
Production and Properties of Microbial Polyhydroxyalkanoates Synthesized from Hydrolysates of Jerusalem Artichoke Tubers and Vegetative Biomass


 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
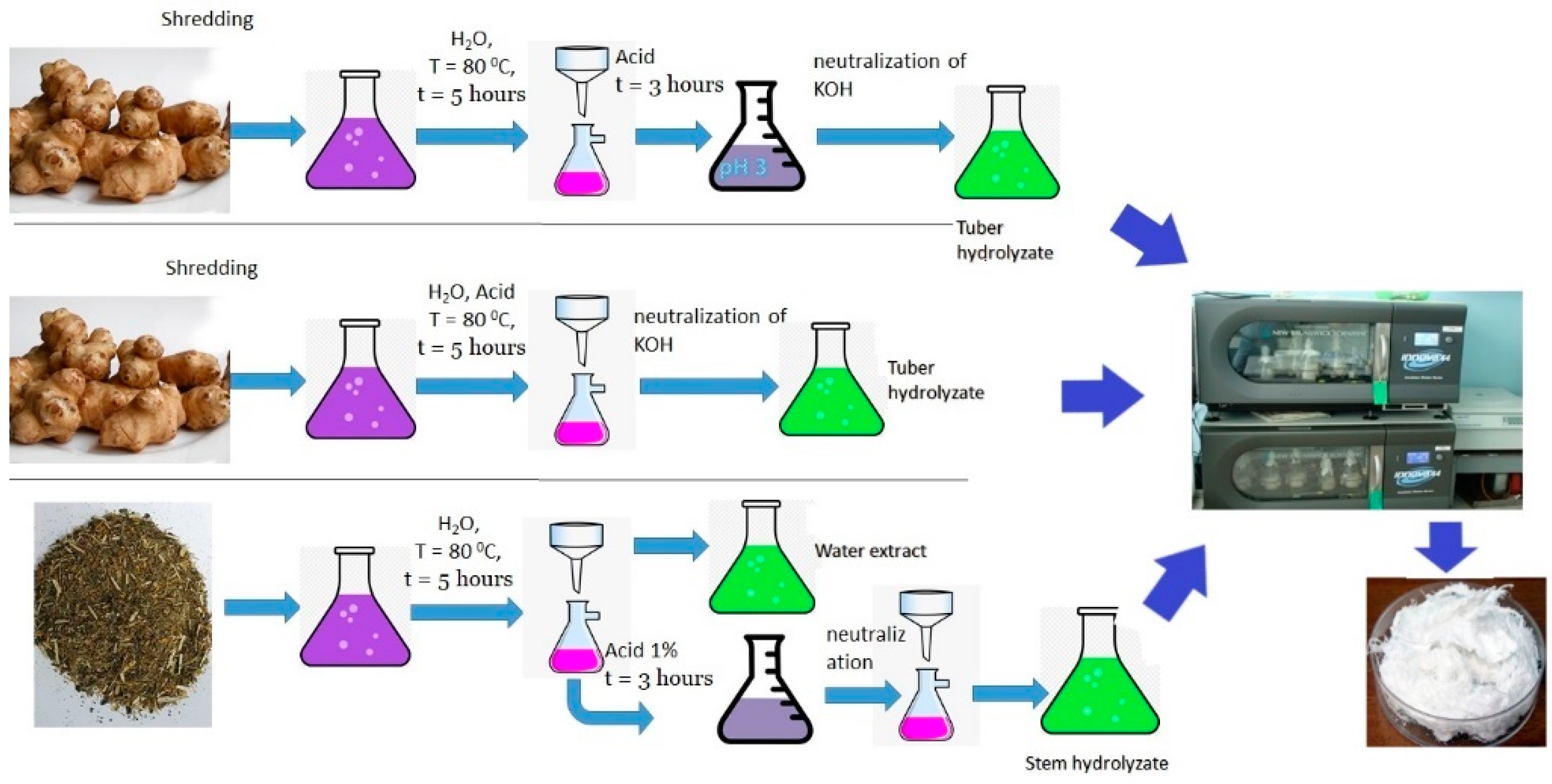
2.2. Production of Sugar-Containing Substrates from Jerusalem Artichoke
2.3. Composition of Jerusalem Artichoke Hydrolysates
2.4. Media and Growth Conditions of Bacterial Strains
2.5. Monitoring Process Parameters
2.6. PHA Recovery from Cell Biomass
2.7. PHA Chemical Composition
2.8. Physicochemical Properties of PHAs
2.9. Production and Investigation of Polymer Films
2.10. Statistics
3. Results and Discussion
3.1. Characterization of Hydrolysates of the JA tubers and Vegetative Biomass
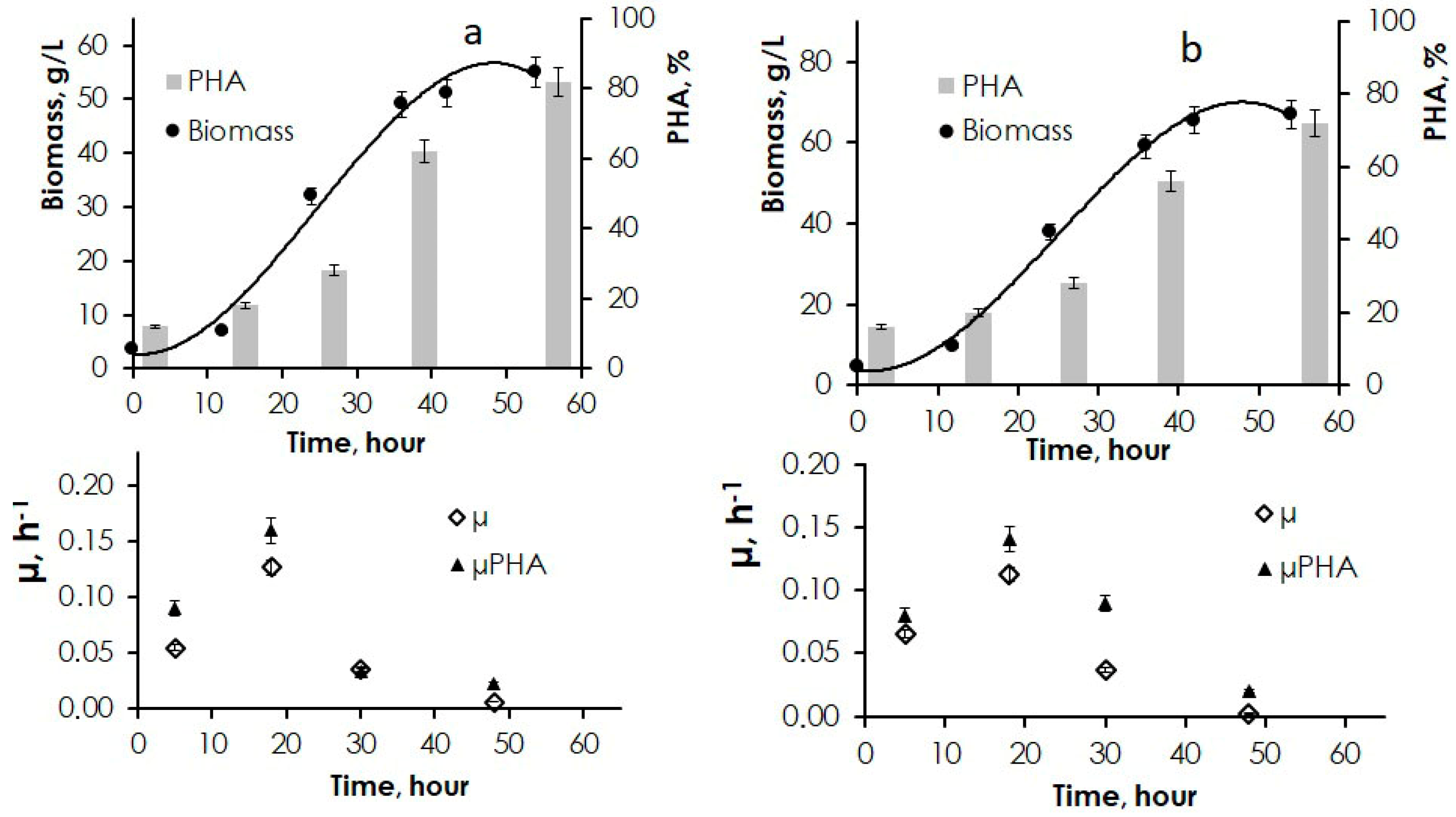
3.2. Production Parameters of PHA Synthesis from Jerusalem Artichoke Hydrolysates
3.3. Structure and Properties of PHAs Synthesized from Jerusalem Artichoke Hydrolysates
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boey, J.Y.; Mohamad, L.; Khok, Y.S.; Tay, G.S.; Baidurah, S. A review of the applications and biodegradation of polyhydroxyalkanoates and poly(lactic acid) and its composites. Polymers 2021, 13, 1544. [Google Scholar] [CrossRef] [PubMed]
- Mannina, G.; Presti, D.; Montiel-Jarillo, G.; Suárez-Ojeda, M.E. Bioplastic recovery from wastewater: A new protocol for polyhydroxyalkanoates (PHA) extraction from mixed microbial cultures. Bioresour. Technol. 2019, 282, 361–369. [Google Scholar] [CrossRef]
- Huang, L.; Chen, Z.; Wen, Q.; Ji, Y.; Wu, Z.; Lee, D.J. Toward flexible regulation of polyhydroxyalkanoate composition based on substrate feeding strategy: Insights into microbial community and metabolic features. Bioresour. Technol. 2020, 296, 122369. [Google Scholar] [CrossRef] [PubMed]
- Ghanbarzadeh, B.; Almasi, H. Biodegradable polymers. In Biodegradation—Life of Science; IntechOpen: Rijeka, Croatia, 2013; pp. 141–185. [Google Scholar]
- Bong, C.P.C.; Alam, M.N.H.Z.; Samsudin, S.A.; Jamaluddin, J.; Adrus, N.; Yusof, A.H.M.; Muis, Z.A.; Hashim, H.; Salleh, M.M.; Abdullah, A.R.; et al. A review on the potential of polyhydroxyalkanoates production from oil-based substrates. J. Environ. Manag. 2021, 298, 113461. [Google Scholar] [CrossRef]
