
Chiral Synthesis of 3-Amino-1-phenylbutane by a Multi-Enzymatic Cascade System



Abstract
:1. Introduction
2. Results and Discussion
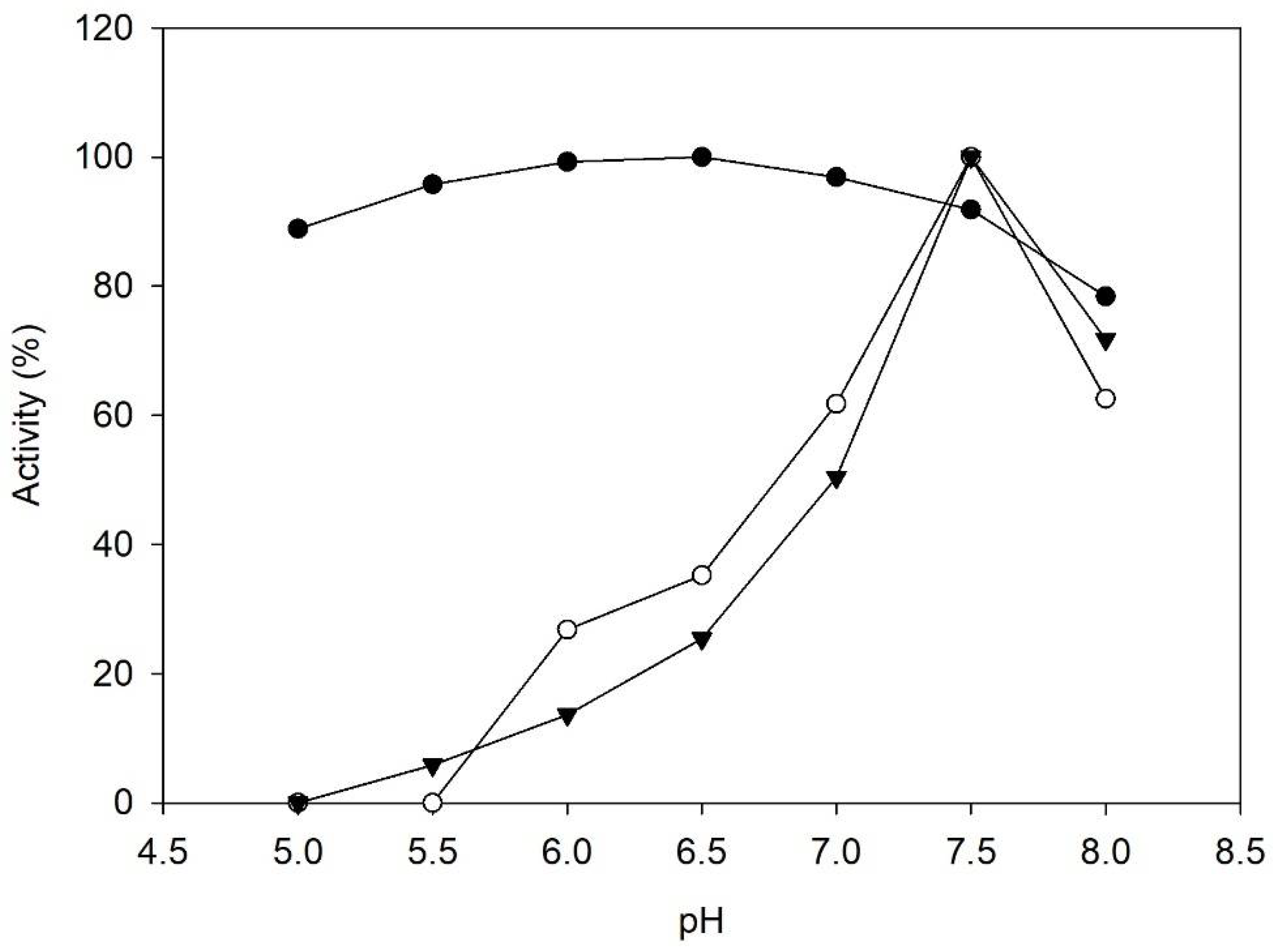
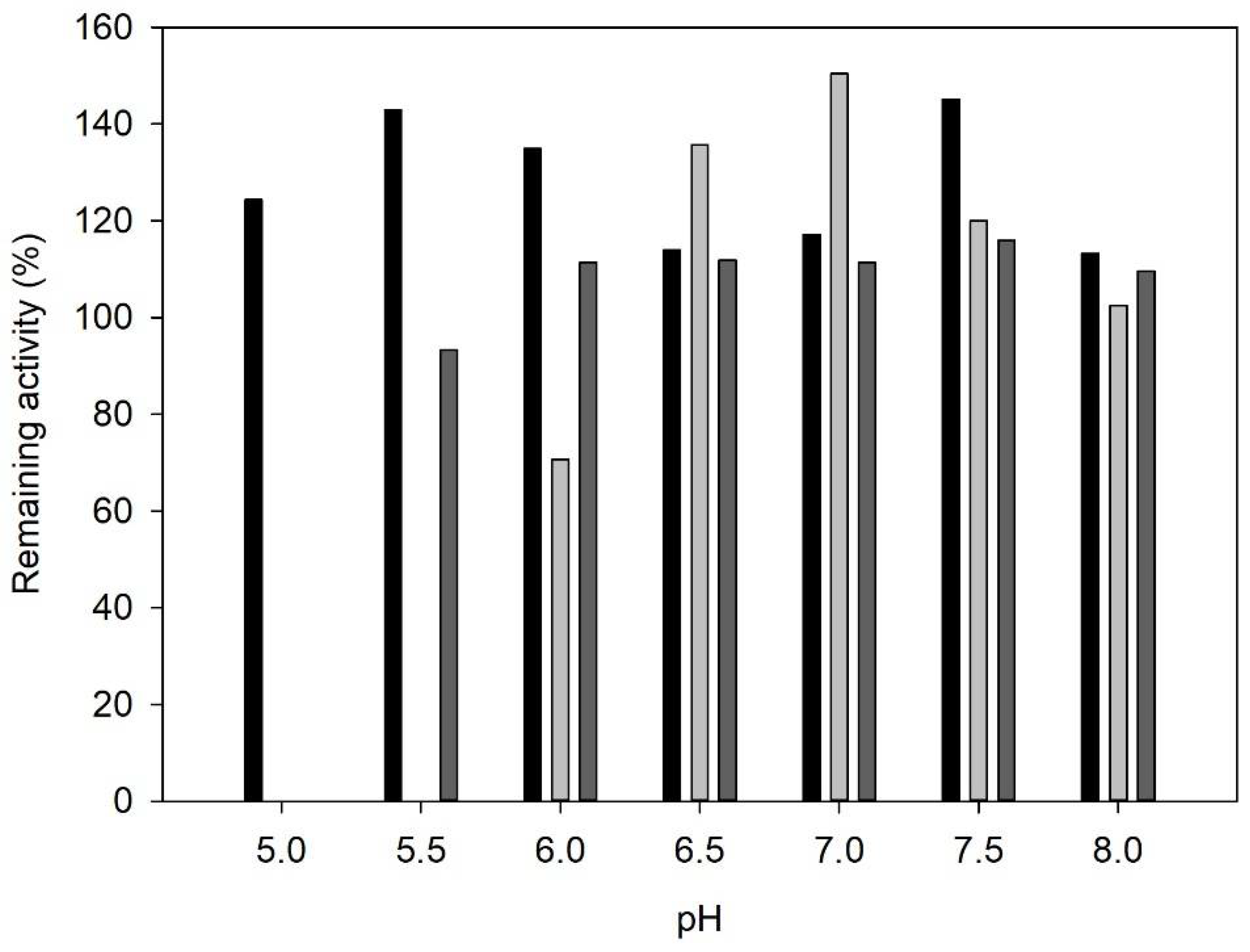
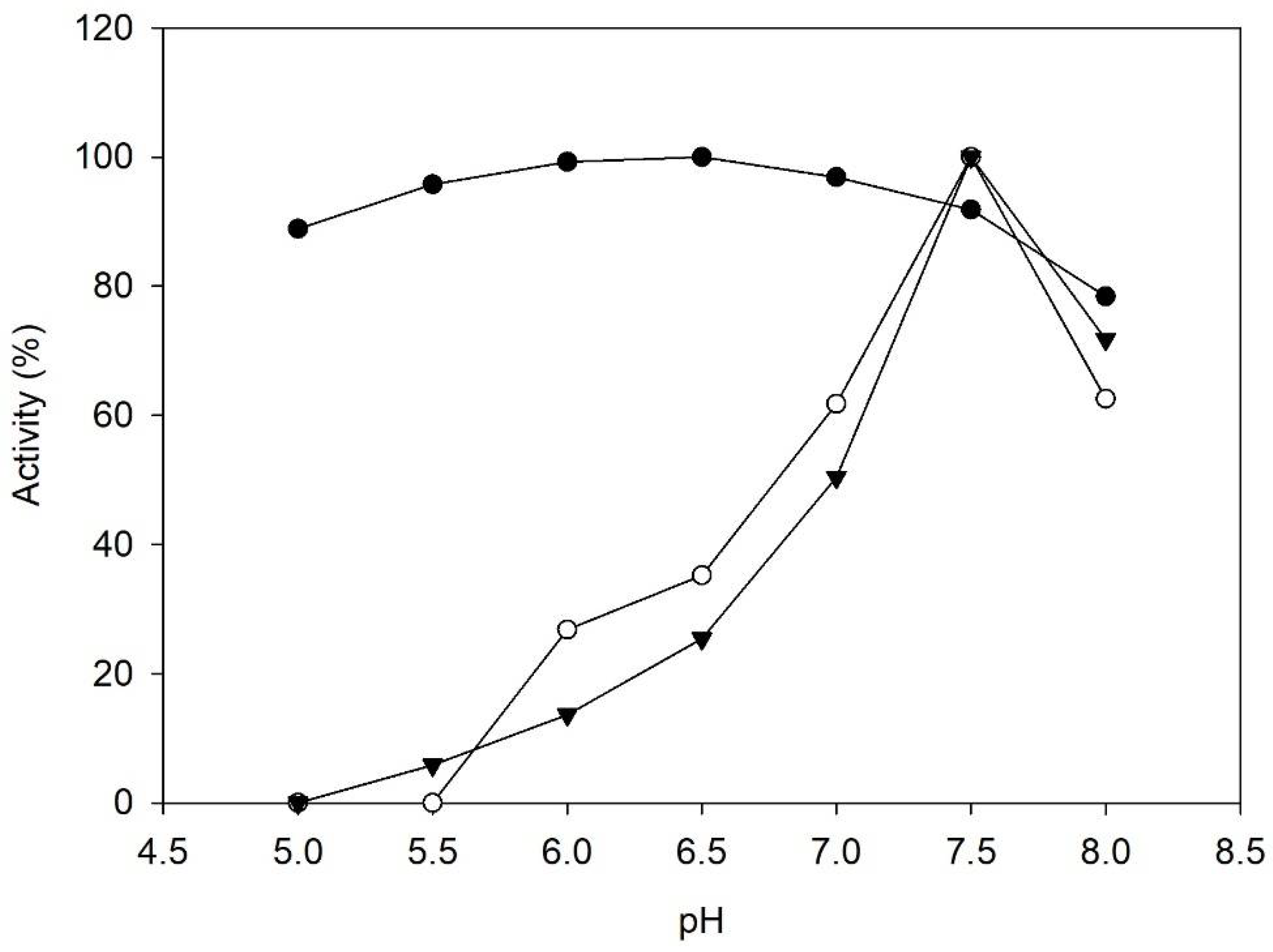
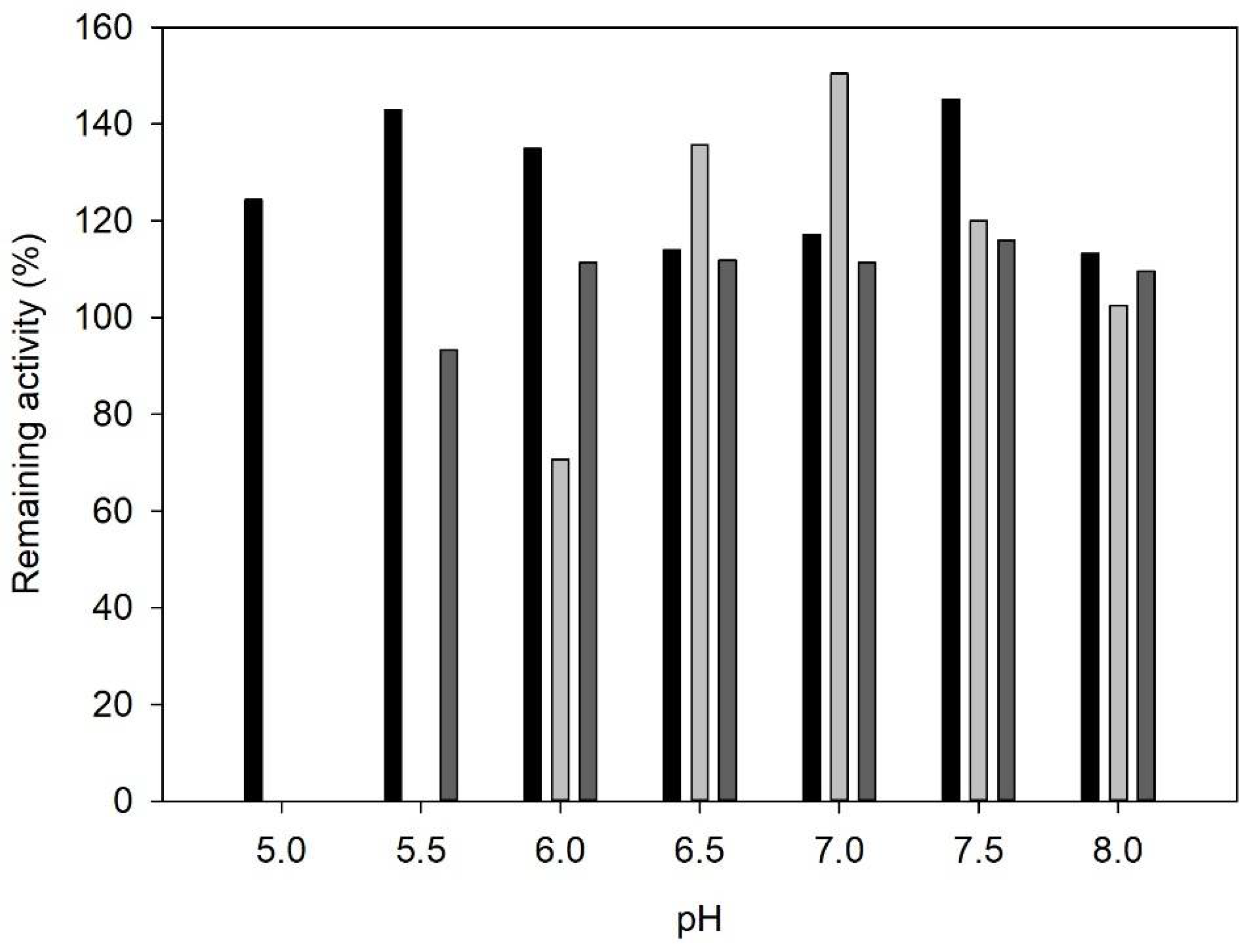
2.1. Enzyme Characterization
2.2. Transaminase Testing
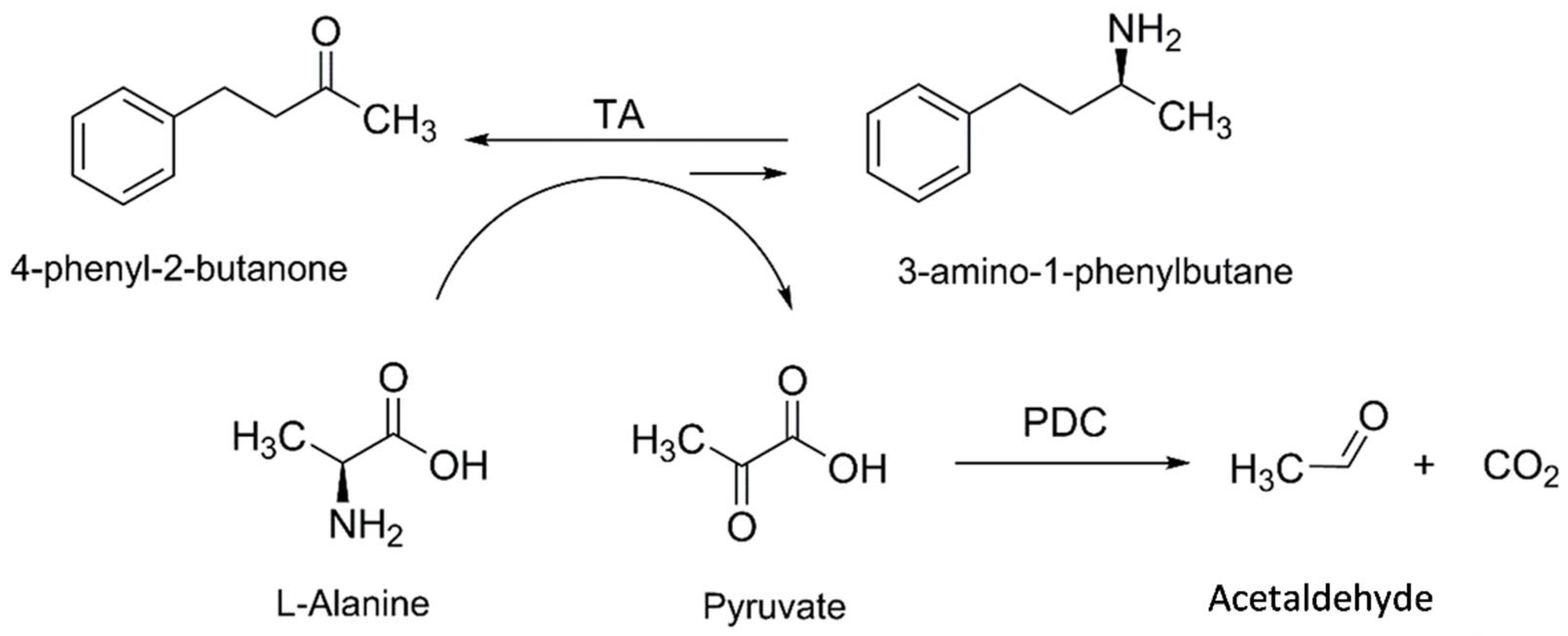
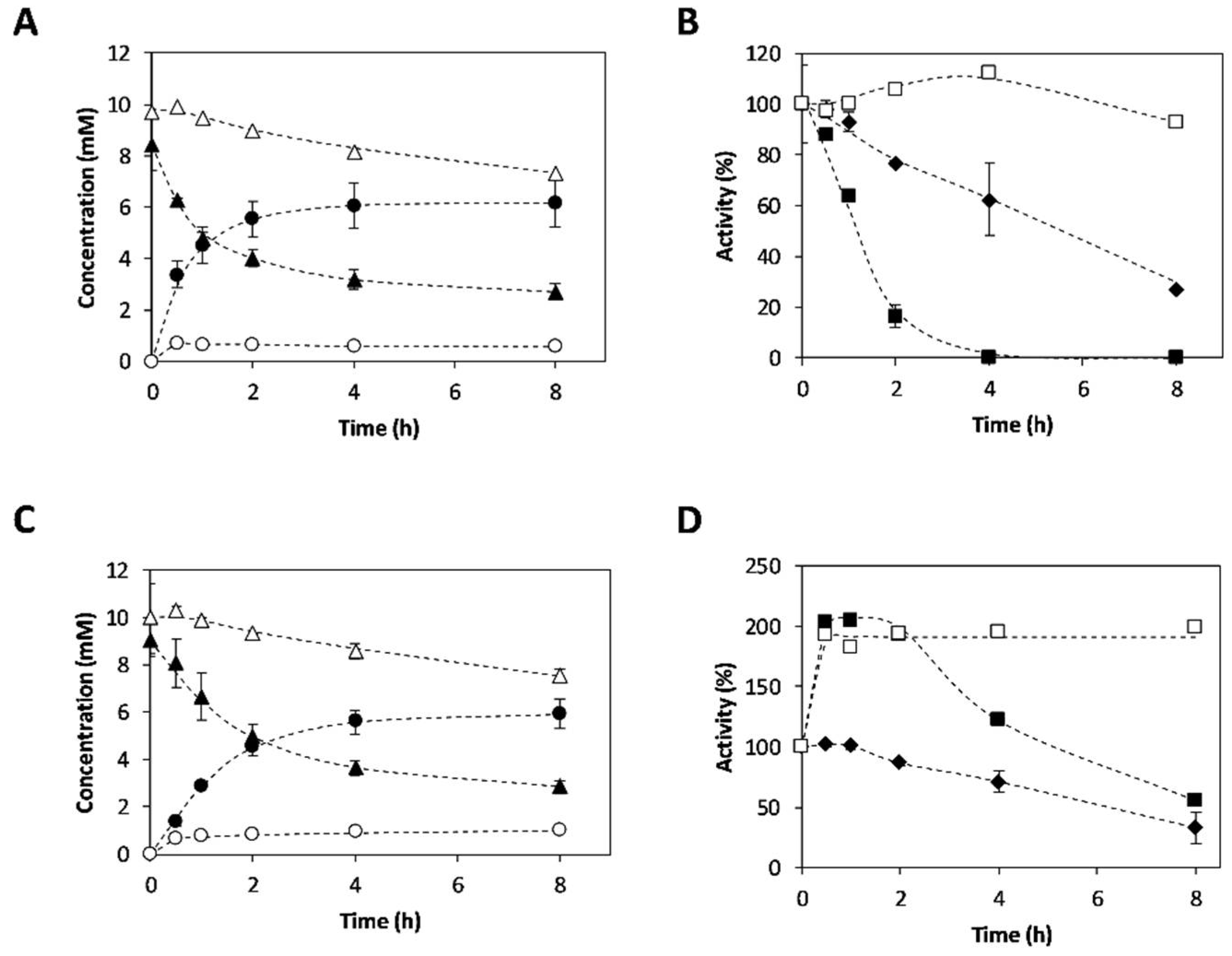
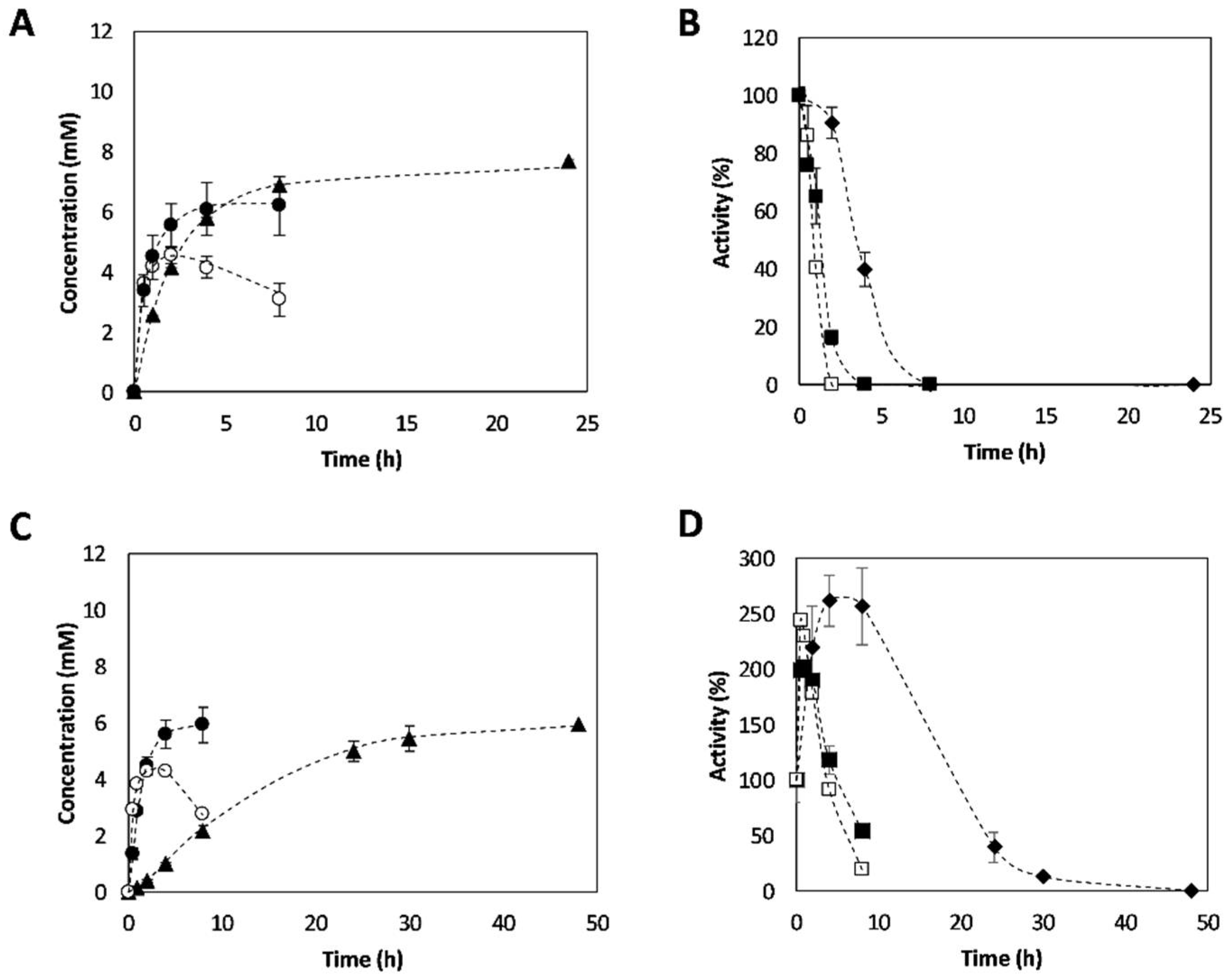
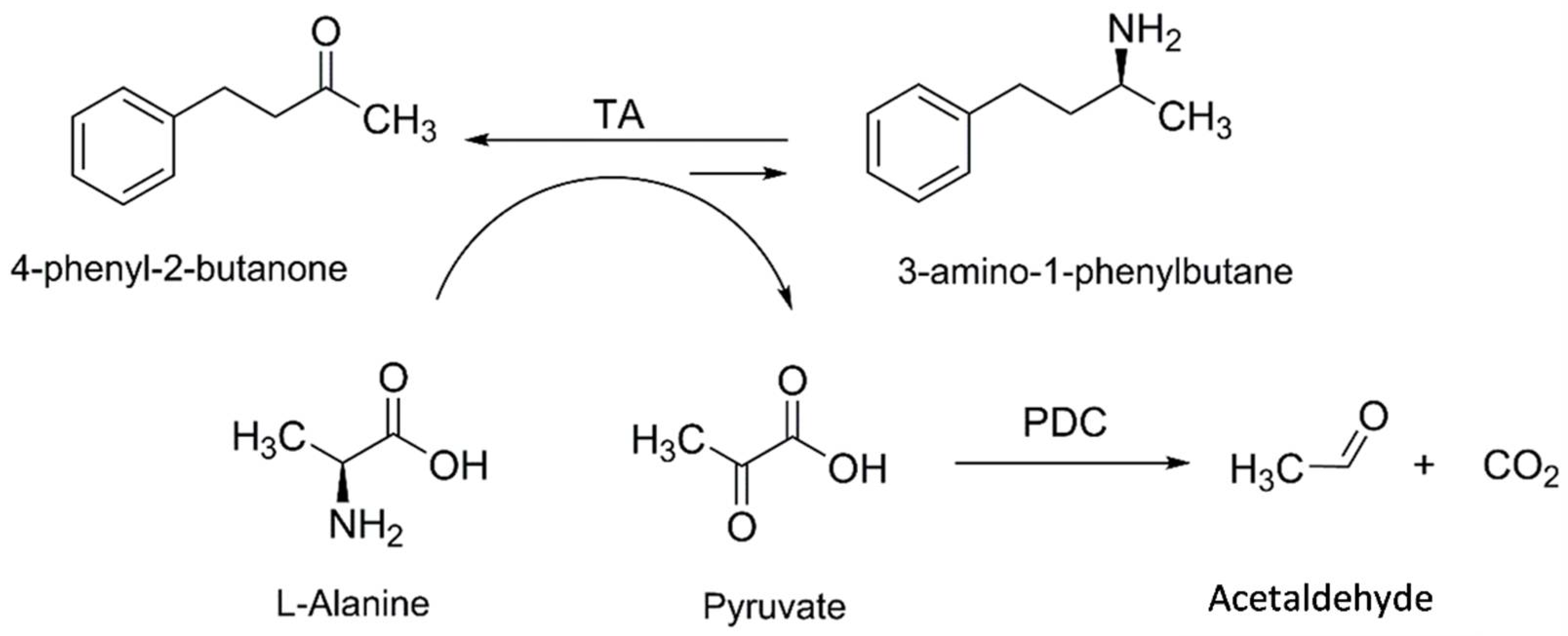
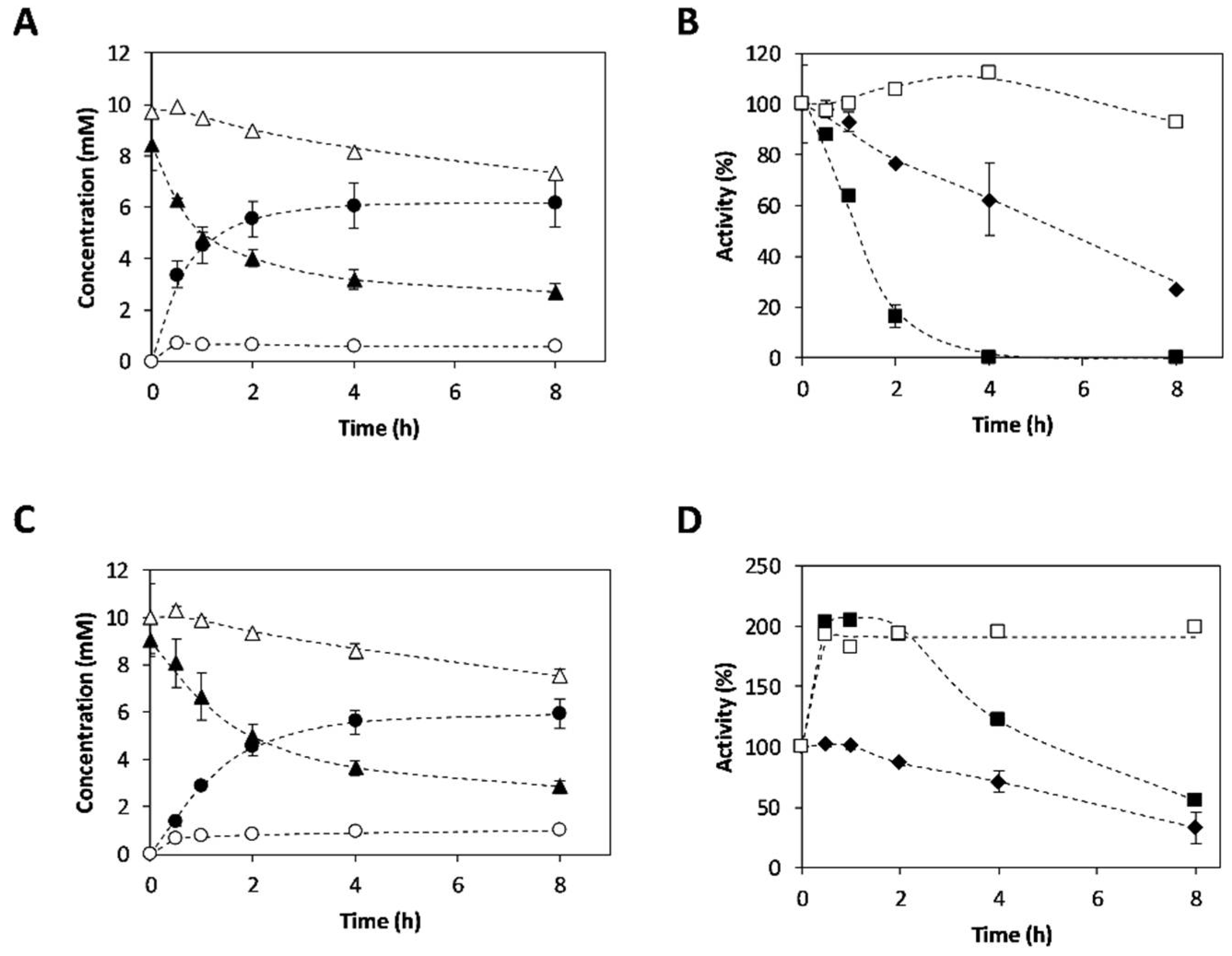
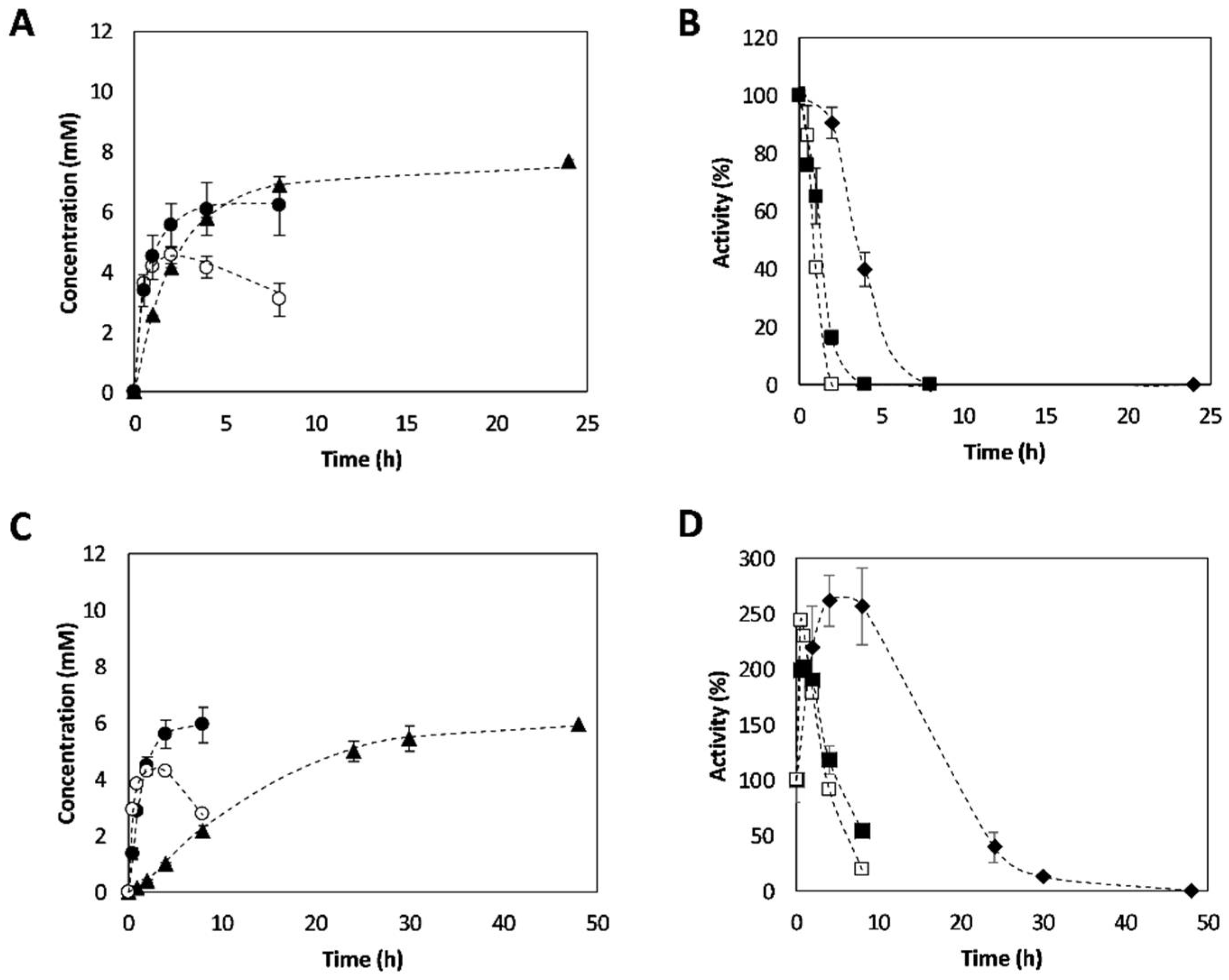
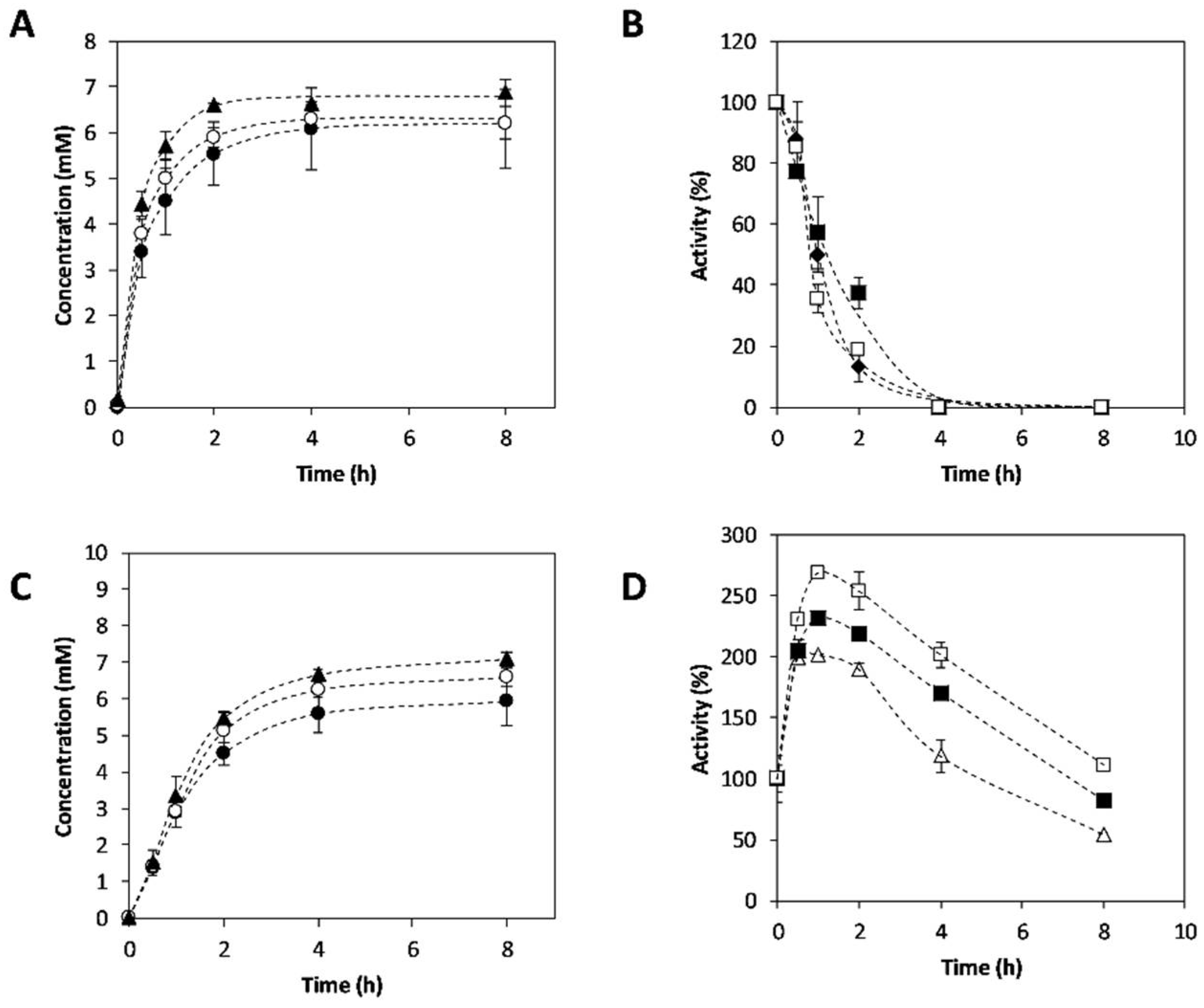
2.3. Synthesis of 3-APB Catalyzed by TA and PDC
2.4. Effect of Temperature on 3-APB Synthesis
2.5. Effect of Amine Donor Concentration on 3-APB Synthesis
2.6. Enantiomeric Excess Determination
3. Materials and Methods
3.1. Chemicals and Enzymes
3.2. Transaminase Activity Assay
3.3. Pyruvate Decarboxylase Activity Assay
3.4. Influence of pH on Enzyme Activity and Stability
3.5. Transaminase Screening
3.6. Asymmetric Synthesis of APB with Transaminase and Pyruvate Decarboxylase
3.7. Temperature and Amine Donor Concentration Effect on 3-APB Synthesis
3.8. Quantification of 3-APB and 4-PB
3.9. Enantiomeric Excess Determination
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jacques, J.; Collet, A.; Wilen, S.H. Enantiomers, Racemates and Resolutions; John Wiley & Sons, Ltd.: New York, NY, USA, 1981; ISBN 0471080586. [Google Scholar]
