
C058 and Other Functional Microorganisms Promote the Synthesis of Extracellular Polymer Substances in Mycelium Biofloc

Abstract
:1. Introduction
2. Results and Discussion
2.1. Changes in EPS Content in the Biofloc
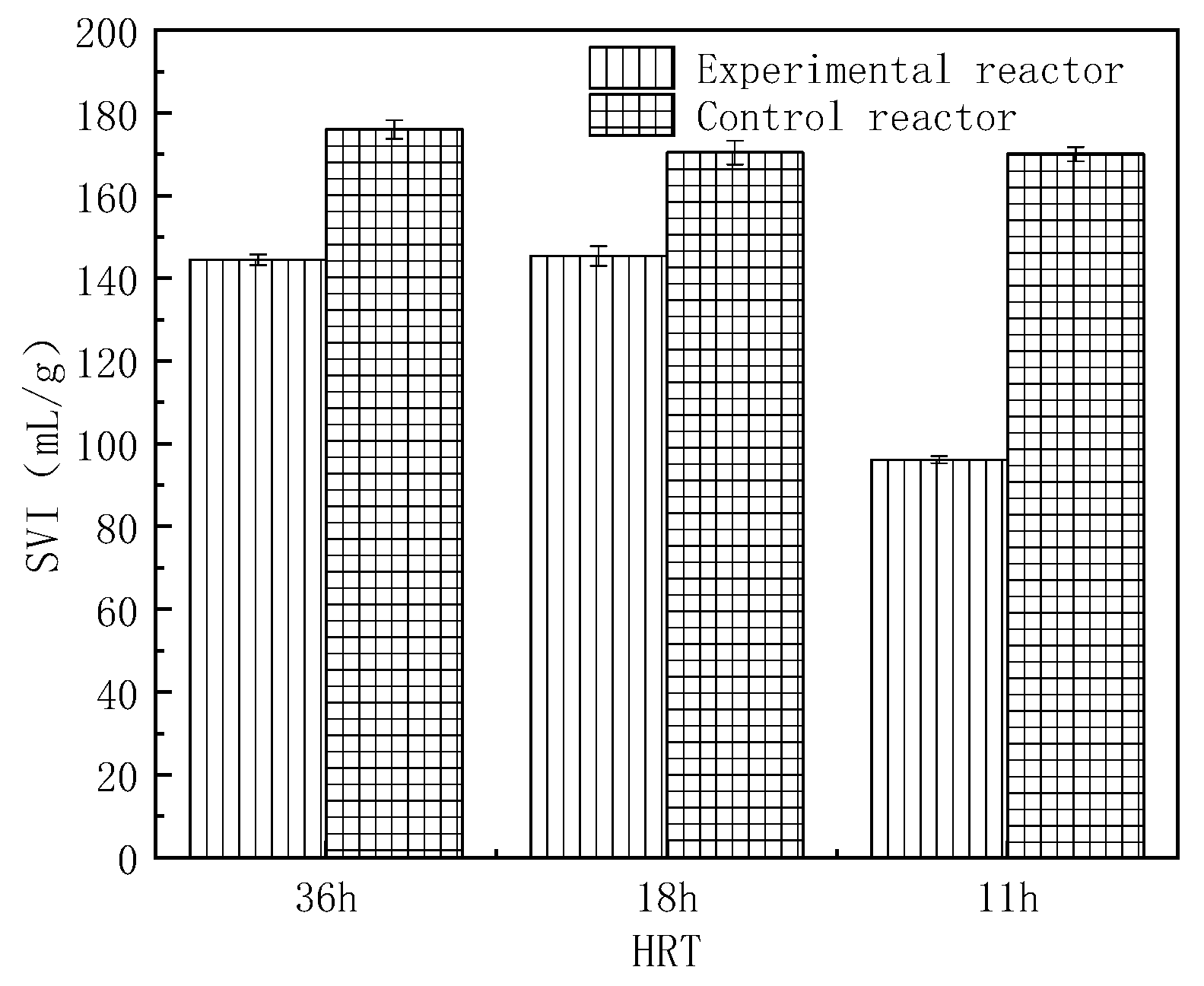
2.2. Sludge Volume Index (SVI) Changes of Biofloc
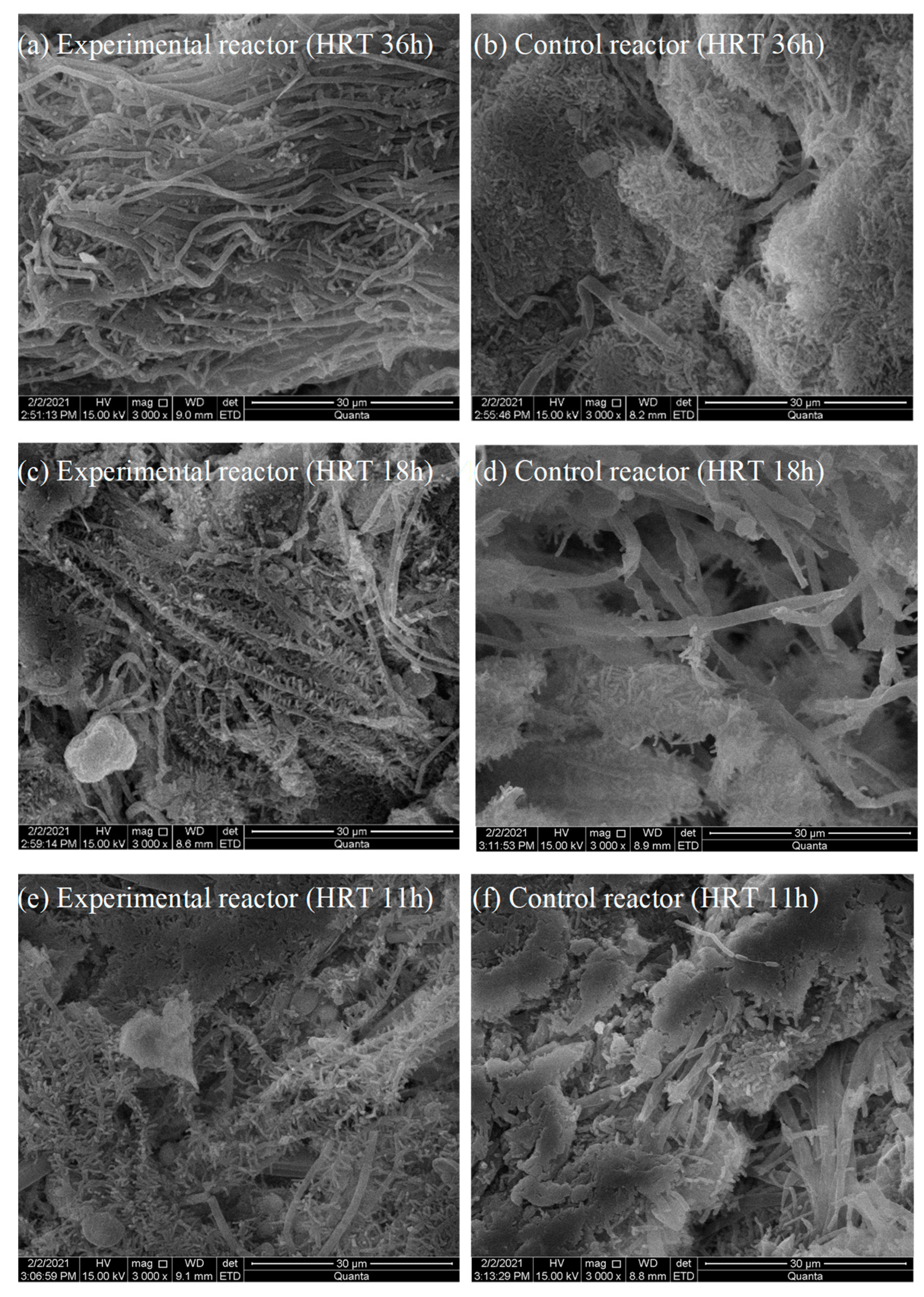
2.3. Changes in the Appearance of Biofloc
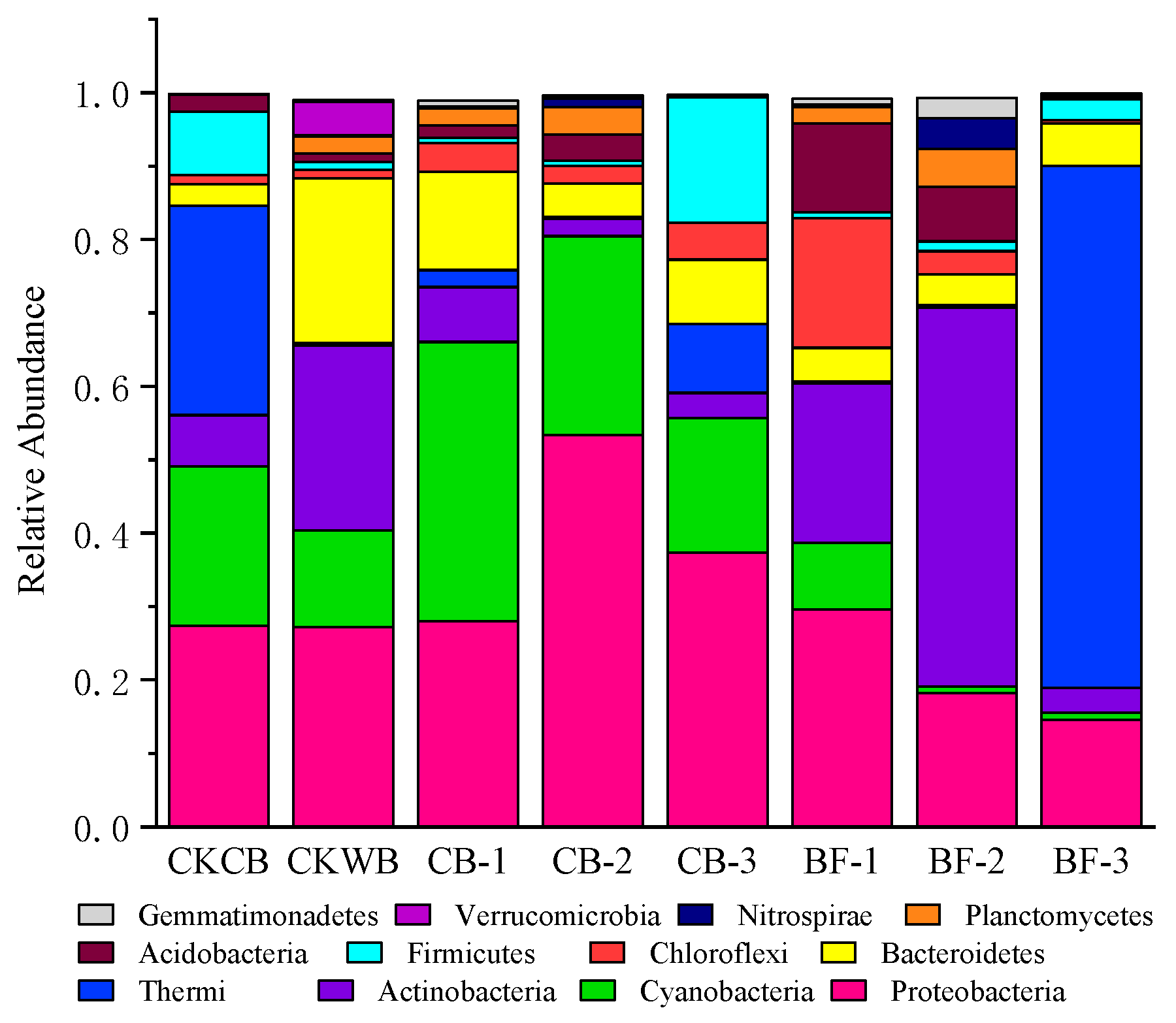
