
Ultrafiltration of Saithe (Pollachius virens) Protein Hydrolysates and Its Effect on Antioxidative Activity

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
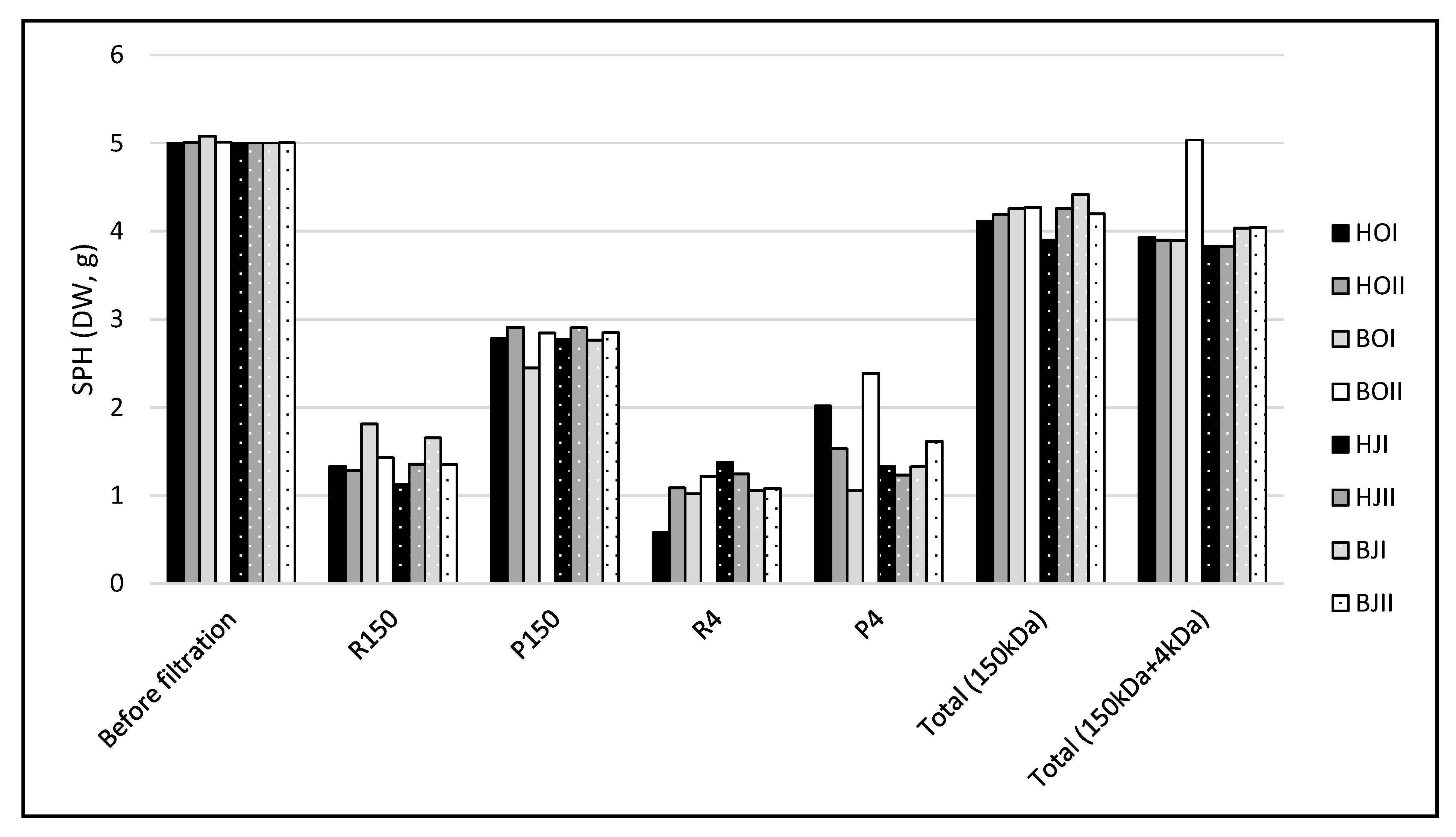
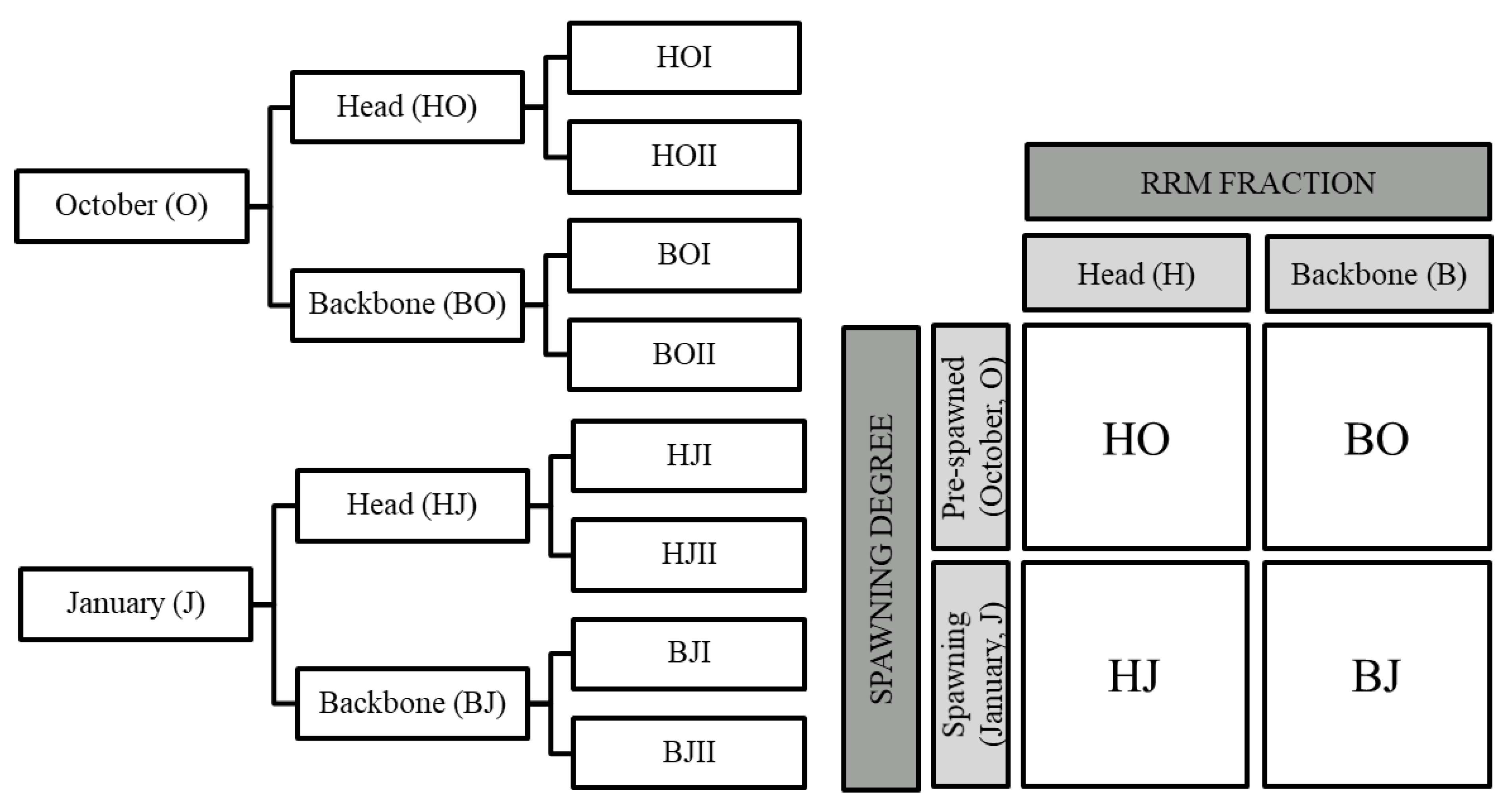
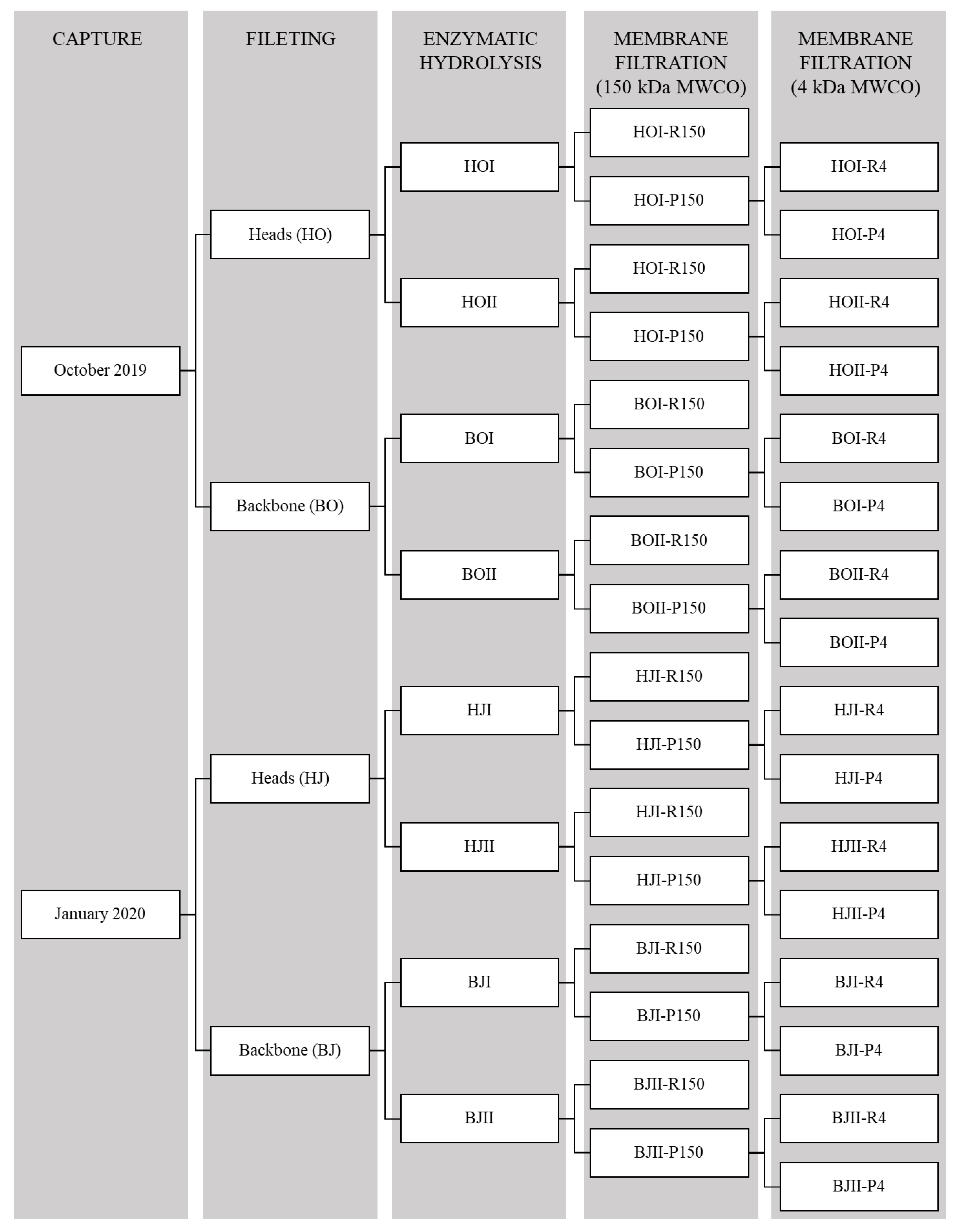
2.1. Membrane Filtration and Protein Flow
2.2. Amino Acid Composition
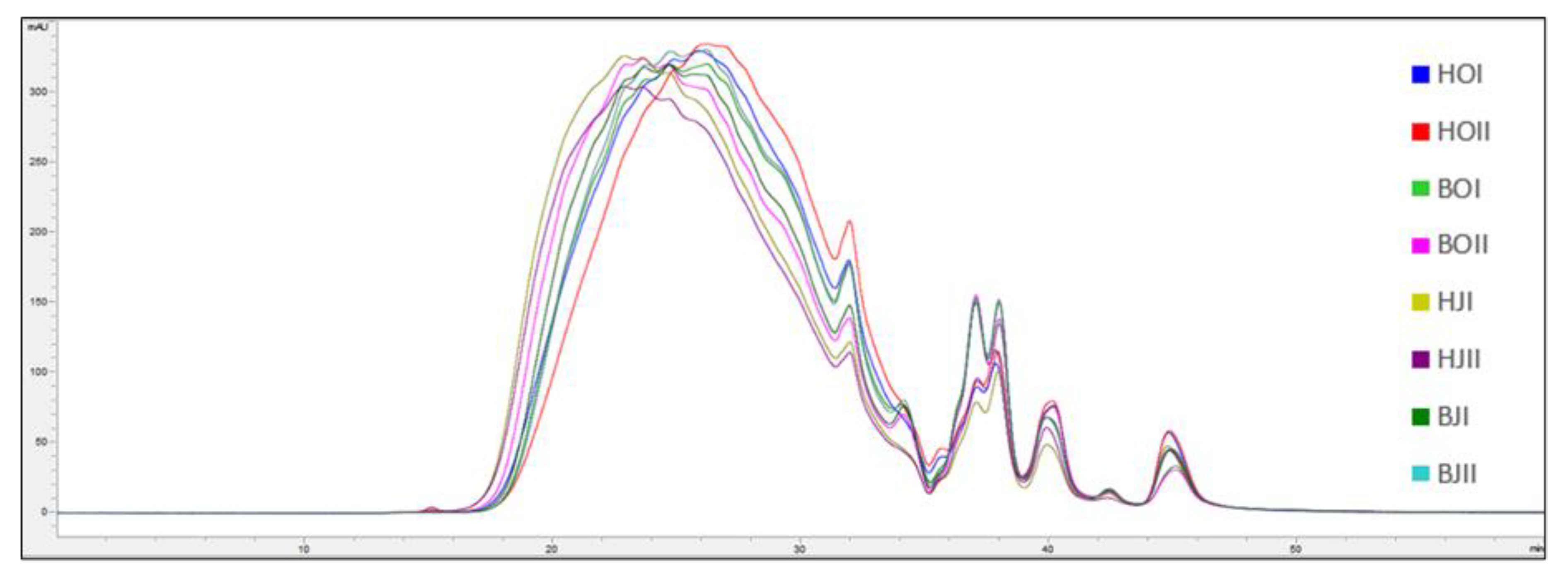
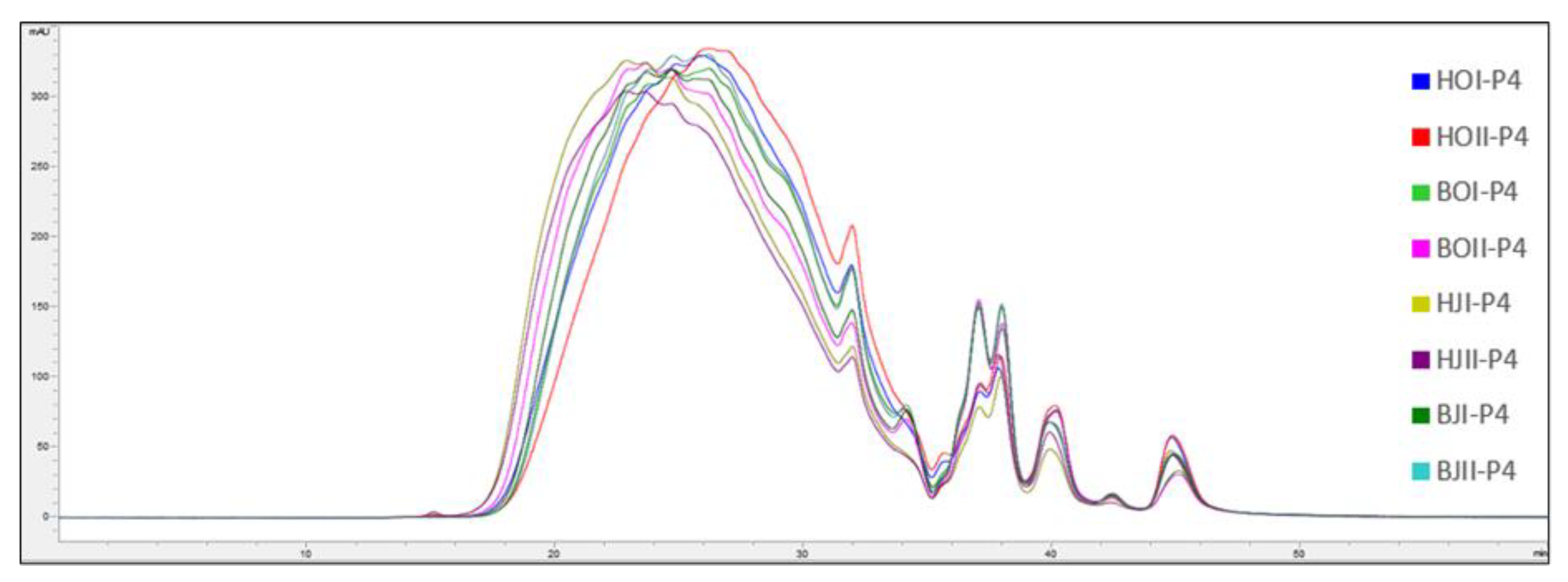
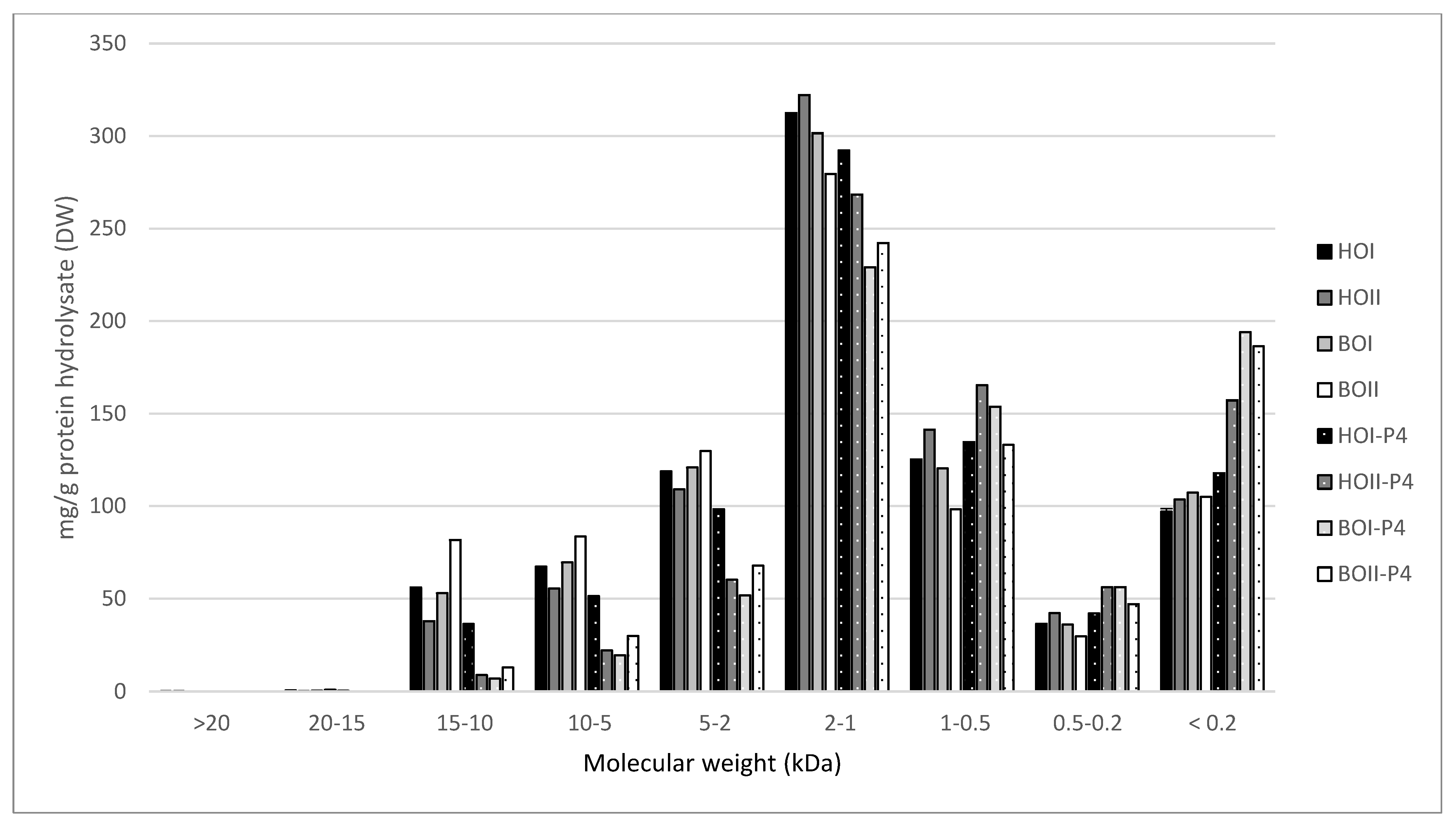
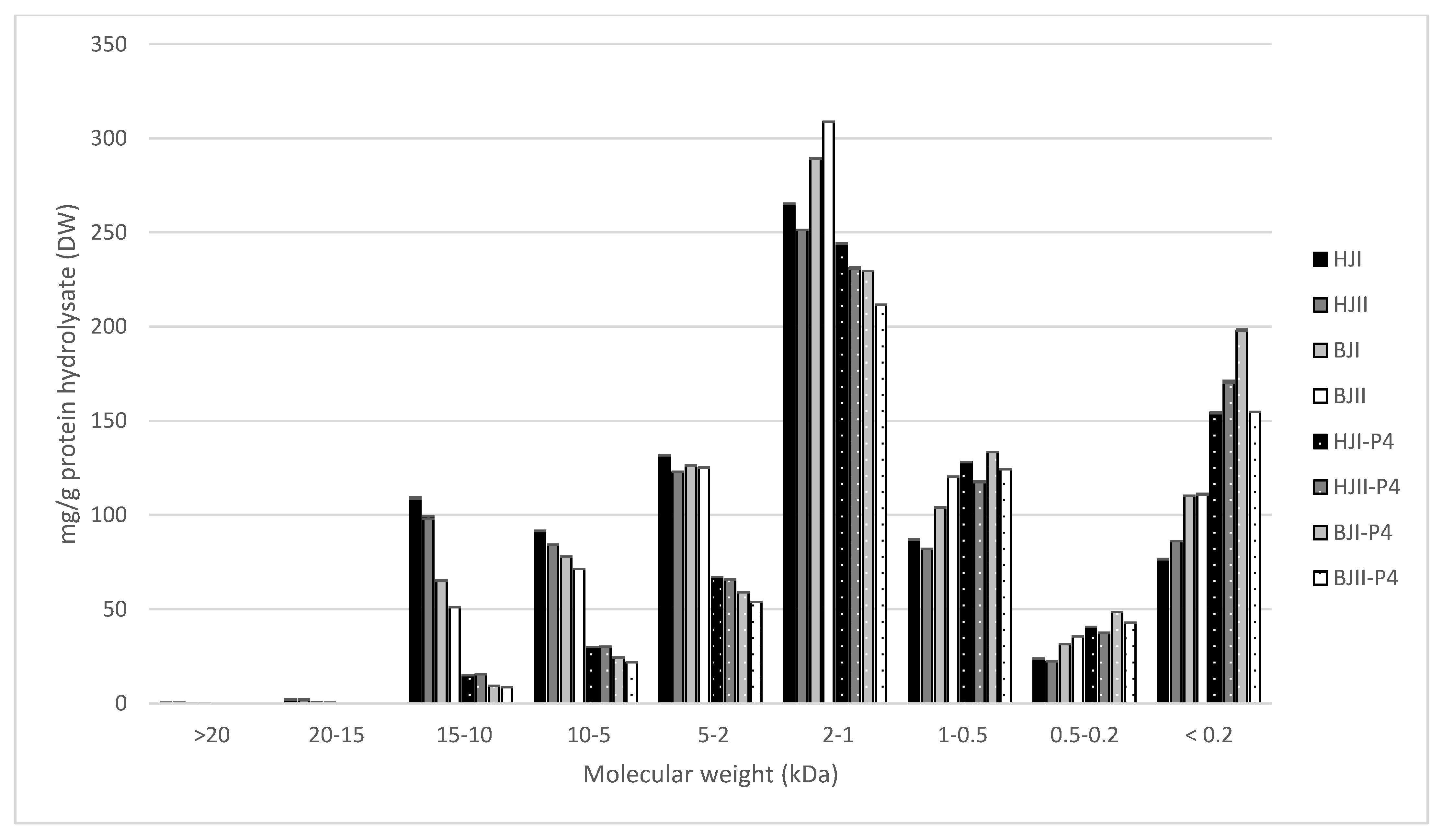
2.3. Molecular Weight Distribution
2.4. Antioxidative Activity
3. Materials and Methods
3.1. Chemicals
3.2. Enzymatic Hydrolysis
3.3. Membrane Ultrafiltration
3.4. Dry Matter
3.5. Antioxidative Assays
3.6. Amino Acid Composition
3.7. Protein Efficiency Ratio (PER) Value
3.8. Molecular Weight Distribution
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghaly, A.; Ramakrishnan, V.; Brooks, M.; Budge, S.; Dave, D. Fish Processing Wastes as a Potential Source of Proteins, Amino Acids and Oils: A Critical Review. J. Microb. Biochem. Technol. 2013, 5, 107–129. [Google Scholar] [CrossRef] [Green Version]
- Richardsen, R.; Myhre, M.; Nystøyl, R.; Strandheim, G.; Marthinussen, A. Analyse Marint Restråstoff 2018. Tilgang og Anvendelse av Marint Restråstoff i Norge; 2019:00475; SINTEF Ocean AS: Oslo, Norway, 2019; p. 47. [Google Scholar]
- Hjellnes, V.; Rustad, T.; Falch, E. Enzymatic hydrolysis of pre-spawned and spawning saithe (Pollachius virens) in bioreactors and its potential for implementation in the Norwegian whitefish industry. 2021; submitted for publication. [Google Scholar]
- Jafarpour, A.; Gomes, R.M.; Gregersen, S.; Sloth, J.J.; Jacobsen, C.; Sørensen, A.-D.M. Characterization of cod (Gadus morhua) frame composition and its valorization by enzymatic hydrolysis. J. Food Compos. Anal. 2020, 89, 103469. [Google Scholar] [CrossRef]
- Hou, H.; Li, B.; Zhao, X.; Zhang, Z.; Li, P. Optimization of enzymatic hydrolysis of Alaska pollock frame for preparing protein hydrolysates with low-bitterness. LWT 2011, 44, 421–428. [Google Scholar] [CrossRef]
