
Reinforced Bioremediation of Excessive Nitrate in Atrazine-Contaminated Soil by Biodegradable Composite Carbon Source


Abstract
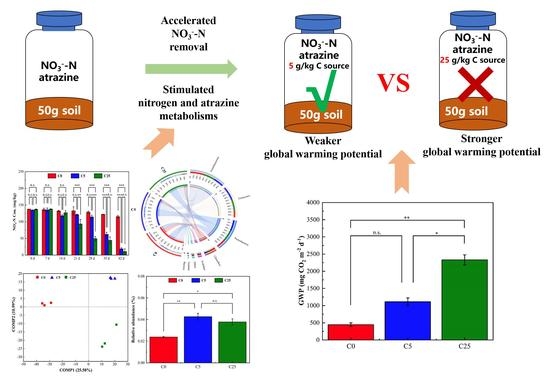
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Soil Sample Collection and Preparation of Composite Carbon Source
2.2. Experimental Design
2.3. Soil DNA Extraction and Illumina MiSeq Sequencing Analysis
2.4. Statistical Analysis
3. Results and Discussion
3.1. Effect of Composite Carbon Source on Soil Denitrification Performance
3.2. Changes of pH and Dissolved Organic Carbon in Different Treatments
3.3. Greenhouse Gas Emissions and Their Global Warming Potential
3.4. Microbial Community Structure
3.4.1. Composition and Diversity of Soil Bacterial and Fungal Community
3.4.2. Effect of Composite Carbon Source on Microbial Functions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castaldelli, G.; Colombani, N.; Soana, E.; Vincenzi, F.; Fano, E.A.; Mastrocicco, M. Reactive nitrogen losses via denitrification assessed in saturated agricultural soils. Geoderma 2019, 337, 91–98. [Google Scholar] [CrossRef]
- Zhang, Z.; Sun, D.; Tang, Y.; Zhu, R.; Li, X.; Gruda, N.; Dong, J.; Duan, Z. Plastic shed soil salinity in China: Current status and next steps. J. Clean. Prod. 2021, 296, 126453. [Google Scholar] [CrossRef]
- Wang, J.; Chu, L. Biological nitrate removal from water and wastewater by solid-phase denitrification process. Biotechnol. Adv. 2016, 34, 1103–1112. [Google Scholar] [CrossRef]
- Dong, L.F.; Smith, C.J.; Papaspyrou, S.; Stott, A.; Osborn, A.M.; Nedwell, D.B. Changes in benthic denitrification, nitrate ammonification, and anammox process rates and nitrate and nitrite reductase gene abundances along an estuarine nutrient gradient (the Colne Estuary, United Kingdom). Appl. Environ. Microbiol. 2009, 75, 3171–3179. [Google Scholar] [CrossRef]
- López-Gutiérrez, J.C.; Henry, S.; Hallet, S.; Martin-Laurent, F.; Catroux, G.; Philippot, L. Quantification of a novel group of nitrate-reducing bacteria in the environment by real-time PCR. J. Microbiol. Methods 2004, 57, 399–407. [Google Scholar] [CrossRef]
- Minick, K.; Pandey, C.; Fox, T.; Subedi, S. Dissimilatory nitrate reduction to ammonium and N2O flux: Effect of soil redox potential and N fertilization in loblolly pine forests. Biol. Fertil. Soils 2016, 52, 601–614. [Google Scholar] [CrossRef]
- Li, H.; Dai, M.; Dai, S.; Dong, X. Current status and environment impact of direct straw return in China’s cropland–A review. Ecotoxicol. Environ. Saf. 2018, 159, 293–300. [Google Scholar] [CrossRef]
- Yang, Z.; Lou, Y.; Pan, H.; Wang, H.; Yang, Q.; Zhuge, Y.; Hu, J. Improved Denitrification Performance of Polybutylene Succinate/Corncob Composite Carbon Source by Proper Pretreatment: Performance, Functional Genes and Microbial Community Structure. Polymers 2023, 15, 801. [Google Scholar] [CrossRef]
- Azam, S.R.; Ma, H.; Xu, B.; Devi, S.; Siddique, M.A.B.; Stanley, S.L.; Bhandari, B.; Zhu, J. Efficacy of ultrasound treatment in the removal of pesticide residues from fresh vegetables: A review. Trends Food Sci. Technol. 2020, 97, 417–432. [Google Scholar] [CrossRef]
