
The Potential Contribution of Biopolymeric Particles in Lung Tissue Regeneration of COVID-19 Patients

 , ,
, ,
Abstract
:1. Introduction
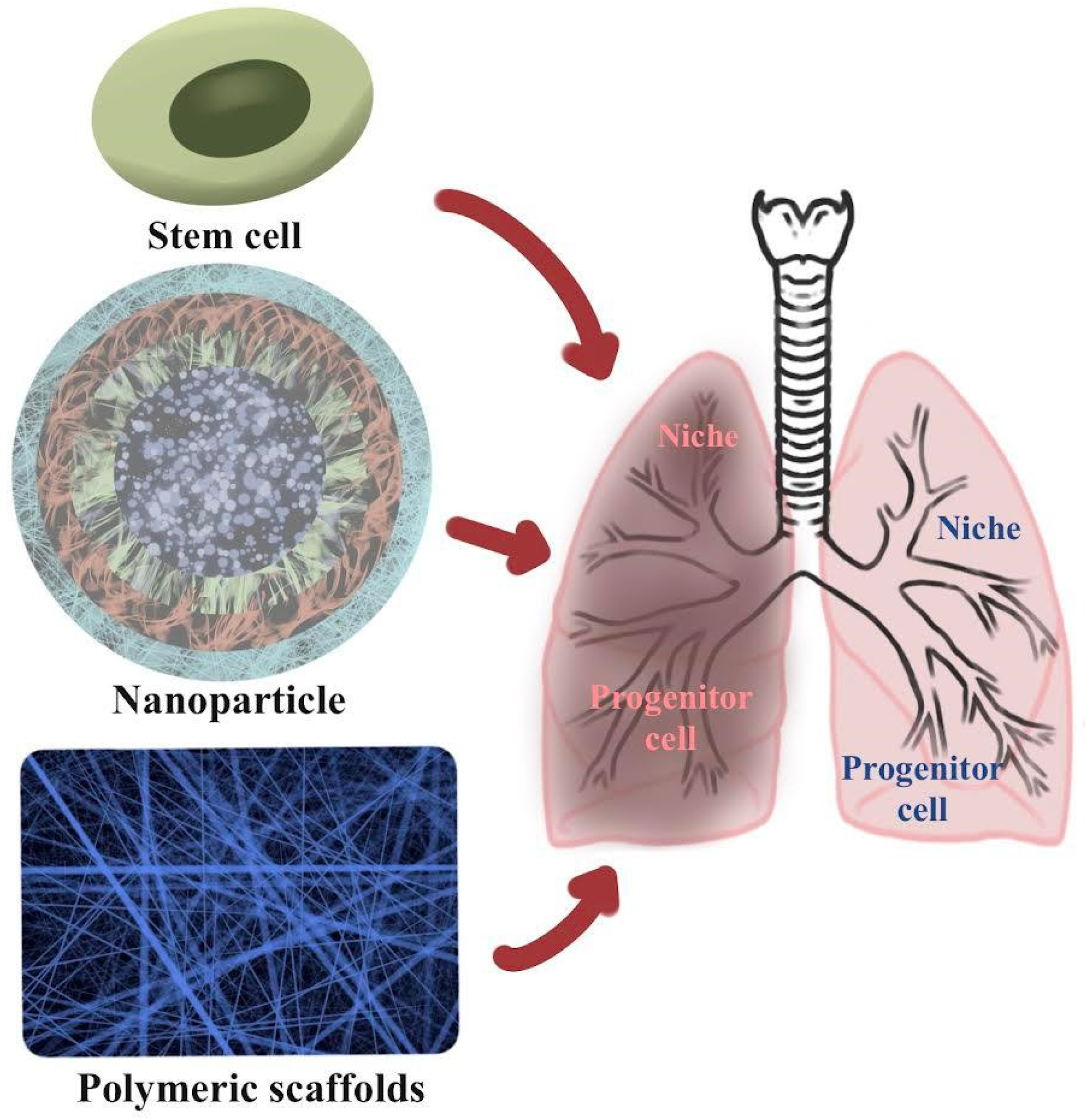
2. Regeneration of Lung Tissues Using Biopolymeric Nanoparticles for COVID-19 Patients
3. Biopolymeric Materials of Stem Cells for Lung Tissue Regeneration
4. Regeneration of Lung Tissue Using Biopolymeric Scaffolds
Application Procedure of the Polymer Scaffolds and Polymer Nanoparticles
5. Discussion
6. Conclusions
7. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| COVID-19 | coronavirus disease 2019 |
| IONPs | magnetic iron oxide nanoparticles |
| Fe2O3 | ferric oxide |
| Fe3O4 | ferrosoferric oxide |
| SARS-CoV-2 | severe acute respiratory syndrome coronavirus 2 |
| RDB | receptor binding domain |
| NPs | nanoparticles |
| MOI | multiplicities of infection |
| MSCs | mesenchymal stem cells |
| RNA | ribonucleic acid |
| NEM | Nanoelectromechanical |
| DNA | deoxyribonucleic acid |
| NRP1 | neuropilin-1 |
| DCs | dendritic cells |
| LNPs | lipid nanoparticles |
| HPD | hesperidin |
| ACE2 | angiotensin-converting enzyme 2 |
| LPS | lipopolysaccharide |
| ARDS | acute respiratory distress syndrome |
| qRT-PCR | quantitative real-time polymerase chain reaction |
| AL-Rem | remdesivir |
| Calu-3 | submucosal gland cell line |
| PLGA | poly (lactic-co-glycolic acid) |
| RdRp | RNA-dependent RNA polymerase |
| NASAR | novo design of UTRs, the optimal combination of 5′ and 3′ UTR |
| TT3 | total triiodothyronine |
| FDA | Food and Drug Administration |
| MC3 | composition of composite material 3 |
| LOAC | lung-on-a-chip |
| NIR-II | second near infrared |
| CT | computed tomography |
| GLCM | gray-level co-occurrence matrix |
| hACE2 | human angiotensin I-converting enzyme 2 |
| IPF | idiopathic pulmonary fibrosis |
| AEC II | type II alveolar epithelial cells |
| PER NPs | surface-engineered nanoparticles |
| MOMC | monocyte-derived multipotent cells |
| AST | astaxanthin |
| TRA | trametinib |
| MMP-2 | matrix metalloproteinase 2 |
| UCNPs | upconverting nanoparticles |
| FITC | fluorescein isothiocyanate |
| LCSTs | lower critical solution temperatures |
| MERS-CoV | Middle East respiratory syndrome coronavirus |
| H1N1 | swine flu |
| LIONs | lipid inorganic nanoparticles |
| IgG | immunoglobulin G |
| ZnO | zinc oxide |
| KIM-1 | kidney injury molecule-1 |
| miRNAs | MicroRNAs |
| LPH | lipid polymer hybrid |
| SSRIs | selective serotonin reuptake inhibitors |
| FH | fluoxetine hydrochloride |
| hsACE2 | a soluble form of hACE2 |
| MHRA | Medicines and Healthcare Products Regulatory Agency |
| ABL-2 | Abelson-related gene |
| IL-1beta | interleukin-1-beta |
| IL-6 | interleukin 6 |
| IL-8 | interleukin-8 |
| Mip-1alpha | macrophage inflammatory protein-1 alpha |
| TNF-alpha | tumor necrosis factor alpha |
| PI4Kiiibeta | PI4KB phosphatidylinositol 4-kinase beta |
| IBD | inflammatory bowel disease |
| AlphaMSH | alpha-melanocyte-stimulating hormone |
| MC1R | melanocortin 1 receptor |
| NO | nitric oxide |
| AFM | atomic force microscopy |
| ROS | reactive oxygen species |
| CCL2 | C–C motif chemokine ligand 2 |
| Th17 | T helper 17 |
| IL-6 | interleukin-6 |
| IL-17 | interleukin-17 |
| IL-18 | interleukin-18 |
| IL-21 | interleukin-21 |
| IL-23 | interleukin-23 |
| EVs | extracellular vesicles |
| SARDS | severe acute respiratory distress syndrome |
| ALI | acute lung injury |
| ASCs | adipose stem cells |
| ICU | intensive care unit |
| FiO2 | fraction of inspired oxygen |
| SaO2 | oxygen saturation |
| PO2 | partial pressure of oxygen |
| LSCs | lung stem cells |
| 7nAChR | vagal-alpha7 nicotinic acetylcholine receptor |
| MSCs-Exo | mesenchymal stem cells and their exosomes |
| BASCs | bronchioalveolar stem cells |
| ALOs | adult lung organoids |
| hiPSC-CMs | human induced pluripotent stem cell-derived cardiomyocytes |
| CDC42 | cell division cycle 42 |
| TGF | transforming growth factor |
| iAT2s | alveolar type 2-like cells |
| UC-MSCs | umbilical cord-mesenchymal stem cells |
| P (VDF-TrFE) | poly (vinylidene fluoride-co-trifluoroethylene) |
| HBD2 | human beta defensin 2 |
| MRC-5 | Medical Research Council cell strain 5 |
| iPSCs | induced pluripotent stem cells |
| COPD | chronic obstructive pulmonary disease |
| cAMP | cyclic adenosine monophosphate |
| MPE | multiphoton excited |
| IPF | idiopathic pulmonary fibrosis |
| PLA | polylactic acid or polylactide |
| GO | graphene oxide |
| DOX | doxorubicin |
| TiO2 | titanium dioxide |
| XRD | X-ray powder diffraction |
| SEM | scanning electron microscopy |
| TEM | transmission electron microscopy |
| SLES | sodium lauryl ether sulfate |
| SDS | sodium dodecyl sulfate |
| GAGs | glycosaminoglycans |
| ECM | extracellular matrix |
| hBECs | human bronchial epithelial cells |
| hLFs | lung fibroblasts |
| NETs | neutrophil extracellular traps |
| MPO | myeloperoxidase |
| PCL | poly(caprolactone) |
| AgNP | silver nanoparticle |
| AuNPs | gold nanoparticle |
| hAFS | human amniotic fluid stem |
| ToF-SIMS | time-of-flight secondary ion mass spectrometry |
| PA-VS | vinyl-sulfonated polyamines |
References
