
Continuous Packed Bed Reactor with Immobilized ?-Galactosidase for Production of Galactooligosaccharides (GOS)



Abstract
:
1. Introduction
2. Results and Discussion
2.1. Immobilization of β-Galactosidase from B. circulans on Glyoxal Agarose Resin
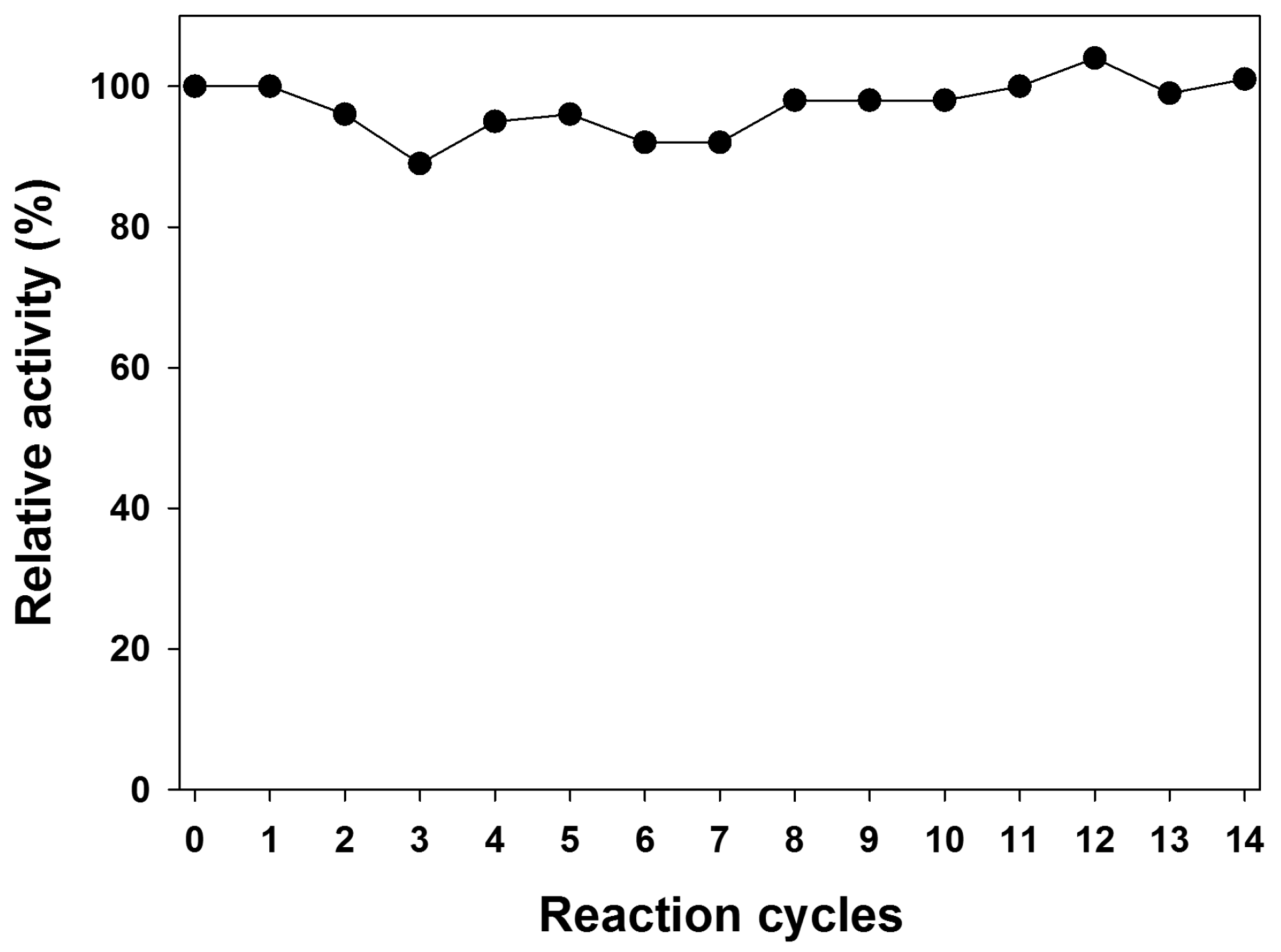
2.2. Operational Stability in Batch Reactor
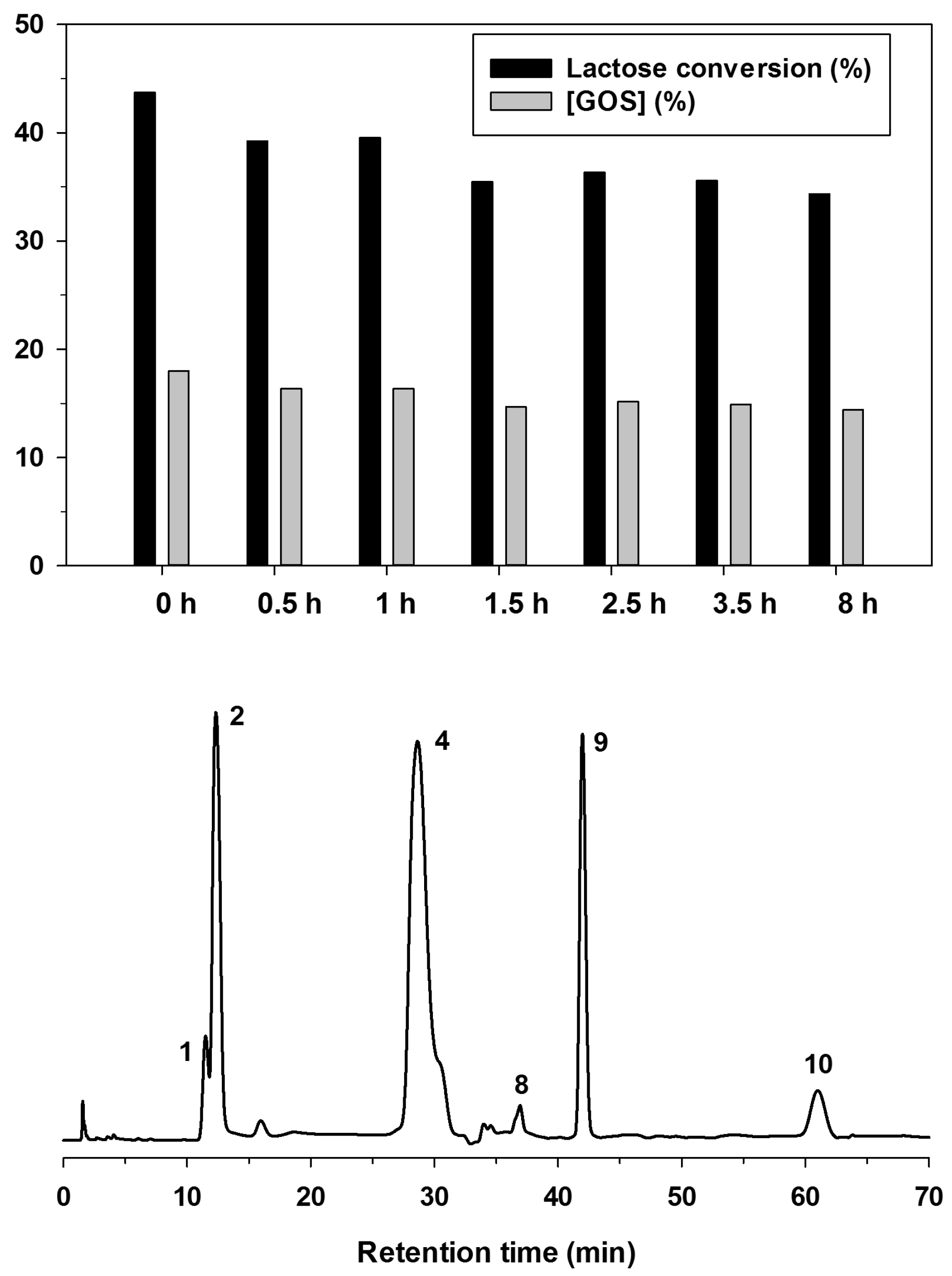
2.3. Effect of Lactose Conversion on GOS Yield with Immobilized B. circulans β-Galactosidase
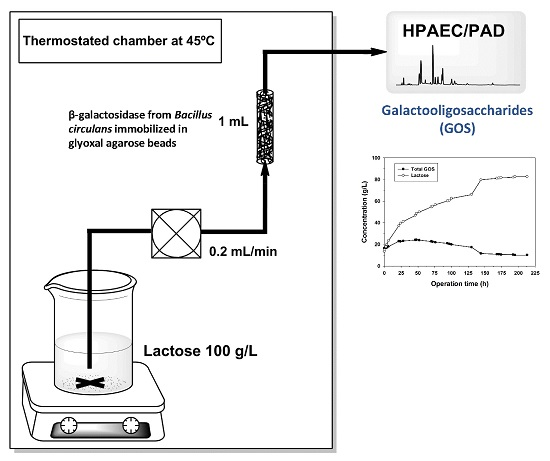
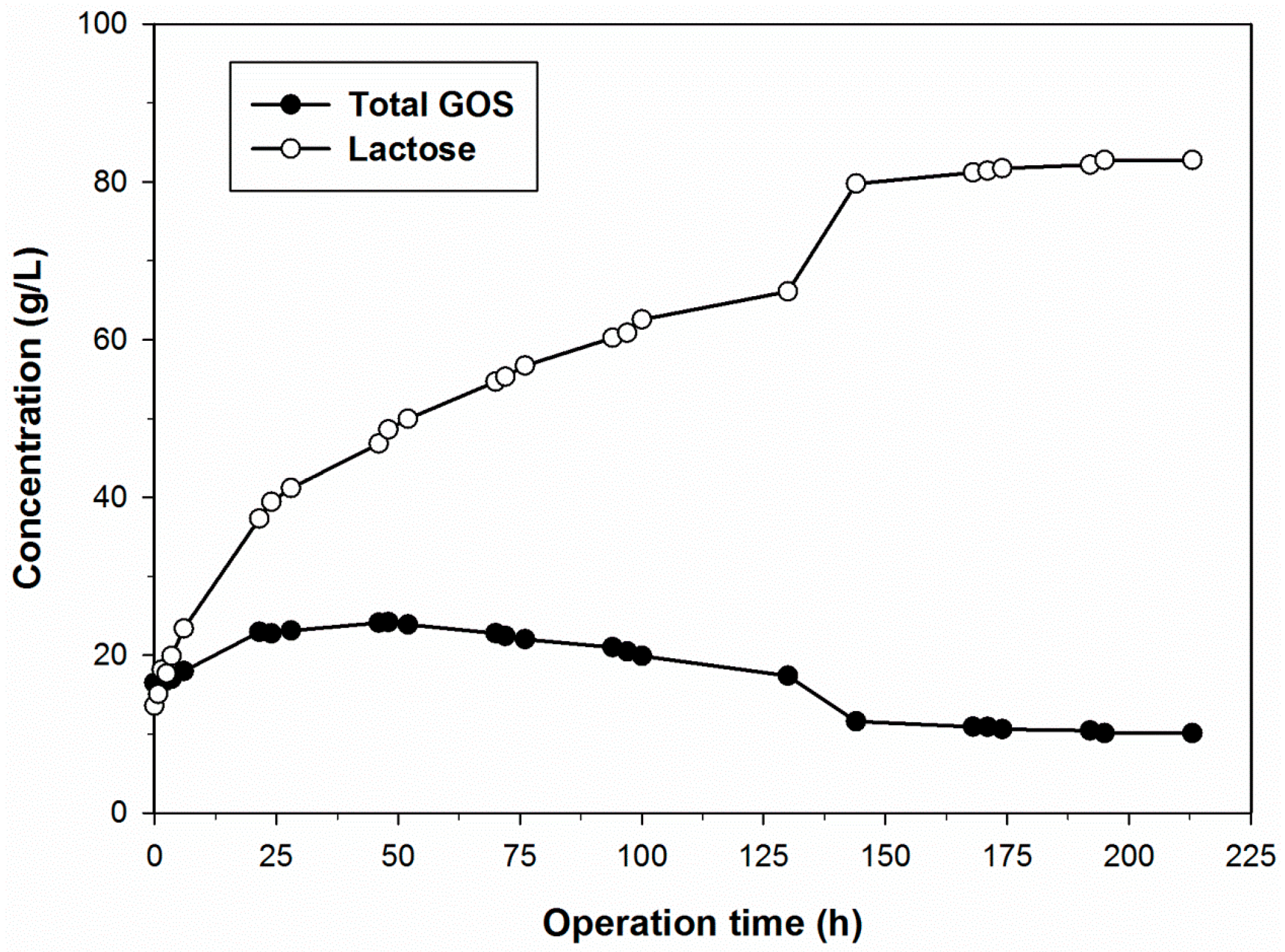
2.4. GOS Synthesis in PBR with Immobilized B. circulans β-Galactosidase
2.5. Specificity of GOS Production in PBR
2.6. Continuous GOS Production using Milk as Feed Solution
3. Materials and Methods
3.1. Materials
3.2. Activity Assay
3.3. Covalent Immobilization
3.4. Operational Stability in Batch Reactor
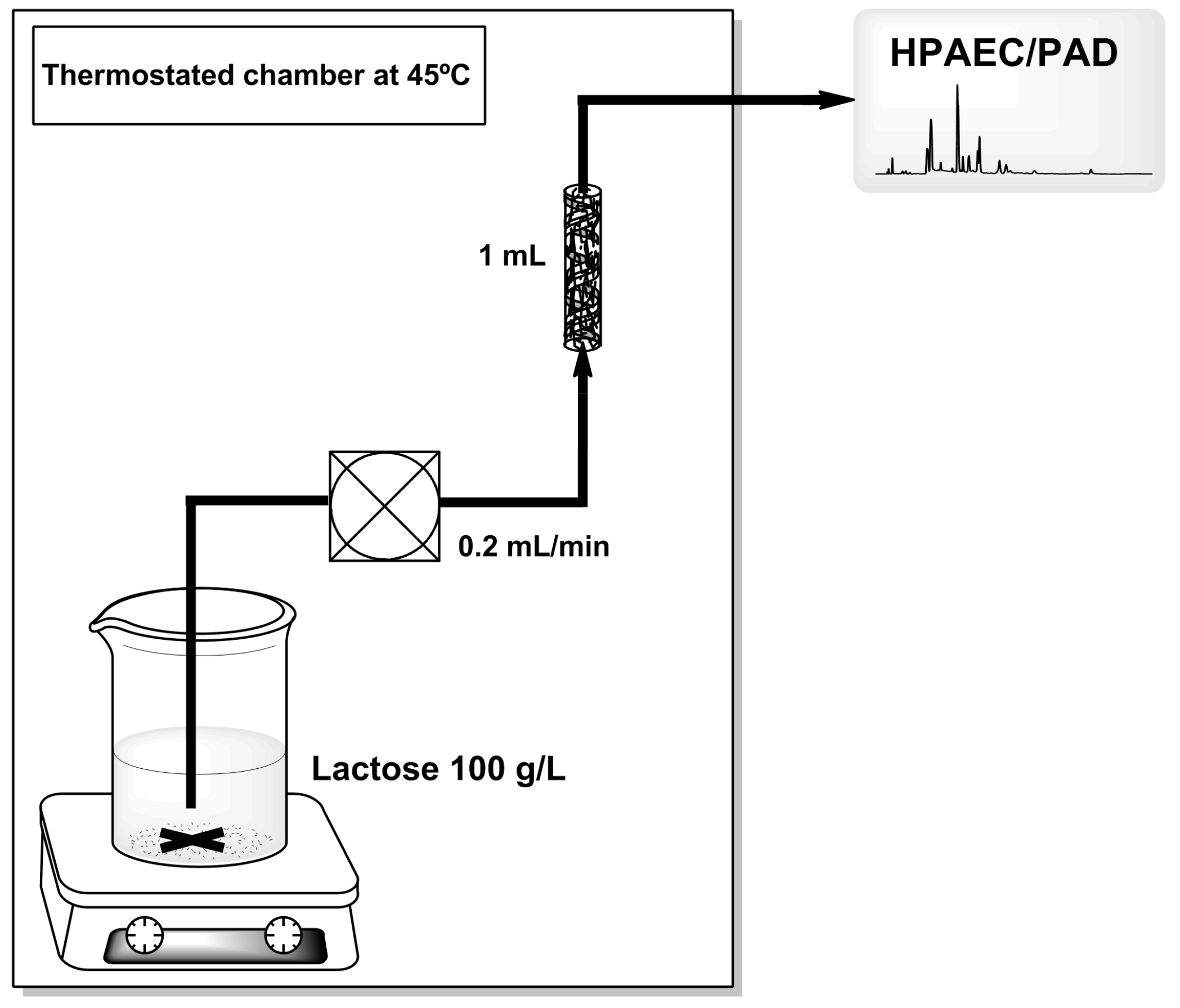
3.5. GOS Production in Continuous Packed-Bed Reactor
3.6. High-Performance Anion-Exchange Chromatography with Pulsed Amperometric Detection (HPAEC-PAD)
3.7. Scanning Electron Microscopy
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Park, A.R.; Oh, D.K. Galacto-oligosaccharide production using microbial β-galactosidase: Current state and perspectives. Appl. Microbiol. Biotechnol. 2010, 85, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Plou, F.J.; Polaina, J.; Sanz-Aparicio, J.; Fernandez-Lobato, M. β-Galactosidases for lactose hydrolysis and galactooligosaccharide synthesis. In Microbial Enzyme Technology in Food Applications; Ray, R.C., Rosell, C.M., Eds.; CRC Press: Boca Raton, FL, USA, 2016; pp. 123–146. [Google Scholar]