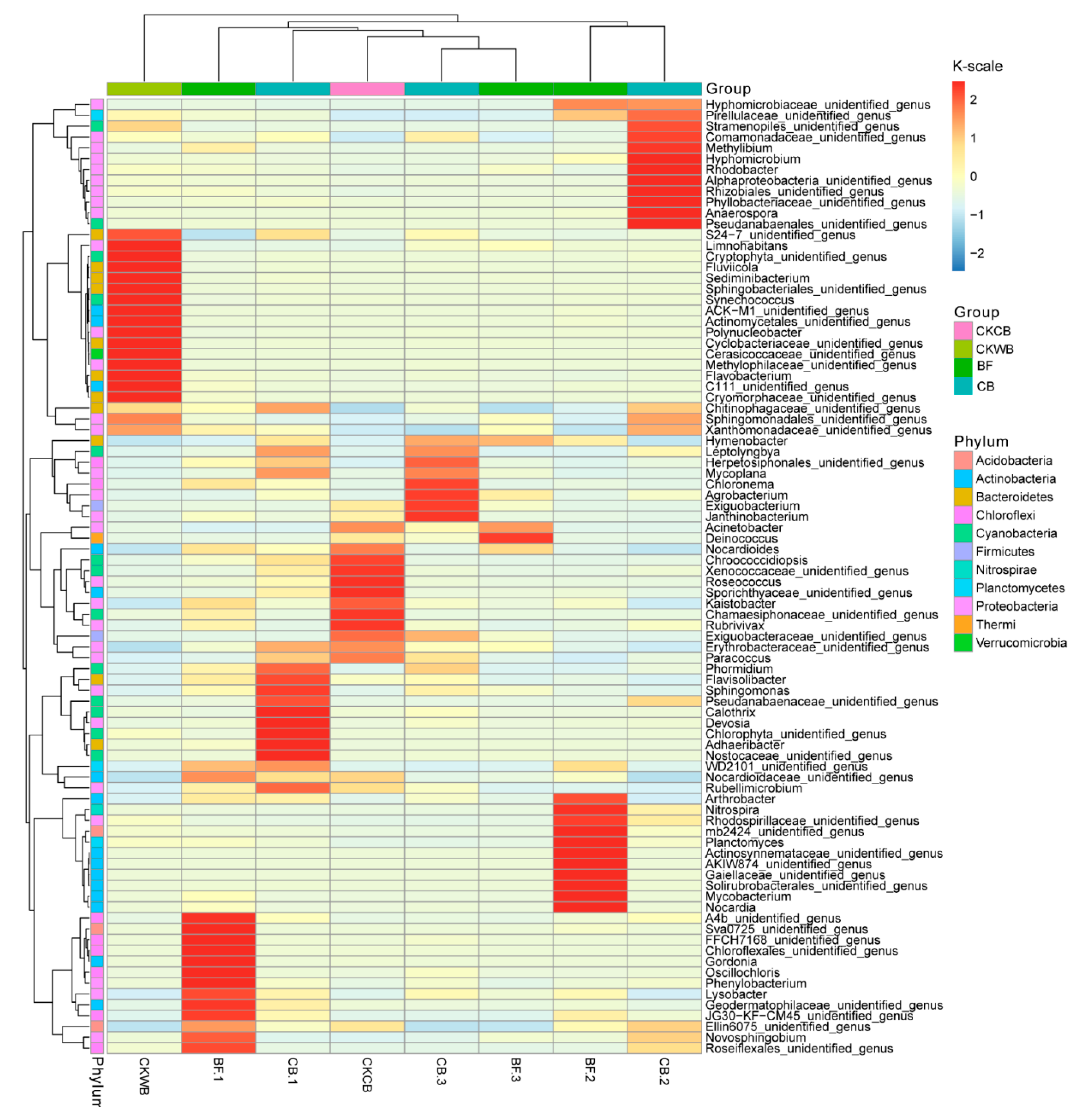
2.4. Changes in the Microbial Community Structure in Biofloc
2.4.1. Changes in Prokaryote in Biofloc
- (1)
- The diversity index of prokaryote
- (2)
- The relative abundance of prokaryotes at the phylum level
- (3)
- The relative abundance of prokaryotes at the genus level
2.4.2. Changes in Eukaryotes in Biofloc
- (1)
- The diversity index of eukaryotes
- (2)