- Whitesell, J.K. C2 symmetry and asymmetric induction. Chem. Rev. 1989, 89, 1581–1590. [Google Scholar] [CrossRef]
- Castagnetto, J.; Canary, J.W. A chiroptically enhanced fluorescent chemosensor. Chem. Commun. 1998, 203–204. [Google Scholar] [CrossRef]
- Arrasate, S.; Lete, E.; Sotomayor, N. Synthesis of enantiomerically enriched amines by chiral ligand mediated addition of organolithium reagents to imines. Tetrahedron Asymmetry 2001, 12, 2077–2082. [Google Scholar] [CrossRef]
- Nugent, T.; El-Shazly, M. Chiral Amine Synthesis—Recent Developments and Trends for Enamide Reduction, Reductive Amination, and Imine Reduction. Adv. Synth. Catal. 2010, 352, 753–819. [Google Scholar] [CrossRef]
- Breuer, M.; Ditrich, K.; Habicher, T.; Hauer, B.; Keßeler, M.; Stürmer, R.; Zelinski, T. Industrial Methods for the Production of Optically Active Intermediates. Angew. Chem. Int. Ed. 2004, 43, 788–824. [Google Scholar] [CrossRef] [PubMed]
- Ghislieri, D.; Turner, N.J. Biocatalytic Approaches to the Synthesis of Enantiomerically Pure Chiral Amines. Top. Catal. 2013, 57, 284–300. [Google Scholar] [CrossRef]
- Kelly, S.A.; Pohle, S.; Wharry, S.; Mix, S.; Allen, C.; Moody, T.S.; Gilmore, B.F. Application of ω-Transaminases in the Pharmaceutical Industry. Chem. Rev. 2017, 118, 349–367. [Google Scholar] [CrossRef]
- Hayashi, H.; Uchida, T. Nitrene Transfer Reactions for Asymmetric C-H Amination: Recent Development. Eur. J. Org. Chem. 2019, 2020, 909–916. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.-S.; Gong, L.-Z. Palladium-Catalyzed Asymmetric Allylic C–H Functionalization: Mechanism, Stereo- and Regioselectivities, and Synthetic Applications. Acc. Chem. Res. 2020, 53, 2841–2854. [Google Scholar] [CrossRef]