- Iqbal, S.; Nguyen, T.H.; Nguyen, T.T.; Maischberger, T.; Haltrich, D. β-Galactosidase from Lactobacillus plantarum WCFS1: Biochemical characterization and formation of prebiotic galacto-oligosaccharides. Carbohydr. Res. 2010, 345, 1408–1416. [Google Scholar] [CrossRef] [PubMed]
- Torres, D.P.; Goncalves, M.; Teixeira, J.A.; Rodrigues, L.R. Galacto-oligosaccharides: Production, properties, applications, and significance as prebiotics. Compr. Rev. Food Sci. Food Saf. 2010, 9, 438–454. [Google Scholar] [CrossRef] [Green Version]
- Lamsal, B.P. Production, health aspects and potential food uses of dairy prebiotic galactooligosaccharides. J. Sci. Food Agric. 2012, 92, 2020–2028. [Google Scholar] [CrossRef] [PubMed]
- Konar, E.; Sarkar, S.; Singhal, R.S. Galactooligosaccharides: Chemistry, production, properties, market status and applications—A review. Trends Carbohydr. Res. 2011, 3, 1–16. [Google Scholar]
- Shadid, R.; Haarman, M.; Knol, J.; Theis, W.; Beermann, C.; Rjosk-Dendorfer, D.; Schendel, D.J.; Koletzko, B.V.; Krauss-Etschmann, S. Effects of galactooligosaccharide and long-chain fructooligosaccharide supplementation during pregnancy on maternal and neonatal microbiota and immunity—A randomized, double-blind, placebo-controlled study. Am. J. Clin. Nutr. 2007, 86, 1426–1437. [Google Scholar] [PubMed]
- Torres-Salas, P.; Del Monte-Martinez, A.; Cutiño-Avila, B.; Rodriguez-Colinas, B.; Alcalde, M.; Ballesteros, A.O.; Plou, F.J. Immobilized biocatalysts: Novel approaches and tools for binding enzymes to supports. Adv. Mater. 2011, 23, 5275–5282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheldon, R.A. Enzyme immobilization: The quest for optimum performance. Adv. Synth. Catal. 2007, 349, 1289–1307. [Google Scholar] [CrossRef]
- Boller, T.; Meier, C.; Menzler, S. Eupergit oxirane acrylic beads: How to make enzymes fit for biocatalysis. Org. Process Res. Dev. 2002, 6, 509–519. [Google Scholar] [CrossRef]
