | <1 | 2 | <1 |
| 65 | LM7 | 0.052 | 100 | 5 | <1 | 2 | <1 |
| 65 | LM4 | 0.014 | 100 | 6 | <1 | 3 | <1 |
| 65 | LM17 | 0.069 | 100 | 14 | 6 | <1 | 2 |
| 65 | LM13 | 0.011 | 100 | 20 | 3 | <1 | 2 |
| 65 | LM41 | 0.015 | 100 | 11 | 10 | 1 | 11 |
| 65 | LM9 | 0.014 | 100 | 11 | 8 | 1 | 3 |
| 65 | LM21 | 0.011 | 100 | 19 | 18 | 2 | 22 |
| 65 | LM30 | 0.023 | 100 | 33 | 20 | 9 | 10 |
| 65 | LM38 | 0.014 | 100 | 40 | 22 | 4 | 16 |
| 65 | LM46 | 0.011 | 100 | 40 | 22 | 25 | 18 |
| 65 | LM48 | 0.012 | 100 | 74 | 57 | 21 | 65 |
| 65 | LM28 | 0.012 | 100 | 75 | 47 | 18 | 102 |
| 65 | LM44 | 0.019 | 100 | 78 | 15 | 43 | 9 |
| 65 | LM11 | 0.019 | 100 | 86 | 18 | 14 | 15 |
| 65 | LM49 | 0.019 | 100 | 92 | 30 | 66 | 74 |
| 65 | LM33 | 0.013 | 100 | 101 | 97 | 13 | 11 |
| 65 | LM50 | 0.035 | 100 | 112 | 39 | 31 | 32 |
| 65 | LM32 | 0.046 | 100 | 114 | 67 | 60 | 110 |
| 65 | LM51 | 0.023 | 100 | 175 | 29 | 59 | 39 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Wu, J.; Zhang, L.; Fan, Z.; Yu, T.; Jiang, F.; Tang, X.; Zhang, Z.; Zhang, W.; Zhang, Q. Doses of Immunogen Contribute to Specificity Spectrums of Antibodies against Aflatoxin. Toxins 2017, 9, 172. https://doi.org/10.3390/toxins9050172
Li P, Wu J, Zhang L, Fan Z, Yu T, Jiang F, Tang X, Zhang Z, Zhang W, Zhang Q. Doses of Immunogen Contribute to Specificity Spectrums of Antibodies against Aflatoxin. Toxins. 2017; 9(5):172. https://doi.org/10.3390/toxins9050172
Chicago/Turabian StyleLi, Peiwu, Jing Wu, Li Zhang, Zhiyong Fan, Tingting Yu, Feng Jiang, Xiaoqian Tang, Zhaowei Zhang, Wen Zhang, and Qi Zhang. 2017. "Doses of Immunogen Contribute to Specificity Spectrums of Antibodies against Aflatoxin" Toxins 9, no. 5: 172. https://doi.org/10.3390/toxins9050172
APA StyleLi, P., Wu, J., Zhang, L., Fan, Z., Yu, T., Jiang, F., Tang, X., Zhang, Z., Zhang, W., & Zhang, Q. (2017). Doses of Immunogen Contribute to Specificity Spectrums of Antibodies against Aflatoxin. Toxins, 9(5), 172. https://doi.org/10.3390/toxins9050172