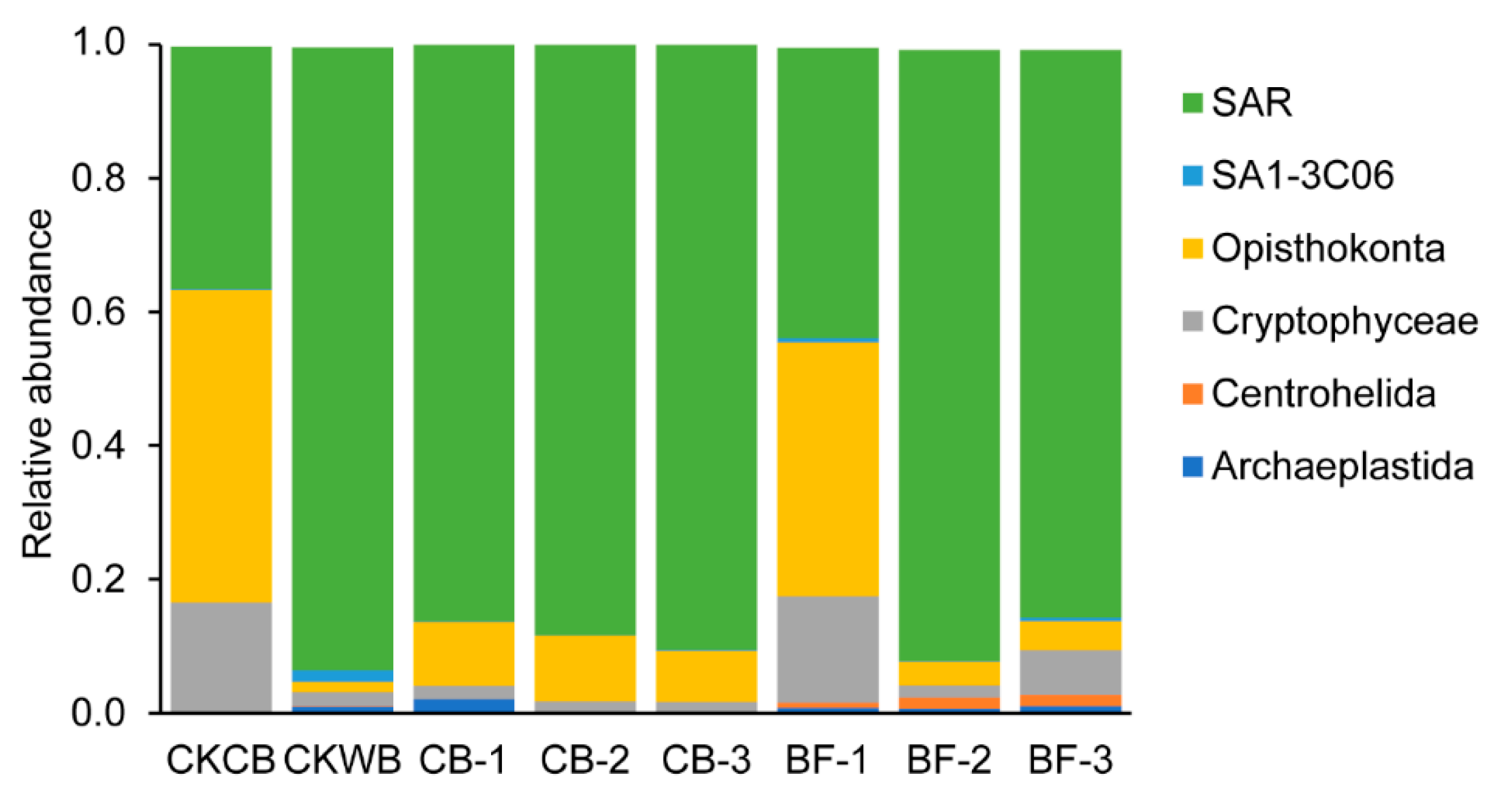
- The relative abundance of eukaryotes at the phylum level
- (3)
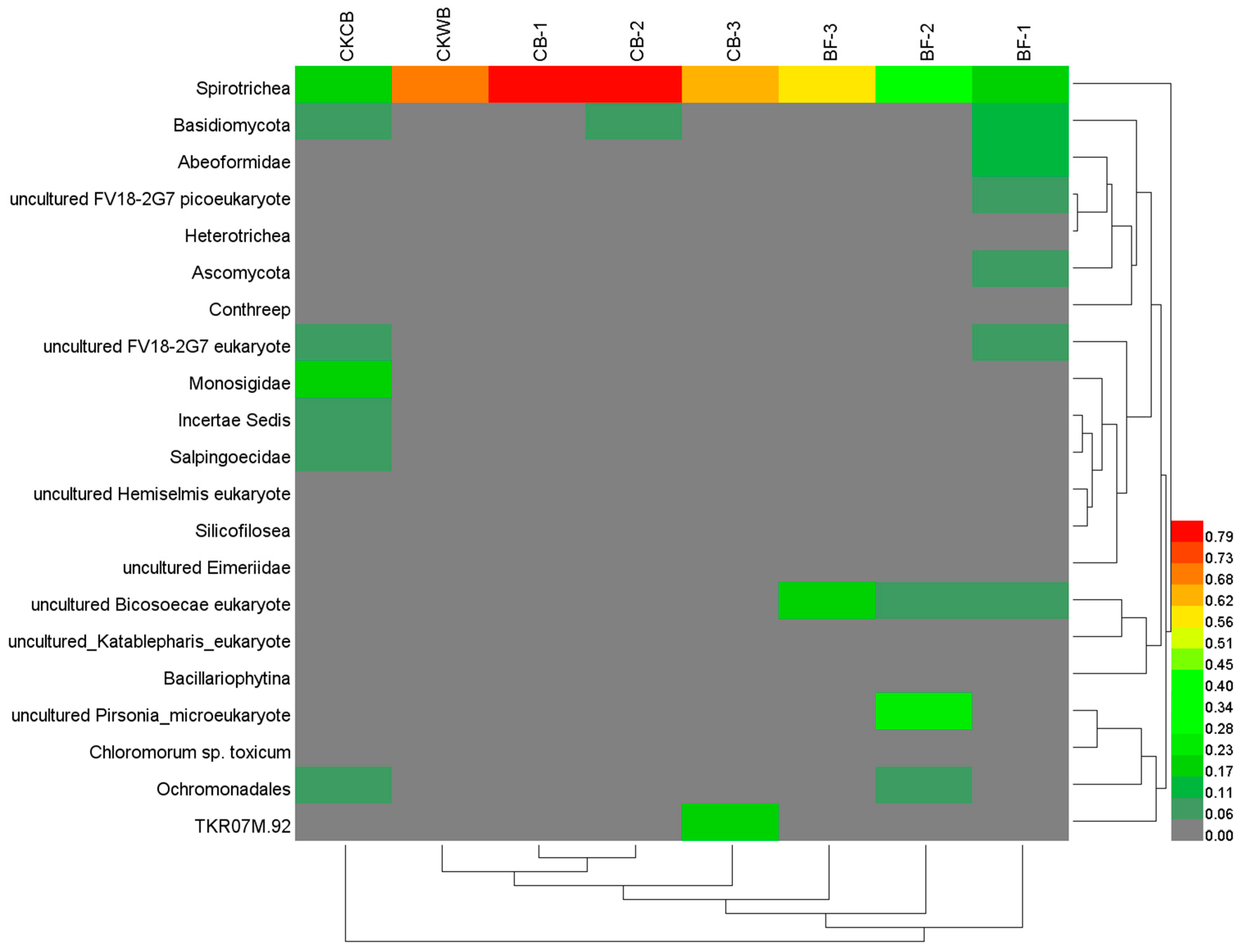
- The relative abundance of eukaryotes at the genus level
3. Materials and Methods
3.1. Experimental Materials
3.2. Experimental Methods
3.2.1. Culture and Collection of C058
3.2.2. Operation of the Reactors
3.3. Analysis Method
3.3.1. Extraction and Determination of EPS
3.3.2. Determination of SVI
3.3.3. SEM Analysis
3.3.4. Analysis of Microbial Community Structure
- (1)
- The 16S rRNA sequence analysis
- (2)
- The 18S rRNA sequence analysis
3.4. Data Processing
4. Conclusions
- (1)
- The EPS contents in MBF were 51.20 mg/g (HRT 36 h), 55.89 mg/g (HRT 18 h), and 33.84 mg/g (HRT 11 h), respectively, higher than the EPS content of OBF under the corresponding HRTs. PN content or PS content also followed the same trend.