- Abdellatif, A.A.; Tawfeek, H.M.; Abdelfattah, A.; Batiha, G.E.-S.; Hetta, H.F. Recent updates in COVID-19 with emphasis on inhalation therapeutics: Nanostructured and targeting systems. J. Drug Deliv. Sci. Technol. 2021, 63, 102435. [Google Scholar] [CrossRef]
- Elashiry, M.; Elsayed, R.; Elashiry, M.M.; Rashid, M.H.; Ara, R.; Arbab, A.S.; Elawady, A.R.; Hamrick, M.; Liu, Y.; Zhi, W.; et al. Proteomic Characterization, Biodistribution, and Functional Studies of Immune-Therapeutic Exosomes: Implications for Inflammatory Lung Diseases. Front. Immunol. 2021, 12, 636222. [Google Scholar] [CrossRef]
- Idris, A.; Davis, A.; Supramaniam, A.; Acharya, D.; Kelly, G.; Tayyar, Y.; West, N.; Zhang, P.; McMillan, C.L.; Soemardy, C.; et al. A SARS-CoV-2 targeted siRNA-nanoparticle therapy for COVID-19. Mol. Ther. 2021, 29, 2219–2226. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Zhao, Z.; Lan, Q.; Zhou, H.; Mai, Z.; Wang, Y.; Ding, X.; Zhang, W.; Pi, J.; Evans, C.E.; et al. Nasal Delivery of Hesperidin/Chitosan Nanoparticles Suppresses Cytokine Storm Syndrome in a Mouse Model of Acute Lung Injury. Front. Pharmacol. 2021, 11, 592238. [Google Scholar] [CrossRef]
- Jing, W.; Procko, E. ACE2-based decoy receptors for SARS coronavirus 2. Proteins Struct. Funct. Bioinform. 2021, 89, 1065–1078. [Google Scholar] [CrossRef]
- Kipshidze, N.; Iversen, P.; Porter, T.R.; Kipshidze, N.; Siddiqui, F.; Dangas, G.; Fareed, J. Targeted, Site-Specific, Delivery Vehicles of Therapeutics for COVID-19 Patients. Brief Review. Clin. Appl. Thromb. 2020, 26, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Muraca, M.; Pessina, A.; Pozzobon, M.; Dominici, M.; Galderisi, U.; Lazzari, L.; Parolini, O.; Lucarelli, E.; Perilongo, G.; Baraldi, E. Mesenchymal stromal cells and their secreted extracellular vesicles as therapeutic tools for COVID-19 pneumonia? J. Control. Release 2020, 325, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Vartak, R.; Patil, S.M.; Saraswat, A.; Patki, M.; Kunda, N.K.; Patel, K. Aerosolized nanoliposomal carrier of remdesivir: An effective alternative for COVID-19 treatment in vitro. Nanomedicine 2021, 16, 1187–1202. [Google Scholar] [CrossRef]
- Wu, J.; Wang, H.; Li, B. Structure-aided ACEI-capped remdesivir-loaded novel PLGA nanoparticles: Toward a computational simulation design for anti-SARS-CoV-2 therapy. Phys. Chem. Chem. Phys. 2020, 22, 28434–28439. [Google Scholar] [CrossRef]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef]
- Moitra, P.; Alafeef, M.; Dighe, K.; Sheffield, Z.; Dahal, D.; Pan, D. Synthesis and characterisation of N-gene targeted NIR-II fluorescent probe for selective localisation of SARS-CoV-2. Chem. Commun. 2021, 57, 6229–6232. [Google Scholar] [CrossRef]
- Costa, C.A. Likely positive effect of alpha-linolenic acid supplementation on the lung and the negative effect of it on the adipose tissue of COVID-19 patients. Nutr. Food Sci. 2021, 51, 878–882. [Google Scholar] [CrossRef]
- Morozov, S.P.; Chernina, V.Y.; Andreychenko, A.E.; Vladzymyrskyy, A.V.; Mokienko, O.A.; Gombolevskiy, V.A. How does artificial intelligence effect on the assessment of lung damage in COVID-19 on chest CT scan? Digit. Diagn. 2021, 2, 27–38. [Google Scholar] [CrossRef]
- Conti, P.; Ronconi, G.; Caraffa, A.; Gallenga, C.; Ross, R.; Frydas, I.; Kritas, S. Induction of pro-inflammatory cytokines (IL-1 and IL-6) and lung inflammation by COVID-19: Anti-inflammatory strategies. J. Biol. Regul. Homeost. Agents 2020, 34, 327–331. [Google Scholar] [PubMed]
- Yasar, H.; Ceylan, M. A novel comparative study for detection of Covid-19 on CT lung images using texture analysis, machine learning, and deep learning methods. Multimed. Tools Appl. 2021, 80, 5423–5447. [Google Scholar] [CrossRef] [PubMed]
- Arrossi, A.V.; Farver, C. The pulmonary pathology of COVID-19. Clevel. Clin. J. Med. 2020, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Micu, C.-E.; Smărăndescu, R.A.; Arghir, I.-A.; Trenchea, M.; Barbu, B.A.; Handra, C.M. Beneficial effect of hyperbaric oxygen therapy in solving a nosocomial COVID-19 infection. Case report and short literature review. Rom. J. Occup. Med. 2021, 72, 47–53. [Google Scholar] [CrossRef]
- Veras, F.P.; Pontelli, M.C.; Silva, C.M.; Toller-Kawahisa, J.E.; de Lima, M.H.; Nascimento, D.C.; Schneider, A.H.; Caetite, D.B.; Rosales, R.; Colón, D.F.; et al. SARS-CoV-2 triggered neutrophil extracellular traps (NETs) mediate COVID-19 pathology. J. Exp. Med. 2020, 217, e20201129. [Google Scholar] [CrossRef]
- Karhadkar, T.R.; Pilling, D.; Gomer, R.H. Serum Amyloid P inhibits single stranded RNA-induced lung inflammation, lung damage, and cytokine storm in mice. PLoS ONE 2021, 16, e0245924. [Google Scholar] [CrossRef]
- Ali, N. Relationship Between COVID-19 Infection and Liver Injury: A Review of Recent Data. Front. Med. 2020, 7, 458. [Google Scholar] [CrossRef]
- Buxton, D.B. Current status of nanotechnology approaches for cardiovascular disease: A personal perspective. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2009, 1, 149–155. [Google Scholar] [CrossRef]
- Aswathy, S.U.; Jarin, T.; Mathews, R.; Nair, L.M.; Rroan, M. CAD Systems for Automatic Detection and Classification of Covid-19 in Nano CT Lung Image by Using Machine Learning Technique. Int. J. Pharm. Res. 2020, 2, 1865–1870. [Google Scholar]
