
Enzymatic Degradation of the Most Common Aliphatic Bio-Polyesters and Evaluation of the Mechanisms Involved: An Extended Study

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
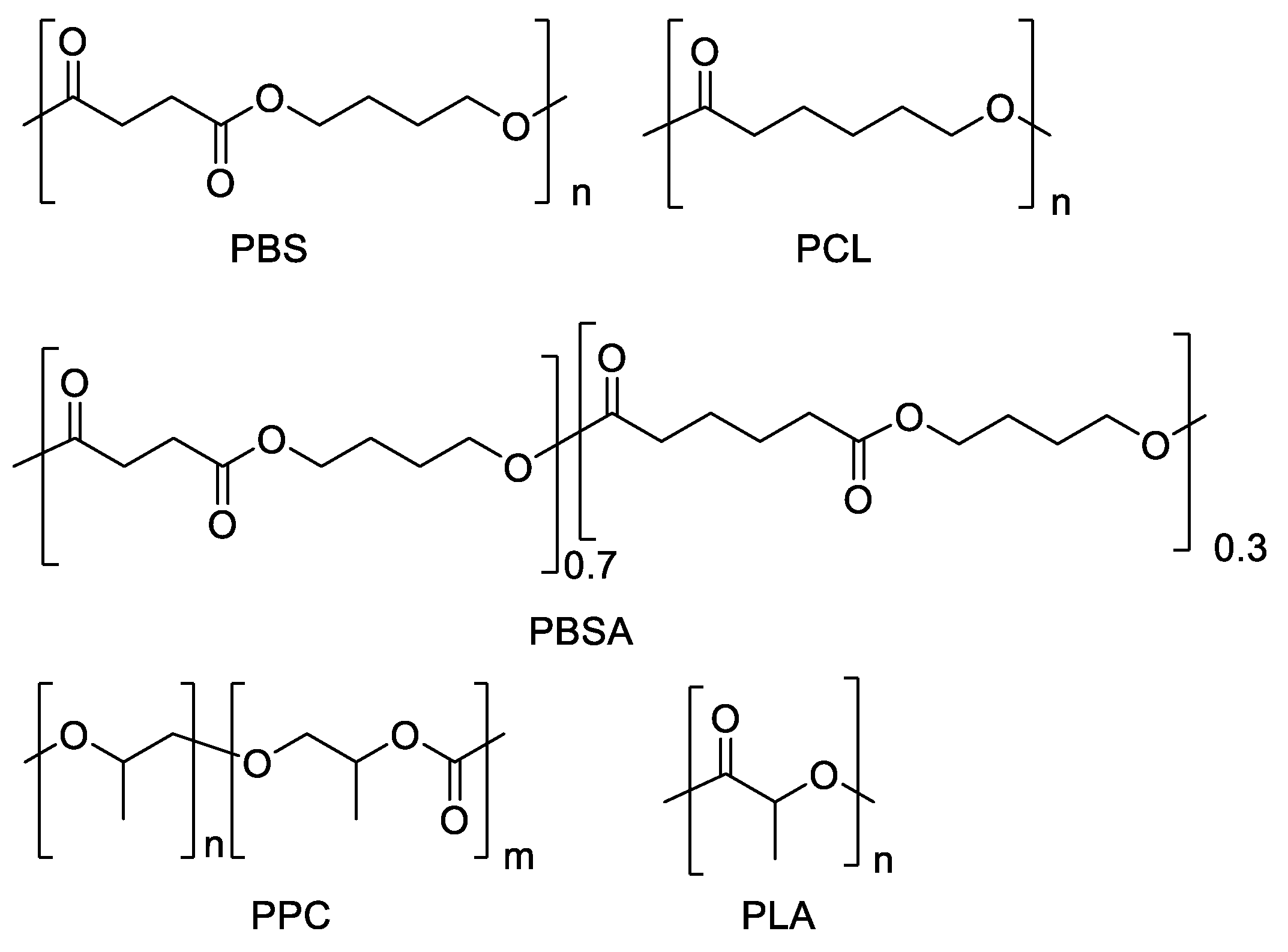
2.1. Polyester Samples and Their Characterization Measurements
2.2. Enzymes and Degradability Tests
2.3. Evaluation of Polyester Enzymatic Degradation Mechanisms
3. Results and Discussion
3.1. Polyester Characterizations
3.2. Degradation of Polyesters by Hydrolytic Enzymes
3.2.1. Enzymes General Screening
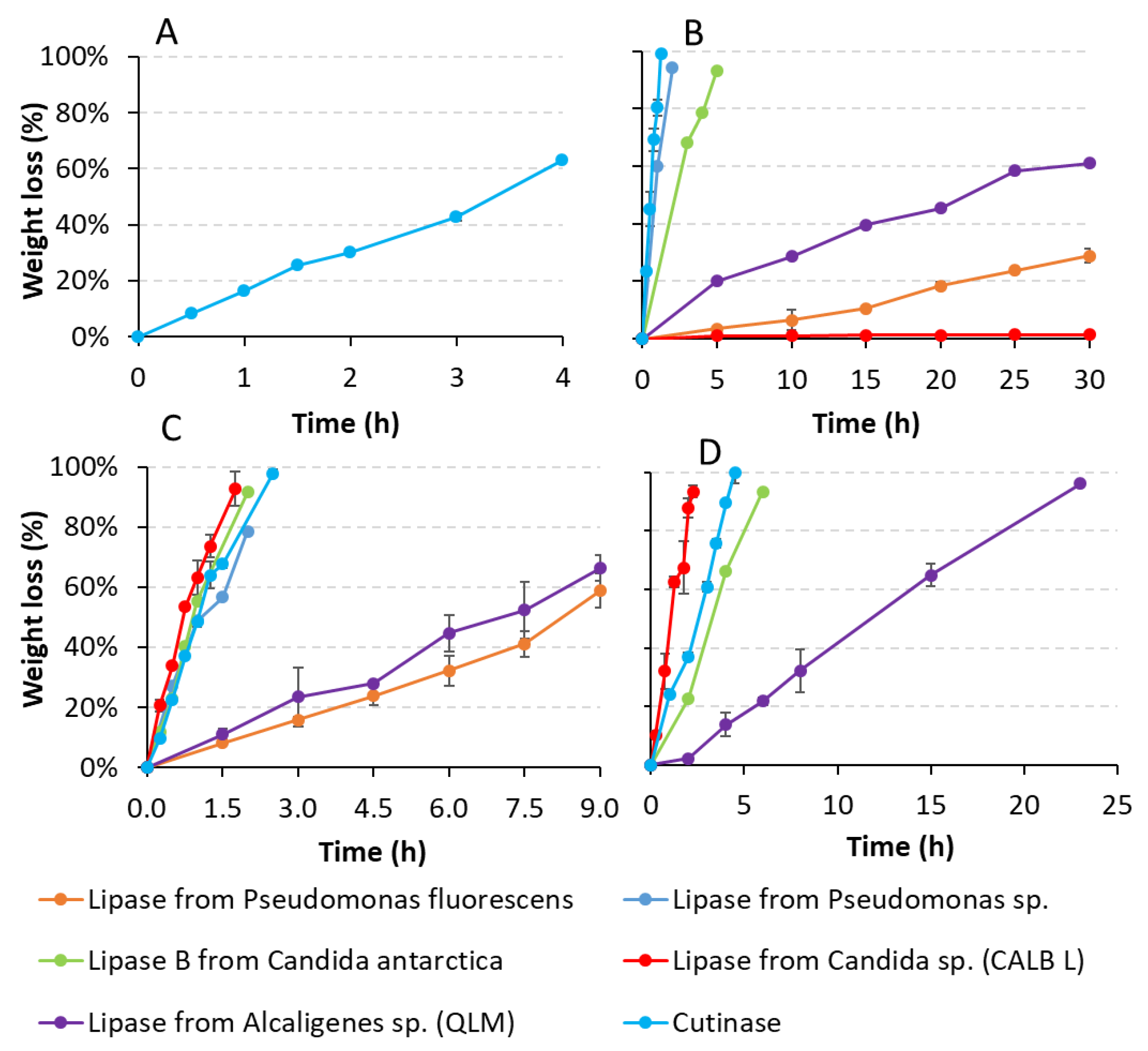
3.2.2. Weight Loss Tests: Specific Polymers and Target Enzymes
3.3. Enzymatic Degradation Mechanisms of PBS, PBSA and PCL Polymers
3.3.1. Solid Residue Films Characterization
3.3.2. Liquid Fraction Characterization: Organic Extraction
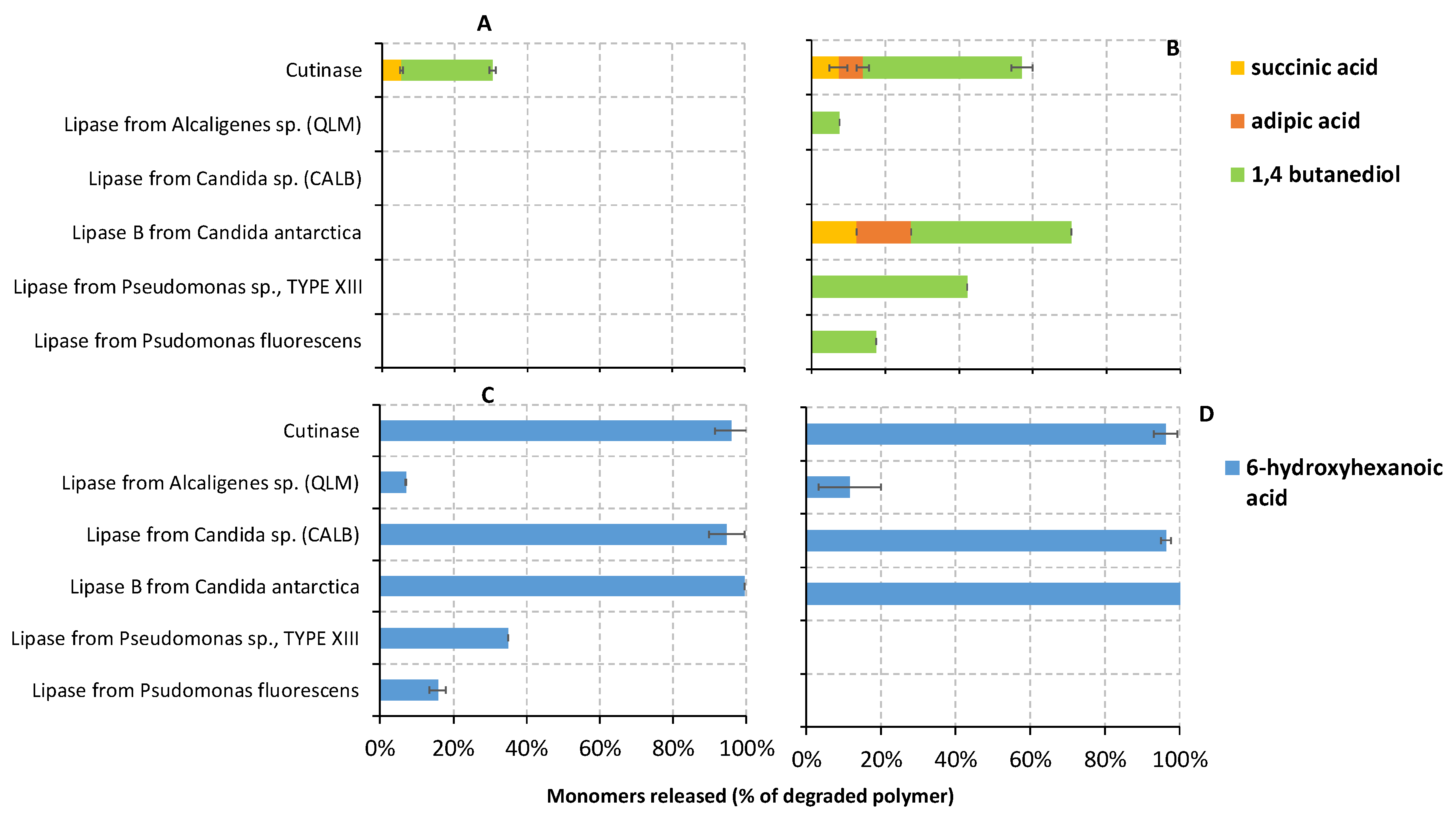
3.3.3. Liquid Fraction Characterization: Water-Soluble Monomers
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gamerith, C.; Vastano, M.; Ghorbanpour, S.M.; Zitzenbacher, S.; Ribitsch, D.; Zumstein, M.T.; Sander, M.; Acero, E.H.; Pellis, A.; Guebitz, G.M. Enzymatic Degradation of Aromatic and Aliphatic Polyesters by P. pastoris Expressed Cutinase 1 from Thermobifida cellulosilytica. Front. Microbiol. 2017, 8, 938. [Google Scholar] [CrossRef] [PubMed]
- Hahladakis, J.N. Delineating and preventing plastic waste leakage in the marine and terrestrial environment. Environ. Sci. Pollut. Res. 2020, 27, 12830–12837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.-C.; Lin, C.-F. CO2 emission from municipal solid waste incinerator: IPCC formula estimation and flue gas measurement. J. Environ. Eng. Manag. 2010, 20, 9–17. [Google Scholar]
- Hoshino, A.; Isono, Y. Degradation of aliphatic polyester films by commercially available lipases with special reference to rapid and complete degradation of poly(L-lactide) film by lipase PL derived from Alcaligenes sp. Biodegradation 2002, 13, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.-A.; Raku, T.; Tokiwa, Y. Hydrolysis of polyesters by serine proteases. Biotechnol. Lett. 2005, 27, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-A.; Tsai, G.-S.; Chen, E.-C.; Wu, T.-M. Thermal degradation behaviors and biodegradability of novel nanocomposites based on various poly[(butylene succinate)-co-adipate] and modified layered double hydroxides. J. Taiwan Inst. Chem. Eng. 2017, 77, 263–270. [Google Scholar] [CrossRef]
- Tokiwa, Y.; Calabia, B.P. Biodegradability and Biodegradation of Polyesters. J. Polym. Environ. 2007, 15, 259–267. [Google Scholar] [CrossRef]
- Meereboer, K.W.; Misra, M.; Mohanty, A.K. Review of recent advances in the biodegradability of polyhydroxyalkanoate (PHA) bioplastics and their composites. Green Chem. 2020, 22, 5519–5558. [Google Scholar] [CrossRef]
- Cook, W.J.; Cameron, J.A.; Bell, J.P.; Huang, S.J. Scanning electron microscopic visualization of biodegradation of polycaprolactones by fungi. J. Polym. Sci. Polym. Lett. Ed. 1981, 19, 159–165. [Google Scholar] [CrossRef]
- Salomez, M.; George, M.; Fabre, P.; Touchaleaume, F.; Cesar, G.; Lajarrige, A.; Gastaldi, E. A comparative study of degradation mechanisms of PHBV and PBSA under laboratory-scale composting conditions. Polym. Degrad. Stab. 2019, 167, 102–113. [Google Scholar] [CrossRef]