- (2)
- The sedimentation performance of MBF was better than that of OBF, attributed to higher EPS PN contents.
- (3)
- MBF was constructed by fungi and bacteria together, and C058 was the main component, promoting the synthesis of EPS.
- (4)
- Compared with OBF, MBF bioaugmented by Cordyceps strain C058 had higher diversity and abundance of microorganisms, realizing a more stable operation of the experimental reactor. More importantly, C058 promoted the growth of some functional bacteria, including Janthinobacterium belonging to Proteobacteria, Phormidium and Leptolyngbya belonging to Cyanobacteria, and Hymenobacter belonging to Bacteroides, which participate in nitrogen fixation and PN and PS production, promoting the EPS synthesis. In addition, C058 also promoted the growth of Spirotrichea belonging to Ciliophora, which benefited floc formation and enhanced the water purification.
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Z.C.; Gao, M.C.; Wei, J.F.; Yang, Y.S.; Zhang, J.; Yu, S.P.; Wang, Y.J. Effect of salinity change on extracellular polymer of anaerobic sludge. Acta Sci. Circumst. 2016, 36, 3273–3281. [Google Scholar]
- Sheng, G.P.; Yu, H.Q.; Li, X.Y. Extracellular polymeric substances (EPS) of microbial aggregates in biological wastewater treatment systems: A review. Biotechnol. Adv. 2010, 28, 882–894. [Google Scholar] [CrossRef]
- More, T.; Yadav, J.; Yan, S.; Tyagi, R.; Surampalli, R. Extracellular polymeric substances of bacteria and their potential environmental applications. J. Environ. Manag. 2014, 144, 1–25. [Google Scholar] [CrossRef]
- Dignac, M.; Urbain, V.; Rybacki, D.; Bruchet, A.; Snidaro, D.; Scribe, P. Chemical description of extracellular polymers: Implication on activated sludge floc structure. Water Sci. Technol. 1998, 38, 45–53. [Google Scholar] [CrossRef]
- Jagaba, A.H.; Kutty, S.; Noor, A.; Haruna Birniwa, A.; Affam, A.; Lawal, I.M.; Usman Kankia, M.; Usman Kilaco, A. A systematic literature review of biocarriers: Central elements for biofilm formation, organic and nutrients removal in sequencing batch biofilm reactor. J. Water Process Eng. 2021, 42, 102178. [Google Scholar] [CrossRef]
- Wang, Z.C.; Gao, M.; Wang, S.; Xin, Y.J.; Ma, D.; She, Z.L.; Wang, Z.; Chang, Q.B.; Ren, Y. Effect of hexavalent chromium on extracellular polymeric substances of granular sludge from an aerobic granular sequencing batch reactor. Chem. Eng. J. 2014, 251, 165–174. [Google Scholar] [CrossRef]
- Basuvaraj, M.; Fein, J.; Liss, S.N. Protein and polysaccharide content of tightly and loosely bound extracellular polymeric substances and the development of a granular activated sludge floc. Water Res. 2015, 82, 104–117. [Google Scholar] [CrossRef]
- Zhu, N.; Liu, L.; Xu, Q.; Chen, G.W.; Wang, G. Resources availability mediated EPS production regulate microbial cluster formation in activated sludge system. Chem. Eng. J. 2015, 279, 129–135. [Google Scholar] [CrossRef]
- Ray, A.J.; Seaborn, G.; Leffler, J.W.; Wilde, S.B.; Lawson, A.; Browdy, C.L. Characterization of microbial communities in minimal-exchange, intensive aquaculture systems and the effects of suspended solids management. Aquaculture 2010, 310, 130–138. [Google Scholar] [CrossRef]
- Zhao, D.H.; Pan, L.Q.; Wang, C. Cleaning effect of biofloc on culture environment and effects of physiological indexes of Penaeus vannamei. Trans. Oceanol. Limnol. 2014, 67–73. [Google Scholar]
- Jagaba, A.H.; Kutty, S.R.M.; Noor, A.; Affam, A.C.; Ghfar, A.A.; Usman, A.K.; Lawal, I.M.; Birniwa, A.H.; Kankia, M.U.; Afolabi, H.K.; et al. Parametric optimization and kinetic modelling for organic matter removal from agro-waste derived paper packaging biorefinery wastewater. Biomass Convers. Biorefin. 2022, 1–18. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, X.; Li, C. Isolation and identification of a bacillus sp. strain and its role in bioflocs for the shrimp culture system. Prog. Fish. Sci. 2016, 37, 111–118. [Google Scholar]