- Rana, M. Polymer-based nano-therapies to combat COVID-19 related respiratory injury: Progress, prospects, and challenges. J. Biomater. Sci. Polym. Ed. 2021, 32, 1219–1249. [Google Scholar] [CrossRef]
- Abduljauwad, S.N.; Habib, T.; Ahmed, H.-U.-R. Nano-clays as Potential Pseudo-antibodies for COVID-19. Nanoscale Res. Lett. 2020, 15, 173. [Google Scholar] [CrossRef] [PubMed]
- Carlon, M.S.; Engels, A.C.; Bosch, B.; Joyeux, L.; da Cunha, M.G.M.C.M.; Vidović, D.; Debyser, Z.; De Boeck, K.; Neyrinck, A.; Deprest, J.A. A novel translational model for fetoscopic intratracheal delivery of nanoparticles in piglets. Prenat. Diagn. 2016, 36, 926–934. [Google Scholar] [CrossRef] [Green Version]
- Chang, X.; Xing, L.; Wang, Y.; Yang, C.-X.; He, Y.-J.; Zhou, T.-J.; Gao, X.-D.; Li, L.; Hao, H.-P.; Jiang, H.-L. Monocyte-derived multipotent cell delivered programmed therapeutics to reverse idiopathic pulmonary fibrosis. Sci. Adv. 2020, 6, eaba3167. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Jin, G.; Ji, C.; He, R.; Lin, M.; Zhao, X.; Li, A.; Lu, T.J.; Xu, F. Non-invasive tracking of hydrogel degradation using upconversion nanoparticles. Acta Biomater. 2017, 55, 410–419. [Google Scholar] [CrossRef]
- Healy, D.; Nash, M.E.; Gorelov, A.; Thompson, K.; Dockery, P.; Beloshapkin, S.; Rochev, Y. Fabrication and Application of Photocrosslinked, Nanometer-Scale, Physically Adsorbed Films for Tissue Culture Regeneration. Macromol. Biosci. 2016, 17, 1600175. [Google Scholar] [CrossRef]
- Chen, M.; Rosenberg, J.; Cai, X.; Hsuan Lee, A.C.; Shi, J.; Nguyen, M.; Wignakumar, T.; Mirle, V.; Edobor, A.J.; Fung, J.; et al. Nanotraps for the containment and clearance of SARS-CoV-2. Matter 2021, 4, 2059–2082. [Google Scholar] [CrossRef]
- Doroudian, M.; O’ Neill, A.; Mac Loughlin, R.; Prina-Mello, A.; Volkov, Y.; Donnelly, S.C. Nanotechnology in pulmonary medicine. Curr. Opin. Pharmacol. 2021, 56, 85–92. [Google Scholar] [CrossRef]
- Erasmus, J.H.; Khandhar, A.P.; O’Connor, M.A.; Walls, A.C.; Hemann, E.A.; Murapa, P.; Archer, J.; Leventhal, S.S.; Fuller, J.T.; Lewis, T.B.; et al. An alphavirus-derived replicon RNA vaccine induces SARS-CoV-2 neutralizing antibody and T cell responses in mice and nonhuman primates. Sci. Transl. Med. 2020, 12, eabc9396. [Google Scholar] [CrossRef]
- Hamdi, M.; Abdel-Bar, H.M.; Elmowafy, E.; El-Khouly, A.; Mansour, M.; Awad, G.A. Investigating the Internalization and COVID-19 Antiviral Computational Analysis of Optimized Nanoscale Zinc Oxide. ACS Omega 2021, 6, 6848–6860. [Google Scholar] [CrossRef]
- Ichimura, T.; Mori, Y.; Aschauer, P.; Padmanabha Das, K.M.; Padera, R.F.; Weins, A.; Nasr, M.L.; Bonventre, J.V. KIM-1/TIM-1 is a Receptor for SARS-CoV-2 in Lung and Kidney. medRxiv 2020. preprint. [Google Scholar] [CrossRef] [PubMed]
- Kalarikkal, S.P.; Sundaram, G.M. Edible plant-derived exosomal microRNAs: Exploiting a cross-kingdom regulatory mechanism for targeting SARS-CoV-2. Toxicol. Appl. Pharm. 2021, 414, 115425. [Google Scholar] [CrossRef] [PubMed]
- Khater, S.E.; El-khouly, A.; Abdel-Bar, H.M.; Al-mahallawi, A.M.; Ghorab, D.M. Fluoxetine hydrochloride loaded lipid polymer hybrid nanoparticles showed possible efficiency against SARS-CoV-2 infection. Int. J. Pharm. 2021, 607, 121023. [Google Scholar] [CrossRef]
- Kim, J.; Mukherjee, A.; Nelson, D.; Jozic, A.; Sahay, G. Rapid generation of circulating and mucosal decoy ACE2 using mRNA nanotherapeutics for the potential treatment of SARS-CoV-2. bioRxiv 2020. preprint. [Google Scholar] [CrossRef]
- Klimek, L.; Novak, N.; Hamelmann, E.; Werfel, T.; Wagenmann, M.; Taube, C.; Bauer, A.; Merk, H.; Rabe, U.; Jung, K.; et al. Severe allergic reactions after COVID-19 vaccination with the Pfizer/BioNTech vaccine in Great Britain and USA. Allergo J. Int. 2021, 30, 51–55. [Google Scholar] [CrossRef]
- Mitrani, M.I.; Bellio, M.A.; Sagel, A.; Saylor, M.; Kapp, W.; VanOsdol, K.; Haskell, G.; Stewart, D.; Abdullah, Z.; Santos, I.; et al. Case Report: Administration of Amniotic Fluid-Derived Nanoparticles in Three Severely Ill COVID-19 Patients. Front. Med. 2021, 8, 583842. [Google Scholar] [CrossRef] [PubMed]
- Mosselhy, D.; Virtanen, J.; Kant, R.; He, W.; Elbahri, M.; Sironen, T. COVID-19 Pandemic: What about the Safety of Anti-Coronavirus Nanoparticles? Nanomaterials 2021, 11, 796. [Google Scholar] [CrossRef]
- Ndeupen, S.; Qin, Z.; Jacobsen, S.; Estanbouli, H.; Bouteau, A.; Igyártó, B.Z. The mRNA-LNP platform’s lipid nanoparticle component used in preclinical vaccine studies is highly inflammatory. bioRxiv 2021. preprint. [Google Scholar] [CrossRef] [PubMed]
- Ngwa, W.; Kumar, R.; Thompson, D.; Lyerly, W.; Moore, R.; Reid, T.-E.; Lowe, H.; Toyang, N. Potential of Flavonoid-Inspired Phytomedicines against COVID-19. Molecules 2020, 25, 2707. [Google Scholar] [CrossRef]
- Peñate-Medina, T.; Damoah, C.; Benezra, M.; Will, O.; Kairemo, K.; Humbert, J.; Sebens, S. Alpha-MSH Targeted Liposomal Nanoparticle for Imaging in Inflammatory Bowel Disease (IBD). Curr. Pharm. Des. 2020, 26, 3840–3846. [Google Scholar] [CrossRef] [PubMed]