- Bhola, S.; Arora, K.; Kulshrestha, S.; Mehariya, S.; Bhatia, R.K.; Kaur, P.; Kumar, P. Established and emerging producers of PHA: Redefining the possibility. Appl. Biochem. Biotechnol. 2021, 193, 3812–3854. [Google Scholar] [CrossRef]
- Khosravi-Darani, K.; Mokhtari, Z.B.; Amai, T.; Tanaka, K. Microbial production of poly (hydroxybutyrate) from C1 carbon sources. Appl. Microbiol. Biotechnol. 2013, 97, 1407–1424. [Google Scholar] [CrossRef] [PubMed]
- Laycock, B.; Halley, P.; Pratt, S.; Werker, A.; Lant, P. The chemomechanical properties of microbial polyhydroxyalkanoates. Prog. Polym. Sci. 2013, 38, 536–583. [Google Scholar] [CrossRef]
- Zheng, J.; Suh, S. Strategies to reduce the global carbon footprint of plastics. Nat. Clim. Change 2019, 9, 374–378. [Google Scholar] [CrossRef]
- Chen, G.Q.; Jiang, X.R.; Guo, Y. Synthetic biology of microbes synthesizing polyhydroxyalkanoates (PHA). Synth. Syst. Biotechnol. 2016, 1, 236–242. [Google Scholar] [CrossRef] [Green Version]
- Koller, M.; Mukherjee, A. Polyhydroxyalkanoates—Linking properties, applications, and end-of-life options. Chem. Biochem. Eng. Q. 2020, 34, 115–129. [Google Scholar] [CrossRef]
- Muhamad, I.I.; Sabbagh, F.; Karim, N.A. Polyhydroxyalkanoates: A valuable secondary metabolite produced in microorganisms and plants. In Plant Secondary Metabolites, Volume Three: Their Roles in Stress Eco-Physiology; Apple Academic Press: Oakville, ON, Canada, 2017; pp. 185–214. [Google Scholar]
- Sabbagh, F.; Muhamad, I.I. Production of poly-hydroxyalkanoate as secondary metabolite with main focus on sustainable energy. Renew. Sustain. Energy Rev. 2017, 72, 95–104. [Google Scholar] [CrossRef]
- Sudesh, K.; Abe, H.; Doi, Y. Synthesis, structure and properties of polyhydroxyalkanoates: Biological polyesters. Prog. Polym. Sci. 2000, 25, 1503–1555. [Google Scholar] [CrossRef]
- Rekhi, P.; Goswami, M.; Ramakrishna, S.; Debnath, M. Polyhydroxyalkanoates Biopolymers toward Decarbonizing Economy and Sustainable Future. Available online: https://pubmed.ncbi.nlm.nih.gov/34645360/ (accessed on 30 November 2021).
