Effect of Water Clustering on the Activity of Candida antarctica Lipase B in Organic Medium



Abstract
:1. Introduction
2. Results and Discussion
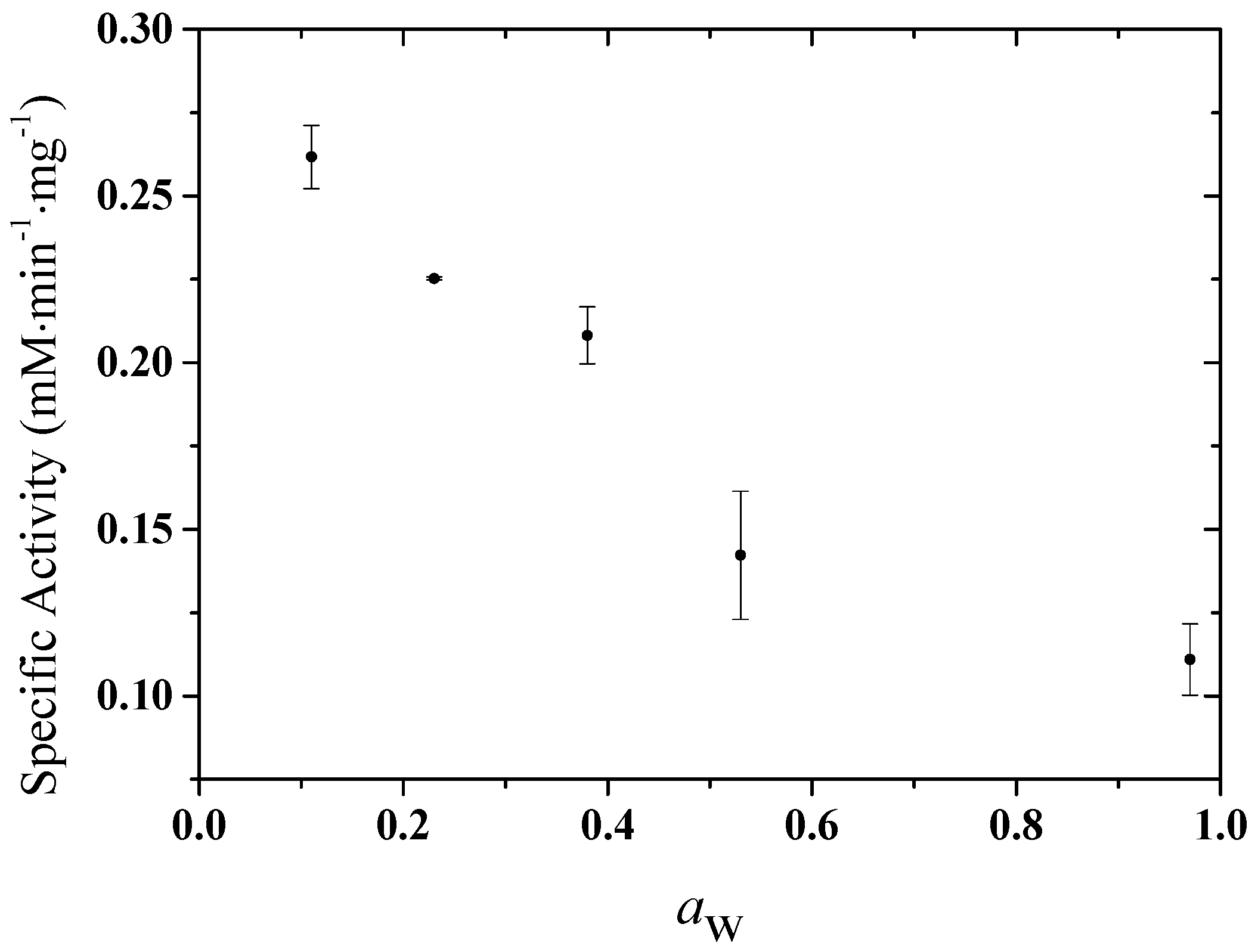
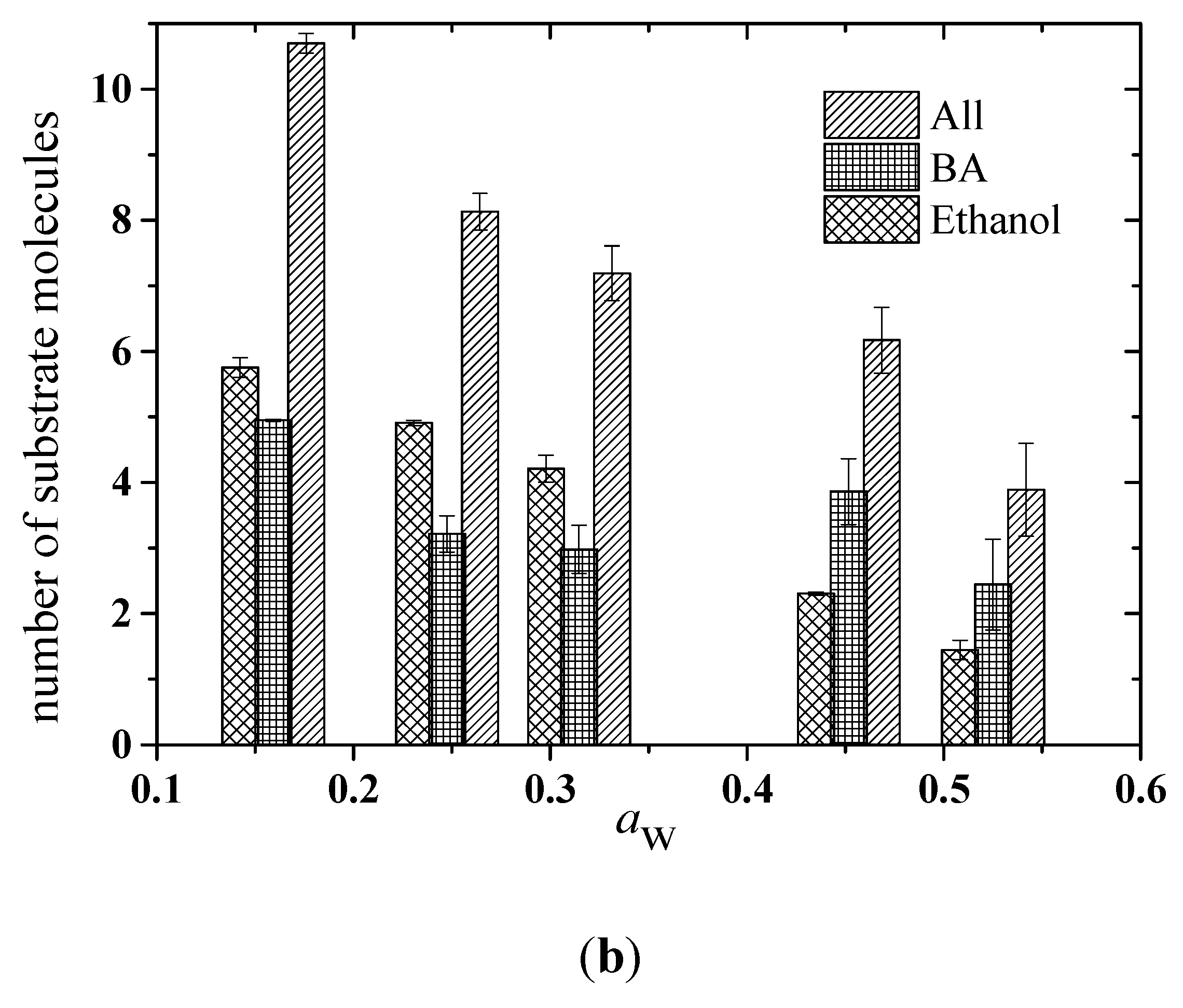
2.1. Experimental Results
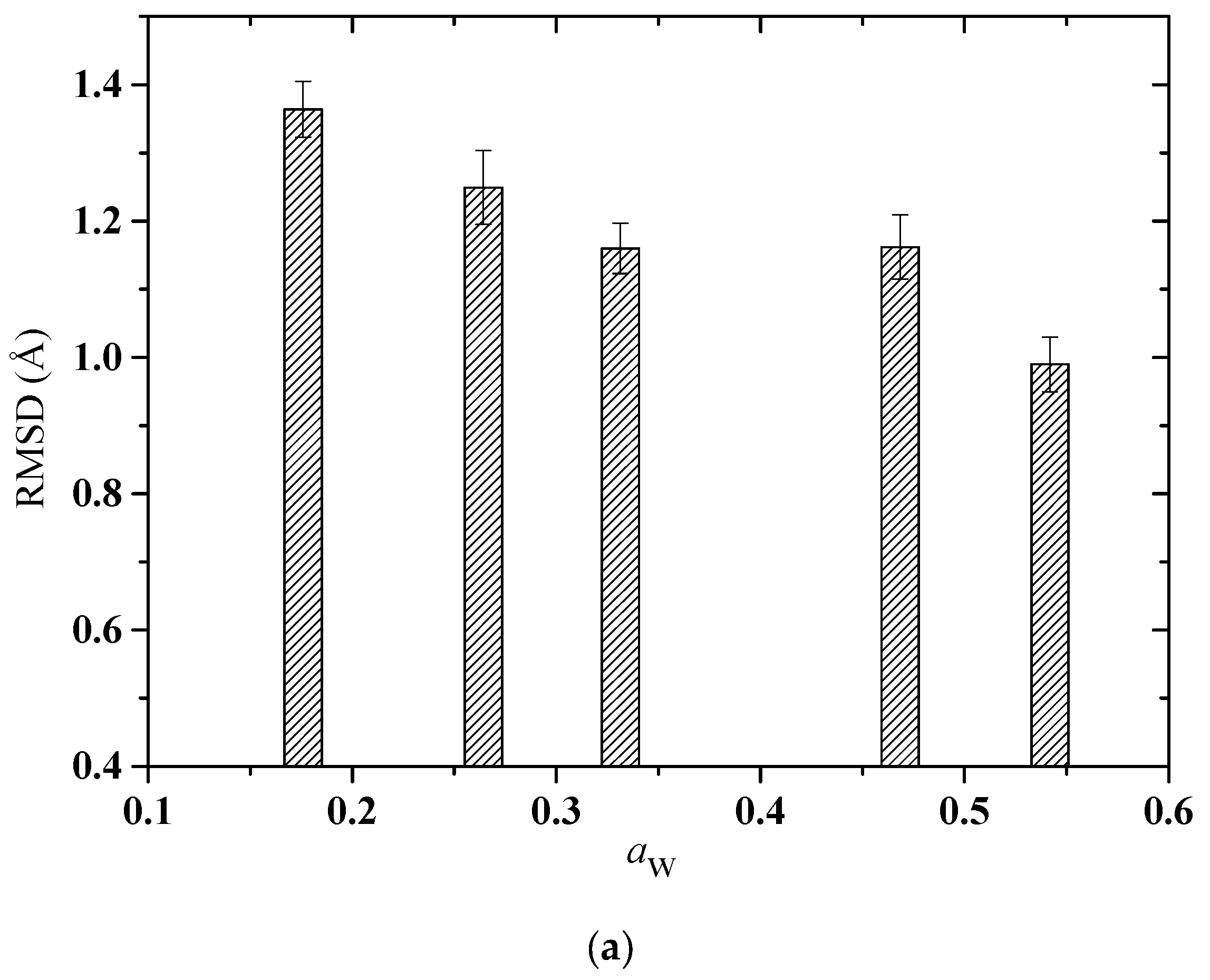
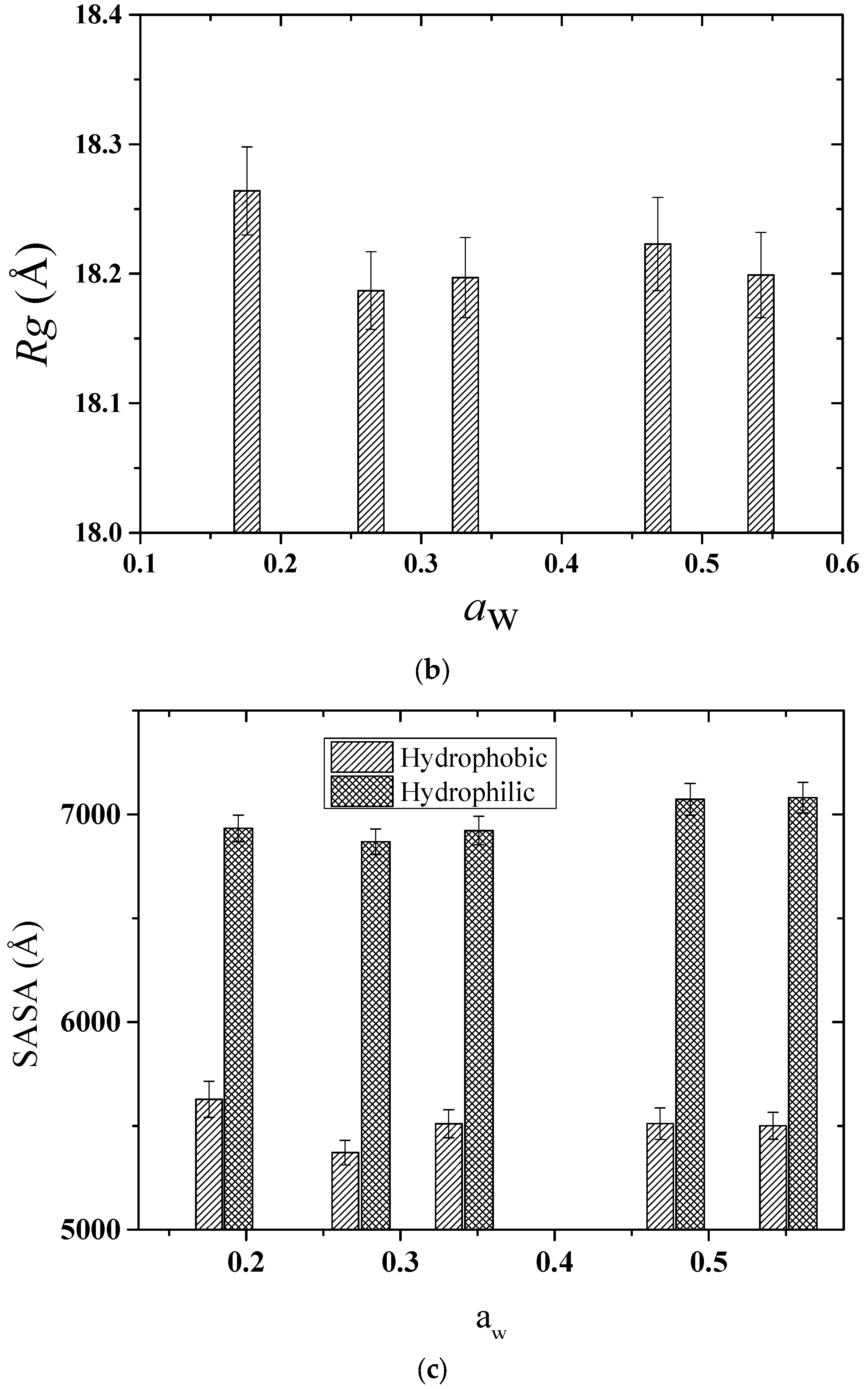
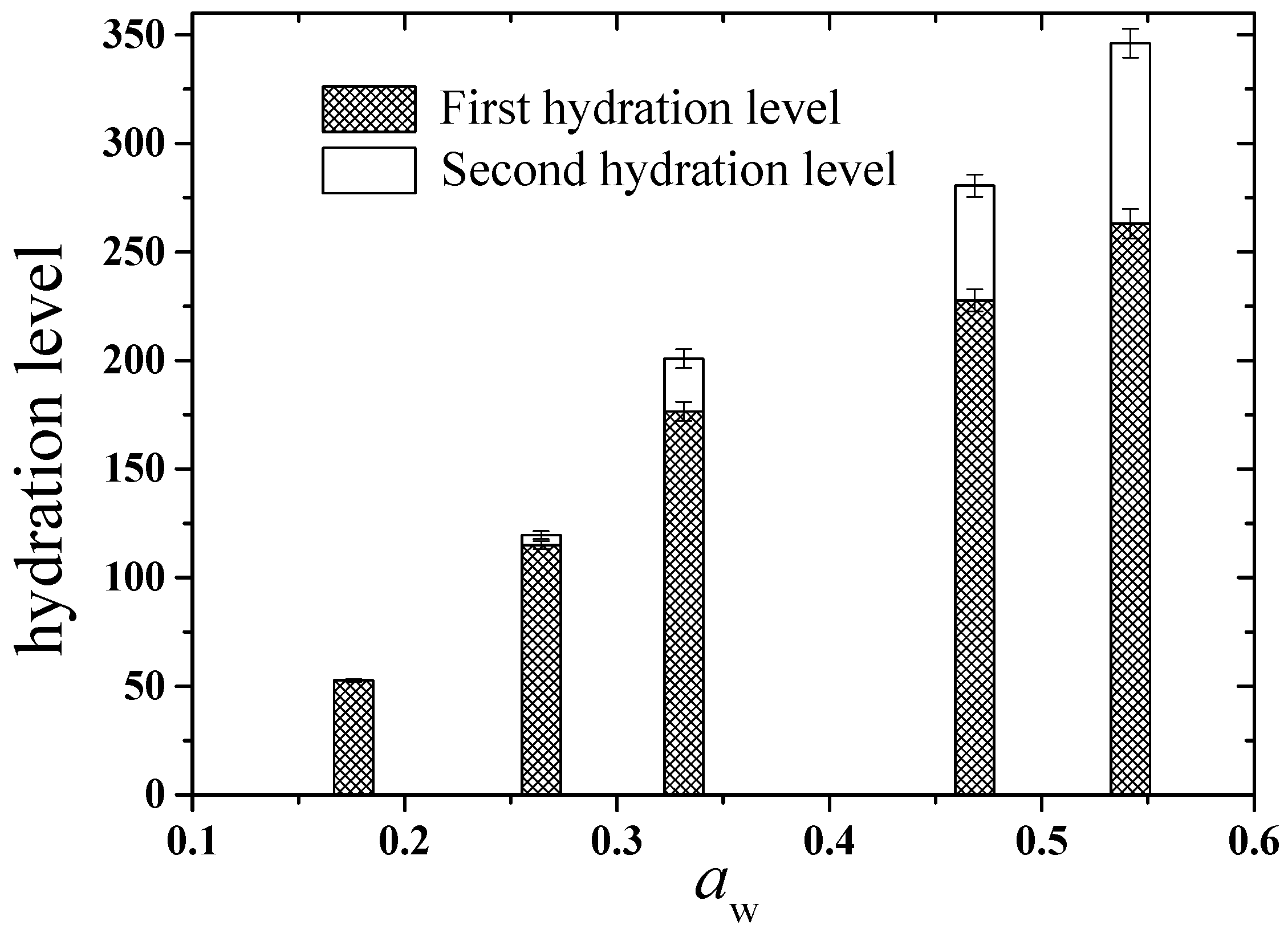
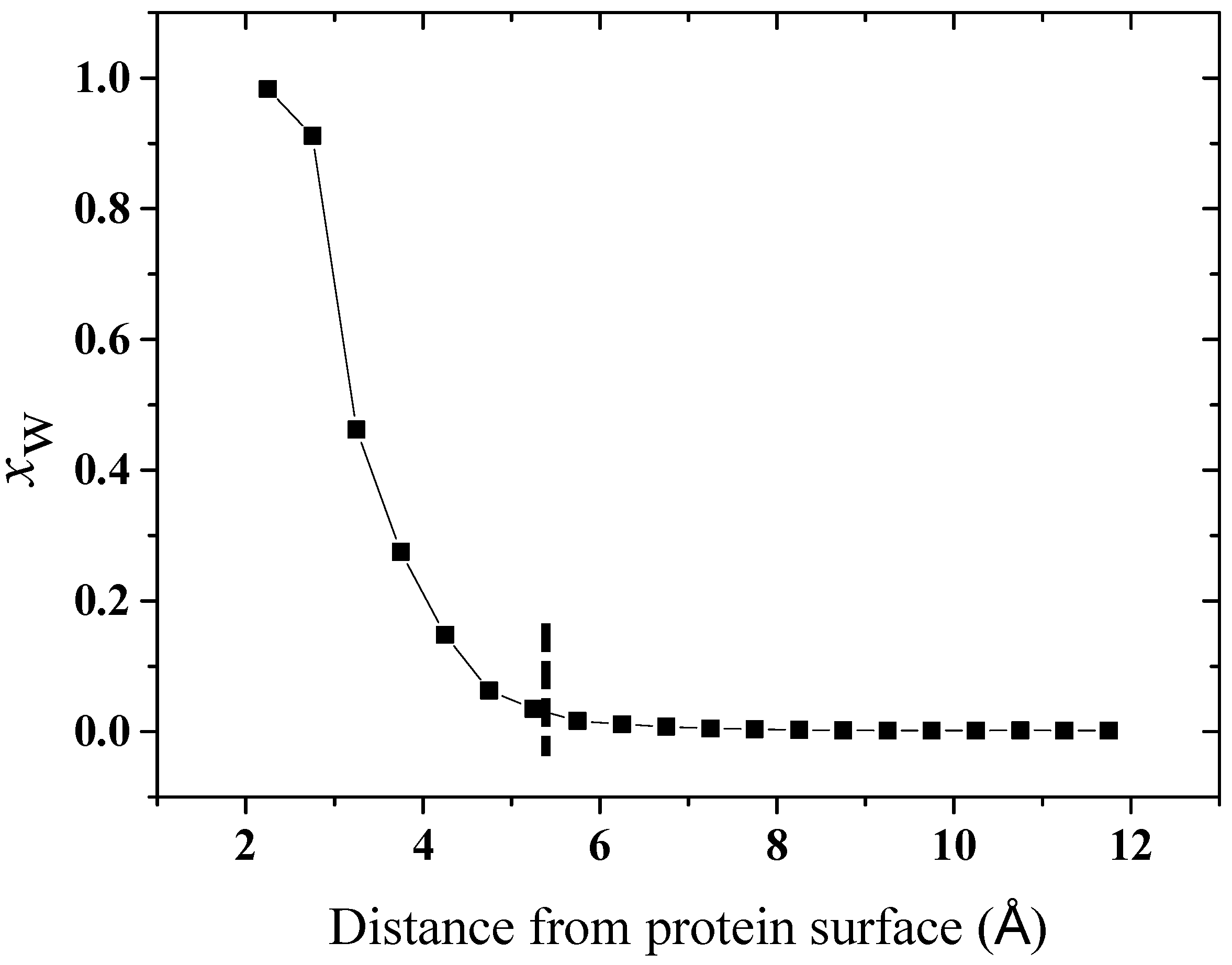
2.2. Computational Results
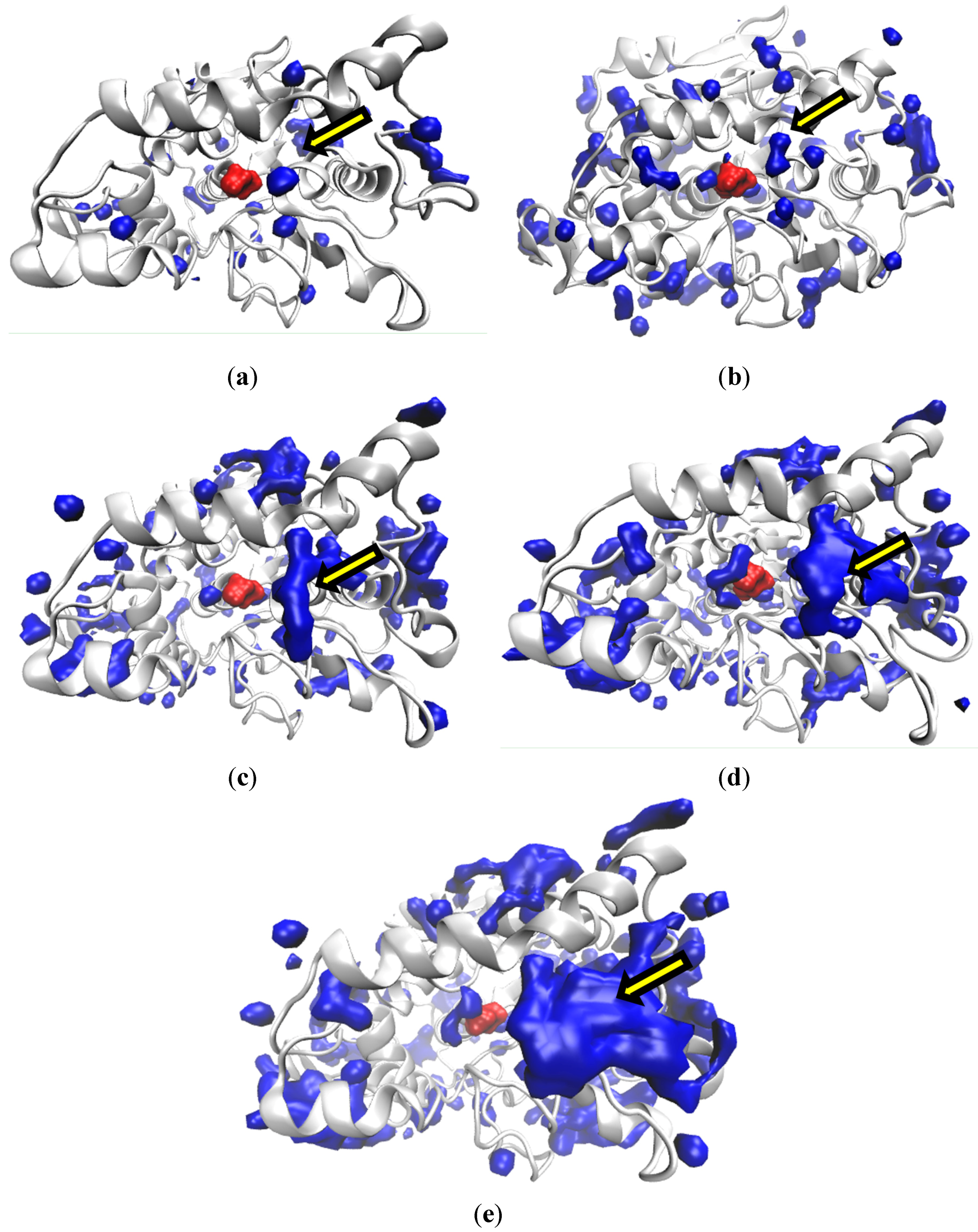
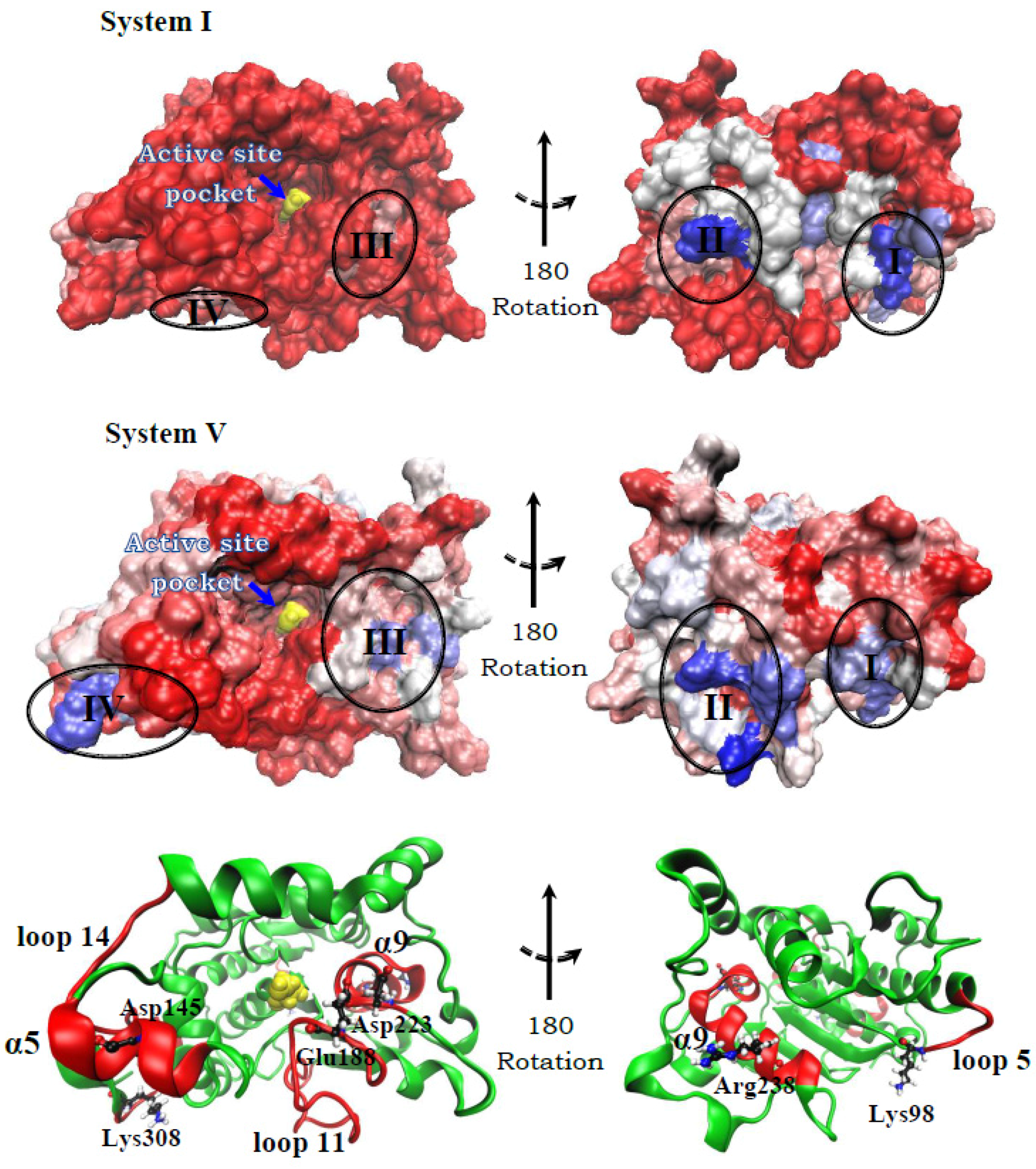
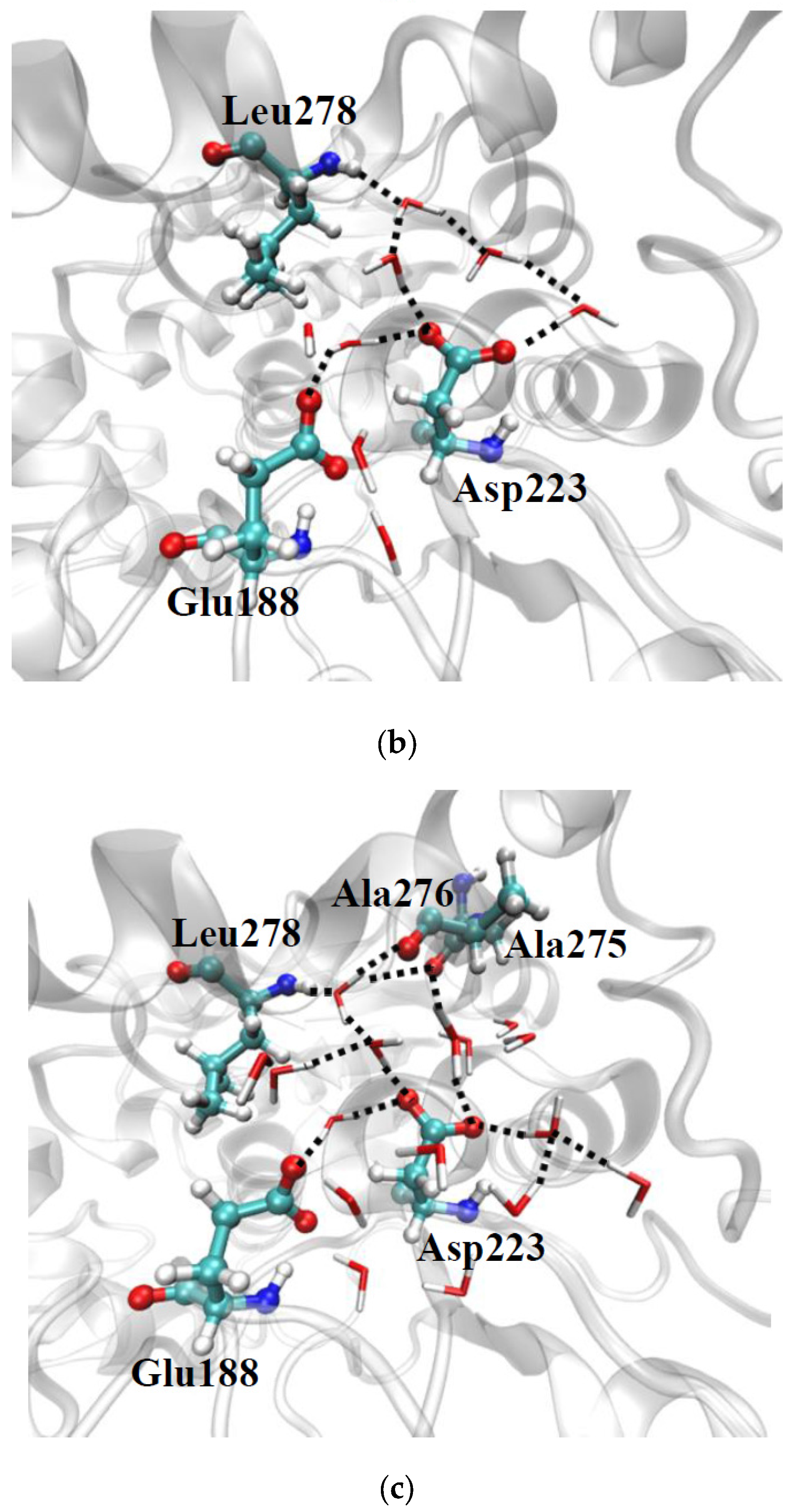
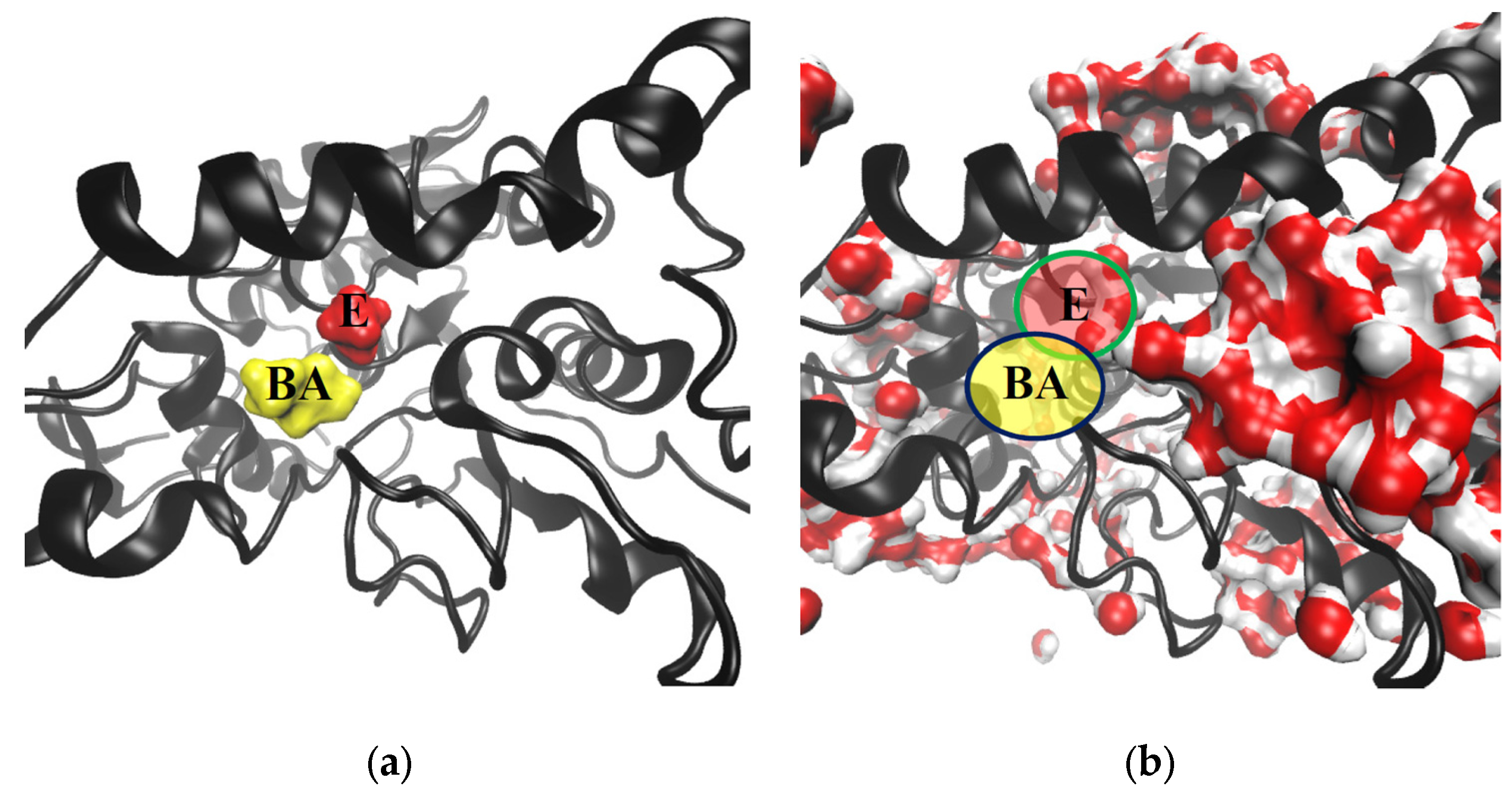
2.2.1. Structure
2.2.2. Hydration Level
3. Methods
3.1. Experimental Procedures
3.1.1. Materials
3.1.2. Esterification Reactions
3.1.3. GC Analysis
3.1.4. Kinetic Parameters Estimation
3.2. Molecular Dynamics Simulations
3.2.1. Structure
3.2.2. System Setup
3.2.3. Simulations
3.2.4. Water activity Calculation
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Houde, A.; Kademi, A.; Leblanc, D. Lipases and Their Industrial Applications: An Overview. Appl. Biochem. Biotechnol. 2004, 118, 155–170. [Google Scholar] [CrossRef]
- Adlercreutz, P. Immobilisation and application of lipases in organic media. Chem. Soc. Rev. 2013, 42, 6406–6436. [Google Scholar] [CrossRef] [PubMed]