- Abdine, R.A.A.; Hedouin, G.; Colobert, F.; Wencel-Delord, J. Metal-Catalyzed Asymmetric Hydrogenation of C═N Bonds. ACS Catal. 2020, 11, 215–247. [Google Scholar] [CrossRef]
- Busto, E.; Gotor-Fernández, V.; Gotor, V. Hydrolases in the Stereoselective Synthesis of N-Heterocyclic Amines and Amino Acid Derivatives. Chem. Rev. 2011, 111, 3998–4035. [Google Scholar] [CrossRef]
- Mathew, S.; Yun, H. ω-Transaminases for the Production of Optically Pure Amines and Unnatural Amino Acids. ACS Catal. 2012, 2, 993–1001. [Google Scholar] [CrossRef]
- Kroutil, W.; Fischereder, E.-M.; Fuchs, C.S.; Lechner, H.; Mutti, F.G.; Pressnitz, D.; Rajagopalan, A.; Sattler, J.H.; Simon, R.C.; Siirola, E. Asymmetric preparation of prim-, sec-, and tert-amines employing selected biocatalysts. Org. Process. Res. Dev. 2013, 17, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Green, A.P.; Turner, N.J.; O’Reilly, E. Chiral Amine Synthesis Using ω-Transaminases: An Amine Donor that Displaces Equilibria and Enables High-Throughput Screening. Angew. Chem. Int. Ed. 2014, 53, 10714–10717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomm, A.; O’Reilly, E. Transaminases for chiral amine synthesis. Curr. Opin. Chem. Biol. 2018, 43, 106–112. [Google Scholar] [CrossRef]