- Gao, G.; Zhu, K.L.; Zhang, Q.Q.; Wang, Z.J.; Huang, J. Simplified fermentation of a functional probiotics and the application in prawn (Litopenaeus vannamei) bio-floc breeding. Prog. Fish. Sci. 2017, 38, 140–147. [Google Scholar]
- Wang, S.H.; Tan, H.X.; Luo, G.Z.; Sun, D.C. Effects of temperature, pH and dissolved oxygen on the construction of biofloc system by Lactobacillus plantarum. Genom. Appl. Biol. 2020, 39, 2628–2637. [Google Scholar]
- Baldrian, P. Interactions of heavy metals with white-rot fungi. Enzyme Microb. Technol. 2003, 32, 78–91. [Google Scholar] [CrossRef]
- Crini, G. Non-conventional low-cost adsorbents for dye removal: A review. Bioresour. Technol. 2006, 97, 1061–1085. [Google Scholar] [CrossRef]
- Zhao, L.H.; Jin, R.F.; Sun, H.J.; Gao, Y.J. Study on treatment of cotton pulp black liquor by white rot fungi and activated sludge. Ecol. Environ. 2009, 18, 28–31. [Google Scholar]
- Wang, L.; Chen, G.Q.; Zeng, G.M.; Zhang, W.Q.; Chen, Y. Research progress on extracellular polymers of fungi and their interaction with heavy Metals. Environ. Pollut. Control 2010, 32, 74–80. [Google Scholar]
- Li, Y.; Yang, Y.C.; Wang, B.E.; Lin, C.; Tao, X.Q.; Du, J.J. Optimization and effect of purification of aquaculture wastewater by Cordyceps sinensis. Adv. Environ. Prot. 2020, 10, 10. [Google Scholar]
- Yang, Y.C. Purification Effect and Mechanism of Cordyceps Sinensis on Aquaculture Wastewater; Zhongkai University of Agriculture and Engineering: Guangzhou, China, 2020. [Google Scholar]
- Zhang, R.Y. Occurrence Regularity of Mycelium Pellets in Activated Sludge System and Its Bioaugmentation Application; Hebei University of Engineering: Handan, China, 2020. [Google Scholar]
- Li, Y.; Chen, Y.C.; Li, D.L.; Zhang, Y.X.; Zhang, W.M. Study on antibacterial and antitumor activity of several fermented extracts of Cordyceps sinensis. Sci. Technol. Food Ind. 2010, 31, 88–90. [Google Scholar]
- Zhou, X.; Huang, M.Q.; Qiu, R.; Li, Z.Q.; Guo, S.H.; Cui, Y.H. Research progress on composition, production and application of Cordyceps sinensis. Priv. Sci. Technol. 2018, 98–99. [Google Scholar]
- Morgan, J.W.; Forster, C.F.; Evison, L. A comparative study of the nature of biopolymers extracted from anaerobic and activated sludges. Water Res. 1990, 24, 743–750. [Google Scholar] [CrossRef]
- Ge, L.Y.; Wang, H.W.; Ma, L.M.; Deng, H.H.; Liu, Y. Study on influencing factors of extracellular polymers in aerobic activated sludge. Environ. Sci. Technol. 2007, 02, 8–9. [Google Scholar]
- Gu, Y.N. Study on the Efficiency and Stability of Aerobic Granular Sludge in the Treatment of Domestic Wastewater; Harbin Institute of Technology: Harbin, China, 2020. [Google Scholar]
- Zhang, X.; Zhang, Q.S.; Xu, Y.; Liu, X. Study on sedimentation and dewatering performance of activated sludge from a sewage treatment plant. Environ. Prot. Sci. 2011, 37, 20–22. [Google Scholar]
- Zhang, L.H.; Li, J.; Guo, J.B.; Jia, Y.P.; Zhang, H.F. Effect of EPS on flocculation settling performance and surface properties of activated sludge. J. Chem. Ind. Eng. 2012, 63, 1865–1871. [Google Scholar]
- Li, H.; Qu, J.; Li, T.; Li, j.; Lin, Q.; Li, X. Pika population density is associated with the composition and diversity of gut microbiota. Front. Microbiol. 2016, 7, 758. [Google Scholar] [CrossRef]
- Tsuneda, S.; Jung, J.; Hayashi, H.; Aikawa, H.; Hirata, A.; Sasaki, H. Influence of extracellular polymers on electrokinetic properties of heterotrophic bacterial cells examined by soft particle electrophoresis theory. Colloids Surf. B Biointerfaces 2003, 29, 181–188. [Google Scholar] [CrossRef]
- Satoshi, T.; Hirotoshi, A.; Hiroshi, H.; Atsushi, Y.; Akira, H. Extracellular polymeric substances responsible for bacterial adhesion onto solid surface. FEMS Microbiol. Lett. 2010, 223, 287–292. [Google Scholar]