- Rizzarelli, P.; Puglisi, C.; Montaudo, G. Soil burial and enzymatic degradation in solution of aliphatic co-polyesters. Polym. Degrad. Stab. 2004, 85, 855–863. [Google Scholar] [CrossRef]
- Bai, Z.; Shi, K.; Su, T.; Wang, Z. Correlation between the chemical structure and enzymatic hydrolysis of Poly(butylene succinate), Poly(butylene adipate), and Poly(butylene suberate). Polym. Degrad. Stab. 2018, 158, 111–118. [Google Scholar] [CrossRef]
- Shah, A.A.; Kato, S.; Shintani, N.; Kamini, N.R.; Nakajima-Kambe, T. Microbial degradation of aliphatic and aliphatic-aromatic co-polyesters. Appl. Microbiol. Biotechnol. 2014, 98, 3437–3447. [Google Scholar] [CrossRef] [PubMed]
- Masaki, K.; Kamini, N.R.; Ikeda, H.; Iefuji, H. Cutinase-Like Enzyme from the Yeast Cryptococcus sp. Strain S-2 Hydrolyzes Polylactic Acid and Other Biodegradable Plastics. Appl. Environ. Microbiol. 2005, 71, 7548–7550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastorino, L.; Pioli, F.; Zilli, M.; Converti, A.; Nicolini, C. Lipase-catalyzed degradation of poly(ε-caprolactone). Enzym. Microb. Technol. 2004, 35, 321–326. [Google Scholar] [CrossRef]
- Wei, X.-F.; Capezza, A.J.; Cui, Y.; Li, L.; Hakonen, A.; Liu, B.; Hedenqvist, M.S. Millions of microplastics released from a biodegradable polymer during biodegradation/enzymatic hydrolysis. Water Res. 2022, 211, 118068. [Google Scholar] [CrossRef] [PubMed]
- Mueller, R. Mechanism of polyester degradation by enzymes. Polym. Prepr. 2007, 48. [Google Scholar]
- Azevedo, H.S.; Reis, R.L. Understanding the Enzymatic Degradation of Biodegradable Polymers and Strategies to Control Their Degradation Rate. In Biodegradable Systems in Tissue Engineering and Regenerative Medicine; CRC Press LLC: Boca Raton, FL, USA, 2005. [Google Scholar]
- Lee, C.W.; Kimura, Y.; Chung, J.-D. Mechanism of enzymatic degradation of poly(butylene succinate). Macromol. Res. 2008, 16, 651–658. [Google Scholar] [CrossRef]
- Shi, K.; Su, T.; Wang, Z. Comparison of poly(butylene succinate) biodegradation by Fusarium solani cutinase and Candida antarctica lipase. Polym. Degrad. Stab. 2019, 164, 55–60. [Google Scholar] [CrossRef]
- Sato, S.; Saika, A.; Shinozaki, Y.; Watanabe, T.; Suzuki, K.; Sameshima-Yamashita, Y.; Fukuoka, T.; Habe, H.; Morita, T.; Kitamoto, H. Degradation profiles of biodegradable plastic films by biodegradable plastic-degrading enzymes from the yeast Pseudozyma antarctica and the fungus Paraphoma sp. B47-9. Polym. Degrad. Stab. 2017, 141, 26–32. [Google Scholar] [CrossRef]
- Chahiniana, H.; Sarda, L. Distinction Between Esterases and Lipases: Comparative Biochemical Properties of Sequence-Related Carboxylesterases. Protein Pept. Lett. 2009, 16, 1149–1161. [Google Scholar] [CrossRef] [PubMed]
- Gan, Z.; Liang, Q.; Zhang, J.; Jing, X. Enzymatic degradation of poly(ε-caprolactone) film in phosphate buffer solution containing lipases. Polym. Degrad. Stab. 1997, 56, 209–213. [Google Scholar] [CrossRef]
- Hwang, Y.; Ree, M.; Kim, H. Enzymatic degradation of poly(propylene carbonate) and poly(propylene carbonate-co-ε-caprolactone) synthesized via CO2 fixation. Catal. Today 2006, 115, 288–294. [Google Scholar] [CrossRef]