- Marin-Navarro, J.; Talens-Perales, D.; Oude-Vrielink, A.; Cañada, F.J.; Polaina, J. Immobilization of thermostable β-galactosidase on epoxy support and its use for lactose hydrolysis and galactooligosaccharides biosynthesis. World J. Microbiol. Biotechnol. 2014, 30, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Shankar, S.K.; Praveen Kumar, S.K.; Mulimani, V.H. Calcium alginate entrapped preparation of α-galactosidase: Its stability and application in hydrolysis of soymilk galactooligosaccharides. J. Ind. Microbiol. Biotechnol. 2011, 38, 1399–1405. [Google Scholar] [CrossRef] [PubMed]
- Genari, A.N.; Passos, F.V.; Passos, F.M.L. Configuration of a bioreactor for milk lactose hydrolysis. J. Dairy Sci. 2003, 86, 2783–2789. [Google Scholar] [CrossRef]
- Foda, M.I.; Lopez-Leiva, M. Continuous production of oligosaccharides from whey using a membrane reactor. Process Biochem. 2000, 35, 581–587. [Google Scholar] [CrossRef]
- Nakkharat, P.; Haltrich, D. Betα-galactosidase from Talaromyces thermophilus immobilized on to Eupergit C for production of galacto-oligosaccharides during lactose hydrolysis in batch and packed-bed reactor. World J. Microbiol. Biotechnol. 2007, 23, 759–764. [Google Scholar] [CrossRef]
- Warmerdam, A.; Benjamins, E.; de Leeuw, T.F.; Broekhuis, T.A.; Boom, R.M.; Janssen, A.E.M. Galacto-oligosaccharide production with immobilized β-galactosidase in a packed-bed reactor vs. free β-galactosidase in a batch reactor. Food Bioprod. Process. 2014, 92, 383–392. [Google Scholar] [CrossRef]
