Abstract
This study was designed to investigate the structure of synthesized hesperidin glycosides (HGs) and evaluate their antibacterial and α-glucosidase inhibitory activities. The preliminary structure of HGs was confirmed by glucoamylase treatment and analyzed on thin layer chromatography (TLC). The LC-MS/MS profiles of HGs showed the important fragments at m/z ratios of 345.21 (added glucose to glucose of rutinose in HG1) and 687.28 (added maltose to glucose of rutinose in HG2), confirming that the structures of HG1 and HG2 were α-glucosyl hesperidin and α-maltosyl hesperidin, respectively. In addition, 1H and 13C-NMR of hesperidin derivatives were performed to identify their α-1,4-glycosidic bonds. The MIC and MBC studies showed that transglycosylated HG1 and HG2 had better antibacterial and bactericidal activities than hesperidin and diosmin, and were more active against Staphylococcus aureus than Escherichia coli. Hesperidin, HG1, HG2, and diosmin inhibited α-glucosidase with IC50 values of 2.75 ± 1.57, 2.48 ± 1.61, 2.36 ± 1.48, and 2.99 ± 1.23 mg/mL, respectively. The inhibition kinetics of HG2 shown by a Lineweaver–Burk plot confirmed HG2 was an α-glucosidase competitive inhibitor with an inhibitor constant, Ki, of 2.20 ± 0.10 mM. Thus, HGs have the potential to be developed into antibacterial drugs and treatments for treating α-glucosidase-associated type 2 diabetes.
1. Introduction
Flavonoids are a large group of polyphenolic compounds that are found in fruit, green leaves and vegetables. Flavonoids are characterized by the presence of 15 carbon atoms in a basic skeleton C6–C3–C6 phenyl-benzopyran structure. They can be found in two different forms: the free aglycone forms and combined sugar molecules as glycosides [1]. Flavonoids consist of six main structural classes: flavones, flavonols, flavanones, flavanonols, flavanols and anthocyanin [2]. Hesperidin, one of the most common flavonoids [3], is bitter and insoluble in water but can be sweetened and made water soluble by transglycosylation with saccharides [4,5,6]. Our previous work on the transglycosylation reaction of p19bBC recombinant cyclodextrin glycosyltransferase (CGTase, E.C.2.4.1.19) found that hesperidin is the best acceptor in the flavonoid group [5]. The transglycosylation of hesperidin using the p19bBC CGTase is able to produce large quantities of HGs (HG1–3) [5,6], but their glycoside structures are yet to be identified.
Hesperidin is widely used in clinical applications as an antioxidant [6,7], anti-inflammatory [8,9], anti-cancer [10,11,12], antibacterial [13,14] and antidiabetic [15]. Several research groups have recently investigated the possible antibacterial activity of hesperidin [13,16]. In plants, hesperidin protects against bacterial, fungal and viral pathogens. Its antibacterial activity depends on its molecular structure, hydrophobicity, solubility, presence or absence of a sugar unit, and the type of sugar in the chemical backbone [17]. By agar diffusion method, das Neves et al. [18] demonstrated that the ethanolic extract of the grapefruit seed and pulp contained hesperidin that inhibited only Gram-positive bacteria; however, this extract was effective against both Gram-positive and Gram-negative bacteria using the broth dilution susceptibility test [19]. Yi et al. [14] evaluated the antibacterial effects of a flavonoid extract of Pericarpium Citri Reticulatae (FEPCR) and its major constituents, including hesperidin, against Escherichia coli, Staphylococcus aureus, Staphylococcus epidermidis, Enterococcus faecalis, Salmonella typhimurium and Enterobacter cloacae using the agar dilution method. This study revealed that hesperidin and FEPCR had a wide range of antibacterial activity that was greater against the Gram-positive than the Gram-negative bacteria. The lowest and highest minimal inhibitory concentrations (MICs) were observed for S. aureus (MIC: 100 µg/mL) and E. cloacae (MIC: 1600 µg/mL), respectively. Given the growing challenge of antibacterial resistance, the search for new drugs should include natural compounds such as medicinal plants.
Hesperidin also inhibits α-glucosidase and decreases the concentration of blood glucose, a property that could be used to treat diabetes mellitus (DM). DM, a major global health challenge, is characterized by reduced insulin secretion and insulin receptor sensitivity resulting in chronic hyperglycaemia [15] and leads to the development of microvascular and macrovascular complications and increased morbidity and mortality [20]. Ahmed et al. [15] reported that hesperidin and naringin increased serum insulin concentrations and reduced serum glucose, blood glycosylated hemoglobin, and lipids in type 2 diabetic rats, consistent with the findings of Od-ek et al. [21], who showed that hesperidin partially inhibited α-glucosidase activity in vitro.
Our previous study on the small-scale biocatalytic production of HGs was performed using recombinant CGTase, derived from p19bBC E. coli cells [6]. This enzyme catalyzes four reactions: Cyclization, coupling, disproportionation and hydrolysis [5,6]. The main catalytic product from its coupling (intermolecular transglycosylation) is functional glycosides that are useful in the food, drug and cosmetic industries. The demand for using glycoside products in those industries has increased because they have better characteristics, such as solubility, stability and bioactivity, than their parent compounds.
This present work aims to use a recombinant CGTase for the upscale synthesis of HGs. We focused on identifying hesperidin and its derivative structures, and their antibacterial and α-glucosidase inhibitory activities. This is the first report that has attempted to elucidate the structure of glycosylated HG1–2 using LC-MS/MS and NMR analyses.
2. Results
2.1. Production and Purification of Hesperidin Glycoside
The reaction mixture for the synthesis of HGs from β-CD as a donor and hesperidin as an acceptor was performed following previously reported methods [5,6]. The synthesis of HGs was scaled-up to 300 mL, and the reaction proceeded under the conditions according to Section 4.2. HG products were then purified using preparative HPLC, since they were separated as three main products at Rt of 4.8 (HG3), 6.2 (HG2) and 7.7 (HG1) min, respectively (Figure 1). These products were lyophilized for further structural analysis and bioactivity determination. The conversion yield of HGs obtained in this study was found to be 38% (1.64 mM) from a 300 mL reaction mixture.