- Remme, J.F.; Austnes, A. Kvalitetsproteiner fra Torskehoder—HEADS UP II; 2020:00322; SINTEF Ocean AS: Oslo, Norway, 2020; p. 21. [Google Scholar]
- Rustad, T. Utilisation of marine by-products. Elec. J. Env. Agricult. Food Chem. 2003, 2, 458–463. [Google Scholar]
- Hoegh-Guldberg, O. The Ocean as a Solution to Climate Change: Five Opportunities for Action; World Resource Institute: Washington, DC, USA, 2019. [Google Scholar]
- Kim, S.-K.; Mendis, E. Bioactive compounds from marine processing byproducts—A review. Food Res. Int. 2006, 39, 383–393. [Google Scholar] [CrossRef]
- Wijesekara, I.; Kim, S.-K. Angiotensin-I-Converting Enzyme (ACE) Inhibitors from Marine Resources: Prospects in the Pharmaceutical Industry. Mar. Drugs 2010, 8, 1080. [Google Scholar] [CrossRef] [Green Version]
- Guérard, F.; Sellos, D.; Le Gal, Y. Fish and Shellfish Upgrading, Traceability. In Marine Biotechnology; Ulber, R., Le Gal, Y., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 127–163. [Google Scholar]
- Jensen, I.-J.; Mæhre, H.K. Preclinical and clinical studies on antioxidative, antihypertensive and cardioprotective effect of marine proteins and peptides—A review. Mar. Drugs 2016, 14, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dale, H.F.; Madsen, L.; Lied, G.A. Fish–derived proteins and their potential to improve human health. Nutr. Rev. 2019, 77, 572–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girgih, A.T.; He, R.; Hasan, F.M.; Udenigwe, C.; Gill, T.A.; Aluko, R.E. Evaluation of the in vitro antioxidant properties of a cod (Gadus morhua) protein hydrolysate and peptide fractions. Food Chem. 2014, 173, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Farvin, K.S.; Andersen, L.L.; Otte, J.; Nielsen, H.H.; Jessen, F.; Jacobsen, C. Antioxidant activity of cod (Gadus morhua) protein hydrolysates: Fractionation and characterisation of peptide fractions. Food Chem. 2016, 204, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Picot, L.; Ravallec, R.; Fouchereau-Péron, M.; Vandanjon, L.; Jaouen, P.; Chaplain-Derouiniot, M.; Guérard, F.; Chabeaud, A.; LeGal, Y.; Alvarez, M.; et al. Impact of ultrafiltration and nanofiltration of an industrial fish protein hydrolysate on its bioactive properties. J. Sci. Food Agric. 2010, 90, 1819–1826. [Google Scholar] [CrossRef] [Green Version]