- Sen, P.; Nath, A.; Bhattacharjee, C.; Chowdhury, R.; Bhattacharya, P. Process engineering studies of free and micro-encapsulated β-galactosidase in batch and packed bed bioreactors for production of galactooligosaccharides. Biochem. Eng. J. 2014, 90, 59–72. [Google Scholar] [CrossRef]
- Rodriguez-Colinas, B.; Fernandez-Arrojo, L.; de Abreu, M.; Urrutia, P.; Fernandez-Lobato, M.; Ballesteros, A.O.; Plou, F.J. On the enzyme specificity for the synthesis of prebiotic galactooligosaccharides. In Advances in Enzyme Biotechnology; Shukla, P., Pletschke, B., Eds.; Springer: New Delhi, India, 2013; pp. 23–39. [Google Scholar]
- Rodriguez-Colinas, B.; Poveda, A.; Jimenez-Barbero, J.; Ballesteros, A.O.; Plou, F.J. Galacto-oligosaccharide synthesis from lactose solution or skim milk using the β-galactosidase from Bacillus circulans. J. Agric. Food Chem. 2012, 60, 6391–6398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mozaffar, Z.; Nakanishi, K.; Matsuno, R. Formation of oligosaccharides during hydrolysis of lactose in milk using β-galactosidase from Bacillus circulans. J. Food Sci. 1985, 50, 1602–1606. [Google Scholar] [CrossRef]
- Rodriguez-Colinas, B.; Fernandez-Arrojo, L.; Ballesteros, A.; Plou, F. Galactooligosaccharides formation during enzymatic hydrolysis of lactose: Towards a prebiotic-enriched milk. Food Chem. 2014, 145, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Mozaffar, Z.; Nakanishi, K.; Matsuno, R. Continuous production of galacto-oligosaccharides from lactose using immobilized betα-galactosidase from Bacillus circulans. Appl. Microbiol. Biotechnol. 1986, 25, 224–228. [Google Scholar]
- Torres, P.; Batista-Viera, F. Immobilization of β-galactosidase from Bacillus circulans onto epoxy-activated acrylic supports. J. Mol. Catal. B Enzym. 2012, 74, 230–235. [Google Scholar] [CrossRef]