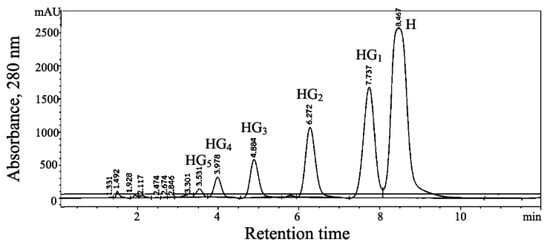
Figure 1.
Detection of hesperidin glycoside products using preparative HPLC chromatography profile of transglycosylation catalyzed using recombinant CGTase from p19bBC cells. The transglycosylation volume of synthesis was scaled up to 300 mL. The reaction mixture was filtered through a 0.45 µm nylon membrane before injecting into preparative HPLC and eluted with acetonitrile:water (20:80 v/v), using a flow rate of 20 mL/min. Hesperidin (H), hesperidin glucoside (HG1), hesperidin maltoside (HG2), hesperidin maltotrioside (HG3), hesperidin maltotetraoside (HG4), and hesperidin maltopentaoside (HG5).
2.2. Structural Analysis of Hesperidin Glycosides
2.2.1. Digestion with Amylolytic Enzyme
Preliminarily structural analysis of synthesized HGs was performed by enzyme treatment, and the final deglucosylated products were then investigated by TLC analysis.
When HG1, HG2 and HG3 were treated with Rhizopus sp. glucoamylase (EC 3.2.1.3), the hesperidin glycoside HG1, HG2 and HG3 spots disappeared while the intensities of hesperidin (the highest Rf value) and glucose spots increased (Figure 2, Lanes 4, 6 and 8). This finding suggested that the glycosidic linkage between the glucose units of HG1, HG2 and HG3 was hydrolyzed and that these glycosidic linkages were of an α-configuration. Thus, our results are consistent with HGs synthesizing of CGTase being specific for α-1,4-linkage transfer.
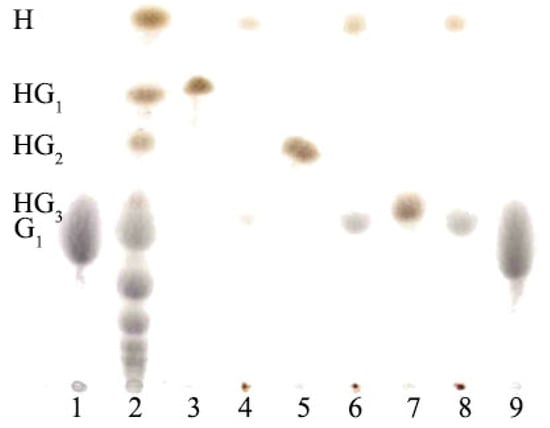
Figure 2.
TLC analysis of glucoamylase treatment of hesperidin glycosides (HGs). Lanes 1, 9 standard glucose (G1); Lane 2, a mixture of products from CGTase reaction for the synthesis of HGs; Lanes 3, 5, and 7, hesperidin glucoside (HG1), hesperidin maltoside (HG2) and hesperidin maltotrioside (HG3), respectively; Lanes 4, 6, and 8, HG1, HG2 and HG3 with glucoamylase treatment; H, hesperidin.
2.2.2. LC-MS/MS Analysis
After HGs were synthesized and purified, their structures were investigated. LC-MS/MS was used in combination with glucoamylase treatment and NMR to determine the possible structures of the obtained HGs by the reaction of recombinant CGTase [22,23]. The LC-MS/MS spectra of hesperidin and each purified HG (HG1 and HG2) were determined by the Triple Quadrupole analyzer. The detector and electrospray mass spectrometry (ESI-MS) were performed in full scan mode and tandem MSn fragmentations. The mass of each purified fraction from preparative HPLC was subjected to ESI-MS1 analysis. The products at Rt of 6.2 and 7.7 min were identified as hesperidin maltoside (HG2, hesperidin + maltose) and hesperidin glucoside (HG1, hesperidin + glucose), with masses [M + H]+ at m/z of 935.30 and 773.26, respectively. Standard hesperidin had a mass at m/z pf 611.09 [6]. The fragmentation patterns of secondary mass spectrometer (MS2) were performed to predict the position of glucose attached to original hesperetin and hesperidin (Figure 3). The results in Figure 3A show the major fragmentation patterns of hesperidin that had m/z of 303.09 and 327.29 are consistent with hesperetin (peak b) and rutinose sugar (peak c), respectively. The fragmentation patterns of HG1 product at Rt 7.7 min (Figure 3B) had important peaks at m/z corresponding to the mass of hesperetin (peak c) and the mass of glucose-glucose in the HG1 structure (m/z of 345.21, peak e). The HG2 product at Rt 6.2 min (Figure 3C) showed the mass of a sugar group linked between maltose and rutinose in the structure of HG2 (m/z of 687.28, peak f). In addition, the fragmentation pattern of HG2 still included the structure of hesperetin (m/z of 303.33, peak b). These results suggest there may be a link at 4″ OH position between the added glucose molecules with the one already attached to hesperetin.
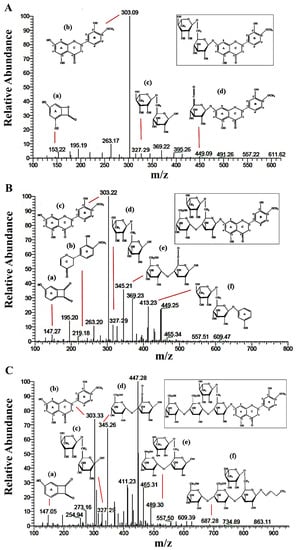
Figure 3.
LC-ESI-MS/MS analysis profile in positive mode ionization of the (A) hesperidin, (B) hesperidin glucoside (HG1) and (C) hesperidin maltoside (HG2).
2.2.3. NMR Analysis
The 1H- and 13C-NMR spectra of hesperetin, hesperidin and HG1–2 were determined.