- Börner, T.; Rämisch, S.; Reddem, E.R.; Bartsch, S.; Vogel, A.; Thunnissen, A.-M.; Adlercreutz, P.; Grey, C. Explaining Operational Instability of Amine Transaminases: Substrate-Induced Inactivation Mechanism and Influence of Quaternary Structure on Enzyme–Cofactor Intermediate Stability. ACS Catal. 2017, 7, 1259–1269. [Google Scholar] [CrossRef]
- Chen, S.; Brocal, J.C.C.; Berglund, P.; Humble, M.S. Characterization of the stability of Vibrio fluvialis JS17 amine transaminase. J. Biotechnol. 2018, 282, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Berglund, P. Transaminase biocatalysis: Optimization and application. Green Chem. 2016, 19, 333–360. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.S.; Park, E.-S.; Shin, J.-S. Features and technical applications of ω-transaminases. Appl. Microbiol. Biotechnol. 2012, 94, 1163–1171. [Google Scholar] [CrossRef]
- Slabu, I.; Galman, J.; Lloyd, R.C.; Turner, N.J. Discovery, Engineering, and Synthetic Application of Transaminase Biocatalysts. ACS Catal. 2017, 7, 8263–8284. [Google Scholar] [CrossRef]
- Patil, M.D.; Grogan, G.; Bommarius, A.; Yun, H. Recent Advances in ω-Transaminase-Mediated Biocatalysis for the Enantioselective Synthesis of Chiral Amines. Catalysts 2018, 8, 254. [Google Scholar] [CrossRef] [Green Version]
- Koszelewski, D.; Tauber, K.; Faber, K.; Kroutil, W. ω-Transaminases for the synthesis of non-racemic α-chiral primary amines. Trends Biotechnol. 2010, 28, 324–332. [Google Scholar] [CrossRef]
- Kaulmann, U.; Smithies, K.; Smith, M.E.; Hailes, H.; Ward, J. Substrate spectrum of ω-transaminase from Chromobacterium violaceum DSM30191 and its potential for biocatalysis. Enzym. Microb. Technol. 2007, 41, 628–637. [Google Scholar] [CrossRef]