- Urrutia, P.; Mateo, C.; Guisan, J.M.; Wilson, L.; Illanes, A. Immobilization of Bacillus circulans β-galactosidase and its application in the synthesis of galacto-oligosaccharides under repeated-batch operation. Biochem. Eng. J. 2013, 77, 41–48. [Google Scholar] [CrossRef]
- Huerta, L.M.; Vera, C.; Guerrero, C.; Wilson, L.; Illanes, A. Synthesis of galacto-oligosaccharides at very high lactose concentrations with immobilized β-galactosidases from Aspergillus oryzae. Process Biochem. 2011, 46, 245–252. [Google Scholar] [CrossRef]
- Bolivar, J.M.; Wilson, L.; Ferrarotti, S.A.; Fernandez-Lafuente, R.; Guisan, J.M.; Mateo, C. Stabilization of a formate dehydrogenase by covalent immobilization on highly activated glyoxyl-agarose supports. Biomacromolecules 2006, 7, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Santos-Moriano, P.; Woodley, J.M.; Plou, F.J. Continuous production of chitooligosaccharides by an immobilized enzyme in a dual-reactor system. J. Mol. Catal. B Enzym. 2016, 133, 211–217. [Google Scholar] [CrossRef]
- Santos-Moriano, P.; Monsalve-Ledesma, L.; Ortega-Munoz, M.; Fernandez-Arrojo, L.; Ballesteros, A.O.; Santoyo-Gonzalez, F.; Plou, F.J. Vinyl sulfone-activated silica for efficient covalent immobilization of alkaline unstable enzymes: Application to levansucrase for fructooligosaccharide synthesis. RSC Adv. 2016, 6, 64175–64181. [Google Scholar] [CrossRef]
- Worsfold, P.J. Classification and chemical characteristics of immobilized enzymes—Technical report. Pure Appl. Chem. 1995, 67, 597–600. [Google Scholar] [CrossRef]
- Fernandez-Arrojo, L.; Rodriguez-Colinas, B.; Gutierrez-Alonso, P.; Fernandez-Lobato, M.; Alcalde, M.; Ballesteros, A.O.; Plou, F.J. Dried alginate-entrapped enzymes (DALGEEs) and their application to the production of fructooligosaccharides. Process Biochem. 2013, 48, 677–682. [Google Scholar] [CrossRef]