According to 1H-NMR (Figure 4B–D), the peaks with doublet signals at 5.0–5.2 ppm (J = 3.66 Hz) were observed with a different height and assigned to the anomeric protons of each glucose, which linked to hesperidin with α-1,4-glycosidic bonds. In contrast, these peaks were not found in the 1H-NMR spectrum of hesperetin, as shown in Figure 4A.
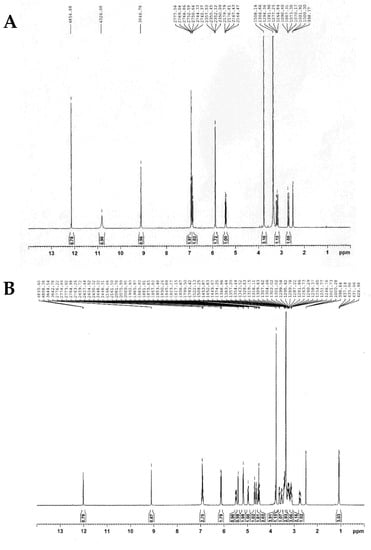
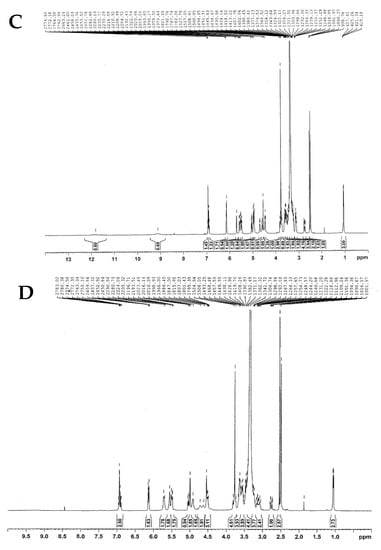
Figure 4.
The 400 MHz 1H-NMR spectra of (A) hesperetin, (B) hesperidin, (C) HG1, and (D) HG2.
In 13C-NMR spectra, carbon signals for glycoside products at Rt of 6.2 and 7.7 min were assigned to hesperidin glycosides. The peaks of HG1–2 with chemical shifts at 60–61 ppm (–CH2OH), 103–197 ppm (aromatic carbon), and 99–101 ppm (α-anomeric carbon of glucose) were observed (Table 1 and Figure S1). From the result, the α-configuration of the D-glucose residue in HG1 was identified by anomeric carbon of the glucose II signal (position C-1*) (Table 1 and Figure 5), which shifted from 92.77 to 101.25 ppm. In addition, comparing the position C-4″ of glucose I of hesperidin and HG1 indicated that the carbon signal of HG1 was deshielded from 70.27 to 79.72 ppm, corresponding to the α-1,4-glycosidic form. For the 13C-NMR spectrum of HG2, another α-1,4-glycosidic bond was confirmed by anomeric carbon of glucose III (the position C-1**), showing a carbon signal at 100.36 ppm. Moreover, position C-4* of HG2 was shifted down from 70.32 to 79.23 ppm compared to HG1. From the combination of these results with enzymatic treatment and LC-MS/MS, the structures of transglycosylated hesperidin were identified as 4G-α-D-glucopyranosyl-hesperidin (HG1) and 4G-α-D-maltopyranosyl-hesperidin (HG2), as illustrated in Figure 5.

Table 1.
13C-NMR analysis of hesperetin, hesperidin and HG products.

2.3. Antibacterial Activity
HG2 showed the lowest MIC values of 4 mg/mL against S. aureus and 8 mg/mL against E. coli (Table 2). In the negative control, sterile water had no inhibitory effect on any bacteria tested. Ampicillin had good antibacterial activity against both organisms.
Table 2.
Minimal inhibitory concentration (MIC) of hesperidin glycosides, diosmin, and standard drug.
The lowest MBC values were 8 mg/mL for HG1 and HG2 against S. aureus, while original hesperidin and diosmin both had MBC values of 32 mg/mL against E. coli and 16 mg/mL against S. aureus (Table 3).
Table 3.
Minimal bactericidal concentration (MBC) of hesperidin glycosides, diosmin, and standard drug.
The MIC and MBC results showed that hesperidin glycosides had a higher antibacterial activity against Gram-positive S. aureus than hesperidin and diosmin. This result was the same as that obtained for ampicillin antibiotic, which showed better activity against Gram-positive than Gram-negative bacteria. Thus, HGs are effective agents for inhibiting S. aureus growth.
2.4. In Vitro α-Glucosidase Inhibitory Activity
As shown in Figure 6, the IC50 values of hesperidin, HG1, HG2 and diosmin were 2.75 ± 1.57, 2.48 ± 1.61, 2.36 ± 1.48 and 2.99 ± 1.23 mg/mL, respectively, whereas acarbose had an IC50 value of 1.64 ± 1.04 mg/mL. The transglycosylated HG1 and HG2 exhibited a higher inhibitory effect than original hesperidin and diosmin. Thus, adding a sugar molecule to hesperidin was a good way to inhibit α-glucosidase activity. To better understand the inhibition of hesperidin, HG2, which gave the best IC50, was chosen to determine the values of kinetic inhibition. The Lineweaver–Burk plot shows that HG2 is a competitive inhibitor of α-glucosidase (Figure 7) with an inhibitory constant (Ki) of 2.20 ± 0.10 mM. For other kinetic parameters, maximum velocity (Vmax) of α-glucosidase assayed under different conditions, with and without HG2 inhibitor, showed the same values of 0.15 ± 0.08 µmole/min. In contrast, the Michaelis constant (Km) of the enzyme on sucrose substrate without HG2 inhibitor was lower (1.34 ± 0.10 mM) compared to the condition with inhibitor (2.34 ± 0.43 mM), and its turnover number (Kcat) was 1.53 × 103 min−1.
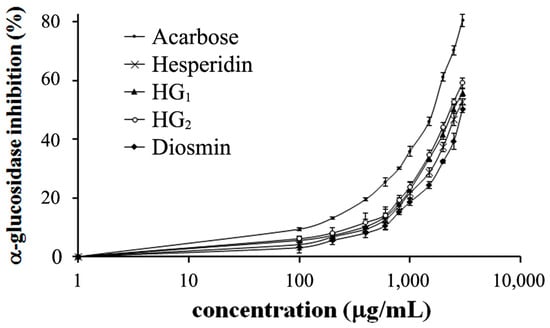
Figure 6.