- Klibanov, A.M. Improving enzymes by using them in organic solvents. Nature 2001, 409, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, P.; Bhunia, B. Industrial application of lipase: A review. Biopharm J. 2015, 1, 41–47. [Google Scholar]
- Soares, C.M.; Teixeira, V.H.; Baptista, A.M. Protein structure and dynamics in nonaqueous solvents: Insights from molecular dynamics simulation studies. Biophys. J. 2003, 84, 1628–1641. [Google Scholar] [CrossRef]
- Xu, Z.; Suzawa, V.; Focht, K.; Clark, D.S.; Dordick, J.S. Enzymatic catalysis and dynamics in low-water environments. Proc. Natl. Acad. Sci. USA 1992, 89, 1100–1104. [Google Scholar]
- Valivety, R.H.; Halling, P.J.; Peilow, A.D.; Macrae, A.R. Lipases from different sources vary widely in dependence of catalytic activity on water activity. Biochim. Biophys. Acta 1992, 1122, 143–146. [Google Scholar] [CrossRef]
- Tsitsimpikou, C.; Daflos, H.; Kolisis, F.N. Comparative studies on the sugar esters synthesis catalysed by Candida antarctica and Candida rugosa lipases in hexane. J. Mol. Catal. B Enzym. 1997, 3, 189–192. [Google Scholar] [CrossRef]