- Fernandez-Arrojo, L.; Santos-Moriano, P.; Rodriguez-Colinas, B.; Ballesteros, A.O.; Plou, F.J. Micro-scale procedure for enzyme immobilization screening and operational stability assays. Biotechnol. Lett. 2015, 37, 1593–1600. [Google Scholar] [CrossRef] [PubMed]
- Plou, F.J.; Alcalde, M.; Ghazi, I.; Fernandez-Arrojo, L.; Ballesteros, A. Efficient fructooligosaccharide synthesis with a fructosyltransferase from Aspergillus aculeatus. In Modern Biocatalysis: Stereoselective and Enviromentally Friendly Reactions; Fessner, W.D., Anthonsen, T., Eds.; Wiley-VCH: Weinheim, Germany, 2009; pp. 153–171. [Google Scholar]
- Linde, D.; Rodriguez-Colinas, B.; Estevez, M.; Poveda, A.; Plou, F.J.; Fernandez-Lobato, M. Analysis of neofructooligosaccharides production mediated by the extracellular b-fructofuranosidase from Xanthophyllomyces dendrorhous. Bioresour. Technol. 2012, 109, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.; Wang, J.; Chen, J.; Yang, N.; Jin, Z.; Xu, X. Production of neo-fructooligosaccharides using free-whole-cell biotransformation by Xanthophyllomyces dendrorhous. Bioresour. Technol. 2010, 101, 7472–7478. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.T.; Plou, F.J.; Alcalde, M.; Ballesteros, A. Immobilization on Eupergit C of cyclodextrin glucosyltransferase (CGTase) and properties of the immobilized biocatalyst. J. Mol. Catal. B Enzym. 2003, 21, 299–308. [Google Scholar] [CrossRef]