- Petri, A.; Colonna, V.; Piccolo, O. Asymmetric synthesis of a high added value chiral amine using immobilized ω-transaminases. Beilstein J. Org. Chem. 2019, 15, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Höhne, M.; Kühl, S.; Robins, K.; Bornscheuer, U. Efficient Asymmetric Synthesis of Chiral Amines by Combining Transaminase and Pyruvate Decarboxylase. ChemBioChem 2008, 9, 363–365. [Google Scholar] [CrossRef]
- Tufvesson, P.; Lima-Ramos, J.; Jensen, J.S.; Al-Haque, N.; Neto, W.L.A.; Woodley, J. Process considerations for the asymmetric synthesis of chiral amines using transaminases. Biotechnol. Bioeng. 2011, 108, 1479–1493. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Kim, B. Asymmetric synthesis of chiral amines with ω-transaminase. Biotechnol. Bioeng. 1999, 65, 206. [Google Scholar] [CrossRef]
- Schell, U.; Wohlgemuth, R.; Ward, J.M. Synthesis of pyridoxamine 5′-phosphate using an MBA:pyruvate transaminase as biocatalyst. J. Mol. Catal. B Enzym. 2009, 59, 279–285. [Google Scholar] [CrossRef]
- Humble, M.S.; Cassimjee, K.E.; Håkansson, M.; Kimbung, Y.R.; Walse, B.; Abedi, V.; Federsel, H.-J.; Berglund, P.; Logan, D.T. Crystal structures of the Chromobacterium violaceum ω-transaminase reveal major structural rearrangements upon binding of coenzyme PLP. FEBS J. 2012, 279, 779–792. [Google Scholar] [CrossRef] [PubMed]