Inhibitory effect of hesperidin, hesperidin glycosides, diosmin and standard acarbose on α-glucosidase from rat intestinal acetone powder. Data shown are mean ± SD of three independent experiments.
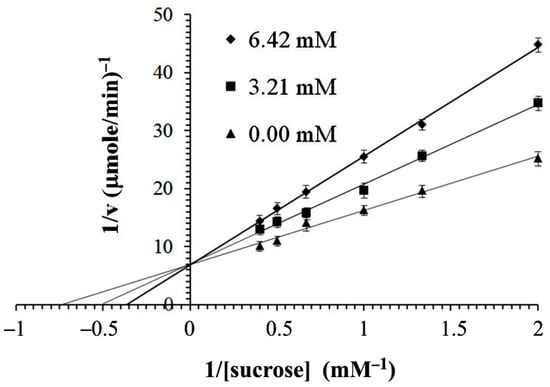
Figure 7.
Lineweaver–Burk plots of α-glucosidase and sucrose in the absence (triangles) and presence (squares and diamonds) of HG2.
3. Discussion
There are many reports on transglycosylation by CGTases in relation to the synthesis of new glycosides [26,27,28,29], but few on the transglycosylation of hesperidin to hesperidin glycosides [30,31]. Kometani et al. [30] reported the formation of HGs in a small scale system at alkaline pH levels. The products with the highest yield (1.23 mM) were purified by HPLC on an ODS column. The structure of the purified hesperidin monoglucoside was identified to be 4G-α-D-glucopyranosyl hesperidin by FAB-MS, α- and β-glucosidase, and glucoamylase treatment. They also found that the transglycosylated hesperidin had higher water soluble and stabilizing properties than hesperidin. Using a small scale system, Go et al. [31] reported a conversion yield of 71.2%. We found that the conversion yield of HGs (38%) was approximately the same as that reported previously for HGs (40%) when using an alkalophilic Bacillus sp. CGTase to catalyze the transglycosylation of glycosylated hesperidin [30].
In this investigation, we performed upscale transglycosylation of hesperidin to glycosides by CGTase, with hesperidin as the glycosyl acceptor. In order to prepare larger quantities of glycosides, the reaction mixtures were scaled up and performed under optimal conditions, and each HG product was characterized by HPLC. Each glycoside product of hesperidin was completely isolated and given only one spot on TLC (Figure 2, lanes 3, 5 and 7). In addition, this is the first report on the structural identification of HGs by glucoamylase treatment and LC-MS/MS analysis. The structure of HGs was preliminarily analyzed in terms of the number of glucose units and their configuration. As a result, HG1 and HG2 could be digested by glucoamylase, an exohydrolase enzyme that catalyzes the hydrolysis of 1,4-linked α-D-glucose residues in glucosyl polysaccharides to liberate glucose units [32]. After hydrolysis by glucoamylase, the TLC results showed that HG1, HG2, and HG3 yielded hesperidin and glucose spots, and the molar ratios of hesperidin to glucose were 1:0.8, 1:2, and 1:3, respectively (Figure 2, lanes 4, 6 and 8). HG1 was also digested by glucoamylase because the linkage between hesperidin and the first glucosyl unit did not directly link to the hesperetin flavonoid ring. However, the first transferred glucose was attached to the C-4″ glucose position of rutinose (a disaccharide) in the hesperidin molecule and the LC-MS/MS and NMR analysis suggested that these glycosides were linked to each glucose via an α-1,4 linkage. The molecular mass of HG1–2 was estimated at 772.26 and 934.30 daltons by ESI-MS1 analysis, which gave a molecular ion at m/z 773 and 935 ([M + H])+. Moreover, the ESI-MS2 spectrum of fragmentation patterns confirmed that all observed peaks were part of the studied glycoside derivatives (Figure 3). Based on all of the results and the NMR data analysis (Figure 4 and Figure 5 and Table 1), the structures of HG1 and HG2 were identified as 4G-α-D-glucopyranosyl- and 4G-α-D-maltopyranosyl-hesperidin, respectively.
We also investigated the antibacterial activities of hesperidin and HGs and found that the latter had greater antibacterial activity than original hesperidin and diosmin, especially against S. aureus. The exact mechanisms driving their antibacterial activities remain unclear, but several mechanisms have been proposed such as interference with bacterial DNA synthesis, bacterial movement, cytoplasmic membrane permeability, and the inhibition of bacterial metalloenzymes [13]. Several studies have evaluated the antibacterial activities of plant flavonoid-rich extracts and pure flavonoids, including hesperidin derivatives [14,19,33]. The ethanolic extract of grapefruit seed and pulp, containing mainly naringin and hesperidin flavanones, significantly inhibited Gram-positive bacteria (agar diffusion) and Gram-positive and Gram-negative bacteria by broth dilution [19]. In contrast to our results, Mandalari et al. [33] found that the flavonoid-rich fractions of bergamot peel had activity against Gram-negative bacteria such as E. coli, Pseudomonas putida, and Salmonella enterica but not against Gram-positive bacteria such as Listeria innocua, B. subtilis, S. aureus, and Lactococcus lactis. The treatment of these fractions with pectinase 62 L [33], which converted flavonoid glycosides into their aglycones, increased their antibacterial activity. Moreover, they studied the antibacterial activities values of pure bergamot flavonoids (aglycone forms), neohesperidin, heperetin, neoeriocitrin, eriodictyol, naringin and naringenin, and all MIC values were 250–1000 μg/mL.
Similar to our work, Lee et al. [34] improved the bioavailability and the water solubility of hesperidin by converting it to hesperetin-7-O-glucoside (Hes-7-G) using Aspergillus sojae naringinase. They also found that Hes-7-G had greater antibacterial activity against Helicobacter pylori than hesperetin, but similar activity to hesperidin; the lower inhibitory effect of hesperetin was due to its poor water solubility, and the antibacterial activity of hesperidin and its derivatives depended on the presence or absence of a sugar moiety and the bacteria tested.