- Pirsaheb, M.; Moradi, N. Sonochemical degradation of pesticides in aqueous solution: Investigation on the influence of operating parameters and degradation pathway-a systematic review. RSC Adv. 2020, 10, 7396–7423. [Google Scholar] [CrossRef]
- Rani, L.; Thapa, K.; Kanojia, N.; Sharma, N.; Singh, S.; Grewal, A.S.; Srivastav, A.L.; Kaushal, J. An extensive review on the consequences of chemical pesticides on human health and environment. J. Clean. Prod. 2021, 283, 124657. [Google Scholar] [CrossRef]
- Liu, Y.; Fan, X.; Zhang, T.; He, W.; Song, F. Effects of the long-term application of atrazine on soil enzyme activity and bacterial community structure in farmlands in China. Environ. Pollut. 2020, 262, 114264. [Google Scholar] [CrossRef] [PubMed]
- Zaya, R.M.; Amini, Z.; Whitaker, A.S.; Kohler, S.L.; Ide, C.F. Atrazine exposure affects growth, body condition and liver health in Xenopus laevis tadpoles. Aquat. Toxicol. 2011, 104, 243–253. [Google Scholar] [CrossRef]
- Banks, M.; Kennedy, A.; Kremer, R.; Eivazi, F. Soil microbial community response to surfactants and herbicides in two soils. Appl. Soil Ecol. 2014, 74, 12–20. [Google Scholar] [CrossRef]
- Ren, X.; Zeng, G.; Tang, L.; Wang, J.; Wan, J.; Wang, J.; Deng, Y.; Liu, Y.; Peng, B. The potential impact on the biodegradation of organic pollutants from composting technology for soil remediation. Waste Manag. 2018, 72, 138–149. [Google Scholar] [CrossRef]
- Chen, Y.; Jiang, Z.; Wu, D.; Wang, H.; Li, J.; Bi, M.; Zhang, Y. Development of a novel bio-organic fertilizer for the removal of atrazine in soil. J. Environ. Manag. 2019, 233, 553–560. [Google Scholar] [CrossRef]
- Raimondo, E.E.; Aparicio, J.D.; Bigliardo, A.L.; Fuentes, M.S.; Benimeli, C.S. Enhanced bioremediation of lindane-contaminated soils through microbial bioaugmentation assisted by biostimulation with sugarcane filter cake. Ecotoxicol. Environ. Saf. 2020, 190, 110143. [Google Scholar] [CrossRef]
- Fang, H.; Lian, J.; Wang, H.; Cai, L.; Yu, Y. Exploring bacterial community structure and function associated with atrazine biodegradation in repeatedly treated soils. J. Hazard Mater. 2015, 286, 457–465. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G. PICRUSt2: An improved and extensible approach for metagenome inference. bioRxiv 2019. [Google Scholar] [CrossRef]
- Matias, T.P.; Braga, J.K.; Brucha, G. Anaerobic biodegradation of atrazine under different redox conditions. Int. J. Eng. Sci. 2019, 6, 227–236. [Google Scholar] [CrossRef]
- Li, Z.; Tang, Z.; Song, Z.; Chen, W.; Tian, D.; Tang, S.; Wang, X.; Wang, J.; Liu, W.; Wang, Y. Variations and controlling factors of soil denitrification rate. Glob. Chang. Biol. 2022, 28, 2133–2145. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; McCarty, G.W.; Lang, M.; Ducey, T.; Hunt, P.; Miller, J. Topographic and physicochemical controls on soil denitrification in prior converted croplands located on the Delmarva Peninsula, USA. Geoderma 2018, 309, 41–49. [Google Scholar] [CrossRef]
