
Polyhydroxybutyrate Metabolism in Azospirillum brasilense and Its Applications, a Review
 ,
,
Abstract
:1. Introduction
2. The Role of the Carbon Source in PHB Production by A. brasilense
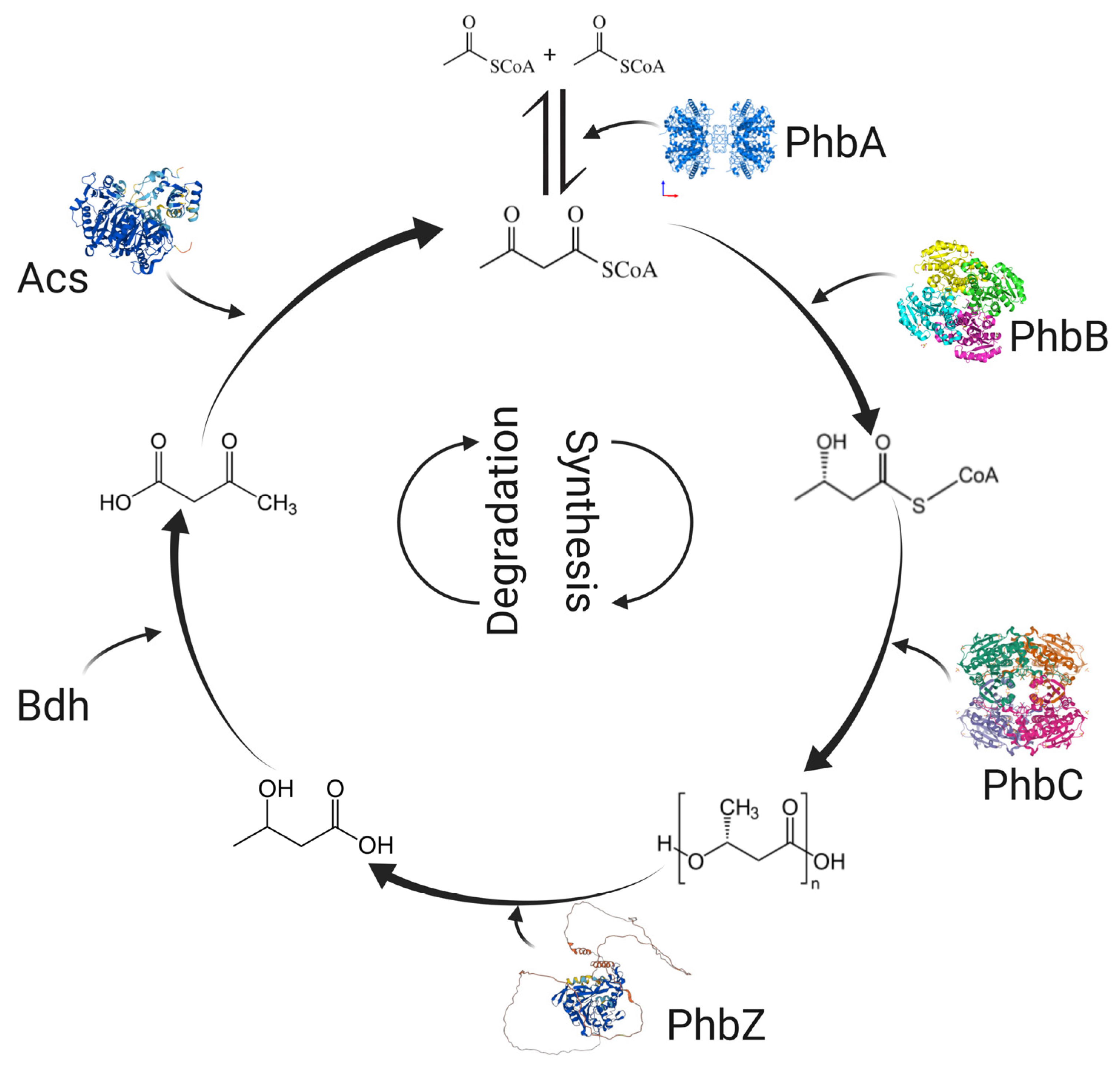
3. PHB Synthesis and Degradation by A. brasilense
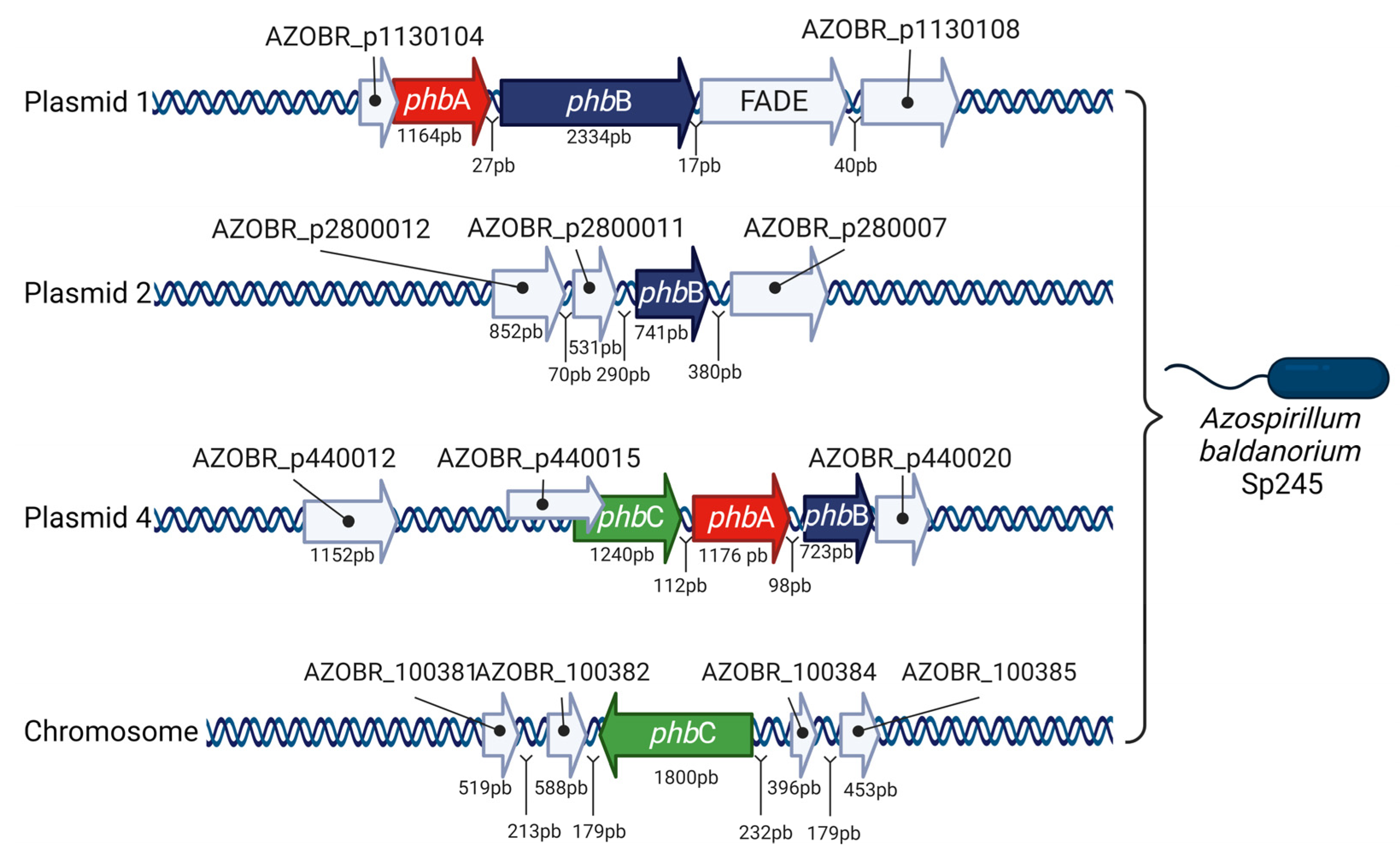
4. Studies on PHB Synthesis and Degradation Genes
5. PHB Metabolism Regulation in A. brasilense Sp7
6. Nitrogen’s Role in PHB Metabolism
7. Flocculation and Cyst Involvement in PHB Production, the Role of Oxygen
8. PHB and Biofilm in A. brasilense
9. Functions of PHB in Azospirillum brasilense
10. PHB Properties and Applications
11. PHB Biodegradability
12. Future Outlooks
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Pereg, L. Azospirillum Cell Aggregation, Attachment, and Plant Interaction. In Handbook for Azospirillum Technical Issues and Protocols, 1st ed.; Cassán, F.D., Okon, Y., Creus, C.M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; Volume 1, pp. 181–197. [Google Scholar] [CrossRef]
- Martínez-Martínez, M.D.L.A.; González-Pedrajo, B.; Dreyfus, G.; Soto-Urzúa, L.; Martínez-Morales, L.J. Phasin PhaP1 is involved in polyhydroxybutyrate granules morphology and in controlling early biopolymer accumulation in Azospirillum brasilense Sp7. AMB Express 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volova, T.G.; Boyandin, A.N.; Vasiliev, A.D.; Karpov, V.A.; Prudnikova, S.V.; Mishukova, O.V.; Boyarskikh, U.A.; Filipenko, M.L.; Rudnev, V.P.; Xuân, B.B.; et al. Biodegradation of polyhydroxyalkanoates (PHAs) in tropical coastal waters and identification of PHA-degrading bacteria. Polym. Degrad. Stab. 2010, 95, 2350–2359. [Google Scholar] [CrossRef]
- Halevas, E.G.; Pantazaki, A.A. Polyhydroxyalkanoates: Chemical structure. In Polyhydroxyalkanoates: Biosynthesis, Chemical Structure and Applications, 1st ed.; Williams, H., Kelly, P., Eds.; Nova Science Publishers Inc.: New York, NY, USA, 2018; Volume 1, pp. 133–166. [Google Scholar]