Hesperidin has potential antihyperglycemic effects [15,21] by inhibiting α-glucosidase activity [35] and could play a future role in treating type-2 DM. The IC50 values of hesperidin, HG1, HG2 and diosmin were 2.75, 2.48, 2.36 and 2.99 mg/mL, respectively, confirming the greater inhibitory effects of HG1 and HG2 (Ki of 2.20 mM). Sahnoun et al. [35] reported that the IC50 of hesperidin for inhibiting the Saccharomyces cerevisiae α-glucosidase from Sigma (St. Louis, Mo, USA) was 111 μM, and that the inhibitory properties of hesperidin, hesperetin, and naringin depended on their structures. For example, the hydroxyl group at 4′-position in the B ring of flavonoids was associated with greater α-glucosidase inhibition than the methoxy group. They found that flavonoids with a 4′-methoxy substitution in the B ring showed the greatest inhibition against α-amylase, followed by 3′-OH-4′-methoxy and 4′-OH substitution; however, this report did not study the effect of the glucose-linked glycoside moiety. Based on computational molecular dynamics and docking simulations, they also found that two hesperetin rings interacted with several residues near the active site on the α-glucosidase, such as Lys155, Asn241, Glu304, Pro309, Phe311, and Arg312, resulting in reversible inhibition of this enzyme in a slope-parabolic mixed-type manner [36].
Od-ek et al. [21] reported that hesperidin decreased goblet cells in diabetic rats when compared to a control group. Goblet cells in the brush border of the small intestinal produce and maintain the protective mucus blanket [37]. Therefore, decreasing the number of goblet cells may lead to a reduction in digestion and absorption in the small intestine and is another potential antihyperglycemic effect of hesperidin. In this work, the standard acarbose, being a well-known α-glucosidase inhibitor, had an IC50 value of 1.64 mg/mL. Kim et al. [38] reported that acarbose is a competitive inhibitor of the rat intestinal α-glucosidase with a Ki of 0.059 μM by using maltose as the substrate. Thus, one of the potential therapeutic approaches to decrease blood glucose and potentially treat type 2 DM is to reduce glucose absorption by inhibiting carbohydrate-hydrolyzing enzymes, e.g., α-amylase, α-glucosidase, and maltase [39]. For hesperidin derivatives, they presented as potential new alternative agents for treating α-glucosidase-associated type 2 diabetes based on their α-glucosidase inhibition. However, more detailed study in this field is required.
4. Materials and Methods
4.1. Chemicals
Hesperidin and diosmin were purchased from Tokyo Chemical Industry Co., Ltd. (Tokyo, Japan). The commercial glucose oxidase kit was obtained from Human mit-diagnostics GmbH (Idstein, Germany). Tryptone and yeast extract were obtained from Difco (Bacton Dickinson and company, Sparks, MD, USA). All other chemicals used were of analytical grade and purchased from Sigma-Aldrich (St. Louis, MO, USA).
4.2. Synthesis and Detection of Hesperidin Glycosides
The p19bBC cells containing CGTase gene from B. circulans A11 were used to prepare recombinant CGTase in the synthesis of the HG products. HG synthesis was carried out using 200 U/mL of CGTase, 1.5% (w/v) β-CD as the glycosyl donor, and 2% (w/v) hesperidin solution containing 80% (v/v) DMSO as the glycosyl acceptor in 0.2 M acetate buffer, pH 5.0 at 60 °C for 6 h [5]. The obtained HG products were detected by thin layer chromatography (TLC) and high performance liquid chromatography (HPLC) [6].
4.3. Production and Product Purification
The production and purification of HGs were performed under the optimized conditions described above. The transglycosylation volume of synthesis was scaled up to 300 mL. The reaction mixture was filtered through a 0.45 µm nylon membrane before injecting the 900 µL mixture into preparative HPLC and eluted with acetonitrile:water (20:80 v/v), using a flow rate of 20 mL/min. Each HG peak was separately collected by preparative HPLC (Agilent Technologies 1260, Böblingen, Germany) using a 7 µm ZORBAX SB-C18 PrepHT column (21.2 × 150 mm) and detected with UV detector at 280 nm. Thereafter, the purified HG was concentrated by evaporation for further characterization.
4.4. Structural Analysis of Hesperidin Glycosides
4.4.1. Digestion with Amylolytic Enzyme
HGs were mixed and dissolved in 0.2 M acetate buffer, pH 5.5. The reactions were incubated with 20 U/mL of Rhizopus sp. glucoamylase (EC 3.2.1.3) (Sigma, St. Louis, Mo, USA) at 37 °C for 24 h. After incubation, the reactions were stopped by boiling for 5 min and then 10-µL aliquots from the reactions were analyzed by TLC using a mobile phase, ethyl acetate/ acetic acid/ water in a 3:1:1 (v/v) ratio [5].
4.4.2. Liquid Chromatography Mass Spectrometer (LC-MS/MS) Analysis
The fragmentation patterns of the synthesized HGs were analyzed by LC-MS/MS (Thermo scientific TSQ Endura™, Waltham, MA, USA) at the Service Unit of the National Centre for Genetic Engineering and Biotechnology (BIOTECH), Thailand. HG products were dissolved in DMSO at concentrations of 1 mg/mL. LC separation was performed on a reverse phase C18 and the retention time of HG2, HG1 and hesperidin was 6.2, 7.7 and 8.4 min, respectively. Then, the products were ionized by electrospray ionization (ESI) source at a constant flow rate of 5 µL/min [40] in order to increase the [M + H]+ signal intensity in the positive ion mode. Quadrupole MS detection and quantitation was done by monitoring at m/z 611 [M + H]+ for hesperidin, m/z 773 [M + H]+ for HG1, and m/z 935 [M + H]+ for HG2. Fragmentation patterns in the mass spectra of each HG were analyzed by tandem mass spectrometry mode and generated as specific mass to charge (m/z).
4.4.3. NMR Analysis
The α-1,4-glycosidic bonds of HGs were identified by 1H and 13C-NMR spectra with Bruker’s AVANCE Tech HD 400 NMR spectrometer (Bruker, Billerica, MA, USA) at the BIOTECH, Thailand. The operation used 400 MHz at room temperature. The NMR spectra were obtained with 3–10 mg of each sample dissolved in DMSO-d6.