- Raj, K.C.; Talarico, L.A.; Ingram, L.O.; Maupin-Furlow, J.A. Cloning and Characterization of the Zymobacter palmae Pyruvate Decarboxylase Gene (pdc) and Comparison to Bacterial Homologues. Appl. Environ. Microbiol. 2002, 68, 2869–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassimjee, K.E.; Humble, M.S.; Miceli, V.; Colomina, C.G.; Berglund, P. Active Site Quantification of an ω-Transaminase by Performing a Half Transamination Reaction. ACS Catal. 2011, 1, 1051–1055. [Google Scholar] [CrossRef]
- Chen, S.; Land, H.; Berglund, P.; Humble, M.S. Stabilization of an amine transaminase for biocatalysis. J. Mol. Catal. B Enzym. 2016, 124, 20–28. [Google Scholar] [CrossRef]
- Mutti, F.G.; Fuchs, C.S.; Pressnitz, D.; Turrini, N.G.; Sattler, J.H.; Lerchner, A.; Skerra, A.; Kroutil, W. Amination of Ketones by Employing Two New (S)-Selective ω-Transaminases and the His-Tagged ω-TA from Vibrio fluvialis. Eur. J. Org. Chem. 2011, 2012, 1003–1007. [Google Scholar] [CrossRef]
- Koszelewski, D.; Göritzer, M.; Clay, D.; Seisser, B.; Kroutil, W. Synthesis of optically active amines employing recombinant ω-transaminases in E. coli cells. ChemCatChem 2010, 2, 73–77. [Google Scholar] [CrossRef]
- Koszelewski, D.; Lavandera, I.; Clay, D.; Rozzell, D.; Kroutil, W. Asymmetric Synthesis of Optically Pure Pharmacologically Relevant Amines Employing ω-Transaminases. Adv. Synth. Catal. 2008, 350, 2761–2766. [Google Scholar] [CrossRef]
- Gundersen, M.T.; Abu, R.; Schürmann, M.; Woodley, J.M. Amine donor and acceptor influence on the thermodynamics of ω-transaminase reactions. Tetrahedron Asymmetry 2015, 26, 567–570. [Google Scholar] [CrossRef]