- Itzigsohn, R.; Yarden, O.; Okon, Y. Polyhydroxyalkanoate analysis in Azospirillum brasilense. Can. J. Micrcobiol. 1995, 41, 73–76. [Google Scholar] [CrossRef]
- Patiño, I.M.E.; Soto, U.L.; Orea, F.M.L.; López, V.D.; Martínez-Morales, L.J. Extraction and NMR determinantion of PHB from Azospirillum brasilense Sp7. JCBPS Spec. Issue 2014, 4, 26–32. [Google Scholar]
- Kamnev, A.A.; Antonyuk, L.P.; Tugarova, A.V.; Tarantilis, P.A.; Polissiou, M.G.; Gardiner, P.H.E. Fourier transform infrared spectroscopic characterisation of heavy metal-induced metabolic changes in the plant-associated soil bacterium Azospirillum brasilense Sp7. J. Mol. Struct. 2002, 610, 127–131. [Google Scholar] [CrossRef]
- Tal, S.; Okon, Y. Production of the reserve material poly-β-hydroxybutyrate and its function in Azospirillum brasilense Cd. Can. J. Microbiol. 1985, 31, 608–613. [Google Scholar] [CrossRef]
- Tal, S.; Smirnoff, P.; Okon, Y. The regulation of poly-β-hydroxybutyrate metabolism in Azospirillum brasilense during balanced growth and starvation. Microbiology 1990, 136, 1191–1196. [Google Scholar] [CrossRef] [Green Version]
- Westby, C.A.; Cutshall, D.S.; Vigil, G.V. Metabolism of various carbon sources by Azospirillum brasilense. J. Bacteriol. 1983, 156, 1369–1372. [Google Scholar] [CrossRef] [Green Version]
- Sadasivan, L.; Neyra, C.A. Flocculation in Azospirillum brasilense and Azospirillum lipoferum: Exopolysaccharides and cyst formation. J. Bacteriol. 1985, 163, 716–723. [Google Scholar] [CrossRef] [Green Version]
- Loh, W.H.; Randles, C.I.; Sharp, W.R.; Miller, R.H. Intermediary carbon metabolism of Azospirillum brasilense. J. Bacteriol. 1984, 158, 264–268. [Google Scholar] [CrossRef] [Green Version]
- Alexandre, G.; Greer, S.E.; Zhulin, I.B. Energy taxis is the dominant behavior in Azospirillum brasilense. J. Bacteriol. 2000, 182, 6042–6048. [Google Scholar] [CrossRef] [Green Version]