4.5. Antibacterial Activity
Cultures of the Gram-positive Staphylococcus aureus ATCC 25923 and Gram-negative Escherichia coli ATCC 25922 were used in the study. The microorganisms were grown on Luria-Bertani (LB) broth at 37 °C for 24 h. A broth microdilution method was performed according to the Clinical and Laboratory Standard Institute (CLSI) [41].
4.5.1. Minimal Inhibitory Concentration (MIC)
The MIC value was determined by the microdilution technique in liquid LB medium by incubating the bacteria in LB broth with variable amounts of the sample being tested [26]. This technique utilizes the microdilution method in a 96-well microplate. Briefly, the 0.5 McFarland cultures (OD600 = 0.1) of S. aureus and E. coli were diluted with LB broth in a ratio of 1:200 (v/v). Two-fold serial dilutions of hesperidin, hesperidin glycosides and diosmin were taken 50 µL in 96 well microplates. After 24 h incubation, 1 mg/mL of resazurin solution (10 µL) was added to each well and the plate was then incubated for 2 h. The MIC value was determined as the lowest concentration of the hesperidin, hesperidin glycosides and diosmin that prevented a color change of resazurin. A blue dye (resazurin) becomes pink (resorufin) when it is reduced by oxidoreductase of viable bacteria in a medium. The MIC value was performed in triplicate experiments and the mean presented as mg/mL [42,43].
4.5.2. Minimal Bactericidal Concentration (MBC)
After the MIC test had been completed, the MBC value was investigated by sub-culturing each well in which no visible growth occurred from a previous MIC test (blue color) to an agar medium. After 24 h incubation at 37 °C, bacterial growth on the agar plate was examined. The lowest concentration at which the hesperidin glycosides eliminated all bacteria represented the MBC. The tests were done in triplicate and the mean MBC was recorded as mg/mL.
4.6. In Vitro α-Glucosidase Inhibitory Activity
α-Glucosidase inhibitory activity [44] was performed using intestinal acetone powder from rats (Sigma, St. Louis, Mo, USA). Hesperidin, hesperidin glycosides and diosmin inhibitors were dissolved in DMSO and prepared at final concentrations of 0.1–2 mg/mL. Then, 100 U/mL of α-glucosidase were pre-incubated with each inhibitor at 37 °C for 30 min, to which sucrose substrate (5.85 mM) in 0.1 M phosphate buffer (pH 6.9) was added. This was incubated for 10 min at 37 °C. After incubation, the reaction was stopped by boiling for 10 min at 100 °C and 10 µL aliquots from each the reaction were added to 1 mL glucose oxidase reagent and incubated for 5 min at 37 °C. The amount of glucose released from this reaction was measured using a glucose oxidase kit at 500 nm. A control reaction was performed without adding inhibitor and acarbose was used as a standard compound for comparison of α-glucosidase inhibitory activity (positive control). All reactions were performed in triplicate experiments. The percentage of α-glucosidase inhibition was calculated by the following Equation (1):
where Acontrol is the absorbance of the enzymatic reaction without inhibitor, and Asample is the absorbance of the enzymatic reaction in presence of the inhibitor. The IC50 was calculated by the curve fitting the data (GraphPad Prism 5.0, GraphPad Software, Inc., San Diego, CA, USA).
Initial rates of reaction were measured at different sucrose concentrations. Lineweaver–Burk plots, 1/V0 vs. 1/[sucrose] with/without HG2 α-glucosidase inhibitor were generated using the Sigma Plot 9.0 program (Systat Software Inc., San Jose, CA, USA) to generate the maximum reaction rate (Vmax) and the Michaelis constant (Km). The inhibition constant (Ki) was calculated using the following Equation (2) [45,46]:
where [I] is the inhibitor concentration.
4.7. Statistical Analyses
Data are presented as the mean ± SD from three independent experiments. Statistical analysis was performed using a Student’s t-test with IBM SPSS Statistic version 26.0 (SPSS Corporation, Chicago, IL, USA). A P-value < 0.05 was considered to be statistically significantly.
5. Conclusions
In conclusion, our structural investigation and upscale production of hesperidin derivatives in the presence of sugar moiety was successfully carried out. Hesperidin glycosides (HG1–2) were shown to be able to prevent bacterial growth and inhibit intestinal α-glucosidase. The best IC50 of α-glucosidase inhibition was acarbose, followed by HG2 and HG1. The present study supports the potential use of HGs as antibacterial and anti-α-glucosidase (antidiabetic) agents.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/catal11050532/s1, Figure S1. The 400 MHz 13C-NMR spectra of the (A) hesperetin, (B) hesperidin, (C) HG1, and (D) HG2.
Author Contributions
Conceptualization, J.K.; formal analysis, T.C., P.R. and J.K.; investigation, T.C. and J.K.; data curation, T.C. and J.K.; project administration, J.K.; funding acquisition, P.R. and J.K.; writing—original draft preparation, T.C. and J.K.; writing—manuscript preparation, J.K.; writing—review and editing, P.R. and J.K. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financially supported by the Thammasat University Research Fund, Contract No. TUFT 059/2563 and partial support from the Phramongkutklao College of Medicine Research Fund.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institute Biosafety Committee of Thammasat University (TU-IBC, Approval No. 041/2563).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article.