- Cardona, E.; Gueguen, Y.; Magré, K.; Lorgeoux, B.; Piquemal, D.; Pierrat, F.; Noguier, F.; Saulnier, D. Bacterial community characterization of water and intestine of the shrimp Litopenaeus stylirostris in a biofloc system. BMC Microbiol. 2016, 16, 157. [Google Scholar] [CrossRef]
- Addo, F.G.; Zhang, S.; Manirakiza, B.; Ohore, E.O.; Yuan, S.D. The impacts of straw substrate on biofloc formation, bacterial community and nutrient removal in shrimp ponds. Bioresour. Technol. 2021, 326, 124727. [Google Scholar] [CrossRef]
- Yang, C.; Wei, Z.; Liu, R.; LI, Q.; Li, B.; Wang, S.; Song, C.; Qiao, C.; Aulchandani, A. Phylogenetic diversity and metabolic potential of activated sludge microbial communities in full-scale wastewater treatment plants. Environ. Sci. Technol. 2011, 45, 7408–7415. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.S.; Parameswaran, P.; Rittmann, B.E. Effects of solids retention time on methanogenesis in anaerobic digestion of thickened mixed sludge. Bioresour. Technol. 2011, 102, 10266–10272. [Google Scholar] [CrossRef]
- Yang, M.; Lu, D.W.; Qin, B.D.; Liu, Q.L.; Zhao, Y.M.; Liu, H.L.; Ma, J. Highly efficient nitrogen removal of a coldness-resistant and low nutrient needed bacterium, Janthinobacterium sp M-11. Bioresour. Technol. 2018, 256, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Bei, Q.C.; Wang, X.J.; Lan, P.; Liu, G.; Lin, X.W.; Liu, Q.; Lin, Z.B.; Liu, B.J.; Zhang, Y.H.; et al. Impacts of Mo application on biological nitrogen fixation and diazotrophic communities in a flooded rice-soil system. Sci. Total Environ. 2019, 649, 686–694. [Google Scholar] [CrossRef] [PubMed]
- Bar-Or, Y.; Shilo, M. Characterization of macromolecular flocculants produced by Phormidium sp. strain J-1 and by Anabaenopsis circularis PCC 6720. Appl. Environ. Microbiol. 1987, 53, 2226–2230. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Choi, W.; Jeon, S.M.; Kim, T.; Park, A.; Kim, J.; Heo, S.J.; Oh, C.; Shim, W.B.; Kang, D.H. Isolation and characterization of Leptolyngbya sp. KIOST-1, a basophilic and euryhaline filamentous cyanobacterium from an open paddle-wheel raceway Arthrospira culture pond in Korea. J. Appl. Microbiol. 2015, 119, 1597–1612. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Kang, D.H. Draft genome sequence of Leptolyngbya sp. KIOST-1, a filamentous cyanobacterium with biotechnological potential for alimentary purposes. Genome Announc. 2016, 4, e00984-16. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.P.; Tang, K.; Li, H.; Cheng, Y.L.; Yang, S.S.; Guo, H.L.; Guo, H.Q.; Yun, X.Y.; Feng, F.Y. Isolation and Identification of thin layer strain L28 and its promoting effect on the growth of Potato seedlings. Microbiol. China 2020, 47, 4050–4058. [Google Scholar]
- Tenney, M.W.; Stumm, W. Chemical flocculation of microorganisms in biological waste treatment. J. Water Pollut. Control Fed. 1965, 37, 1370–1388. [Google Scholar]
- Chen, Y.H. Composition and Function Analysis of Extracellular Polymers during Activated Sludge Aggregation; Chongqing University: Chongqing, China, 2019. [Google Scholar]
- Andersen, P. The quantitative importance of the microbial loop in the marine pelagic: A case study from the North Bering/Chukchi seas. Arch. Hydrobiol. Beih. 1988, 31, 243–251. [Google Scholar]
- Nielsen, T.G.; Hansen, B. Plankton community structure and carbon cycling on the western coast of Greenland during and after the sedimentation of a diatom bloom. Mar. Ecol. Prog. Ser. 1995, 125, 239–257. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weerman, E.J.; Van Der Geest, H.G.; Van Der Meulen, M.D.; Manders, E.M.M.; Koppel, J.V.D.; Herman, P.M.J.; Admiraal, A.W. Ciliates as engineers of phototrophic biofilms. Freshw. Biol. 2011, 56, 1358–1369. [Google Scholar] [CrossRef]