- Alcover, N.; Carceller, A.; Alvaro, G.; Guillén, M. Zymobacter palmae pyruvate decarboxylase production process development: Cloning in Escherichia coli, fed-batch culture and purification. Eng. Life Sci. 2019, 19, 502–512. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transaminase | PDC | pH | [APB] (mM) |
|---|---|---|---|
| CviTA | + | 7.5 | 6.2 |
| − | 0.6 | ||
| VflTA | + | 7.5 | 5.9 |
| − | 1.0 |
| Transaminase | PDC | Final [APB] (mM) | Conversion (%) | Yield (%) | Selectivity (%) | STY (mmol·L−1·h−1) |
|---|---|---|---|---|---|---|
| CviTA | Yes | 6.20 ± 0.98 | 72.0 ± 3.1 | 64.2 ± 10.1 | 88.7 ± 10.2 | 0.78 ± 0.12 |
| No | 0.58 ± 0.00 | 23.0 ± 9.5 | 6.1 ± 1.0 | 34.1 ± 18.3 | 0.07 ± 0.00 | |
| VflTA | Yes | 5.92 ± 0.64 | 71.0 ± 3.0 | 60.8 ± 6.6 | 86.1 ± 12.9 | 0.74 ± 0.08 |
| No | 1.01 ± 0.05 | 22.9 ± 8.8 | 10.5 ± 2.0 | 57.4 ± 30.8 | 0.13 ± 0.01 |
| Transaminase | Temperature (°C) | Reaction Time (h) | Final APB (mM) | Conversion (%) | Yield (%) | Selectivity (%) | STY (mmol·L−1·h−1) |
|---|---|---|---|---|---|---|---|
| CviTA | 15 | 24 | 7.68 ± 0.05 | 73.7 ± 0.5 | 75.4 ± 0.5 | 102.3 ± 1.4 | 0.32 ± 0.00 |
| 30 | 8 | 6.20 ± 0.98 | 72.0 ± 3.1 | 64.2 ± 10.1 | 88.7 ± 10.2 | 0.78 ± 0.12 | |
| 40 | 8 | 3.08 ± 0.54 | 61.7 ± 1.3 | 30.2 ± 5.3 | 48.8 ± 7.6 | 0.38 ± 0.07 | |
| VflTA | 15 | 48 | 5.92 ± 0.50 | 71.8 ± 1.3 | 52.2 ± 4.4 | 80.9 ± 5.4 | 0.12 ± 0.01 |
| 30 | 8 | 5.92 ± 0.64 | 71.0 ± 3.0 | 60.8 ± 6.6 | 86.1 ± 12.9 | 0.74 ± 0.08 | |