- Pan, F.; Chapman, S.J.; Li, Y.; Yao, H. Straw amendment to paddy soil stimulates denitrification but biochar amendment promotes anaerobic ammonia oxidation. J. Soils Sediments 2017, 17, 2428–2437. [Google Scholar] [CrossRef]
- Kabra, A.N.; Ji, M.-K.; Choi, J.; Kim, J.R.; Govindwar, S.P.; Jeon, B.-H. Toxicity of atrazine and its bioaccumulation and biodegradation in a green microalga, Chlamydomonas mexicana. Environ. Sci. Pollut. Res. 2014, 21, 12270–12278. [Google Scholar] [CrossRef]
- Kourtev, P.S.; Nakatsu, C.H.; Konopka, A. Responses of the anaerobic bacterial community to addition of organic C in chromium (VI)-and iron (III)-amended microcosms. Appl. Environ. Microbiol. 2006, 72, 628–637. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Zhen, Z.; Liang, Y.; Li, J.; Yang, J.; Zhong, L.; Zhao, L.; Li, Y.; Luo, C.; Ren, L. Changes in atrazine speciation and the degradation pathway in red soil during the vermiremediation process. J. Hazard Mater. 2019, 364, 710–719. [Google Scholar] [CrossRef]
- Ferri, M.V.W.; Gomes, J.; Dick, D.P.; Souza, R.F.d.; Vidal, R.A. Sorption of acetochlor herbicide by soil samples, humic acids and humin from an argisol under no-till and conventional tillage systems. Rev. Bras. Cienc. Solo 2005, 29, 705–714. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, N. Atrazine and its metabolites degradation in mineral salts medium and soil using an enrichment culture. Environ. Monit. Assess. 2016, 188, 142. [Google Scholar] [CrossRef]
- Ghafari, S.; Hasan, M.; Aroua, M.K. Bio-electrochemical removal of nitrate from water and wastewater—A review. Bioresour. Technol. 2008, 99, 3965–3974. [Google Scholar] [CrossRef]
- Takeshita, V.; Mendes, K.F.; Alonso, F.G.; Tornisielo, V.L. Effect of organic matter on the behavior and control effectiveness of herbicides in soil. Planta Daninha 2019, 37, 1–17. [Google Scholar] [CrossRef]
- Tejada, M.; Benítez, C. Flazasulfuron behavior in a soil amended with different organic wastes. Appl. Soil Ecol. 2017, 117, 81–87. [Google Scholar] [CrossRef]
- Le Mer, J.; Roger, P. Production, oxidation, emission and consumption of methane by soils: A review. Eur. J. Soil Biol. 2001, 37, 25–50. [Google Scholar] [CrossRef]
- Kuypers, M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef]
- Lee, Y.-Y.; Choi, H.; Cho, K.-S. Effects of carbon source, C/N ratio, nitrate, temperature, and pH on N2O emission and functional denitrifying genes during heterotrophic denitrification. J. Environ. Sci. Health A 2019, 54, 16–29. [Google Scholar] [CrossRef]
- Hayakawa, A.; Akiyama, H.; Sudo, S.; Yagi, K. N2O and NO emissions from an Andisol field as influenced by pelleted poultry manure. Soil Biol. Biochem. 2009, 41, 521–529. [Google Scholar] [CrossRef]
- Qiu, Q.; Wu, L.; Ouyang, Z.; Li, B.; Xu, Y.; Wu, S.; Gregorich, E. Effects of plant-derived dissolved organic matter (DOM) on soil CO2 and N2O emissions and soil carbon and nitrogen sequestrations. Appl. Soil Ecol. 2015, 96, 122–130. [Google Scholar] [CrossRef]
- Paterson, E.; Sim, A. Soil-specific response functions of organic matter mineralization to the availability of labile carbon. Glob. Chang. Biol. 2013, 19, 1562–1571. [Google Scholar] [CrossRef]