- Rabiei, M.; Kashanian, S.; Samavati, S.S.; Derakhshankhah, H.; Jamasb, S.; McInnes, S.J. Characteristics of SARS-CoV2 that may be useful for nanoparticle pulmonary drug delivery. J. Drug Target. 2021, 1–11, Epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Rao, L.; Xia, S.; Xu, W.; Tian, R.; Yu, G.; Gu, C.; Pan, P.; Meng, Q.-F.; Cai, X.; Qu, D.; et al. Decoy nanoparticles protect against COVID-19 by concurrently adsorbing viruses and inflammatory cytokines. Proc. Natl. Acad. Sci. USA 2020, 117, 27141–27147. [Google Scholar] [CrossRef] [PubMed]
- Seyfoori, A.; Barough, M.S.; Mokarram, P.; Ahmadi, M.; Mehrbod, P.; Sheidary, A.; Madrakian, T.; Kiumarsi, M.; Walsh, T.; McAlinden, K.; et al. Emerging Advances of Nanotechnology in Drug and Vaccine Delivery against Viral Associated Respiratory Infectious Diseases (VARID). Int. J. Mol. Sci. 2021, 22, 6937. [Google Scholar] [CrossRef] [PubMed]
- Shurbaji, S.; El-Sherbiny, I.M.; Alser, M.; Ali, I.H.; Kordi, H.; Al-Sadi, A.; Popelka, A.; Benslimane, F.; Yacoub, M.; Yalcin, H.C. Nitric Oxide Releasing Hydrogel Nanoparticles Decreases Epithelial Cell Injuries Associated With Airway Reopening. Front. Bioeng. Biotechnol. 2021, 8, 579788. [Google Scholar] [CrossRef]
- Si, X.A.; Talaat, M.; Su, W.-C.; Xi, J. Inhalation dosimetry of nasally inhaled respiratory aerosols in the human respiratory tract with locally remodeled conducting lungs. Inhal. Toxicol. 2021, 33, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Tahmasebi, S.; El-Esawi, M.A.; Mahmoud, Z.H.; Timoshin, A.; Valizadeh, H.; Roshangar, L.; Varshoch, M.; Vaez, A.; Aslani, S.; Navashenaq, J.G.; et al. Immunomodulatory effects of nanocurcumin on Th17 cell responses in mild and severe COVID-19 patients. J. Cell. Physiol. 2021, 236, 5325–5338. [Google Scholar] [CrossRef]
- Abo-Zeid, Y.; Ismail, N.S.; McLean, G.R.; Hamdy, N.M. A molecular docking study repurposes FDA approved iron oxide nanoparticles to treat and control COVID-19 infection. Eur. J. Pharm. Sci. 2020, 153, 105465. [Google Scholar] [CrossRef]
- Abo-Zeid, Y.; Williams, G.R.; Touabi, L.; McLean, G.R. An investigation of rhinovirus infection on cellular uptake of poly (glycerol-adipate) nanoparticles. Int. J. Pharm. 2020, 589, 119826. [Google Scholar] [CrossRef]
- Molinaro, R.; Pasto, A.; Taraballi, F.; Giordano, F.; Azzi, J.A.; Tasciotti, E.; Corbo, C. Biomimetic Nanoparticles Potentiate the Anti-Inflammatory Properties of Dexamethasone and Reduce the Cytokine Storm Syndrome: An Additional Weapon against COVID-19? Nanomaterials 2020, 10, 2301. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Hou, X.; Yan, J.; Zhang, C.; Li, W.; Zhao, W.; Du, S.; Dong, Y. Leveraging mRNA Sequences and Nanoparticles to Deliver SARS-CoV-2 Antigens In Vivo. Adv. Mater. 2020, 32, e2004452. [Google Scholar] [CrossRef] [PubMed]
- Arefi, S.M.A.; Yang, C.W.T.; Sin, D.D.; Feng, J.J. Simulation of nanoparticle transport and adsorption in a microfluidic lung-on-a-chip device. Biomicrofluidics 2020, 14, 044117. [Google Scholar] [CrossRef]
- Nasajpour, A.; Ansari, S.; Rinoldi, C.; Rad, A.S.; Aghaloo, T.; Shin, S.R.; Mishra, Y.K.; Adelung, R.; Święszkowski, W.; Annabi, N.; et al. A Multifunctional Biopolymeric Periodontal Membrane with Osteogenic and Antibacterial Characteristics. Adv. Funct. Mater. 2018, 28, 1703437. [Google Scholar] [CrossRef]
- Mufamadi, M.S. Nanotechnology shows promise for next-generation vaccines in the fight against COVID-19. MRS Bull. 2020, 45, 981–982. [Google Scholar] [CrossRef]
- Xu, Y.; Xiang, J.; Zhao, H.; Liang, H.; Huang, J.; Li, Y.; Pan, J.; Zhou, H.; Zhang, X.; Wang, J.H.; et al. Human amniotic fluid stem cells labeled with up-conversion nanoparticles for imaging-monitored repairing of acute lung injury. Biomaterials 2016, 100, 91–100. [Google Scholar] [CrossRef]
- Pang, S.; Gao, Y.; Wang, F.; Wang, Y.; Cao, M.; Zhang, W.; Liang, Y.; Song, M.; Jiang, G. Toxicity of silver nanoparticles on wound healing: A case study of zebrafish fin regeneration model. Sci. Total Environ. 2020, 717, 137178. [Google Scholar] [CrossRef]
- Mârza, S.M.; Magyari, K.; Bogdan, S.; Moldovan, M.L.; Peştean, C.; Nagy, A.; Tăbăran, F.; Licarete, E.; Suarasan, S.; Dreanca, A.; et al. Skin wound regeneration with bioactive glass-gold nanoparticles ointment. Biomed. Mater. 2019, 14, 025011. [Google Scholar] [CrossRef]
- Nho, R. Pathological effects of nano-sized particles on the respiratory system. Nanomed. Nanotechnol. Biol. Med. 2020, 29, 102242. [Google Scholar] [CrossRef]
- Hanot-Roy, M.; Tubeuf, E.; Guilbert, A.; Bado-Nilles, A.; Vigneron, P.; Trouiller, B.; Braun, A.; Lacroix, G. Oxidative stress pathways involved in cytotoxicity and genotoxicity of titanium dioxide (TiO2) nanoparticles on cells constitutive of alveolo-capillary barrier in vitro. Toxicol. In Vitro 2016, 33, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Veith, L.; Vennemann, A.; Breitenstein, D.; Engelhard, C.; Wiemann, M.; Hagenhoff, B. Detection of SiO2 nanoparticles in lung tissue by ToF-SIMS imaging and fluorescence microscopy. Analyst 2017, 142, 2631–2639. [Google Scholar] [CrossRef]
- Yamamoto, K.; Yoshida, T.; Hayashida, T.; Izumi, H.; Morimoto, Y. Comparison of Rat Lung Tissue Following Inhalation and Intratracheal Administration of Metal Oxide Nanoparticles. Microsc. Microanal. 2016, 22, 1064–1065. [Google Scholar] [CrossRef] [Green Version]