- Biffinger, J.C.; Barlow, D.E.; Cockrell, A.L.; Cusick, K.D.; Hervey, W.J.; Fitzgerald, L.A.; Nadeau, L.J.; Hung, C.S.; Crookes-Goodson, W.J.; Russell, J.N. The applicability of Impranil®DLN for gauging the biodegradation of polyurethanes. Polym. Degrad. Stab. 2015, 120, 178–185. [Google Scholar] [CrossRef]
- Murray, E.; Thompson, B.C.; Sayyar, S.; Wallace, G.G. Enzymatic degradation of graphene/polycaprolactone materials for tissue engineering. Polym. Degrad. Stab. 2015, 111, 71–77. [Google Scholar] [CrossRef]
- Khan, I.; Dutta, J.R.; Ganesan, R. Lactobacillus sps. lipase mediated poly (ε-caprolactone) degradation. Int. J. Biol. Macromol. 2017, 95, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Gigli, M.; Negroni, A.; Soccio, M.; Zanaroli, G.; Lotti, N.; Fava, F.; Munari, A. Influence of chemical and architectural modifications on the enzymatic hydrolysis of poly(butylene succinate). Green Chem. 2012, 14, 2885–2893. [Google Scholar] [CrossRef]
- Ding, M.; Zhang, M.; Yang, J.; Qiu, J.-H. Study on the enzymatic degradation of PBS and its alcohol acid modified copolymer. Biodegradation 2011, 23, 127–132. [Google Scholar] [CrossRef]
- Gautam, R.; Bassi, A.S.; Yanful, E.K. Candida rugosa lipase-catalyzed polyurethane degradation in aqueous medium. Biotechnol. Lett. 2007, 29, 1081–1086. [Google Scholar] [CrossRef]
- Jbilou, F.; Dole, P.; Degraeve, P.; Ladavière, C.; Joly, C. A green method for polybutylene succinate recycling: Depolymerization catalyzed by lipase B from Candida antarctica during reactive extrusion. Eur. Polym. J. 2015, 68, 207–215. [Google Scholar] [CrossRef]
- Nakajima-Kambe, T.; Edwinoliver, N.G.; Maeda, H.; Thirunavukarasu, K.; Gowthaman, M.K.; Masaki, K.; Mahalingam, S.; Kamini, N.R. Purification, cloning and expression of an Aspergillus niger lipase for degradation of poly(lactic acid) and poly(ε-caprolactone). Polym. Degrad. Stab. 2012, 97, 139–144. [Google Scholar] [CrossRef]
- Bai, Z.; Liu, Y.; Su, T.; Wang, Z. Effect of Hydroxyl Monomers on the Enzymatic Degradation of Poly(ethylene succinate), Poly(butylene succinate), and Poly(hexylene succinate). Polymers 2018, 10, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartnikowski, M.; Dargaville, T.R.; Ivanovski, S.; Hutmacher, D.W. Degradation mechanisms of polycaprolactone in the context of chemistry, geometry and environment. Prog. Polym. Sci. 2019, 96, 1–20. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, T.; Long, L.; Zhang, R.; Ding, S. Efficient enzymatic degradation of poly (ε-caprolactone) by an engineered bifunctional lipase-cutinase. Polym. Degrad. Stab. 2019, 160, 120–125. [Google Scholar] [CrossRef]
- Hu, X.; Gao, Z.; Wang, Z.; Su, T.; Yang, L.; Li, P. Enzymatic degradation of poly(butylene succinate) by cutinase cloned from Fusarium solani. Polym. Degrad. Stab. 2016, 134, 211–219. [Google Scholar] [CrossRef]
- Bornscheuer, U.T.; Pohl, M. Improved biocatalysts by directed evolution and rational protein design. Curr. Opin. Chem. Biol. 2001, 5, 137–143. [Google Scholar] [CrossRef]
- Flores-Gallegos, A.C.; Delgado-García, M.; Ascacio-Valdés, J.A.; Villareal-Morales, S.; Michel-Michel, M.R.; Aguilar-González, C.N.; Rodríguez-Herrera, R. Hydrolases of Halophilic Origin with Importance for the Food Industry. In Enzymes in Food Biotechnology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 197–219. [Google Scholar] [CrossRef]