- Mora-pale, J.M.; Munguia, S.P.; Gonzalez-Mejia, J.C.; Dordick, J.S.; Barzana, E. The Lipase-Catalyzed Hydrolysis of Lutein Diesters in Non-Aqueous Media is Favored at Extremely Low Water Activities. Biotechnol. Bioeng. 2007, 98, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Miyawaki, Y.; Adachi, S.; Nakanishi, K.; Matsuno, R. Equilibrium constant for lipase-catalyzed condensation of mannose and lauric acid in water-miscible organic solvents. Enzym. Microb. Technol. 2001, 29, 494–498. [Google Scholar] [CrossRef]
- Chamouleau, F.; Coulon, D.; Girardin, M.; Ghoul, M. Influence of water activity and water content on sugar esters lipase-catalyzed synthesis in organic media. J. Mol. Catal. B Enzym. 2001, 11, 949–954. [Google Scholar] [CrossRef]
- Humeau, C.; Girardin, M.; Rovel, B.; Miclo, A. Effect of the thermodynamic water activity and the reaction medium hydrophobicity on the enzymatic synthesis of ascorbyl palmitate. J. Biotechnol. 1998, 63, 1–8. [Google Scholar] [CrossRef]
- Jin, Z.; Liang, S.; Zhang, X.; Han, S.; Ren, C.; Lin, Y.; Zheng, S. Synthesisof Fructose Laurate Esters Catalyzed by a CALB-displaying Pichia pastoris Whole-cell Biocatalyst in a Non-aqueous System. Biotechnol. Bioeng. 2013, 18, 365–374. [Google Scholar] [CrossRef]