- Lierová, A.; Kašparová, J.; Pejchal, J.; Kubelkova, K.; Jeličová, M.; Palarčík, J.; Korecká, L.; Bílková, Z.; Šinkorová, Z. Attenuation of Radiation-Induced Lung Injury by Hyaluronic Acid Nanoparticles. Front. Pharmacol. 2020, 11, 1199. [Google Scholar] [CrossRef] [PubMed]
- Chrzanowski, W.; Kim, S.; McClements, L. Can Stem Cells Beat COVID-19: Advancing Stem Cells and Extracellular Vesicles Toward Mainstream Medicine for Lung Injuries Associated With SARS-CoV-2 Infections. Front. Bioeng. Biotechnol. 2020, 8, 554. [Google Scholar] [CrossRef]
- Taghavi-Farahabadi, M.; Mahmoudi, M.; Soudi, S.; Hashemi, S.M. Hypothesis for the management and treatment of the COVID-19-induced acute respiratory distress syndrome and lung injury using mesenchymal stem cell-derived exosomes. Med. Hypotheses 2020, 144, 109865. [Google Scholar] [CrossRef]
- Atluri, S.; Manocha, V.; Boddu, N.; Bhati, S.; Syed, Z.A.; Diwan, S. Safety and Effectiveness of Intravascular Mesenchymal Stem Cells to Treat Organ Failure and Possible Application in COVID-19 Complications. Pain Physician 2020, 23, S391–S420. [Google Scholar] [CrossRef] [PubMed]
- Zayed, M.; Iohara, K. Immunomodulation and Regeneration Properties of Dental Pulp Stem Cells: A Potential Therapy to Treat Coronavirus Disease 2019. Cell Transplant. 2020, 29, 963689720952089. [Google Scholar] [CrossRef]
- Desai, D.; Shende, P. Nanoconjugates-Based Stem Cell Therapy for the Management of COVID-19. Stem Cell Rev. Rep. 2021, 17, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Al-Khawaga, S.; Abdelalim, E.M. Potential application of mesenchymal stem cells and their exosomes in lung injury: An emerging therapeutic option for COVID-19 patients. Stem Cell Res. Ther. 2020, 11, 437. [Google Scholar] [CrossRef]
- Fang, Y.; Liu, H.; Huang, H.; Li, H.; Saqi, A.; Qiang, L.; Que, J. Distinct stem/progenitor cells proliferate to regenerate the trachea, intrapulmonary airways and alveoli in COVID-19 patients. Cell Res. 2020, 30, 705–707. [Google Scholar] [CrossRef]
- Rogers, C.J.; Harman, R.J.; Bunnell, B.A.; Schreiber, M.A.; Xiang, C.; Wang, F.-S.; Santidrian, A.F.; Minev, B.R. Rationale for the clinical use of adipose-derived mesenchymal stem cells for COVID-19 patients. J. Transl. Med. 2020, 18, 203. [Google Scholar] [CrossRef]
- Liu, S.; Peng, D.; Qiu, H.; Yang, K.; Fu, Z.; Zou, L. Mesenchymal stem cells as a potential therapy for COVID-19. Stem Cell Res. Ther. 2020, 11, 169. [Google Scholar] [CrossRef] [PubMed]
- Gentile, P.; Sterodimas, A. Adipose Stem Cells (ASCs) and Stromal Vascular Fraction (SVF) as a Potential Therapy in Combating (COVID-19)-Disease. Aging Dis. 2020, 11, 465–469. [Google Scholar] [CrossRef]
- Klimczak, A. Perspectives on mesenchymal stem/progenitor cells and their derivates as potential therapies for lung damage caused by COVID-19. World J. Stem Cells 2020, 12, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Kashte, S.; Gupta, M.; Rodriguez, H.; Gautam, S.S.; Kadam, S. Mesenchymal stem cells and exosome therapy for COVID-19: Current status and future perspective. Hum. Cell 2020, 33, 907–918. [Google Scholar] [CrossRef]
- Can, A.; Coskun, H. The rationale of using mesenchymal stem cells in patients with COVID-19-related acute respiratory distress syndrome: What to expect. Stem Cells Transl. Med. 2020, 9, 1287–1302. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Jiang, Y.; Zhu, M.; Chen, L.; Zhou, X.; Zhou, C.; Ye, P.; Chen, X.; Wang, B.; Xu, Z.; et al. Clinical study using mesenchymal stem cells for the treatment of patients with severe COVID-19. Front. Med. 2020, 14, 664–673. [Google Scholar] [CrossRef]
- Xiao, K.; Hou, F.; Huang, X.; Li, B.; Qian, Z.R.; Xie, L. Mesenchymal stem cells: Current clinical progress in ARDS and COVID-19. Stem Cell Res. Ther. 2020, 11, 305. [Google Scholar] [CrossRef]
- Yen, B.L.; Yen, M.; Wang, L.; Liu, K.; Sytwu, H. Current status of mesenchymal stem cell therapy for immune/inflammatory lung disorders: Gleaning insights for possible use in COVID-19. Stem Cells Transl. Med. 2020, 9, 1163–1173. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, C.; Zhang, C.; Li, Q.; Chen, J.; Cheng, L.; Zhou, J.; Su, X.; Song, Y. Vagal-α7nAChR signaling promotes lung stem cells regeneration via fibroblast growth factor 10 during lung injury repair. Stem Cell Res. Ther. 2020, 11, 230. [Google Scholar] [CrossRef]
- Duan, F.; Guo, L.; Yang, L.; Han, Y.; Thakur, A.; Nilsson-Payant, B.E.; Wang, P.; Zhang, Z.; Ma, C.Y.; Zhou, X.; et al. Modeling COVID-19 with Human Pluripotent Stem Cell-Derived Cells Reveals Synergistic Effects of Anti-inflammatory Macrophages with ACE2 Inhibition Against SARS-CoV-2. Res. Sq. 2020. preprint. [Google Scholar] [CrossRef]
- Parekh, K.R.; Nawroth, J.; Pai, A.; Busch, S.M.; Senger, C.N.; Ryan, A.L. Stem Cells and Lung Regeneration. Am. J. Physiol. Physiol. 2020, 319, C675–C693. [Google Scholar] [CrossRef]