- Oda, Y.; Yonetsu, A.; Urakami, T.; Tonomura, K. Degradation of Polylactide by Commercial Proteases. J. Polym. Environ. 2000, 8, 29–32. [Google Scholar] [CrossRef]
- Kawai, F.; Nakadai, K.; Nishioka, E.; Nakajima, H.; Ohara, H.; Masaki, K.; Iefuji, H. Different enantioselectivity of two types of poly(lactic acid) depolymerases toward poly(l-lactic acid) and poly(d-lactic acid). Polym. Degrad. Stab. 2011, 96, 1342–1348. [Google Scholar] [CrossRef]
- Totaro, G.; Sisti, L.; Vannini, M.; Marchese, P.; Tassoni, A.; Lenucci, M.S.; Lamborghini, M.; Kalia, S.; Celli, A. A new route of valorization of rice endosperm by-product: Production of polymeric biocomposites. Compos. Part B Eng. 2018, 139, 195–202. [Google Scholar] [CrossRef]
- Sisti, L.; Totaro, G.; Cionci, N.B.; Di Gioia, D.; Celli, A.; Verney, V.; Leroux, F. Olive Mill Wastewater Valorization in Multifunctional Biopolymer Composites for Antibacterial Packaging Application. Int. J. Mol. Sci. 2019, 20, 2376. [Google Scholar] [CrossRef] [Green Version]
- Sisti, L.; Totaro, G.; Celli, A.; Marek, A.A.; Verney, V.; Leroux, F. Chain extender effect of 3-(4-hydroxyphenyl)propionic acid/layered double hydroxide in biopolyesters containing the succinate moiety. New J. Chem. 2020, 44, 10127–10136. [Google Scholar] [CrossRef]
- Labet, M.; Thielemans, W. Synthesis of polycaprolactone: A review. Chem. Soc. Rev. 2009, 38, 3484–3504. [Google Scholar] [CrossRef] [PubMed]
- Kweon, H.; Yoo, M.K.; Park, I.K.; Kim, T.H.; Lee, H.C.; Lee, H.-S.; Oh, J.-S.; Akaike, T.; Cho, C.-S. A novel degradable polycaprolactone networks for tissue engineering. Biomaterials 2003, 24, 801–808. [Google Scholar] [CrossRef]
- Farah, S.; Anderson, D.G.; Langer, R. Physical and mechanical properties of PLA, and their functions in widespread applications—A comprehensive review. Adv. Drug Deliv. Rev. 2016, 107, 367–392. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-Y.; Weng, Y.-X.; Wang, W.; Huang, Z.-G.; Wang, Y.-Z. Modification of poly(propylene carbonate) with chain extender ADR-4368 to improve its thermal, barrier, and mechanical properties. Polym. Test. 2016, 54, 301–307. [Google Scholar] [CrossRef]
- Chen, Y.; Tan, L.; Chen, L.; Yang, Y.; Wang, X. Study on biodegradable aromatic/aliphatic copolyesters. Braz. J. Chem. Eng. 2008, 25, 321–335. [Google Scholar] [CrossRef]
- Sisti, L.; Kalia, S.; Totaro, G.; Vannini, M.; Negroni, A.; Zanaroli, G.; Celli, A. Enzymatically treated curaua fibers in poly(butylene succinate)-based biocomposites. J. Environ. Chem. Eng. 2018, 6, 4452–4458. [Google Scholar] [CrossRef]
- Seggiani, M.; Altieri, R.; Cinelli, P.; Esposito, A.; Lazzeri, A. Thermoplastic Blends Based on Poly(Butylene Succinate-co-Adipate) and Different Collagen Hydrolysates from Tanning Industry: I—Processing and Thermo-mechanical Properties. J. Polym. Environ. 2020, 29, 392–403. [Google Scholar] [CrossRef]
- Aliotta, L.; Vannozzi, A.; Canesi, I.; Cinelli, P.; Coltelli, M.-B.; Lazzeri, A. Poly(lactic acid) (PLA)/Poly(butylene succinate-co-adipate) (PBSA) Compatibilized Binary Biobased Blends: Melt Fluidity, Morphological, Thermo-Mechanical and Micromechanical Analysis. Polymers 2021, 13, 218. [Google Scholar] [CrossRef]