- Ngo, D.-H.; Vo, T.-S.; Ryu, B.; Kim, S.-K. Angiotensin- I- converting enzyme (ACE) inhibitory peptides from Pacific cod skin gelatin using ultrafiltration membranes. Process. Biochem. 2016, 51, 1622–1628. [Google Scholar] [CrossRef] [Green Version]
- Ween, O.; Stangeland, J.K.; Fylling, T.S.; Aas, G.H. Nutritional and functional properties of fishmeal produced from fresh by-products of cod (Gadus morhua L.) and saithe (Pollachius virens). Heliyon 2017, 3, e00343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, I.-J.; Eysturskarð, J.; Madetoja, M.; Eilertsen, K.-E. The potential of cod hydrolyzate to inhibit blood pressure in spontaneously hypertensive rats. Nutr. Res. 2014, 34, 168–173. [Google Scholar] [CrossRef]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Pihlanto, A. Antioxidative peptides derived from milk proteins. Int. Dairy J. 2006, 16, 1306–1314. [Google Scholar] [CrossRef]
- Gulcin, I. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [Green Version]
- Branen, A.L. Toxicology and biochemistry of butylated hydroxyanisole and butylated hydroxytoluene. J. Am. Oil Chem. Soc. 1975, 52, 59–63. [Google Scholar] [CrossRef]
- Huang, D.; Ou, .A.B.; Prior, R.L. The Chemistry behind Antioxidant Capacity Assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- FAO. The Future of Food and Agriculture—Trends And Challenges; FAO: Rome, Italy, 2017. [Google Scholar]
- Udenigwe, C.; Aluko, R.E. Food Protein-Derived Bioactive Peptides: Production, Processing, and Potential Health Benefits. J. Food Sci. 2011, 77, R11–R24. [Google Scholar] [CrossRef]
- Kaur, N.; Sharma, P.; Jaimni, S.; Kehinde, B.A.; Kaur, S. Recent developments in purification techniques and industrial applications for whey valorization: A review. Chem. Eng. Commun. 2019, 207, 123–138. [Google Scholar] [CrossRef]