- Secundo, F.; Carrea, G.; Soregaroli, C.; Varinelli, D.; Morrone, R.; Molecolare, R.; Bianco, M. Activity of Different Candida antarctica Lipase B Formulations in Organic Solvents. Biotechnol. Bioeng. 2001, 73, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Banik, S.D.; Nordblad, M.; Woodley, J.M.; Peters, G.H. A Correlation between the Activity of Candida antarctica Lipase B and Di ff erences in Binding Free Energies of Organic Solvent and Substrate. ACS Catal. 2016, 6, 6350–6361. [Google Scholar] [CrossRef]
- Ates, S.; Türk, B.; Bayraktar, E.; Güvenc, A. Enhanced ethyl butyrate production using immobilized lipase. Artif. Cells Nanomed. Biotechnol. 2013, 41, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-nogales, J.M.; Roura, E.; Contreras, E. Biosynthesis of ethyl butyrate using immobilized lipase: A statistical approach. Process Biochem. 2005, 40, 63–68. [Google Scholar] [CrossRef]
- Baptista, M. Water Dependent Properties of Cutinase in Nonaqueous Solvents: A Computational Study of Enantioselectivity. Biophys. J. 2005, 89, 999–1008. [Google Scholar]
- Micaêlo, N.M.; Soares, C.M. Modeling hydration mechanisms of enzymes in nonpolar and polar organic solvents. FEBS J. 2007, 274, 2424–2436. [Google Scholar] [CrossRef] [PubMed]