- Guarino, V.; Cirillo, V.; Taddei, P.; Alvarez-Perez, M.A.; Ambrosio, L. Tuning Size Scale and Crystallinity of PCL Electrospun Fibres via Solvent Permittivity to Address hMSC Response. Macromol. Biosci. 2011, 11, 1694–1705. [Google Scholar] [CrossRef]
- Sisti, L.; Belcari, J.; Mazzocchetti, L.; Totaro, G.; Vannini, M.; Giorgini, L.; Zucchelli, A.; Celli, A. Multicomponent reinforcing system for poly(butylene succinate): Composites containing poly(l-lactide) electrospun mats loaded with graphene. Polym. Test. 2016, 50, 283–291. [Google Scholar] [CrossRef]
- Bureepukdee, C.; Suttireungwong, S.; Seadan, M. A study on reactive blending of (poly lactic acid) and poly (butylene succinate co adipate). IOP Conf. Ser. Mater. Sci. Eng. 2015, 87, 012070. [Google Scholar] [CrossRef] [Green Version]
- Fei, B.; Chen, C.; Peng, S.; Zhao, X.; Wang, X.; Dong, L. FTIR study of poly(propylene carbonate)/bisphenol A blends. Polym. Int. 2004, 53, 2092–2098. [Google Scholar] [CrossRef]
- Hu, X.; Su, T.; Li, P.; Wang, Z. Blending modification of PBS/PLA and its enzymatic degradation. Polym. Bull. 2017, 75, 533–546. [Google Scholar] [CrossRef]
- Li, X.; Cui, J.; Liu, Y.; Ye, F.; Jin, J.; Xie, X.; Li, A.; Jia, L.; Zhao, Y.; Yang, L. The Effect of Molecular Weight on the Physical Properties and In Vitro Enzymatic Degradation Behavior of Poly(ε-caprolactone). Sci. Adv. Mater. 2019, 11, 1369–1375. [Google Scholar] [CrossRef]
- Williams, D.F. Enzymic Hydrolysis of Polylactic Acid. Eng. Med. 1981, 10, 5–7. [Google Scholar] [CrossRef]
- Ollis, D.L.; Cheah, E.; Cygler, M.; Dijkstra, B.; Frolow, F.; Franken, S.M.; Harel, M.; Remington, S.J.; Silman, I.; Schrag, J. The α/β hydrolase fold. Protein Eng. Des. Sel. 1992, 5, 197–211. [Google Scholar] [CrossRef] [Green Version]
- Ping, L.-F.; Chen, X.-Y.; Yuan, X.-L.; Zhang, M.; Chai, Y.-J.; Shan, S.-D. Application and comparison in biosynthesis and biodegradation by Fusarium solani and Aspergillus fumigatus cutinases. Int. J. Biol. Macromol. 2017, 104, 1238–1245. [Google Scholar] [CrossRef]
- Tserki, V.; Matzinos, P.; Pavlidou, E.; Vachliotis, D.; Panayiotou, C. Biodegradable aliphatic polyesters. Part I. Properties and biodegradation of poly(butylene succinate-co-butylene adipate). Polym. Degrad. Stab. 2006, 91, 367–376. [Google Scholar] [CrossRef]
- Shi, K.; Jing, J.; Song, L.; Su, T.; Wang, Z. Enzymatic hydrolysis of polyester: Degradation of poly(ε-caprolactone) by Candida antarctica lipase and Fusarium solani cutinase. Int. J. Biol. Macromol. 2020, 144, 183–189. [Google Scholar] [CrossRef]
- Marten, E.; Müller, R.-J.; Deckwer, W.-D. Studies on the enzymatic hydrolysis of polyesters I. Low molecular mass model esters and aliphatic polyesters. Polym. Degrad. Stab. 2003, 80, 485–501. [Google Scholar] [CrossRef]
- Fields, R.D.; Rodriguez, F.; Finn, R.K. Microbial degradation of polyesters: Polycaprolactone degraded by P. pullulans. J. Appl. Polym. Sci. 1974, 18, 3571–3579. [Google Scholar] [CrossRef]
- Bikiaris, D.N.; Papageorgiou, G.Z.; Achilias, D.S. Synthesis and comparative biodegradability studies of three poly(alkylene succinate)s. Polym. Degrad. Stab. 2006, 91, 31–43. [Google Scholar] [CrossRef]