- Chen, G.Q. Plastics completely synthesized by bacteria: Polyhydroxyalkanoates. In Plastics from Bacteria. Natural Functions and Applications; Springer: Berlin/Heidelberg, Germany, 2010; pp. 17–37. [Google Scholar]
- Volova, T.G.; Shishatskaya, E.I.; Sinskey, A.J. Degradable Polymers: Production, Properties, Applications; Nova Science Pub Inc.: New York, NY, USA, 2013; p. 380. [Google Scholar]
- Volova, T.G.; Vinnik, Y.S.; Shishatskaya, E.I.; Markelova, N.M.; Zaikov, G.E. Natural-Based Polymers for Biomedical Applications; Apple Academic Press: New York, NY, USA, 2017; p. 460. [Google Scholar]
- Volova, T.G.; Shishatskaya, E.I.; Prudnikova, S.V.; Zhila, N.O.; Boyandin, A.N. New Generation Formulations of Agrochemicals: Current Trends and Future Priorities; Apple Academic Press: Boca Raton, FL, USA, 2020; p. 286. [Google Scholar]
- Tarrahi, R.; Fathi, Z.; Özgür, M.; Seydibeyoğlu, K.; Doustkhah, E.; Khataee, A. Polyhydroxyalkanoates (PHA): From production to nanoarchitecture. Int. J. Biol. Macromol. 2020, 146, 596–619. [Google Scholar] [CrossRef]
- Ansari, S.; Sami, N.; Yasin, D.; Ahmad, N.; Fatma, T. Biomedical applications of environmental friendly poly-hydroxyalkanoates. Int. J. Biol. Macromol. 2021, 183, 549–563. [Google Scholar] [CrossRef] [PubMed]
- Hinchliffe, J.D.; Madappura, A.P.; Mohamed, S.M.D.S.; Roy, I. Biomedical applications of bacteria-derived polymers. Polymers 2021, 13, 108117. [Google Scholar] [CrossRef]
- Chen, G.Q.; Chen, X.Y.; Wu, F.Q.; Chen, J.C. Polyhydroxyalkanoates (PHA) toward cost competitiveness and functionality. Adv. Ind. Eng. Polym. Res. 2020, 3, 1–7. [Google Scholar] [CrossRef]
- Saratale, R.G.; Cho, S.-K.; Saratale, G.D.; Kadam, A.A.; Ghodake, G.S.; Kumar, M.; Bharagava, R.N.; Kumar, G.; Kim, D.S.; Mulla, S.I.; et al. A comprehensive overview and recent advances on polyhydroxyalkanoates (PHA) production using various organic waste streams. Bioresour. Technol. 2021, 325, 124685. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, D.; Illman, B.; Crooks, C. Differential sensitivity of polyhydroxyalkanoate producing bacteria to fermentation inhibitors and comparison of polyhydroxybutyrate production from Burkholderia cepacia and Pseudomonas pseudoflava. BMC Res. Notes 2013, 6, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Oh, Y.H.; Jang, Y.A.; Kang, K.H.; David, Y.; Yu, J.H.; Song, B.K.; Choi, J.; Chang, Y.K.; Joo, J.C.; et al. Recombinant Ralstonia eutropha engineered to utilize xylose and its use for the production of poly (3-hydroxybutyrate) from sunflower stalk hydrolysate solution. Microb. Cell Fact. 2016, 15, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, S.K.; Gurav, R.; Choi, T.R.; Jung, H.R.; Yang, S.Y.; Moon, Y.M.; Song, H.S.; Jeon, J.M.; Choi, K.Y.; Yang, Y.H. Bioconversion of plant biomass hydrolysate into bioplastic (polyhydroxyalkanoates) using Ralstonia eutropha 5119. Bioresour. Technol. 2019, 271, 306–315. [Google Scholar] [CrossRef]
- Saratale, G.D.; Bhosale, R.; Shobana, S.; Banu, J.R.; Pugazhendhi, A.; Mahmoud, E.; Sirohi, R.; Bhatia, S.K.; Atabani, A.E.; Mulone, V.; et al. A review on valorization of spent coffee grounds (SCG) towards biopolymers and biocatalysts production. Bioresour. Technol. 2020, 314, 123800. [Google Scholar] [CrossRef] [PubMed]
- Yustinah, N.; Hidayat, N.; Alamsyah, R.; Roslan, A.M.; Hermansyah, H.; Gozan, M. Production of polyhydroxybutyrate from oil palm empty fruit bunch (OPEFB) hydrolysates by Bacillus cereussuaeda B-001. Biocatal. Agric. Biotechnol. 2019, 18, 101019. [Google Scholar] [CrossRef]
- Ahn, J.; Jho, E.H.; Nam, K. Effect of acid-digested rice straw waste feeding methods on the 3HV fraction of bacterial poly (3-hydroxybutyrate-co-3-hydroxyvalerate) production. Process Biochem. 2016, 51, 2119–2126. [Google Scholar] [CrossRef]
- Tanamool, V.; Soemphol, W. Polyhydroxyalkanoate (PHA) synthesis by newly bacterial isolates using non-detoxified rice husk hydrolysate. Asia Pac. J. Sci. Technol. 2016, 21, 404–410. [Google Scholar]
- Soto, L.R.; Byrne, E.; van Niel, E.W.J.; Sayed, M.; Villanueva, C.C.; Hatti-Kaul, R. Hydrogen and polyhydroxybutyrate production from wheat straw hydrolysate using Caldicellulosiruptor species and Ralstonia eutropha in a coupled process. Bioresour. Technol. 2019, 272, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.O.; Kanekar, P.P.; Jog, J.P.; Sarnaik, S.S.; Nilegaonkar, S.S. Production of copolymer, poly(hydroxybutyrate-co-hydroxyvalerate) by Halomonas campisalis MCM B-1027 using agro-wastes. Int. J. Biol. Macromol. 2015, 72, 784–789. [Google Scholar] [CrossRef]
- Saratale, R.G.; Cho, S.K.; Ghodake, G.S.; Shin, H.S.; Saratale, G.D.; Park, Y.; Lee, H.S.; Bharagava, R.N.; Kim, D.S. Utilization of noxious weed water hyacinth biomass as a potential feedstock for biopolymers production: A novel approach. Polymers 2020, 12, 1704. [Google Scholar] [CrossRef]
- Saratale, G.D.; Saratale, R.G.; Varjani, S.; Cho, S.K.; Ghodake, G.S.; Kadam, A.; Mulla, S.I.; Bharagava, R.N.; Kim, D.S.; Shin, H.S. Development of ultrasound aided chemical pretreatment methods to enrich saccharification of wheat waste biomass for polyhydroxybutyrate production and its characterization. Ind. Crops Prod. 2020, 150, 112425. [Google Scholar] [CrossRef]
- Koutinas, A.A.; Garcia, I.L.; Kopsahelis, N.; Papanikolaou, S.; Webb, C.; Villar, M.A.; López, J.A. Production of fermentation feedstock from Jerusalem artichoke tubers and its potential for polyhydroxybutyrate synthesis. Waste Biomass Valor. 2013, 4, 359–370. [Google Scholar] [CrossRef]