- Wang, N.; Yu, J.-G.; Zhao, Y.-H.; Chang, Z.-Z.; Shi, X.-X.; Ma, L.Q.; Li, H.-B. Straw enhanced CO2 and CH4 but decreased N2O emissions from flooded paddy soils: Changes in microbial community compositions. Atmos. Environ. 2018, 174, 171–179. [Google Scholar] [CrossRef]
- Thangarajan, R.; Bolan, N.S.; Tian, G.; Naidu, R.; Kunhikrishnan, A. Role of organic amendment application on greenhouse gas emission from soil. Sci. Total Environ. 2013, 465, 72–96. [Google Scholar] [CrossRef]
- Cui, Y.-F.; Jun, M.; Wang, Q.-X.; Zhang, W.-M.; Cheng, X.-Y.; Chen, W.-F. Effects of straw and biochar addition on soil nitrogen, carbon, and super rice yield in cold waterlogged paddy soils of North China. J. Integr. Agric. 2017, 16, 1064–1074. [Google Scholar] [CrossRef]
- Yang, J.; Guo, W.; Wang, F.; Wang, F.; Zhang, L.; Zhou, B.; Xing, S.; Yang, W. Dynamics and influencing factors of soluble organic nitrogen in paddy soil under different long-term fertilization treatments. Soil Tillage Res. 2021, 212, 105077. [Google Scholar] [CrossRef]
- Banerjee, S.; Kirkby, C.A.; Schmutter, D.; Bissett, A.; Kirkegaard, J.A.; Richardson, A.E. Network analysis reveals functional redundancy and keystone taxa amongst bacterial and fungal communities during organic matter decomposition in an arable soil. Soil Biol. Biochem. 2016, 97, 188–198. [Google Scholar] [CrossRef]
- Li, X.; Wu, S.; Fan, H.; Dong, Y.; Wang, Y.; Bai, Z.; Jing, C.; Zhuang, X. Phylogenetic distance affects the artificial microbial consortia’s effectiveness and colonization during the bioremediation of polluted soil with Cr (VI) and atrazine. J. Hazard Mater. 2023, 454, 131460. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yang, B.; Wang, H.; He, F.; Gao, Y.; Scheel, R.A. Soil microbial community toxic response to atrazine and its residues under atrazine and lead contamination. Environ. Sci. Pollut. Res. 2015, 22, 996–1007. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Lou, Y.; Pan, H.; Wang, H.; Yang, Q.; Sun, Y.; Zhuge, Y. Reinforced Bioremediation of Excessive Nitrate in Atrazine-Contaminated Soil by Biodegradable Composite Carbon Source. Polymers 2023, 15, 2765. https://doi.org/10.3390/polym15132765
Yang Z, Lou Y, Pan H, Wang H, Yang Q, Sun Y, Zhuge Y. Reinforced Bioremediation of Excessive Nitrate in Atrazine-Contaminated Soil by Biodegradable Composite Carbon Source. Polymers. 2023; 15(13):2765. https://doi.org/10.3390/polym15132765
Chicago/Turabian StyleYang, Zhongchen, Yanhong Lou, Hong Pan, Hui Wang, Quangang Yang, Yajie Sun, and Yuping Zhuge. 2023. "Reinforced Bioremediation of Excessive Nitrate in Atrazine-Contaminated Soil by Biodegradable Composite Carbon Source" Polymers 15, no. 13: 2765. https://doi.org/10.3390/polym15132765
APA StyleYang, Z., Lou, Y., Pan, H., Wang, H., Yang, Q., Sun, Y., & Zhuge, Y. (2023). Reinforced Bioremediation of Excessive Nitrate in Atrazine-Contaminated Soil by Biodegradable Composite Carbon Source. Polymers, 15(13), 2765. https://doi.org/10.3390/polym15132765