- Okon, Y.; Albrecht, S.L.; Burris, R.H. Factors affecting growth and nitrogen fixation of Spirillum lipoferum. J. Bacteriol. 1976, 127, 1248–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neyra, C.A.; Döbereiner, J.; Lalande, R.; Knowles, R. Denitrification by N2-fixing Spirillum lipoferum. Can. J. Microbiol. 1977, 23, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Drets, G.; Del Gallo, M.; Burpee, C.; Burris, R.H. Catabolism of carbohydrates and organic acids and expression of nitrogenase by Azospirilla. J. Bacteriol. 1984, 159, 80–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tal, S.; Smirnoff, P.; Okon, Y. Purification and characterization of d-β-hydroxybutyrate dehydrogenase from Azospirillum brasilense Cd. Microbiology 1990, 136, 645–649. [Google Scholar] [CrossRef] [Green Version]
- Okon, Y.; Itzigsohn, R. Poly-β-hydroxybutyrate metabolism in Azospirillum brasilense and the ecological role of PHB in the rhizosphere. FEMS Microbiol. 1992, 9, 131–139. [Google Scholar] [CrossRef]
- Manna, A.; Pal, S.; Paul, A.K. Occurrence of poly-3-hydroxybutyrate in Azospirillum sp. Folia Microbiol. 1997, 42, 629–634. [Google Scholar] [CrossRef]
- Narancic, T.; Scollica, E.; Kenny, S.T.; Gibbons, H.; Carr, E.; Brennan, L.; Cafney, G.; Wynne, K.; Murphy, C.; Raberg, M.; et al. Understanding the physiological roles of polyhydroxybutyrate (PHB) in Rhodospirillum rubrum S1 under aerobic chemoheterotrophic conditions. Appl. Microbiol. Biotechnol. 2016, 100, 8901–8912. [Google Scholar] [CrossRef]
- Prieto, A.; Escapa, I.F.; Martínez, V.; Dinjaski, N.; Herencias, C.; de la Peña, F.; Tarazona, N.; Revelles, O. A holistic view of polyhydroxyalkanoate metabolism in Pseudomonas putida. Environ. Microbiol. 2016, 18, 341–357. [Google Scholar] [CrossRef] [Green Version]
- Ochsner, A.M.; Sonntag, F.; Buchhaupt, M.; Schrader, J.; Vorholt, J.J. Methylobacterium extorquens: Methylotrophy and biotechnological applications. Appl. Microbiol. Biotechnol. 2015, 99, 517–534. [Google Scholar] [CrossRef]