- Kakhana, B.M.; Arasimovich, V.V. Biokhimiya Topinambura (Biochemistry of Jerusalem Artichoke); Shtinitsa: Kishinev, Moldova, 1974; p. 88. (In Russian) [Google Scholar]
- Pasko, N.M. Jerusalem artichoke—For feed, technical, food, medicinal and environmental purposes, 3-ya Vsesoyuznaya nauchno-proizvodstvennaya konferentsiya “Topinambur i topisolnechnik—Problem vozdelivaniya i ispolzovaniya. In Proceedings of the 3rd All-Union State Scientific and Production Conference “Jerusalem artichoke and topisflower—Problems of cultivation and use”, Odessa, Ukraine, 7–11 October 1991; p. 134. (In Russian). [Google Scholar]
- Corrado, I.; Cascelli, N.; Ntasi, G.; Birolo, L.; Sannia, G.; Pezzella, C. Optimization of inulin hydrolysis by Penicillium lanosocoeruleum inulinases and efficient conversion into polyhydroxyalkanoates. Front. Bioeng. Biotechnol. 2021, 9, 1–17. [Google Scholar] [CrossRef]
- Ricca, E.; Calabro, V.; Curcio, S.; Iorio, G. Fructose production by chicory inulin enzymatic hydrolysis: A kinetic study and reaction mechanism. Process Biochem. 2009, 44, 466–470. [Google Scholar] [CrossRef]
- Bajpai, P.K.; Bajpai, P. Cultivation and utilization of Jerusalem artichoke for ethanol, single cell protein, and high-fructose syrup production. Enzym. Microb. Technol. 1991, 13, 359–362. [Google Scholar] [CrossRef]
- Guiraud, J.P.; Daurelles, J.; Galzy, P. Alcohol production from Jerusalem artichoke using yeasts with inulinase activity. Biotechnol. Bioeng. 1991, 23, 1461–1465. [Google Scholar] [CrossRef]
- Bekers, M.; Grube, M.; Upite, D.; Kaminska, E.; Linde, R.; Scherbaka, R.; Danilevich, A. Carbohydrates in Jerusalem artichoke powder suspension. Nutr. Food Sci. 2007, 37, 42–49. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Z.; Liang, L.; Wen, S.; Liu, C.; Xu, X. A comparative study of high-performance liquid chromatography and colorimetric method for inulin determination. Eur. Food Res. Technol. 2010, 230, 701–706. [Google Scholar] [CrossRef]
- Chen, L.; Xin, C.; Deng, P.; Ren, J.; Liang, H.; Bai, F. Butanol production from hydrolysate of Jerusalem artichoke juice by Clostridium acetobutylicum L7. Chin. J. Biotechnol. 2010, 26, 991–996. (In Chinese) [Google Scholar]
- Zhao, X.; Wu, S.; Hu, C.; Wang, Q.; Hua, Y.; Zhao, Z.K. Lipid production from Jerusalem artichoke by Rhodosporidium toruloides Y4. J. Ind. Microbiol. Biotechnol. 2010, 37, 581–585. [Google Scholar] [CrossRef]
- Zhao, C.H.; Chi, Z.; Zhang, F.; Guo, F.J.; Li, M.; Song, W.B.; Chi, Z.M. Direct conversion of inulin and extract of tubers of Jerusalem artichoke into single cell oil by co-cultures of Rhodotorula mucilaginosa TJY15a and immobilized inulinase-producing yeast cells. Bioresour. Technol. 2011, 102, 6128–6133. [Google Scholar] [CrossRef]
- Choi, H.Y.; Ryu, H.K.; Park, K.M.; Lee, E.G.; Lee, H.; Kim, S.W.; Choi, E.S. Direct lactic acid fermentation of Jerusalem artichoke tuber extract using Lactobacillus paracasei without acidic or enzymatic inulin hydrolysis. Bioresour. Technol. 2012, 114, 745–747. [Google Scholar] [CrossRef]
- Khatun, M.M.; Liu, C.G.; Zhao, X.Q.; Yuan, W.J.; Bai, F.W. Consolidated ethanol production from Jerusalem artichoke tubers at elevated temperature by Saccharomyces cerevisiae engineered with inulinase expression through cell surface display. J. Ind. Microbiol. Biotechnol. 2017, 44, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Zhang, Y.; Zhu, Y.; Sha, Y.; Xu, Z.; Feng, X.; Li, S.; Xu, H. Improving poly-(γ-glutamic acid) production from a glutamic acid-independent strain from inulin substrate by consolidated bioprocessing. Bioprocess Biosyst. Eng. 2019, 42, 1711–1720. [Google Scholar] [CrossRef] [PubMed]
- Haas, C.; Steinwandter, V.; De Apodaca, E.D.; Madurga, B.M.; Smerilli, M.; Dietrich, T.; Neureiter, M. Production of PHB from chicory roots—Comparison of three Cupriavidus necator strains. Chem. Biochem. Eng. Q. 2015, 29, 99–112. [Google Scholar] [CrossRef]
- Emelina, T.N. Kompleksnaya Pererabotka Vegetativnoi Chasti Topinambura s Polucheniem Produktov Mikrobnogo Sinteza (Complex Processing of the Vegetative Part of Jerusalem Artichoke to Obtain Products of Microbial Synthesis). Ph.D. Thesis, Dissertation for the Degree of Candidate of Biological Sciences, Siberian State Technological University, Krasnoyarsk, Russia, 2003. (In Russian). [Google Scholar]
- Emelina, T.N.; Ryazanova, T.V.; Volova, T.G. The use of substrates from the vegetative part of Jerusalem artichoke in biotechnology. In Proceedings of the Mezhdunarodnaya Nauchno-Prakticheskaya Konferentsiya “Rastitelnie Resursi dlya Zdorovya Cheloveka (Vozdelyvanie, Pererabotka, Marketing)” (International Scientific and Practical Conference “Plant Resources for Human Health (Cultivation, Processing, Marketing)”), Moscow, Russia, 23–27 September 2002; pp. 351–354. (In Russian). [Google Scholar]
- Stasishina, G.N.; Volova, T.G. Shtamm bakterii Alcaligenes eutrophus B5786-Produtsent (The Producing Strain of the Bacteria Alcaligenes eutrophus B5786). RF Patent No. 2053292, BI. No. 1, 27 January 1996. (In Russian). [Google Scholar]
- Volova, T.G.; Kozhevnikov, I.V.; Dolgopolova, Y.B.; Trusova, M.Y.; Kalacheva, G.S.; Aref’eva, Y.V. Physiological and biochemical characteristics and capacity for polyhydroxyalkanoates synthesis in a glucose-utilizing strain of hydrogen-oxidizing bacteria, Ralstonia eutropha B8562. Microbiol. 2005, 74, 788–794. [Google Scholar] [CrossRef]