- Gao, R.; Yu, Q.; Shen, Y.; Chu, Q.; Chen, G.; Fen, S.; Yang, M.; Yuan, L.; McClements, D.J.; Sun, Q. Production, bioactive properties, and potential applications of fish protein hydrolysates: Developments and challenges. Trends Food Sci. Technol. 2021, 110, 687–699. [Google Scholar] [CrossRef]
- Hjellnes, V.; Rustad, T.; Falch, E. The value chain of the white fish industry in Norway: History, current status and possibilities for improvement—A review. Reg. Stud. Mar. Sci. 2020, 36, 101293. [Google Scholar] [CrossRef]
- Bacchin, P.; Aimar, P.; Field, R. Critical and sustainable fluxes: Theory, experiments and applications. J. Membr. Sci. 2006, 281, 42–69. [Google Scholar] [CrossRef] [Green Version]
- Araujo, J.; Sica, P.; Costa, C.; Márquez, M.C. Enzymatic Hydrolysis of Fish Waste as an Alternative to Produce High Value-Added Products. Waste Biomass Valorization 2020, 12, 847–855. [Google Scholar] [CrossRef]
- Gildberg, A.; A Arnesen, J.; Carlehög, M. Utilisation of cod backbone by biochemical fractionation. Process. Biochem. 2002, 38, 475–480. [Google Scholar] [CrossRef]
- Karami, Z.; Akbari-Adergani, B. Bioactive food derived peptides: A review on correlation between structure of bioactive peptides and their functional properties. J. Food Sci. Technol. 2019, 56, 535–547. [Google Scholar] [CrossRef]
- Jensen, I.-J.; Larsen, R.; Rustad, T.; Eilertsen, K.-E. Nutritional content and bioactive properties of wild and farmed cod (Gadus morhua L.) subjected to food preparation. J. Food Compos. Anal. 2013, 31, 212–216. [Google Scholar] [CrossRef]
- Coultate, T.P. Food: The Chemistry of its Components; Royal Society of Chemistry: London, UK, 2009. [Google Scholar]
- Branden, C.I.; Tooze, J. The Building Blocks. In Introduction to Protein Structure, 12nd ed.; Garland Science: New York, NY, USA, 2012; pp. 1–12. [Google Scholar]
- Slizyte, R.; Daukšas, E.; Falch, E.; Storrø, I.; Rustad, T. Characteristics of protein fractions generated from hydrolysed cod (Gadus morhua) by-products. Process. Biochem. 2005, 40, 2021–2033. [Google Scholar] [CrossRef]
- HoleČek, M. Histidine in Health and Disease: Metabolism, Physiological Importance, and Use as a Supplement. Nutrients 2020, 12, 848. [Google Scholar] [CrossRef] [Green Version]