- Valverde, C.; Lligadas, G.; Ronda, J.C.; Galià, M.; Cádiz, V. Hydrolytic and enzymatic degradation studies of aliphatic 10-undecenoic acid-based polyesters. Polym. Degrad. Stab. 2018, 155, 84–94. [Google Scholar] [CrossRef]
- Li, F.; Hu, X.; Guo, Z.; Wang, Z.; Wang, Y.; Liu, D.; Xia, H.; Chen, S. Purification and characterization of a novel poly(butylene succinate)-degrading enzyme from Aspergillus sp. XH0501-a. World J. Microbiol. Biotechnol. 2011, 27, 2591–2596. [Google Scholar] [CrossRef]
- Honda, N.; Taniguchi, I.; Miyamoto, M.; Kimura, Y. Reaction Mechanism of Enzymatic Degradation of Poly(butylene succinate-co-terephthalate) (PBST) with a Lipase Originated from Pseudomonas cepacia. Macromol. Biosci. 2003, 3, 189–197. [Google Scholar] [CrossRef]
- Chisholm, M.H.; Navarro-Llobet, D.; Zhou, Z. Poly(propylene carbonate). 1. More about Poly(propylene carbonate) Formed from the Copolymerization of Propylene Oxide and Carbon Dioxide Employing a Zinc Glutarate Catalyst. Macromolecules 2002, 35, 6494–6504. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Supplier | General Screening Conditions | Weight Loss Test Conditions | ||||
|---|---|---|---|---|---|---|---|
| Tested Polymer | pH; T (°C) | Enzyme Concentration (U/mL) | Tested Polymer | pH; T (°C) | Enzyme Concentration (U/mL) | ||
| Amano Lipase from Pseudomonas fluorescens | A | PBS; PBSA; PCL6500D; PCL6800D; PLA; PPC | 8; 40 | 50 | PBSA; PCL6500D | 8; 40 | 10 |
| Lipase from Pseudomonas sp., TYPE XIII | A | PBS; PBSA; PCL6500D; PCL6800D; PLA; PPC | 7; 40 | 10 | PBSA; PCL6500D | 7; 40 | 10 |
| Lipase from Rhizopus niveus | A | PBS; PBSA; PCL6500D; PCL6800D; PLA; PPC | 7; 40 | 100 | Not tested | ||
| Lipase from Rhizopus oryzae (Lipase F-AP 15), LIGHT B | A | PBS; PBSA; PCL6500D; PCL6800D; PLA; PPC | 7; 40 | 100 | PCL6500D | 7; 40 | 10 |
| Lipase B from Candida antarctica; recombinant from Aspergillus oryzae | A | PBS; PBSA; PCL6500D; PCL6800D; PLA; PPC | 7; 40 | 10 | PBSA; PCL6500D; PCL6800D | 7; 40 | 10 |
| Lipase from Candida sp., recombinant from Aspergillus niger (NZ CALB L) | A | PBS; PBSA; PCL6500D; PCL6800D; PLA; PPC | 8; 40 | 100 | PBSA; PCL6500D; PCL6800D | 8; 40 | 10 |
| Lipase from Candida rugosa, Type VII | A | PBS; PBSA; PCL6500D; PCL6800D; PLA; PPC | 7; 40 | 100 | Not tested | ||
| Lipase from Candida rugosa | A | PBS; PBSA; PCL6500D; PCL6800D; PLA; PPC | 8; 40 | 100 | Not tested | ||
| Lipase from Candida cylindracea | A | PBS; PBSA; PCL6500D; PCL6800D; PLA; PPC | 7; 30 | 50 | Not tested | ||
| Lipase from hog pancreas | A | PBS; PBSA; PCL6500D; PCL6800D; PLA; PPC | 8; 40 | 100 | Not tested | ||
| Lipase from Alcaligenes sp. (QLM) | B | PBS; PBSA; PCL6500D; PCL6800D; PLA; PPC | 8; 40 | 100 | PBSA; PCL6500D; PCL6800D | 8; 40 | 10 |
| Cutinase (NZ 51032) | B | PBS; PBSA; PCL6500D; PCL6800D; PLA; PPC | 8; 40 | 100 | PBS; PBSA; PCL6500D; PCL6800D | 8; 40 | 10 |
| Proteinasi K from Tritirachium album | C | PBS; PBSA; PCL6500D; PCL6800D; PLA; PPC | 8; 40 | 100 | Not tested | ||
| GPC | DSC | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Cooling Scan | 2nd Heating Scan | ||||||||
| Sample | Mw (×10−3 g/mol) | PD | Tc (°C) | ΔHc (J/g) | Tg (°C) | Tcc (°C) | ΔHcc (J/g) | Tm (°C) | ΔHm (J/g) |
| PBS | 186 | 2.5 | 63 | 50 | −31 | / | / | 116 | 53 |
| PBSA | 195 | 2.4 | 41 | 38 | −45 | / | / | 87 | 34 |