- Schröder, C.; Rudas, T.; Boresch, S.; Steinhausera, O. Simulation studies of the protein-water interface. I. Properties at the molecular resolution. J. Chem. Phys. 2006, 124, 234907. [Google Scholar] [CrossRef] [PubMed]
- Gruber, C.C.; Pleiss, J. Enzymatic Lipase B from Candida antarctica binds to hydrophobic substrate—Water interfaces via hydrophobic anchors surrounding the active site entrance. J. Mol. Catal. B Enzym. 2012, 84, 48–54. [Google Scholar] [CrossRef]
- Kulschewski, T.; Pleiss, J. Binding of Solvent Molecules to a Protein Surface in Binary Mixtures Follows a Competitive Langmuir Model. Langmuir 2016, 32, 8960–8968. [Google Scholar] [CrossRef] [PubMed]
- Goderis, H.L.; Ampe, G.; Feyten, M.P.; Fouwe, B.L.; Guffens, W.M.; van Cauwenbergh, S.M.; Tobback, P.P. Lipase-catalysed ester exchange reaction in organic media with controlled humidity. Biotechnol. Bioeng. 1987, 30, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Uppenberg, J.; Hansen, M.T.; Patkar, S.; Jones, T.A. The sequence, crystal structure determination and refinement of two crystal forms of lipase B from Candida antarctica. Structure 1994, 2, 293–308. [Google Scholar] [CrossRef]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kalé, L.; Schulten, K. Scalable molecular dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef] [PubMed]