- Weber, S.D.; Ludwig, W.; Schleifer, K.H.; Fried, J. Microbial composition and structure of aerobic granular sewage biofilms. Appl. Environ. Microbiol. 2007, 73, 6233–6240. [Google Scholar] [CrossRef] [Green Version]
- Schryver, P.D.; Verstraete, W. Nitrogen removal from aquaculture pond water by heterotrophic nitrogen assimilation in lab-scale sequencing batch reactors. Bioresour. Technol. 2009, 99, 1162–1167. [Google Scholar] [CrossRef]
- Zhang, X.; Bishop, P.L.; Kinkle, B.K. Comparison of extraction methods for quantifying extracellular polymers in biofilms. Water Sci. Technol. 1999, 39, 211–218. [Google Scholar] [CrossRef]
- Bo, F.; Palmgren, R.; Keiding, K.; Nielsen, P.H. Extraction of extracellular polymers from activated sludge using a cation exchange resin. Water Res. 1996, 30, 1749–1758. [Google Scholar]
- Eaton, A.D.; Clesceri, L.S.; Greenberg, A.E.; Franson, M.A.H. Standard methods for the examination of water and wastewater. Am. J. Public Health Nations Health 1966, 56, 387–388. [Google Scholar]
- Shen, P.; Chen, X.D. Microbiology Experiment; Higher Education Press: Beijing, China, 2007. [Google Scholar]
- Tamaki, H.; Wright, C.L.; Li, X.; Lin, Q.; Hwang, C.; Wang, S.; Thimmapuram, J.; Kamagata, Y.; Liu, W.T. Analysis of 16S rRNA amplicon sequencing options on the Roche/454 Next-Generation Titanium Sequencing Platform. PLoS ONE 2011, 6, e25263. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Peng, Y.Z.; Wang, R.D. The role of extracellular polymers substance in biological treatment of wastewater. J. Nat. Sci. Heilongjiang Univ. 2016, 33, 515–520. [Google Scholar]
- Ni, J.J.; Li, X.J.; He, Z.L.; Xu, M.Y. A novel method to determine the minimum number of sequences required for reliable microbial community analysis. J. Microbiol. Methods 2017, 139, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Lejzerowicz, F.; Esling, P.; Pillet, L.; Wilding, T.A.; Black, K.D.; Pawlowski, J. High-throughput sequencing and morphology perform equally well for benthic monitoring of marine ecosystems. Sci. Rep. 2015, 5, 13932. [Google Scholar] [CrossRef] [PubMed] [Green Version]









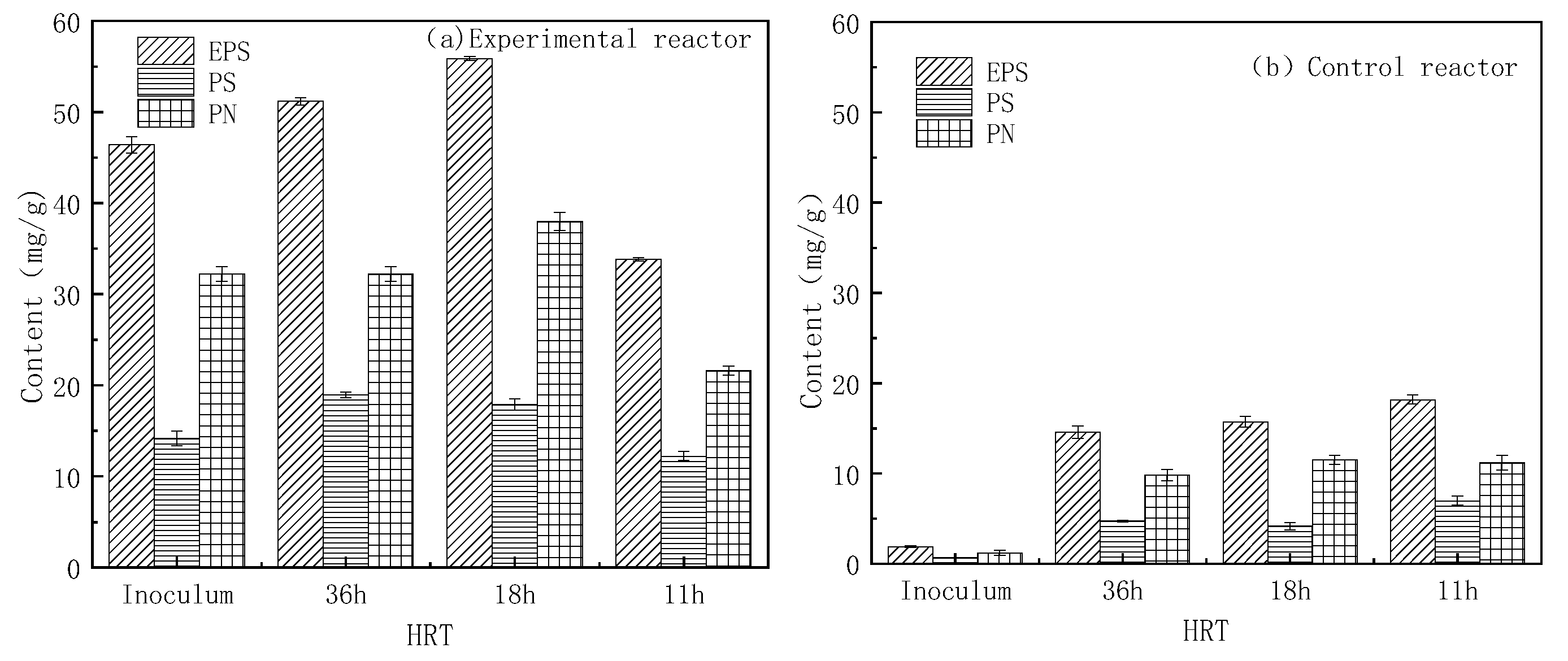{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chao1 | Shannon | Simpson | |
|---|---|---|---|
| CKCB | 1701.59 | 6.16 | 0.969 |
| CB-1 | 2868.92 | 8.42 | 0.988 |
| CB-2 | 2720.18 | 8.27 | 0.987 |
| CB-3 | 1906.67 | 6.16 | 0.942 |
| CKWB | 2477.06 | 7.80 | 0.988 |
| BF-1 | 2242.12 | 7.82 | 0.981 |
| BF-2 | 2059.89 | 6.67 | 0.915 |
| BF-3 | 1489.73 | 3.97 | 0.745 |
| Chao1 | Shannon | Simpson | |
|---|---|---|---|
| CKCB | 290.25 | 4.43 | 0.92 |
| CB-1 | 387.69 | 2.94 | 0.63 |
| CB-2 | 259.12 | 2.21 | 0.52 |
| CB-3 | 393.45 | 3.17 | 0.75 |
| CKWB | 492.11 | 4.06 | 0.87 |
| BF-1 | 481.27 | 4.93 | 0.93 |
| BF-2 | 372.07 | 4.16 | 0.88 |
| BF-3 | 353.57 | 3.75 | 0.86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Luo, W.; Liu, W.; Yang, Y.; Lei, Z.; Tao, X.; Wang, B. C058 and Other Functional Microorganisms Promote the Synthesis of Extracellular Polymer Substances in Mycelium Biofloc. Catalysts 2022, 12, 693. https://doi.org/10.3390/catal12070693
Li Y, Luo W, Liu W, Yang Y, Lei Z, Tao X, Wang B. C058 and Other Functional Microorganisms Promote the Synthesis of Extracellular Polymer Substances in Mycelium Biofloc. Catalysts. 2022; 12(7):693. https://doi.org/10.3390/catal12070693
Chicago/Turabian StyleLi, Yiyong, Wanyi Luo, Wen Liu, Yongcong Yang, Zexiang Lei, Xueqin Tao, and Baoe Wang. 2022. "C058 and Other Functional Microorganisms Promote the Synthesis of Extracellular Polymer Substances in Mycelium Biofloc" Catalysts 12, no. 7: 693. https://doi.org/10.3390/catal12070693
APA StyleLi, Y., Luo, W., Liu, W., Yang, Y., Lei, Z., Tao, X., & Wang, B. (2022). C058 and Other Functional Microorganisms Promote the Synthesis of Extracellular Polymer Substances in Mycelium Biofloc. Catalysts, 12(7), 693. https://doi.org/10.3390/catal12070693