| PCL6500D | 153 | 1.5 | 21 | 49 | −61 | / | / | 58 | 45 |
| PCL6800D | 216 | 1.5 | 14 | 50 | nd | / | / | 57 | 47 |
| PLA | 202 | 2.0 | / | / | 59 | 116 | 25 | 152 | 24 |
| PPC | 184 | 3.1 | / | / | 17 | / | / | / | / |
| Enzyme | PBS | PBSA | PCL 6500D | PCL 6800D | PLA | PPC |
|---|---|---|---|---|---|---|
| Amano Lipase from Pseudomonas fluorescens | * | 100% (>24 h) | 100% (6–24 h) | * | * | * |
| Lipase from Pseudomonas sp. | * | 100% (4–6 h) | 100% (2–4 h) | * | * | * |
| Lipase from Rhizopus niveus | * | * | * | * | * | * |
| Lipase from Rhizopus oryzae | * | * | 100% (6–24 h) | * | * | * |
| Lipase B from Candida antarctica | * | 100% (>24 h) | 100% (1–2 h) | 100% (6–24 h) | * | * |
| Lipase from Candida sp. (CALB) | * | 100% (6–24 h) | 100% (0.3–1 h) | 100% (2–4 h) | * | * |
| Lipase from Candida rugosa, Type VII | * | * | * | * | * | * |
| Lipase from Candida rugosa | * | * | * | * | * | * |
| Lipase from Candida cylindracea | * | * | * | * | * | * |
| Lipase from hog pancreas | * | * | * | * | * | * |
| Lipase from Alcaligenes sp. (QLM) | * | 100% (1–2 h) | 100% (1–2 h) | 100 (6–24 h) | * | * |
| Cutinase | 100% (2–4 h) | 100% (1–2 h) | 100% (1–2 h) | 100% (4–6 h) | * | * |
| Proteinasi K | * | * | * | * | * | * |
| Enzyme (10 U/mL) | PBS (mg/h/cm2) | PBSA (mg/h/cm2) | PCL6500D (mg/h/cm2) | PCL6800D (mg/h/cm2) |
|---|---|---|---|---|
| Lipase from Pseudomonas fluorescens | Not tested | 0.10 ± 0.01 | 0.54 ± 0.01 | Not tested |
| Lipase from Pseudomonas sp. | Not tested | 1.91 | 3.13 | Not tested |
| Lipase B from Candida antarctica | Not tested | 1.17 | 3.52 | 1.41 |
| Lipase from Candida sp. (CALB) | Not tested | 0.00 ± 0.00 | 5.17 ± 0.51 | 4.78 ± 0.30 |
| Lipase from Alcaligenes sp. (QLM) | Not tested | 0.16 ± 0.00 | 0.72 ± 0.02 | 0.33 ± 0.03 |
| Cutinase | 0.35 ± 0.01 | 6.38 ± 0.66 | 4.57 ± 0.05 | 1.96 ± 0.15 |
| 1H NMR | GPC | ||||||
|---|---|---|---|---|---|---|---|
| Sample | Enzyme | Time (h) | Wt Loss (%) | OH End-Groups (mol%) | Mn (×10−3 g/mol) | Mw (×10−3 g/mol) | PD |
| PBS | Control | 30 | / | 1.1 | 74 | 186 | 2.5 |
| Cutinase | 1 | 16 | 1.7 | 73 | 178 | 2.4 | |
| 2 | 30 | 1.5 | 70 | 173 | 2.5 | ||
| 4 | 63 | 1.8 | 68 | 176 | 2.6 | ||
| PBSA | Control | 30 | / | 1.1 | 81 | 195 | 2.4 |
| Cutinase | 0.5 | 45 | 1.6 | 67 | 164 | 2.5 | |
| 1 | 80 | 2.2 | 72 | 181 | 2.6 | ||
| Lipase from P. fluorescens | 15 | 11 | 1.6 | 62 | 163 | 2.6 | |
| 25 | 24 | 1.9 | 61 | 157 | 2.6 | ||
| 30 | 29 | 1.8 | 61 | 155 | 2.5 | ||
| Lipase from Alcaligenes sp. (QLM) | 10 | 29 | 1.6 | 63 | 160 | 2.6 | |
| 20 | 45 | 2.1 | 62 | 162 | 2.6 | ||
| 30 | 61 | 1.9 | 60 | 164 | 2.7 | ||
| Lipase from Pseudomonas sp. | 1 | 60 | na | 64 | 160 | 2.5 | |
| 2 | 94 | na | 61 | 152 | 2.5 | ||
| Lipase B from C. antarctica | 3 | 68 | na | 64 | 164 | 2.6 | |
| 4 | 78 | na | 62 | 152 | 2.5 | ||
| 5 | 93 | na | 57 | 155 | 2.7 | ||
| PCL 6500D | Control | 30 | / | 0.5 | 102 | 153 | 1.5 |
| Cutinase | 0.5 | 23 | 0.8 | 87 | 133 | 1.6 | |
| 1 | 48 | 1.2 | 90 | 141 | 1.6 | ||
| 1.5 | 68 | 1.6 | 81 | 130 | 1.6 | ||
| Lipase from Candida sp. (CALB) | 0.3 | 21 | 0.6 | 95 | 146 | 1.5 | |
| 0.8 | 53 | 0.6 | 91 | 138 | 1.5 | ||
| 1.3 | 74 | 1.2 | 90 | 140 | 1.6 | ||
| Lipase from P. fluorescens | 3 | 16 | 0.6 | 94 | 146 | 1.5 | |