- Rajapakse, N.; Mendis, E.; Jung, W.-K.; Je, J.-Y.; Kim, S.-K. Purification of a radical scavenging peptide from fermented mussel sauce and its antioxidant properties. Food Res. Int. 2005, 38, 175–182. [Google Scholar] [CrossRef]
- Marcuse, R. The effect of some amino acids on the oxidation of linoleic acid and its methyl ester. J. Am. Oil Chem. Soc. 1962, 39, 97–103. [Google Scholar] [CrossRef]
- Chen, H.-M.; Muramoto, K.; Yamauchi, F.; Fujimoto, K.; Nokihara, K. Antioxidative Properties of Histidine-Containing Peptides Designed from Peptide Fragments Found in the Digests of a Soybean Protein. J. Agric. Food Chem. 1998, 46, 49–53. [Google Scholar] [CrossRef]
- Siemensma, A.D.; Weijer, W.J.; Bak, H.J. The importance of peptide lengths in hypoallergenic infant formulae. Trends Food Sci. Technol. 1993, 4, 16–21. [Google Scholar] [CrossRef]
- Chabeaud, A.; Vandanjon, L.; Bourseau, P.; Jaouen, P.; Chaplain-Derouiniot, M.; Guerard, F. Performances of ultrafiltration membranes for fractionating a fish protein hydrolysate: Application to the refining of bioactive peptidic fractions. Sep. Purif. Technol. 2009, 66, 463–471. [Google Scholar] [CrossRef]
- Chabeaud, A.; Vandanjon, L.; Bourseau, P.; Jaouen, P.; Guérard, F. Fractionation by ultrafiltration of a saithe protein hydrolysate (Pollachius virens): Effect of material and molecular weight cut-off on the membrane performances. J. Food Eng. 2009, 91, 408–414. [Google Scholar] [CrossRef]
- Damodaran, S.; Parkin, K.L.; Fennema, O.R. Amino acids, peptides, and proteins. In Fennema’s Food Chemistry, 4th ed.; Damodaran, S., Ed.; CRC Press: Boca Raton, FL, USA, 2008; pp. 217–331. [Google Scholar]
- Jeon, Y.-J.; Byun, H.-G.; Kim, S.-K. Improvement of functional properties of cod frame protein hydrolysates using ultrafiltration membranes. Process. Biochem. 1999, 35, 471–478. [Google Scholar] [CrossRef]