- Volova, T.G.; Trusova, M.Y.; Kalacheva, G.S.; Kozhevnicov, I.V. Physiological-biochemical properties and the ability to synthesize polyhydroxyalkanoates of the glucose-utilizing strain of the hydrogen bacterium Ralstonia eutropha B8562. Appl. Microbiol. Biotechnol. 2006, 73, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Volova, T.G.; Shishatskaya, E.I. Cupriavidus eutrophus Shtamm Bakterii VKPM B-10646—Produtsent Poligidroksialkanoatov i Sposob Ikh Polucheniya (Cupriavidus eutrophus Bacterial Strain VKPM B-10646—A Producer of Polyhydroxyalkanoates and a Method of Their Production). RF Patent No. 2439143, 10 January 2012. (In Russian). [Google Scholar]
- Vandamme, P.; Coenye, T. Taxonomy of the genus Cupriavidus: A tale of lost and found. Int. J. Syst. Evol. Microbiol. 2004, 54, 2285–2289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlegel, H.G.; Kaltwasser, H.; Gottschalk, G. A submersion method for culture of hydrogen-oxidizing bacteria: Growth physiological studies. Arch. Mikrobiol. 1961, 38, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Zavarzin, G.A. Vodorodnyye Bakterii i Karboksidobakterii (Hydrogen Bacteria and Carboxydobacteria); Nauka: Moscow, Russia, 1978; p. 203. (In Russian) [Google Scholar]
- Tanaka, K.; Ishizaki, A.; Kanamaru, T.; Kawano, T. Production of poly (D-3-hydroxybutyrate) from CO2, H2, and O2 by high cell density autotrophic cultivation of Alcaligenes eutrophus. Biotechnol. Bioeng. 1995, 45, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Miyawaki, K.; Yamaguchi, A.; Khosravi-Darani, K.; Matsusaki, H. Cell growth and P(3HB) accumulation from CO2 of a carbon monoxide-tolerant hydrogen-oxidizing bacterium, Ideonella sp. O-1. Appl. Microbiol. Biotechnol. 2011, 92, 1161–1169. [Google Scholar] [CrossRef]
- Volova, T.G. Hydrogen-Based Biosynthesis; Nova Science Pub Inc.: Hauppauge, NY, USA, 2009; p. 287. [Google Scholar]
- Moreau, C.; Belgacem, M.N.; Gandini, A. Recent catalytic advances in the chemistry of substituted furans from carbohydrates and in the ensuing polymers. Top. Catal. 2004, 27, 11–30. [Google Scholar] [CrossRef]
- Chheda, J.N.; Huber, G.W.; Dumesic, J.A. Liquid-phase catalytic processing of biomass derived oxygenated hydrocarbons to fuels and chemicals. Angew. Chem. Int. Ed. 2007, 46, 7164–7183. [Google Scholar] [CrossRef]
- Marzo, M.; Gerrasini, A.; Carniti, P. Hydrolysis of disaccharides over solid acid catalysts under green conditions. Carbohydr. Res. 2012, 347, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Chesnokov, N.V.; Yatsenkova, O.V.; Chydina, A.I.; Skripnikov, A.M.; Kuznetsov, B.N. The study of the sucrose hydrolysis with acid catalysts. J. Sib. Fed. Univ. Chem. 2012, 5, 311–319. (In Russian) [Google Scholar]
- Samokish, I.I.; Zyablitseva, N.S.; Kompantsev, V.A. Sposob Polucheniya Fruktozosoderzhaschego Produkta iz Topinambura (Method for Producing Fructose-Containing Product from Jerusalem Artichoke). Patent RU No. 2118369 C1, 27 August 1998. (In Russian). [Google Scholar]
- Chuprova, N.A.; Ryazanova, T.V. Production of bioethanol from the vegetative part of Jerusalem artichoke. Chem. Veg Raw 2010, 2, 49–52. (In Russian) [Google Scholar]
- Sharkov, V.I.; Kuybina, N.I.; Solovyeva, Y.P.; Pavlova, T.A. Kolichestvennyi Khimicheskiy Analiz Rastitelnogo Syriya (Quantitative Chemical Analysis of Vegetable Raw Materials); Lesnaya Promyshlennost: Moscow, Russia, 1976; p. 72. (In Russian) [Google Scholar]
- Obolenskaya, A.V.; Elnitskaya, Z.P.; Leonovich, A.A. Loboratornie Raboti po Khimii Drevesini i Cellulozi (Laboratory Work on the Chemistry of Wood and Cellulose); Ekologiya: Moscow, Russia, 1991; p. 320. (In Russian) [Google Scholar]
- Ermakov, A.I.; Arasimovich, V.V.; Smirnova-Ikonnikova, M.I.; Yarosh, N.P.; Lukovnikova, G.A. Metody Biokhimicheskogo Issledovaniya Rastenii (Methods of Biochemical Plant Research); Kolos: Leningrad, Russia, 1972; p. 456. (In Russian) [Google Scholar]
- Volova, T.; Demidenko, A.; Kiselev, E.; Baranovskiy, S.; Shishatskaya, E.; Zhila, N. Polyhydroxyalkanoate synthesis based on glycerol and implementation of the process under conditions of pilot production. Appl. Microbiol. Biotechnol. 2019, 103, 225–237. [Google Scholar] [CrossRef] [Green Version]
- Volova, T.; Kiselev, E.; Zhila, N.; Shishatskaya, E. Synthesis of polyhydroxyalkanoates by hydrogen-oxidizing bacteria in a pilot production process. Biomacromolecules 2019, 20, 3261–3270. [Google Scholar] [CrossRef]
- Kiselev, E.G. Techniko-Technologicheskiye Osnovi Biosinteza Rezervnikh Poligidroksialkanoatov Vodorodnimi Bakteriyami (Technical and Technological Bases of Biosynthesis of Reserve Polyhydroxyalkanoates by Hydrogen Bacteria). Ph.D. Thesis Summary of the Candidate of Sciences Thesis, Institute of Biophysics SB RAS, Krasnoyarsk, Russia, 28 December 2012. (In Russian). [Google Scholar]
- Braunegg, G.; Sonnleitner, B.; Lafferty, R.M. A rapid gas chromatographic method for the determination of poly-b-hydroxybutyric acid in microbial biomass. Eur. J. Appl. Microbiol. Biotechnol. 1978, 6, 29–37. [Google Scholar] [CrossRef]
- ISO 468; Surface Roughness—Parameters, Their Values, and General Rules for Specifying Requirements. International Organization for Standardization: Geneva, Switzerland, 1982.