- Alzahrani, F.A.; Saadeldin, I.M.; Ahmad, A.; Kumar, D.; Azhar, E.I.; Siddiqui, A.J.; Kurdi, B.; Sajini, A.; Alrefaei, A.F.; Jahan, S. The Potential Use of Mesenchymal Stem Cells and Their Derived Exosomes as Immunomodulatory Agents for COVID-19 Patients. Stem Cells Int. 2020, 2020, 8835986. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Tang, M.; Liu, Q.; Han, X.; Jin, H.; Zhu, H.; Li, Y.; He, L.; Ji, H.; Zhou, B. Bi-directional differentiation of single bronchioalveolar stem cells during lung repair. Cell Discov. 2020, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Tindle, C.; Fuller, M.; Fonseca, A.; Taheri, S.; Ibeawuchi, S.-R.; Beutler, N.; Katkar, G.D.; Claire, A.; Castillo, V.; Hernandez, M.; et al. Adult stem cell-derived complete lung organoid models emulate lung disease in COVID-19. eLife 2021, 10, e66417. [Google Scholar] [CrossRef]
- Wu, H.; Yu, Y.; Huang, H.; Hu, Y.; Fu, S.; Wang, Z.; Shi, M.; Zhao, X.; Yuan, J.; Li, J.; et al. Progressive Pulmonary Fibrosis Is Caused by Elevated Mechanical Tension on Alveolar Stem Cells. Cell 2020, 180, 107–121.e17. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.; Volckaert, T.; Redente, E.F.; Hopkins, S.; Klinkhammer, K.; Wasnick, R.; Chao, C.-M.; Yuan, J.; Zhang, J.-S.; Yao, C.; et al. FGF10-FGFR2B Signaling Generates Basal Cells and Drives Alveolar Epithelial Regeneration by Bronchial Epithelial Stem Cells after Lung Injury. Stem Cell Rep. 2019, 12, 1041–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, V.; Thakur, S.; Rao, J.; Arakeri, G.; Brennan, P.A.; Jadhav, S.; Sayeed, M.S.; Rao, G. Mesenchymal stem cells-bridge catalyst between innate and adaptive immunity in COVID 19. Med. Hypotheses 2020, 143, 109845. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Hume, A.J.; Abo, K.M.; Werder, R.B.; Villacorta-Martin, C.; Alysandratos, K.; Beermann, M.L.; Simone-Roach, C.; Lindstrom-Vautrin, J.; Olejnik, J.; et al. SARS-CoV-2 Infection of Pluripotent Stem Cell-derived Human Lung Alveolar Type 2 Cells Elicits a Rapid Epithelial-Intrinsic Inflammatory Response. Cell Stem Cell 2020, 27, 962–973.e7. [Google Scholar] [CrossRef]
- Shi, L.; Huang, H.; Lu, X.; Yan, X.; Jiang, X.; Xu, R.; Wang, S.; Zhang, C.; Yuan, X.; Xu, Z.; et al. Effect of human umbilical cord-derived mesenchymal stem cells on lung damage in severe COVID-19 patients: A randomized, double-blind, placebo-controlled phase 2 trial. Signal Transduct. Target. Ther. 2021, 6, 58. [Google Scholar] [CrossRef]
- Askenase, P.W. COVID-19 therapy with mesenchymal stromal cells (MSC) and convalescent plasma must consider exosome involvement: Do the exosomes in convalescent plasma antagonize the weak immune antibodies? J. Extracell. Vesicles 2020, 10, e12004. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liu, K.; Cui, G.; Huang, X.; Yao, S.; Guo, W.; Qin, Z.; Li, Y.; Yang, R.; Pu, W.; et al. Lung regeneration by multipotent stem cells residing at the bronchioalveolar-duct junction. Nat. Genet. 2019, 51, 728–738. [Google Scholar] [CrossRef]
- Miller, A.J.; Dye, B.R.; Ferrer-Torres, D.; Hill, D.R.; Overeem, A.W.; Shea, L.D.; Spence, J.R. Generation of lung organoids from human pluripotent stem cells in vitro. Nat. Protoc. 2019, 14, 518–540. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Chang, W.; Meng, S.; Xu, X.; Xie, J.; Guo, F.; Yang, Y.; Qiu, H.; Liu, L. Mesenchymal stem cells induce dendritic cell immune tolerance via paracrine hepatocyte growth factor to alleviate acute lung injury. Stem Cell Res. Ther. 2019, 10, 372. [Google Scholar] [CrossRef] [Green Version]
- McQualter, J.L. Endogenous lung stem cells for lung regeneration. Expert Opin. Biol. Ther. 2019, 19, 539–546. [Google Scholar] [CrossRef]
- Lee, J.-H.; Rawlins, E.L. Developmental mechanisms and adult stem cells for therapeutic lung regeneration. Dev. Biol. 2018, 433, 166–176. [Google Scholar] [CrossRef]
- Nikolić, M.Z.; Johnson, J.-A.; Sun, D.; Caritg, O.; Laresgoiti, U.; Brady, J.; Allen, G.; Giangreco, A.; Rawlins, E.L. Development of a genetically modifiable epithelial in-vitro culture system from human embryonic lung epithelial stem cells: Towards human lung regeneration in end-stage respiratory failure. Lancet 2017, 389, S74. [Google Scholar] [CrossRef]
- El-Badrawy, M.K.; Shalabi, N.M.; Mohamed, M.A.; Ragab, A.; Abdelwahab, H.W. Stem Cells and Lung Regeneration. Int. J. Stem Cells 2016, 9, 31–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kathiriya, J.J.; Brumwell, A.N.; Jackson, J.R.; Tang, X.; Chapman, H.A. Distinct Airway Epithelial Stem Cells Hide among Club Cells but Mobilize to Promote Alveolar Regeneration. Cell Stem Cell 2020, 26, 346–358.e4. [Google Scholar] [CrossRef]
- Beers, M.F.; Moodley, Y. When Is an Alveolar Type 2 Cell an Alveolar Type 2 Cell? A Conundrum for Lung Stem Cell Biology and Regenerative Medicine. Am. J. Respir. Cell Mol. Biol. 2017, 57, 18–27. [Google Scholar] [CrossRef]
- Wang, J.; Huang, R.; Xu, Q.; Zheng, G.; Qiu, G.; Ge, M.; Shu, Q.; Xu, J. Mesenchymal Stem Cell–Derived Extracellular Vesicles Alleviate Acute Lung Injury Via Transfer of miR-27a-3p. Crit. Care Med. 2020, 48, e599–e610. [Google Scholar] [CrossRef] [PubMed]
- Azimi, B.; Sorayani Bafqi, M.S.; Fusco, A.; Ricci, C.; Gallone, G.C.; Bagherzadeh, R.; Donnarumma, G.; Uddin, M.J.; Latifi, M.; Lazzeri, A.; et al. Electrospun ZnO/poly(vinylidene fluoride-trifluoroethylene) scaffolds for lung tissue engineering. Tissue Eng. Part A 2020, 26, 1312–1331. [Google Scholar] [CrossRef]