- Dávalos, A.; Gómez-Cordovés, A.C.; Bartolomé, B. Extending Applicability of the Oxygen Radical Absorbance Capacity (ORAC−Fluorescein) Assay. J. Agric. Food Chem. 2003, 52, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Saiga, A.; Tanabe, S.; Nishimura, T. Antioxidant activity of peptides obtained from porcine myofibrillar proteins by protease treatment. J. Agric. Food Chem. 2003, 51, 3661–3667. [Google Scholar] [CrossRef]
- Kristinová, V.; Mozuraityte, R.; Storrø, I.; Rustad, T. Antioxidant Activity of Phenolic Acids in Lipid Oxidation Catalyzed by Different Prooxidants. J. Agric. Food Chem. 2009, 57, 10377–10385. [Google Scholar] [CrossRef]
- AOAC. Association of Official Analytical Chemists. Official Methods of Analysis, 15th ed.; AOAC: Arlington, VA, USA; Washington, DC, USA, 1990. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free. Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Nenadis, N.; Wang, L.-F.; Tsimidou, M.; Zhang, H.-Y. Estimation of Scavenging Activity of Phenolic Compounds Using the ABTS•+Assay. J. Agric. Food Chem. 2004, 52, 4669–4674. [Google Scholar] [CrossRef] [PubMed]
- Nenadis, N.; Lazaridou, A.O.; Tsimidou, M. Use of Reference Compounds in Antioxidant Activity Assessment. J. Agric. Food Chem. 2007, 55, 5452–5460. [Google Scholar] [CrossRef]
- Benzie, I.; Strain, J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, I.-J.; Abrahamsen, H.; Maehre, H.K.; Elvevoll, E.O. Changes in Antioxidative Capacity of Saithe (Pollachius virens) and Shrimp (Pandalus borealis) duringin VitroDigestion. J. Agric. Food Chem. 2009, 57, 10928–10932. [Google Scholar] [CrossRef]
- Blackburn, S. Amino Acid Determination. Methods and Techniques; Edwars Arnold Ltd.: London, UK, 1968; p. 271. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BJII | BJI | HJII | HJI | BOII | BOI | HOII | HOI | Backbone January (RRM) | Head January (RRM) | Backbone October (RRM) | Head October (RRM) | AA |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 6.08 ± 0.59 | 5.91 ± 1.59 | 8.18 ± 0.15 | 7.53 ± 0.36 | 5.45 ± 1.22 | 5.34 ± 0.59 | 8.18 ± 0.32 | 6.68 ± 0.10 | 9.63 ± 0.49 | 10.01 ± 0.13 | 11.57 ± 0.73 | 7.88 ± 0.31 | Asp h |
| 8.93 ± 0.91 | 8.86 ± 2.45 | 12.35 ± 0.26 | 11.48 ± 0.59 | 7.73 ± 1.75 | 7.56 ± 0.85 | 12.19 ± 0.43 | 9.78 ± 0.18 | 14.47 ± 0.89 | 15.30 ± 0.18 | 17.13 ± 1.16 | 11.98 ± 0.51 | Glu |