- Emelyanova, I.Z. Khimiko-Tekhnologicheskii Kontrol Gidroliznogo Proizvodstva (Chemical and Technological Control of Hydrolysis Manufacture); Lesnaya Promishlennost: Moscow, Russia, 1973; p. 120. (In Russian) [Google Scholar]
- Öngen-Baysal, G.; Sukan, S.S. Production of inulinase by mixed culture of Aspergillus niger and Kluyveromyces marxianus. Biotechnol. Lett. 1996, 18, 1431–1434. [Google Scholar] [CrossRef]
- Sirisansaneeyakul, S.; Worawuthiyanan, N.; Vanichsriratana, W.; Srinophakun, P.; Chisti, Y. Production of fructose from inulin using. World J. Microbiol. Biotechnol. 2007, 23, 543–552. [Google Scholar] [CrossRef]
- Koutinas, A.A.; Xu, Y.; Wang, R.; Webb, C. Polyhydroxybutyrate production from a novel feedstock derived from a wheatbased biorefinery. Enzym. Microb. Technol. 2007, 40, 1035–1044. [Google Scholar] [CrossRef]
- Zhila, N.O.; Sapozhnikova, K.Y.; Kiselev, E.G.; Vasiliev, A.D.; Nemtsev, I.V.; Shishatskaya, E.I.; Volova, T.G. Properties of degradable polyhydroxyalkanoates (PHA) synthesized by a new strain of Cupriavidus necator IBP-21 on various C-substrates. Polymers 2021, 13, 3142. [Google Scholar] [CrossRef]
- Zhila, N.O.; Kalacheva, G.S.; Volova, T.G. Fatty acid composition and polyhydroxyalkanoates production by Cupriavidus eutrophus B-10646 cells grown on different carbon sources. Process Biochem. 2015, 50, 69–78. [Google Scholar] [CrossRef]
- Holmes, P.A. Applications of PHB—A microbially produced biodegradable thermoplastic. Phys. Technol. 1985, 16, 32–36. [Google Scholar] [CrossRef]
- Kim, B.C.; Lee, S.Y.; Chang, Y.K.; Chang, Y.K.; Woo, S.I. Production of poly(3-hydroxybutyric-co-3-polyhydroxyvaleric acid) by fed-batch culture of Alcaligenes eutrophus with substrate control using on-line glucose analyze. Enzym. Microb. Technol. 1994, 16, 556–561. [Google Scholar] [CrossRef]
- Ryu, H.W.; Cho, K.S.; Kim, B.S.; Chang, H.N.; Shim, H.J. Mass production of poly(3-hydroxybutyrate) in fed-batch culture of Ralstonia eutropha with nitrogen and phosphate limitation. J. Microbiol. Biotechnol. 1999, 9, 751–756. [Google Scholar]
- Chanprateep, S.; Kulpreecha, S. Production and characterization of biodegradable terpolymer poly(3-hydroxybutyrate-co-3-hydroxyvalerate-co-4-hydroxybutyrate) by Alcaligenes sp. A-04. J. Biosci. Bioeng. 2006, 101, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Volova, T.G.; Kiselev, E.G.; Vinogradova, O.; Nikolaeva, E.; Chistyakov, A.; Sukovatyi, A.; Shishatskaya, E.I. A glucose-utilizing strain, Cupriavidus eutrophus B-10646: Growth kinetics, characterization and synthesis of multicomponent PHAs. PLoS ONE 2014, 9, e87551. [Google Scholar]
- Volova, T.; Kiselev, E.; Nemtsev, I.; Lukyanenko, A.; Sukovatyi, A.; Kuzmin, A.; Ryltseva, G.; Shishatskaya, E. Properties of degradable PHAs with different monomer compositions. Int. J. Biol. Macromol. 2021, 182, 98–114. [Google Scholar] [CrossRef] [PubMed]
- Bera, A.; Dubey, S.; Bhayani, K.; Mondal, D.; Mishra, S.; Ghosh, P.K. Microbial synthesis of polyhydroxyalkanoate using seaweed-derived crude levulinic acid as co-nutrient. Int. J. Biol. Macromol. 2015, 72, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Poley, L.H.; Siqueira, A.P.L.; da Silva, M.G.; Sanchez, R.; Prioli, R.; Mansanares, A.M.; Vargas, H. Photothermal methods and atomic force microscopy images applied to the study of poly(3-hydroxybutyrate) and poly(3-hydroxybutyrate-co-3-hydroxyvalerate) dense membranes. J. Appl. Polym. Sci. 2005, 97, 1491–1497. [Google Scholar] [CrossRef]
- Zhila, N.O.; Shishatskaya, E.I. Properties of PHA bi-, ter-, and quarter-polymers containing 4-hydroxybutyrate monomer units. Int. J. Biol. Macromol. 2018, 111, 1019–1026. [Google Scholar] [CrossRef] [Green Version]
- Chanprateep, S.; Buasri, K.; Muangwong, A.; Utiswannakul, P. Biosynthesis and biocompatibility of biodegradable poly(3-hydroxybutyrate-co-4-hydroxybutyrate). Polym. Degrad. Stab. 2010, 95, 2003–2010. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Content, % | |
|---|---|---|
| Tubers | Vegetative Biomass | |
| Moisture content | 74.0 ± 3.7 | 12.0 ± 0.6 |