- Rezaei, F.S.; Khorshidian, A.; Beram, F.M.; Derakhshani, A.; Esmaeili, J.; Barati, A. 3D printed chitosan/polycaprolactone scaffold for lung tissue engineering: Hope to be useful for COVID-19 studies. RSC Adv. 2021, 11, 19508–19520. [Google Scholar] [CrossRef]
- Shulimzon, T.R.; Giladi, S.; Zilberman, M. Catheter Injectable Hydrogel-Based Scaffolds for Tissue Engineering Applications in lung disease. Isr. Med Assoc. J. IMAJ 2020, 22, 736–740. [Google Scholar]
- Bölükbas, D.A.; De Santis, M.M.; Alsafadi, H.N.; Doryab, A.; Wagner, D.E. The Preparation of Decellularized Mouse Lung Matrix Scaffolds for Analysis of Lung Regenerative Cell Potential. In Methods in Molecular Biology; Humana Press: New York, NY, USA, 2019; pp. 275–295. [Google Scholar] [CrossRef]
- Gilpin, S.E.; Wagner, D.E. Acellular human lung scaffolds to model lung disease and tissue regeneration. Eur. Respir. Rev. 2018, 27, 180021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilodeau, C.; Shojaie, S.; Goltsis, O.; Wang, J.; Luo, D.; Ackerley, C.; Rogers, I.M.; Cox, B.; Post, M. TP63 basal cells are indispensable during endoderm differentiation into proximal airway cells on acellular lung scaffolds. NPJ Regen. Med. 2021, 6, 12. [Google Scholar] [CrossRef]
- Piccinini, E.; Allegretto, J.A.; Scotto, J.; Cantillo, A.L.; Fenoy, G.E.; Marmisollé, W.A.; Azzaroni, O. Surface Engineering of Graphene through Heterobifunctional Supramolecular-Covalent Scaffolds for Rapid COVID-19 Biomarker Detection. ACS Appl. Mater. Interfaces 2021, 13, 43696–43707. [Google Scholar] [CrossRef] [PubMed]
- Kchaou, M.; Alquraish, M.; Abuhasel, K.; Abdullah, A.; Ali, A. Electrospun Nanofibrous Scaffolds: Review of Current Progress in the Properties and Manufacturing Process, and Possible Applications for COVID-19. Polymers 2021, 13, 916. [Google Scholar] [CrossRef]
- Ohata, K.; Ott, H.C. Human-scale lung regeneration based on decellularized matrix scaffolds as a biologic platform. Surg. Today 2020, 50, 633–643. [Google Scholar] [CrossRef]
- Zuo, H.; Cattani-Cavalieri, I.; Valença, S.S.; Musheshe, N.; Schmidt, M. Function of cAMP scaffolds in obstructive lung disease: Focus on epithelial-to-mesenchymal transition and oxidative stress. Br. J. Pharmacol. 2019, 176, 2402–2415. [Google Scholar] [CrossRef] [Green Version]
- Tisler, M.; Alkmin, S.; Chang, H.-Y.; Leet, J.; Bernau, K.; Sandbo, N.; Campagnola, P.J. Analysis of fibroblast migration dynamics in idiopathic pulmonary fibrosis using image-based scaffolds of the lung extracellular matrix. Am. J. Physiol. Cell. Mol. Physiol. 2020, 318, L276–L286. [Google Scholar] [CrossRef] [PubMed]
- Samadi, S.; Moradkhani, M.; Beheshti, H.; Irani, M.; Aliabadi, M. Fabrication of chitosan/poly(lactic acid)/graphene oxide/TiO2 composite nanofibrous scaffolds for sustained delivery of doxorubicin and treatment of lung cancer. Int. J. Biol. Macromol. 2018, 110, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Scarritt, M.E.; Pashos, N.C.; Motherwell, J.M.; Eagle, Z.; Burkett, B.J.; Gregory, A.N.; Mostany, R.; Weiss, D.J.; Alvarez, D.F.; Bunnell, B.A. Re-endothelialization of rat lung scaffolds through passive, gravity-driven seeding of segment-specific pulmonary endothelial cells. J. Tissue Eng. Regen. Med. 2018, 12, e786–e806. [Google Scholar] [CrossRef] [PubMed]
- Stahl, E.C.; Bonvillain, R.W.; Skillen, C.D.; Burger, B.L.; Hara, H.; Lee, W.; Trygg, C.B.; Didier, P.J.; Grasperge, B.F.; Pashos, N.; et al. Evaluation of the host immune response to decellularized lung scaffolds derived from α-Gal knockout pigs in a non-human primate model. Biomaterials 2018, 187, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Ju, Z.; Yu, J.; Qiao, Y.; Hou, C.; Wang, C.; Hei, F. Decellularized Rat Lung Scaffolds Using Sodium Lauryl Ether Sulfate for Tissue Engineering. ASAIO J. 2018, 64, 406–414. [Google Scholar] [CrossRef]
- Zvarova, B.; Uhl, F.E.; Uriarte, J.J.; Borg, Z.D.; Coffey, A.L.; Bonenfant, N.R.; Weiss, D.J.; Wagner, D.E. Residual Detergent Detection Method for Nondestructive Cytocompatibility Evaluation of Decellularized Whole Lung Scaffolds. Tissue Eng. Part C Methods 2016, 22, 418–428. [Google Scholar] [CrossRef] [Green Version]
- Binzel, D.W.; Li, X.; Burns, N.; Khan, E.; Lee, W.; Chen, L.; Ellipilli, S.; Miles, W.; Ho, Y.; Guo, P. Thermostability, Tunability, and Tenacity of RNA as Rubbery Anionic Biopolymeric Materials in Nanotechnology and Nanomedicine-Specific Cancer Targeting with Undetectable Toxicity. Chem. Rev. 2021, 121, 7398–7467. [Google Scholar] [CrossRef]
- Godin, L.M.; Sandri, B.; Wagner, D.; Meyer, C.M.; Price, A.P.; Akinnola, I.; Weiss, D.J.; Panoskaltsis-Mortari, A. Decreased Laminin Expression by Human Lung Epithelial Cells and Fibroblasts Cultured in Acellular Lung Scaffolds from Aged Mice. PLoS ONE 2016, 11, e0150966. [Google Scholar] [CrossRef] [Green Version]
- Le, A.V.; Hatachi, G.; Beloiartsev, A.; Ghaedi, M.; Engler, A.J.; Baevova, P.; Niklason, L.E.; Calle, E.A. Efficient and Functional Endothelial Repopulation of Whole Lung Organ Scaffolds. ACS Biomater. Sci. Eng. 2017, 3, 2000–2010. [Google Scholar] [CrossRef]