- Binay, B.; Alagöz, D.; Yildirim, D.; Çelik, A.; Tükel, S.S. Highly stable and reusable immobilized formate dehydrogenases: Promising biocatalysts for in situ regeneration of NADH. Beilstein J. Org. Chem. 2016, 12, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, R.M.; Costa, J.B.; Serp, P.; Loureiro, J.M.; Faria, J.L.; Silva, C.G.; Tavares, A.P.M. A strategy for improving peroxidase stability via immobilization on surface modified multi-walled carbon nanotubes. J. Chem. Technol. Biotechnol. 2015, 90, 1570–1578. [Google Scholar] [CrossRef]
- Albayrak, N.; Yang, S.T. Production of galacto-oligosaccharides from lactose by Aspergillus oryzae betα-galactosidase immobilized on cotton cloth. Biotechnol. Bioeng. 2002, 77, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.J.; Park, J.M.; Yang, J.W. Continuous production of galacto-oligosaccharides from lactose by Bullera singularis beta galactosidase immobilized in chitosan beads. Process Biochem. 1998, 33, 787–792. [Google Scholar] [CrossRef]
- Zheng, P.; Yu, H.; Sun, Z.; Ni, Y.; Zhang, W.; Fan, Y.; Xu, Y. Production of galacto-oligosaccharides by immobilized recombinant β-galactosidase from Aspergillus candidus. Biotechnol. J. 2006, 1, 1464–1470. [Google Scholar] [CrossRef] [PubMed]
- Plou, F.J.; Rodriguez-Colinas, B.; Fernandez-Arrojo, L.; Ballesteros, A. Low-lactose, prebiotic-enriched milk. In Probiotics, Prebiotics, and Synbiotics; Watson, R.R., Preedy, V.R., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 47–57. [Google Scholar]
- Rodriguez-Colinas, B.; De Abreu, M.A.; Fernandez-Arrojo, L.; De Beer, R.; Poveda, A.; Jimenez-Barbero, J.; Haltrich, D.; Ballesteros, A.O.; Fernandez-Lobato, M.; Plou, F.J. Production of galacto-oligosaccharides by the β-galactosidase from Kluyveromyces lactis: Comparative analysis of permeabilized cells versus soluble enzyme. J. Agric. Food Chem. 2011, 59, 10477–10484. [Google Scholar] [CrossRef] [PubMed]