- AMacKerell, D.; Bashford, D.J.; Bellott, M.; Dunbrack, R.L.; Evanseck, J.D.; Field, M.J.; Fischer, S.; Gao, J.; Guo, H.; Ha, S.; et al. All-Atom Empirical Potential for Molecular Modeling and Dynamics Studies of Proteins. J. Phys. Chem. B 1998, 102, 3586–3616. [Google Scholar] [CrossRef] [PubMed]
- Mackerell, A.D.; Feig, M.; Brooks, C.L. Extending the treatment of backbone energetics in protein force fields: Limitations of gas-phase quantum mechanics in reproducing protein conformational distributions in molecular dynamics simulations. J. Comput. Chem. 2004, 25, 1400–1415. [Google Scholar] [CrossRef] [PubMed]
- Vorobyov, I.; Anisimov, V.M.; Greene, S.; Venable, R.M.; Moser, A.; Pastor, R.W.; MacKerell, A.D. Additive and classical drude polarizable force fields for linear and cyclic ethers. J. Chem. Theory Comput. 2007, 3, 1120–1133. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926. [Google Scholar] [CrossRef]
- Vanquelef, E.; Simon, S.; Marquant, G.; Garcia, E.; Klimerak, G.; Delepine, J.C.; Cieplak, P.; Dupradeau, F.Y. R.E.D. Server: A web service for deriving RESP and ESP charges and building force field libraries for new molecules and molecular fragments. Nucleic Acids Res. 2011, 39, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Wedberg, R.; Abildskov, J.; Peters, G.H. Protein dynamics in organic media at varying water activity studied by molecular dynamics simulation. J. Phys. Chem. B 2012, 116, 2575–2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixit, S.B.; Chipot, C.; Poincare, H.; Vandœu, V. Can Absolute Free Energies of Association Be Estimated from Molecular Mechanical Simulations? The Biotin—Streptavidin System Revisited. J. Phys. Chem. A 2001, 105, 9795–9799. [Google Scholar] [CrossRef]
- Frenkel, D.; Smit, B. Understanding Molecular Simulation; Academic Press: Cambridge, MA, USA, 2002. [Google Scholar]
- Zacharias, M.; Straatsma, T.P.; Mccammon, J.A.; Zacharias, M.; Straatsma, T.P.; Mccammon, J.A. Separation-shifted scaling, a new scaling method for Lennard-Jones interactions in thermodynamic integration Separation-shifted scaling, a new scaling method for Lennard-Jones interactions in thermodynamic integration. J. Chem. Phys. 1994, 100, 9025–9031. [Google Scholar] [CrossRef]
- Gonçalves, P.F.B.; Stassen, H. Calculation of the free energy of solvation from molecular dynamics simulations. Pure Appl. Chem. 2004, 76, 231–240. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | nw | nMTBE | xw | Bulk xw | aw | Specific Activity |