| 0.02 ± 0.02 | 0.02 ± 0.02 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.01 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.03 ± 0.02 | 0.04 ± 0.03 | 0.00 ± 0.00 | 0.11 ± 0.01 | 0.00 ± 0.00 | Asn |
| 0.80 ± 0.08 | 0.69 ± 0.31 | 1.07 ± 0.09 | 0.79 ± 0.08 | 0.52 ± 0.27 | 0.81 ± 0.10 | 0.85 ± 0.04 | 0.42 ± 0.14 | 1.23 ± 0.57 | 2.00 ± 0.04 | 0.13 ± 0.04 | 1.43 ± 0.07 | His e |
| 2.82 ± 0.36 | 2.56 ± 0.80 | 4.49 ± 0.07 | 4.32 ± 0.23 | 2.30 ± 0.53 | 2.35 ± 0.26 | 4.98 ± 0.24 | 3.71 ± 0.28 | 4.12 ± 0.44 | 5.86 ± 0.07 | 4.40 ± 0.30 | 4.38 ± 0.21 | Ser |
| 0.16 ± 0.05 | 0.19 ± 0.13 | 0.17 ± 0.01 | 0.15 ± 0.03 | 0.14 ± 0.03 | 0.20 ± 0.05 | 0.22 ± 0.00 | 0.12 ± 0.02 | 0.25 ± 0.08 | 0.31 ± 0.01 | 0.18 ± 0.05 | 0.20 ± 0.04 | Gln |
| 5.87 ± 0.61 | 6.02 ± 1.89 | 12.70 ± 0.27 | 13.00 ± 0.63 | 5.11 ± 1.20 | 5.03 ± 0.59 | 14.27 ± 0.53 | 11.01 ± 0.37 | 7.99 ± 0.63 | 17.04 ± 0.26 | 8.33 ± 0.34 | 11.71 ± 0.70 | Gly/Arg h |
| 2.61 ± 0.25 | 2.63 ± 0.81 | 4.03 ± 0.06 | 3.59 ± 0.39 | 2.34 ± 0.54 | 2.31 ± 0.28 | 4.12 ± 0.19 | 3.31 ± 0.15 | 4.44 ± 0.30 | 5.26 ± 0.06 | 5.28 ± 0.37 | 2.55 ± 1.28 | Thr e |
| 4.04 ± 0.42 | 4.06 ± 1.19 | 6.53 ± 0.15 | 6.26 ± 0.29 | 3.71 ± 0.83 | 3.61 ± 0.40 | 6.82 ± 0.34 | 5.52 ± 0.07 | 5.97 ± 0.30 | 8.58 ± 0.13 | 7.06 ± 0.35 | 6.30 ± 0.32 | Ala |
| 0.52 ± 0.07 | 0.28 ± 0.12 | 1.14 ± 0.23 | 1.16 ± 0.07 | 0.37 ± 0.23 | 0.63 ± 0.07 | 1.84 ± 0.03 | 0.50 ± 0.47 | 1.75 ± 0.86 | 2.52 ± 0.06 | 0.02 ± 0.00 | 2.20 ± 0.15 | Tyr ce |
| 1.56 ± 0.15 | 1.27 ± 0.45 | 2.53 ± 0.07 | 2.35 ± 0.00 | 1.24 ± 0.35 | 1.38 ± 0.15 | 2.59 ± 0.06 | 1.93 ± 0.17 | 2.84 ± 0.45 | 3.25 ± 0.05 | 2.92 ± 0.23 | 2.61 ± 0.09 | Met e,h |
| 2.48 ± 0.23 | 2.44 ± 0.61 | 3.51 ± 0.08 | 3.22 ± 0.12 | 2.30 ± 0.52 | 2.23 ± 0.25 | 3.63 ± 0.15 | 2.94 ± 0.09 | 4.20 ± 0.22 | 4.25 ± 0.05 | 5.18 ± 0.30 | 3.46 ± 0.13 | Val e,h |
| 1.79 ± 0.18 | 1.56 ± 0.50 | 2.71 ± 0.11 | 2.52 ± 0.16 | 1.39 ± 0.30 | 1.49 ± 0.17 | 2.89 ± 0.11 | 2.10 ± 0.22 | 3.38 ± 0.54 | 3.83 ± 0.05 | 3.27 ± 0.30 | 3.23 ± 0.12 | Phe h |