| Solids including | 26.0 ± 0.9 | 88.0 ± 4.4 |
| Minerals | 4.2 ± 0.3 | 8.5 ± 0.6 |
| Monosaccharides | 6.4 ± 0.6 | 7.9 ± 0.7 |
| Oligosaccharides | 12.7 ± 1.0 | 18.7 ± 1.1 |
| Crude protein | 2.5 ± 0.3 | - |
| Pectic substances | 1.2 ± 0.1 | - |
| Easy-to-hydrolyze polysaccharides, % CDW | 34.1 ± 2.4 | 10.7 ± 0.7 |
| Hard-to-hydrolyze polysaccharides, % CDW | 12.3 ± 0.9 | 23.5 ± 1.9 |
| Lignins, % CDW | 9.1 ± 0.4 | 16.2 ± 0.9 |
| Parameter, % | Aqueous Extraction at 80 °C Followed by Acid Hydrolysis at 60 °C | Simultaneous Extraction and Acid Hydrolysis at 80 °C |
|---|---|---|
| Tuber hydrolysates: | ||
| Reducing substances (RS) | 59.8 ± 4.2 | 73.2 ± 5.1 |
| Monosaccharides (% of RS): | 49.1 ± 2.9 | 66.0 ± 3.3 |
| galactose | - | - |
| glucose | 4.3 ± 0.3 | 7.8 ± 0.4 |
| fructose | 44.7 ± 2.2 | 58.2 ± 2.3 |
| Vegetative biomass hydrolysates: | ||
| Reducing substances (RS) | 31.0 ± 1.2 | 11.5 ± 0.7 |
| Monosaccharides (% of RS): | ||
| galactose | 15.9 ± 0.8 | 1.1 ± 0.1 |
| glucose | 34.1 ± 1.3 | 44.6 ± 1.3 |
| fructose | 45.1 ± 1.2 | 38.1 ± 1.3 |
| Element | Tubers | Vegetative Biomass | ||
|---|---|---|---|---|
| Aqueous Extraction at 80 °C Followed by Acid Hydrolysis at 60 °C | Simultaneous Extraction and Acid Hydrolysis at 80 °C | Aqueous Extraction at 80 °C Followed by Acid Hydrolysis at 60 °C | Simultaneous Extraction and Acid Hydrolysis at 80 °C | |
| S | 59 | 1127 | 89 | 1324.00 |
| P | 286 | 211 | 121 | 110 |
| K | 2944 | 2900 | 1570 | 830 |
| Na | 32 | 35 | 30 | 79 |
| Ca | 73 | 88 | 122 | 121 |
| Mg | 72 | 112 | 135 | 138 |
| Fe | 0.07 | 4.9 | 5.8 | 48.0 |
| Cu | 0.23 | 0.38 | 0.27 | 0.46 |
| Zn | 0.05 | 2.1 | 0.75 | 5.11 |
| Mn | 0.17 | 1.12 | 1.55 | 14.3 |
| Cr | 0.0063 | 0.081 | 0.38 | 7.5 |
| Ni | 0.05 | 0.1 | 0.07 | 72.3 |
| B | 0.3 | 0.31 | 0.21 | 0.48 |
| Mo | 0.01 | 0.04 | 0.01 | 0.23 |
| PHA Producing Strain | X, g/L | PHA, % | Culture Conditions | Type of Hydrolysate | Reference |
|---|---|---|---|---|---|
| Cupriavidus necator B-10646 | 7.0 | 70.0 | Shake flasks, 72 h | JA tuber hydrolysates, aqueous extraction at 80 °C followed by acid hydrolysis at 60 °C | This study |
| Ralstonia eutropha B5786 | 5.7 | 62.2 | Shake flasks, 72 h | JA tuber hydrolysates, Aqueous extraction at 80 °C followed by acid hydrolysis at 60 °C | This study |
| Ralstonia eutropha B8562 | 6.9 | 67.8 | Shake flasks, 72 h | JA tuber hydrolysates, Aqueous extraction at 80 °C followed by acid hydrolysis at 60 °C | This study |
| Cupriavidus necator B-10646 | 5.6 | 63.0 | Shake flasks, 72 h | JA tuber hydrolysates, Simultaneous extraction and acid hydrolysis at 80 °C | This study |
| Ralstonia eutropha B5786 | 3.4 | 49.1 | Shake flasks, 72 h | JA tuber hydrolysates, Simultaneous extraction and acid hydrolysis at 80 °C | This study |
| Ralstonia eutropha B8562 | 5.2 | 61.8 | Shake flasks, 72 h | JA tuber hydrolysates, Simultaneous extraction and acid hydrolysis at 80 °C | This study |
| Cupriavidus necator B-10646 | 6.9 | 67.9 | Shake flasks, 72 h | JA vegetative biomass hydrolysates, Aqueous extraction at 80 °C followed by acid hydrolysis at 60 °C | This study |
| Ralstonia eutropha B5786 | 5.3 | 57.7 | Shake flasks, 72 h | JA vegetative biomass hydrolysates, Aqueous extraction at 80 °C followed by acid hydrolysis at 60 °C | This study |
| Ralstonia eutropha B8562 | 6.4 | 60.0 | Shake flasks, 72 h | JA vegetative biomass hydrolysates, Aqueous extraction at 80 °C followed by acid hydrolysis at 60 °C | This study |
| Cupriavidus necator B-10646 | 5.6 | 64.3 | Shake flasks, 72 h | JA vegetative biomass hydrolysates, Simultaneous extraction and acid hydrolysis at 80 °C | This study |
| Ralstonia eutropha B5786 | 3.4 | 48.9 | Shake flasks, 72 h | JA vegetative biomass hydrolysates, Simultaneous extraction and acid hydrolysis at 80 °C | This study |
| Ralstonia eutropha B8562 | 5.3 | 60.8 | Shake flasks, 72 h | JA vegetative biomass hydrolysates, Simultaneous extraction and acid hydrolysis at 80 °C | This study |
| Ralstonia eutropha Z-1 | 6–7 | 60–70 | Shake flasks, 72–96 h | JA vegetative biomass acid hydrolysate | [53] |