- Balestrini, J.L.; Gard, A.L.; Liu, A.; Leiby, K.L.; Schwan, J.; Kunkemoeller, B.; Calle, E.A.; Sivarapatna, A.; Lin, T.; Dimitrievska, S.; et al. Production of decellularized porcine lung scaffolds for use in tissue engineering. Integr. Biol. 2015, 7, 1598–1610. [Google Scholar] [CrossRef] [Green Version]
- Pothen, J.J.; Rajendran, V.; Wagner, D.; Weiss, D.J.; Smith, B.J.; Ma, B.; Bates, J.H. A Computational Model of Cellular Engraftment on Lung Scaffolds. Biores. Open Access 2016, 5, 308–319. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.-J.; Hsu, C.-H.; Ling, T.-Y.; Liu, L.; Lin, T.-C.; Jakfar, S.; Young, I.-C.; Lin, F.-H. The preparation of cell-containing microbubble scaffolds to mimic alveoli structure as a 3D drug-screening system for lung cancer. Biofabrication 2020, 12, 025031. [Google Scholar] [CrossRef] [PubMed]
- Dhamecha, D.; Le, D.Q.; Movsas, R.; Gonsalves, A.; Menon, J.U. Porous Biopolymeric Microspheres With Controllable Pore Diameters for Tissue Engineered Lung Tumor Model Development. Front. Bioeng. Biotechnol. 2020, 8, 799. [Google Scholar] [CrossRef] [PubMed]
- Chiarelli, L.R.; Degiacomi, G.; Egorova, A.; Makarov, V.; Pasca, M.R. Nitric oxide-releasing compounds for the treatment of lung infections. Drug Discov. Today 2021, 26, 542–550. [Google Scholar] [CrossRef]
- Abrami, M.; Chiarappa, G.; Farra, R.; Grassi, G.; Marizza, P.; Grassi, M. Use of low-field NMR for the characterization of gels and biological tissues. ADMET DMPK 2018, 6, 34–46. [Google Scholar] [CrossRef] [Green Version]
- Filippi, M.; Born, G.; Chaaban, M.; Scherberich, A. Natural Biopolymeric Scaffolds in Bone Regeneration. Front. Bioeng. Biotechnol. 2020, 8, 474. [Google Scholar] [CrossRef]
- Patel, M.; Min, J.H.; Hong, M.; Lee, H.J.; Kang, S.; Yi, S.; Koh, W. Culture of neural stem cells on the conductive and microgrooved biopolymeric scaffolds fabricated via electrospun fiber-template lithography (EFTL). Biomed. Mater. 2020, 15, 045007. [Google Scholar] [CrossRef]
- Meng, Z.; He, J.; Li, J.; Su, Y.; Li, D. Melt-based, solvent-free additive manufacturing of biodegradable biopolymeric scaffolds with designer microstructures for tailored mechanical/biological properties and clinical applications. Virtual Phys. Prototyp. 2020, 15, 417–444. [Google Scholar] [CrossRef]
- Chin, A.R.; Gao, J.; Wang, Y.; Taboas, J.M.; Almarza, A.J. Regenerative Potential of Various Soft Biopolymeric Scaffolds in the Temporomandibular Joint Condyle. J. Oral Maxillofac. Surg. 2018, 76, 2019–2026. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nanoparticle Types | Methods/Roles | Ref. |
|---|---|---|
| PCL | Electrospinning of PCL and ZnO particles creates the membranes | [54] |
| LNPs | employed to carry plasmid DNA into the nucleus of host cells in lung tissues | [55] |
| PLGA | used to create porous substrates for complex cell cultures | [124] |
| TT3 | nanoparticles enhance the expression of putative SARS-CoV-2 antigens | [52] |
| MC3 | elicits almost two orders of magnitude more antigen-specific antibodies | [52] |
| UCNPs | deterioration of the hydrogels was assessed utilizing a simple and non-invasive in vivo hydrogel tracking methodology | [27] |
| LPH | boost the medicine’s efficacy | [35] |
| PLA | PLA/chitosan ratio, TiO2/DOX content, and GO/TiO2/DOX content influence DOX release from nanofibrous scaffolds | [113] |
| TW-37 | TW-37, a novel KIM-1-mediated endocytosis inhibitor, and anti-Kim-1 antibodies both reduced absorptions | [33] |
| Nanocurcumin | therapeutic effects on the quantity and reactivity of Th17 cells in COVID-19 patients | [48] |
| Nanoparticle Types | Methods/Roles | Ref. |
|---|---|---|
| IONPs | Interaction of magnetic IONPs (Fe2O3 and Fe3O4) with the SARS-CoV-2 spike protein | [49] |
| PER NPs | MOMCs have many PER NPs on their surface that target the lungs | [26] |
| LIONs | The LION/repRNA-CoV2S vaccination elicited anti-SARS-CoV-2 S protein IgG antibody isotypes suggestive of a type 1 T helper cell response | [31] |
| ZnO | interaction of ZnO NPs with COVID-19 targets such as the ACE2 receptor | [32] |
| AgNP | are often used to treat acute and chronic wounds | [57] |
| AuNPs | potential to accelerate wound healing by stimulating tissue regeneration, connective tissue development, and angiogenesis | [58] |
| anti-CoV NPs | addresses nano–bio interactions, in vitro (lung cells), and in vivo assessments, as well as human ramifications | [39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbas, M.; Alqahtani, M.S.; Almohiy, H.M.; Alqahtani, F.F.; Alhifzi, R.; Jambi, L.K. The Potential Contribution of Biopolymeric Particles in Lung Tissue Regeneration of COVID-19 Patients. Polymers 2021, 13, 4011. https://doi.org/10.3390/polym13224011
Abbas M, Alqahtani MS, Almohiy HM, Alqahtani FF, Alhifzi R, Jambi LK. The Potential Contribution of Biopolymeric Particles in Lung Tissue Regeneration of COVID-19 Patients. Polymers. 2021; 13(22):4011. https://doi.org/10.3390/polym13224011
Chicago/Turabian StyleAbbas, Mohamed, Mohammed S. Alqahtani, Hussain M. Almohiy, Fawaz F. Alqahtani, Roaa Alhifzi, and Layal K. Jambi. 2021. "The Potential Contribution of Biopolymeric Particles in Lung Tissue Regeneration of COVID-19 Patients" Polymers 13, no. 22: 4011. https://doi.org/10.3390/polym13224011