| 1.99 ± 0.19 | 1.90 ± 0.48 | 2.44 ± 0.04 | 2.22 ± 0.13 | 1.74 ± 0.39 | 1.73 ± 0.19 | 2.54 ± 0.10 | 2.03 ± 0.05 | 3.89 ± 0.22 | 3.25 ± 0.05 | 4.69 ± 0.33 | 2.59 ± 0.09 | Ile e,h |
| 3.91 ± 0.37 | 3.75 ± 0.96 | 4.89 ± 0.09 | 4.39 ± 0.23 | 3.51 ± 0.78 | 3.44 ± 0.38 | 4.90 ± 0.18 | 3.99 ± 0.07 | 7.16 ± 0.53 | 5.86 ± 0.02 | 8.57 ± 0.57 | 4.81 ± 0.17 | Leu e,h |
| 5.95 ± 0.56 | 5.64 ± 1.57 | 6.57 ± 0.13 | 5.99 ± 0.28 | 5.16 ± 1.16 | 5.03 ± 0.54 | 6.47 ± 0.26 | 5.13 ± 0.10 | 7.88 ± 0.73 | 7.15 ± 0.14 | 9.07 ± 0.59 | 5.97 ± 0.21 | Lys e |
| 1.53 | 1.44 | 2.05 | 1.85 | 1.32 | 1.31 | 2.08 | 1.62 | 2.62 | 2.55 | 3.04 | 1.93 | PER * |
| Sample | ABTS (µmol/g) | FRAP (µmol/g) | ORAC (µmol/g) |
|---|---|---|---|
| Hydrolysate | - | - | - |
| H(SPH) | 58.33 ± 0.64 h3 | 5.16 ± 0.40 h1 | 380.50 ± 36.12 h1 |
| B(SPH) | 63.57 ± 1.20 b3 | 12.57 ± 0.40 b2 | 557.22 ± 72.47 b2 |
| Permeate | - | - | - |
| H(P4) | 42.38 ± 0.64 h1 | 8.06 ± 0.35 h2 | 394.97 ± 12.50 h1 |
| B(P4) | 55.37 ± 1.15 b1 | 10.70 ± 0.44 b1 | 547.79 ± 42.45 b2 |
| Retentate | - | - | - |
| H(R4) | 45.45 ± 0.94 h2 | 7.58 ± 0.35 h2 | |
| B(R4) | 59.72 ± 0.80 b2 | 12.55 ± 0.28 b2 | 354.39 ± 14.48 b1 |
| H(R150) | 42.23 ± 0.64 h1 | 14.70 ± 0.55 h3 | |
| B(R150) | 56.07 ± 0.66 b1,b2 | 15.55 ± 0.56 b3 | 331.12 ± 52.82 b1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hjellnes, V.; Rustad, T.; Jensen, I.-J.; Eiken, E.; Pettersen, S.M.; Falch, E. Ultrafiltration of Saithe (Pollachius virens) Protein Hydrolysates and Its Effect on Antioxidative Activity. Catalysts 2021, 11, 1053. https://doi.org/10.3390/catal11091053
Hjellnes V, Rustad T, Jensen I-J, Eiken E, Pettersen SM, Falch E. Ultrafiltration of Saithe (Pollachius virens) Protein Hydrolysates and Its Effect on Antioxidative Activity. Catalysts. 2021; 11(9):1053. https://doi.org/10.3390/catal11091053
Chicago/Turabian StyleHjellnes, Veronica, Turid Rustad, Ida-Johanne Jensen, Elin Eiken, Stine Marie Pettersen, and Eva Falch. 2021. "Ultrafiltration of Saithe (Pollachius virens) Protein Hydrolysates and Its Effect on Antioxidative Activity" Catalysts 11, no. 9: 1053. https://doi.org/10.3390/catal11091053