Acknowledgments
T.C. was directly supported for personal expenses by the Research Assistant Scholarship from Thammasat University, Year 2561–2563. We also thank the Drug Discovery and Development Center, and Center of Scientific Equipment for Advanced Research, Thammasat University for providing a special service rate for preparative HPLC.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Villiers, A.; Venter, P.; Pasch, H. Recent advances and trends in the liquid-chromatography-mass spectrometry analysis of flavonoids. J. Chromatogr. A 2016, 1430, 16–78. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Biochem. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed]
- Kuntic, V.; Brboric, J.; Holclajtner-Antunović, I.; Uskoković-Marković, S. Evaluating the bioactive effects of flavonoid hesperidin: A new literature data survey. Vojnosanit. Pregl. 2014, 71, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Kometani, T.; Terada, Y.; Nishimura, T.; Nakae, T.; Takii, H.; Okada, S. Acceptor specificity of cyclodextrin glucanotransferase from an alkalophilic Bacillus species and synthesis of glucosyl rhamnose. Biosci. Biotechnol. Biochem. 1996, 60, 1176–1178. [Google Scholar] [CrossRef] [PubMed]
- Chaisin, T.; Kaulpiboon, J.; Poomipark, N. Acceptor specificity of recombinant cyclodextrin glycosyltransferase from Bacillus circulans A11. STA 2018, 23, 86–97. [Google Scholar]
- Poomipark, N.; Chaisin, T.; Kaulpiboon, J. Synthesis and evaluation of antioxidant and β-glucuronidase inhibitory activity of hesperidin glycosides. Agric. Nat. Resour. 2020, 54, 165–172. [Google Scholar]
- Elavarasan, J.; Velusamy, P.; Ganesan, T.; Ramakrishnan, S.; Rajasekaran, D.; Kalaiselvi, P. Hesperidin-mediated expression of Nrf2 and upregulation of antioxidant status in senescent rat heart. J. Pharm. Pharmacol. 2012, 64, 1472–1482. [Google Scholar] [CrossRef]
- Kang, S.; Park, K.; Park, H.S.; Lee, D.H.; Kim, J.; Nagappan, A.; Kim, E.H.; Lee, W.; Shin, S.; Park, M.; et al. Anti-inflammatory effect of flavonoids isolated from Korea Citrus aurantium L. on lipopolysaccharide-induced mouse macrophage RAW 264.7 cells by blocking of nuclear factor-kappa B (NF-kB) and mitogen-activated protein kinase (MAPK) signalling pathways. Food Chem. 2011, 129, 1721–1728. [Google Scholar] [CrossRef]
- Parhiz, H.; Roohbakhsh, A.; Soltani, F.; Rezaee, R.; Iranshahi, M. Antioxidant and anti-inflammatory properties of the citrus flavonoids hesperidin and hesperetin: An updated review of their molecular mechanisms and experimental models. Phytother. Res. 2015, 29, 323–331. [Google Scholar] [CrossRef]
- Cincin, Z.; Kiran, B.; Baran, Y.; Cakmakoglu, B. Hesperidin promotes programmed cell death by downregulation of nongenomic estrogen receptor signalling pathway in endometrial cancer cells. Biomed. Pharmacother. 2018, 103, 336–345. [Google Scholar] [CrossRef]
- Cincin, Z.; Unlu, M.; Kiran, B.; Sinem Bireller, E.; Baran, Y.; Cakmakoglu, B. Anti-proliferative, apoptotic and signal transduction effects of hesperidin in non-small cell lung cancer cells. Cell Oncol. 2015, 38, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, H.; Zhang, J.; Gao, J.; Ge, X.; Lou, G. Hesperidin inhibits HeLa cell proliferation through apoptosis mediated by endoplasmic reticulum stress pathways and cell cycle arrest. BMC Cancer 2015, 15, 682. [Google Scholar] [CrossRef]
- Iranshahi, M.; Rezaee, R.; Parhiz, H.; Roohbakhsh, A.; Soltani, F. Protective effects of flavonoids against microbes and toxins: The cases of hesperidin and hesperetin. Life Sci. 2015, 137, 125–132. [Google Scholar] [CrossRef]
- Yi, Z.; Yu, Y.; Liang, Y.; Zeng, B. In vitro antioxidant and antimicrobial activities of the extract of pericarpium citri reticulatae of a new citrus cultivar and its main flavonoids. Food Sci. Technol. 2008, 41, 597–603. [Google Scholar] [CrossRef]
- Ahmed, O.M.; Mahmoud, A.M.; Abdel-Moneim, A.; Ashour, M.B. Antidiabetic effects of hesperidin and naringin in type 2 diabetic rats. Diabetol. Croat. 2012, 41, 53–67. [Google Scholar]
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial activity of flavonoids and their structure-activity relationship: An update review. Phytother. Res. 2019, 33, 13–40. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [PubMed]
- Das Neves, R.N.; Carvalho, F.; Carvalho, M.; Fernandes, E.; Soares, E.; Bastos, M.D.L.; Pereira, M.D.L. Protective activity of hesperidin and lipoic acid against sodium arsenite acute toxicity in mice. Toxicol. Pathol. 2004, 32, 527–535. [Google Scholar] [CrossRef]
- Cvetnić, Z.; Vladimir-Knezević, S. Antimicrobial activity of grapefruit seed and pulp ethanolic extract. Acta Pharm. 2004, 54, 243–250. [Google Scholar]
- Taskinen, M.R. Diabetic dyslipidemia. Atheroscl. Suppl. 2002, 3, 47–51. [Google Scholar] [CrossRef]
- Od-ek, P.; Phoungpetchara, I.; Hanchang, W.; Tunsophon, S. Inhibitory effect of hesperidin on glucosidase activity and small intestinal morphology in diabetic rats. Naresuan Univ. J. Sci. Technol. 2016, 24, 88–99. [Google Scholar]
- Cuyckens, F.; Claeys, M. Mass spectrometry in the structural analysis of flavonoids. J. Mass Spectrom. 2004, 39, 1–15. [Google Scholar] [CrossRef]
- Tsimogiannis, D.; Samiotaki, M.; Panayotou, G.; Oreopoulou, V. Characterization of flavonoid subgroups and hydroxy substitution by HPLC-MS/MS. Molecules 2007, 12, 593–606. [Google Scholar] [CrossRef]