|---|---|---|---|---|---|---|
| I | 54 | 1788 | 0.029 | 0.0007 | 0.18 | 0.26172 |
| II | 120 | 1788 | 0.063 | 0.0011 | 0.26 | 0.22522 |
| III | 204 | 1788 | 0.102 | 0.0014 | 0.33 | 0.20815 |
| IV | 286 | 1788 | 0.138 | 0.0020 | 0.47 | 0.1422 |
| V | 354 | 1752 | 0.168 | 0.0023 | 0.54 | 0.11095 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dutta Banik, S.; Nordblad, M.; Woodley, J.M.; Peters, G.H. Effect of Water Clustering on the Activity of Candida antarctica Lipase B in Organic Medium. Catalysts 2017, 7, 227. https://doi.org/10.3390/catal7080227
Dutta Banik S, Nordblad M, Woodley JM, Peters GH. Effect of Water Clustering on the Activity of Candida antarctica Lipase B in Organic Medium. Catalysts. 2017; 7(8):227. https://doi.org/10.3390/catal7080227
Chicago/Turabian StyleDutta Banik, Sindrila, Mathias Nordblad, John M. Woodley, and Günther H. Peters. 2017. "Effect of Water Clustering on the Activity of Candida antarctica Lipase B in Organic Medium" Catalysts 7, no. 8: 227. https://doi.org/10.3390/catal7080227
APA StyleDutta Banik, S., Nordblad, M., Woodley, J. M., & Peters, G. H. (2017). Effect of Water Clustering on the Activity of Candida antarctica Lipase B in Organic Medium. Catalysts, 7(8), 227. https://doi.org/10.3390/catal7080227