- Urrutia, P.; Rodriguez-Colinas, B.; Fernandez-Arrojo, L.; Ballesteros, A.O.; Wilson, L.; Illanes, A.; Plou, F.J. Detailed analysis of galactooligosaccharides synthesis with β-galactosidase from Aspergillus oryzae. J. Agric. Food Chem. 2013, 61, 1081–1087. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Operation Time 1 (h) | Gal (g/L) | Glc (g/L) | Lact (g/L) | 4′-Gal-Lact 2 (g/L) | Tetrasaccharide 3 (g/L) | Other GOS (g/L) | Total GOS (g/L) |
|---|---|---|---|---|---|---|---|
| 0 | 26.1 | 43.9 | 13.6 | 3.1 | 0.7 | 12.6 | 16.5 |
| 48 | 4.1 | 23.0 | 48.6 | 15.7 | 5.3 | 3.3 | 24.3 |
| 213 | 0.3 | 6.7 | 83.0 | 9.0 | 0.7 | 0.3 | 10.0 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez-Colinas, B.; Fernandez-Arrojo, L.; Santos-Moriano, P.; Ballesteros, A.O.; Plou, F.J. Continuous Packed Bed Reactor with Immobilized ?-Galactosidase for Production of Galactooligosaccharides (GOS). Catalysts 2016, 6, 189. https://doi.org/10.3390/catal6120189
Rodriguez-Colinas B, Fernandez-Arrojo L, Santos-Moriano P, Ballesteros AO, Plou FJ. Continuous Packed Bed Reactor with Immobilized ?-Galactosidase for Production of Galactooligosaccharides (GOS). Catalysts. 2016; 6(12):189. https://doi.org/10.3390/catal6120189
Chicago/Turabian StyleRodriguez-Colinas, Barbara, Lucia Fernandez-Arrojo, Paloma Santos-Moriano, Antonio O. Ballesteros, and Francisco J. Plou. 2016. "Continuous Packed Bed Reactor with Immobilized ?-Galactosidase for Production of Galactooligosaccharides (GOS)" Catalysts 6, no. 12: 189. https://doi.org/10.3390/catal6120189
APA StyleRodriguez-Colinas, B., Fernandez-Arrojo, L., Santos-Moriano, P., Ballesteros, A. O., & Plou, F. J. (2016). Continuous Packed Bed Reactor with Immobilized ?-Galactosidase for Production of Galactooligosaccharides (GOS). Catalysts, 6(12), 189. https://doi.org/10.3390/catal6120189