| 6 | 32 | 1.4 | 87 | 138 | 1.6 | ||
| 9 | 58 | 1.3 | 88 | 142 | 1.6 | ||
| Lipase from Alcaligenes sp. (QLM) | 3 | 23 | 1.2 | 80 | 122 | 1.5 | |
| 6 | 45 | 1.4 | 78 | 123 | 1.6 | ||
| 9 | 66 | 1.7 | 78 | 118 | 1.5 | ||
| Lipase from Pseudomonas sp. | 0.5 | 27 | na | 79 | 125 | 1.6 | |
| 1.5 | 57 | na | 80 | 124 | 1.5 | ||
| 2 | 79 | na | 80 | 123 | 1.5 | ||
| Lipase B from C. antarctica | 0.5 | 23 | na | 77 | 119 | 1.5 | |
| 0.8 | 40 | na | 85 | 129 | 1.5 | ||
| 1 | 55 | na | 82 | 127 | 1.5 | ||
| 2 | 92 | na | 75 | 118 | 1.6 | ||
| PCL 6800D | Control | 30 | / | 1.0 | 120 | 197 | 1.64 |
| Cutinase | 1 | 24 | 1.3 | 115 | 189 | 1.64 | |
| 3 | 61 | 1.0 | 114 | 185 | 1.63 | ||
| 4 | 89 | 1.2 | 121 | 197 | 1.63 | ||
| Lipase from Candida sp. (CALB) | 0.8 | 32 | 1.4 | 110 | 180 | 1.63 | |
| 1.8 | 67 | 1.4 | 115 | 189 | 1.65 | ||
| 2 | 88 | 1.2 | 123 | 188 | 1.53 | ||
| Lipase from Alcaligenes sp. (QLM) | 4 | 14 | 1.2 | 102 | 170 | 1.67 | |
| 8 | 32 | 1.2 | 102 | 172 | 1.68 | ||
| 15 | 65 | 1.2 | 107 | 183 | 1.71 | ||
| Lipase B from C. antarctica | 2 | 23 | na | 99 | 172 | 1.74 | |
| 4 | 66 | na | 91 | 153 | 1.69 | ||
| 6 | 93 | na | 94 | 161 | 1.72 | ||
| Polymer | Enzyme | GPC | 1H NMR | Prevalent Cleaving Action Mode * | |||
|---|---|---|---|---|---|---|---|
| Time (h) | Wt. Loss (%) | Mw (g/mol) | PD | Notes | |||
| PBS | Cutinase | 4 | 63 | 383 | 1.2 | na | Endo-type scission |
| PBSA | Cutinase | 1.3 | 99 | na | na | Oligomers traces + BD | Exo-type scission |
| Lipase from Pseudomonas fluorescens | 30 | 29 | 484 | 1.3 | Oligomers + BD BA/BS ↑ | Endo-type scission | |
| Lipase from Alcaligenes sp. (QLM) | 30 | 61 | 646 | 1.4 | Oligomers + BD BA/BS ↑ | Endo-type scission | |
| PCL6500D | Cutinase | 2.5 | 98 | 431 | 1.3 | HA | Exo-type scission |
| Lipase from Candida sp. (CALB) | 1.8 | 93 | 428 | 1.3 | HA | Exo-type scission | |
| Lipase from Pseudomonas fluorescens | 9 | 59 | 306 | 1.1 | Oligomers + HA | Endo-type scission | |
| Lipase from Alcaligenes sp. (QLM) | 9 | 66 | 297 | 1.2 | Oligomers + HA | Endo-type scission | |
| PCL6800D | Cutinase | 4.5 | 100 | 370 | 1.2 | Oligomers traces + HA | Exo-type scission |
| Lipase from Candida sp. (CALB) | 2.3 | 93 | 982 | 1.5 | Oligomers + HA | Exo-type scission | |
| Lipase from Alcaligenes sp. (QLM) | 23 | 96 | 256 | 1.5 | Oligomers +HA | Endo-type scission | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosato, A.; Romano, A.; Totaro, G.; Celli, A.; Fava, F.; Zanaroli, G.; Sisti, L. Enzymatic Degradation of the Most Common Aliphatic Bio-Polyesters and Evaluation of the Mechanisms Involved: An Extended Study. Polymers 2022, 14, 1850. https://doi.org/10.3390/polym14091850
Rosato A, Romano A, Totaro G, Celli A, Fava F, Zanaroli G, Sisti L. Enzymatic Degradation of the Most Common Aliphatic Bio-Polyesters and Evaluation of the Mechanisms Involved: An Extended Study. Polymers. 2022; 14(9):1850. https://doi.org/10.3390/polym14091850
Chicago/Turabian StyleRosato, Antonella, Angela Romano, Grazia Totaro, Annamaria Celli, Fabio Fava, Giulio Zanaroli, and Laura Sisti. 2022. "Enzymatic Degradation of the Most Common Aliphatic Bio-Polyesters and Evaluation of the Mechanisms Involved: An Extended Study" Polymers 14, no. 9: 1850. https://doi.org/10.3390/polym14091850