| Cupriavidus necator DSM 428 | 11.3 | 66 | Bioreactor, 120 h | Hydrolysate from chicory roots | [51] |
| Cupriavidus necator DSM 545 | 14 | 78 | Bioreactor, 72 h | Hydrolysate from chicory roots | [51] |
| Cupriavidus necator DSM 531 | 3.5 | 46 | Bioreactor, 120 h | Hydrolysate from chicory roots | [51] |
| Cupriavidus necator NCIMB 11599 | 3.5–20.8 | 10–70% | Shake flasks, 18–35 h | Wheat hydrolysates and fungal extract | [81] |
| Cupriavidus necator DSM 4058 | 5.3–9.2 | 15–52 | Shake flasks, 32–56 h | JA tubers hydrolysate and yeast extract | [36] |
| Cupriavidus necator DSM 428 | 3.9 | 82 | 120 h | Inulin and fungal inulinase mixture | [39] |
| Strain | Culture Conditions | Number Average Molecular Weight, Mn, kDa | Weigh Average Molecular Weight, Mw, kDa | Polydispersity, Ð | Degree of Crystallinity, Cx, % | Melting Point, Tmelt, °C | Thermal Degradation Temperature, Tdegr, °C |
|---|---|---|---|---|---|---|---|
| P(3HB) | |||||||
| Hydrolysate of JA tubers | |||||||
| C. necator B-10646 | Shake flasks, 72 h | 104 | 432 | 4.2 | 71 | 176 | 293 |
| R. eutropha B5786 | Shake flasks, 72 h | 121 | 480 | 4.0 | 72 | 178 | 290 |
| R. eutropha B8562 | Shake flasks, 72 h | 111 | 466 | 4.2 | 72 | 178 | 296 |
| Hydrolysate of JA vegetative biomass | |||||||
| C. necator B-10646 | Shake flasks, 72 h | 116 | 448 | 3.9 | 70 | 175 | 280 |
| R. eutropha B5786 | Shake flasks, 72 h | 110 | 430 | 4.0 | 75 | 171 | 290 |
| R. eutropha B8562 | Shake flasks, 72 h | 103 | 409 | 4.0 | 69 | 177 | 281 |
| Hydrolysate of JA tubers | |||||||
| P(3HB-co-9.9 mol.%3HV) | |||||||
| C. necator B-10646 | Shake flasks, 72 h; addition of sodium valerate (1 g/L) | 129 | 620 | 4.8 | 60 | 170 | 275 |
| P(3HB-co-37.4 mol.%3HV) | |||||||
| C. necator B-10646 | Shake flasks, 72 h; addition of sodium valerate (2 g/L) | 194 | 1170 | 6.0 | 49 | 174 | 283 |
| P(3HB-co-11.9 mol.%4HB) | |||||||
| C. necator B-10646 | Shake flasks, 72 h; addition of ε-caprolactone (2 g/L) | 147 | 527 | 3.6 | 54 | 170 | 285 |
| P(3HB-co-21.1 mol.%4HB) | |||||||
| C. necator B-10646 | Shake flasks, 72 h; addition of ε-caprolactone (4 g/L) | 162 | 550 | 3.4 | 46 | 169 | 284 |
| Porosity | Surface Roughness: | ||||
|---|---|---|---|---|---|
| Average Pore Area, µm2 | Number of Pores, Pores/1000 µm2 | Total Pores Area, µm2/1000 µm2 | Arithmetic Mean Surface Roughness, (Sa) nm | Root Mean Square Roughness, (Sq) nm | Peak-to-Valley Height, (Sz) nm |
| P(3HB) | |||||
| 7.9 | 20.8 | 164 | 163.10 | 211.01 | 1047.38 |
| P(3HB-co-9.9 mol.%3HV) | |||||
| 30.8 | 25.6 | 788 | 244.00 | 318,71 | 1780.01 |
| P(3HB-co-37.4 mol.%3HV) | |||||
| 4.7 | 49.6 | 235 | 196.35 | 244.61 | 1038.91 |
| P(3HB-co-11.9 mol.%4HB) | |||||
| 3.7 | 20.8 | 77 | 111.94 | 141.98 | 605.22 |
| P(3HB-co-21.1 mol.%4HB) | |||||
| 1.7 | 103.2 | 173 | 131.20 | 167.60 | 641.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volova, T.G.; Kiselev, E.G.; Demidenko, A.V.; Zhila, N.O.; Nemtsev, I.V.; Lukyanenko, A.V. Production and Properties of Microbial Polyhydroxyalkanoates Synthesized from Hydrolysates of Jerusalem Artichoke Tubers and Vegetative Biomass. Polymers 2022, 14, 132. https://doi.org/10.3390/polym14010132
Volova TG, Kiselev EG, Demidenko AV, Zhila NO, Nemtsev IV, Lukyanenko AV. Production and Properties of Microbial Polyhydroxyalkanoates Synthesized from Hydrolysates of Jerusalem Artichoke Tubers and Vegetative Biomass. Polymers. 2022; 14(1):132. https://doi.org/10.3390/polym14010132
Chicago/Turabian StyleVolova, Tatiana G., Evgeniy G. Kiselev, Alexey V. Demidenko, Natalia O. Zhila, Ivan V. Nemtsev, and Anna V. Lukyanenko. 2022. "Production and Properties of Microbial Polyhydroxyalkanoates Synthesized from Hydrolysates of Jerusalem Artichoke Tubers and Vegetative Biomass" Polymers 14, no. 1: 132. https://doi.org/10.3390/polym14010132
APA StyleVolova, T. G., Kiselev, E. G., Demidenko, A. V., Zhila, N. O., Nemtsev, I. V., & Lukyanenko, A. V. (2022). Production and Properties of Microbial Polyhydroxyalkanoates Synthesized from Hydrolysates of Jerusalem Artichoke Tubers and Vegetative Biomass. Polymers, 14(1), 132. https://doi.org/10.3390/polym14010132