- Colson, P.; King, R.R. The 13C-n.m.r. spectra of disaccharides of d-glucose, d-galactose, and l-rhamnose as models for immununological polysaccharides. Carbohydr. Res. 1976, 47, 1–13. [Google Scholar] [CrossRef]
- Kitty, S.C.T.; Izabela, K.; Jian, Z. Identification and quantification of phenolics in Australian native mint (Mentha australis R. Br.). Food Chem. 2016, 192, 698–705. [Google Scholar]
- Charoensapyanan, R.; Ito, K.; Rudeekulthamrong, P.; Kaulpiboon, J. Enzymatic synthesis of propyl-α-glycosides and their application as emulsifying and antibacterial agents. Biotechnol. Bioprocess Eng. 2016, 21, 389–401. [Google Scholar] [CrossRef]
- Aramsangtienchai, P.; Chavasiri, W.; Ito, K.; Pongsawasdi, P. Synthesis of epicatechin glucosides by a β-cyclodextrin glycosyltransferase. J. Mol. Catal. B-Enzym. 2011, 73, 27–34. [Google Scholar] [CrossRef]
- Khummanee, N.; Rudeekulthamrong, R.; Kaulpiboon, J. Cyclodextrin glycosyltransferase-catalyzed synthesis of pinoresinol-α-D-glucoside having antioxidant and anti-inflammatory activities. Appl. Biochem. Microbiol. 2019, 55, 360–370. [Google Scholar] [CrossRef]
- Kaulpiboon, J.; Rudeekulthamrong, P. Biosynthesis of methyl glucoside and its antibacterial activity against Staphylococcus aureus and Escherichia coli. Bioact. Carbohydr. Diet. Fibre 2019, 20, 100197. [Google Scholar] [CrossRef]
- Kometani, T.; Terada, Y.; Nishimura, T.; Takii, H.; Okada, S. Transglycosylation to hesperidin by cyclodextrin glucanotransferase from an alkalophilic Bacillus species in alkaline pH and properties of hesperidin glycosides. Biosci. Biotechnol. Biochem. 1994, 58, 1990–1994. [Google Scholar] [CrossRef]
- Go, Y.H.; Kim, T.K.; Lee, K.W.; Lee, Y.H. Functional characteristics of cyclodextrin glucanotransferase from alkalophilic Bacillus sp. BL-31 highly specific for intermolecular transglycosylation of bioflavonoids. J. Microbiol. Biotechnol. 2007, 17, 1550–1553. [Google Scholar] [PubMed]
- Chiba, S. Molecular mechanism in α-glucosidase and glucoamylase. Biosci. Biotechnol. Biochem. 1997, 61, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Bennett, R.N.; Bisignano, G.; Trombetta, D.; Saija, A.; Faulds, C.B.; Gasson, M.J.; Narbad, A. Antimicrobial activity of flavonoids extracted from bergamot (Citrus bergamia Risso) peel, a byproduct of the essential oil industry. J. Appl. Microbiol. 2007, 103, 2056–2064. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Huh, J.Y.; Nam, S.H.; Moon, S.K.; Lee, S.B. Enzymatic bioconversion of citrus hesperidin by Aspergillus sojae naringinase: Enhanced solubility of hesperetin-7-O-glucoside with in vitro inhibition of human intestinal maltase, HMG-CoA reductase, and growth of Helicobacter pylori. Food Chem. 2012, 135, 2253–2259. [Google Scholar] [CrossRef] [PubMed]
- Sahnoun, M.; Trabelsi, S.; Bejar, S. Citrus flavonoids collectively dominate the α-amylase and α-glucosidase inhibitions. Biologia 2017, 72, 764–773. [Google Scholar] [CrossRef]
- Gong, Y.; Qin, X.-Y.; Zhai, Y.-Y.; Hao, H.; Lee, J.; Park, Y.-D. Inhibitory effect of hesperetin on α-glucosidase: Molecular dynamics simulation integrating inhibition kinetics. Int. J. Biol. Macromol. 2017, 101, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Specian, R.D.; Oliver, M.G. Functional biology of intestinal goblet cells. Am. J. Physiol. Cell Physiol. 1991, 260, C183–C193. [Google Scholar] [CrossRef]
- Kim, M.J.; Lee, S.B.; Lee, H.S.; Lee, S.Y.; Baek, J.S.; Kim, D.; Moon, T.W.; Robyt, J.F.; Park, K.H. Comparative study of the inhibition of alpha-glucosidase, alpha-amylase, and cyclomaltodextrin glucanosyltransferase by acarbose, isoacarbose, and acarviosine-glucose. Arch. Biochem. Biophys. 1999, 371, 277–283. [Google Scholar] [CrossRef]
- Laoufi, H.; Benariba, N.; Adjdir, S.; Djaziri, R. In vitro α-amylase and α-glucosidase inhibitory activity of Ononis angustissima extracts. J. App. Pharm. Sci. 2017, 7, 191–198. [Google Scholar]
- Cuyckens, F.; Ma, Y.L.; Pocsfalvi, G.; Claeysi, M. Tandem mass spectral strategies for the structural characterization of flavonoid glycosides. Analusis 2000, 28, 888–895. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standard Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, Supplement M100, 31st ed.; Wayne, P., Ed.; Clinical and Laboratory Standard Institute: Wayne, PA, USA, 2021. [Google Scholar]
- Awwad, A.; Abdeen, A. Green synthesis of silver nanoparticles using carob leaf extract and its antibacterial activity. Int. J. Ind. Chem. 2013, 4, 29. [Google Scholar] [CrossRef]
- Bhalodia, N.R.; Shukla, V.J. Antibacterial and antifungal activities from leaf extracts of Cassia fistula L.: An ethnomedicinal plant. J. Adv. Pharm. Technol. Res. 2011, 2, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Damsud, T.; Grace, M.; Adisakwattana, S.; Phuwapraisirisan, P. Orthosiphol A from the aerial parts of Orthosiphon aristatus is putatively responsible for hypoglycemic effect via α-glucosidase inhibition. Nat. Prod. Commun. 2014, 9, 639–641. [Google Scholar] [CrossRef] [PubMed]
- Palmer, T. Enzyme inhibition. In Understanding Enzymes; Acford, R.J., Ed.; Ellis Horwood Limited Publishers: Chichester, UK, 1981; pp. 142–169. [Google Scholar]
- Mechchate, H.; Es-Safi, I.; Louba, A.; Alqahtani, A.S.; Nasr, F.A.; Noman, O.M.; Farooq, M.; Alharbi, M.S.; Alqahtani, A.; Bari, A.; et al. In vitro alpha-amylase and alpha-glucosidase inhibitory activity and in vivo antidiabetic activity of Withania frutescens L. Foliar extract. Molecules 2021, 26, 293. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).