| 40 | 8 | 2.30 ± 0.45 | 48.4 ± 2.1 | 24.1 ± 4.7 | 46.5 ± 7.1 | 0.29 ± 0.06 |
| Transaminase | Initial L-Alanine (mM) | Final APB (mM) | Conversion (%) | Yield (%) | Selectivity (%) | STY (mmol·L−1·h−1) |
|---|---|---|---|---|---|---|
| CviTA | 200 | 6.20 ± 0.98 | 72.0 ± 3.1 | 64.2 ± 10.1 | 88.7 ± 10.2 | 0.78 ± 0.12 |
| 400 | 6.22 ± 0.35 | 76.4 ± 3.6 | 59.5 ± 3.3 | 88.7 ± 10.2 | 0.78 ± 0.12 | |
| 600 | 6.88 ± 0.06 | 77.8 ± 1.6 | 71.7 ± 1.6 | 48.8 ± 7.6 | 0.38 ± 0.07 | |
| VflTA | 200 | 5.92 ± 0.64 | 71.0 ± 3.0 | 60.8 ± 6.6 | 86.1 ± 12.9 | 0.74 ± 0.08 |
| 400 | 6.60 ± 0.25 | 78.4 ± 4.4 | 68.4 ± 2.6 | 87.8 ± 8.2 | 0.82 ± 0.03 | |
| 600 | 7.08 ± 0.18 | 80.0 ± 6.2 | 73.2 ± 2.0 | 91.8 ± 4.6 | 0.89 ± 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alcover, N.; Álvaro, G.; Guillén, M. Chiral Synthesis of 3-Amino-1-phenylbutane by a Multi-Enzymatic Cascade System. Catalysts 2021, 11, 973. https://doi.org/10.3390/catal11080973
Alcover N, Álvaro G, Guillén M. Chiral Synthesis of 3-Amino-1-phenylbutane by a Multi-Enzymatic Cascade System. Catalysts. 2021; 11(8):973. https://doi.org/10.3390/catal11080973
Chicago/Turabian StyleAlcover, Natàlia, Gregorio Álvaro, and Marina Guillén. 2021. "Chiral Synthesis of 3-Amino-1-phenylbutane by a Multi-Enzymatic Cascade System" Catalysts 11, no. 8: 973. https://doi.org/10.3390/catal11080973
APA StyleAlcover, N., Álvaro, G., & Guillén, M. (2021). Chiral Synthesis of 3-Amino-1-phenylbutane by a Multi-Enzymatic Cascade System. Catalysts, 11(8), 973. https://doi.org/10.3390/catal11080973