- Quelas, J.I.; Mesa, S.; Mongiardini, E.J.; Jendrossek, D.; Lodeiro, A.R. Regulation of polyhydroxybutyrate synthesis in the soil bacterium Bradyrhizobium diazoefficiens. Appl. Environ. Microbiol. 2016, 82, 4299–4308. [Google Scholar] [CrossRef] [Green Version]
- Sreedevi, S.; Unni, K.N.; Sajith, S.; Priji, P.; Josh, M.S.; Benjamin, S. Bioplastics: Advances in polyhydroxybutyrate research. In Advances in Polymer Science; Abe, A., Albertsson, A.C., Coates, G.W., Genzer, J., Kobayashi, S., Lee, K.S., Leibler, L., Long, T.E., Möller, M., Okay, O., et al., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; Volume 1, pp. 1–30. [Google Scholar] [CrossRef]
- Soto, L.R.; Byrne, E.; Van Niel, E.W.; Sayed, M.; Villanueva, C.C.; Hatti-Kaul, R. Hydrogen and polyhydroxybutyrate production from wheat straw hydrolysate using Caldicellulosiruptor species and Ralstonia eutropha in a coupled process. Bioresour. Technol. 2009, 272, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Ayub, N.D.; Pettinari, M.J.; Ruiz, J.A.; López, N.I. A polyhydroxybutyrate-producing Pseudomonas sp. isolated from Antarctic environments with high stress resistance. Curr. Microbiol. 2004, 49, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Ayub, N.D.; Pettinari, M.J.; Méndez, B.S.; López, N.I. Impaired polyhydroxybutyrate biosynthesis from glucose in Pseudomonas sp. 14-3 is due to a defective beta-ketothiolase gene. FEMS Microbiol. Lett. 2006, 264, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghoddosi, F.; Golzar, H.; Yazdian, F.; Khosravi-Darani, K.; Vasheghani-Farahani, E. Effect of carbon sources for PHB production in bubble column bioreactor: Emphasis on improvement of methane uptake. J. Environ. Chem. Eng. 2019, 7, 102978. [Google Scholar] [CrossRef]
- Manju, J.; Prabakaran, P. Effect of carbon sources in the production of polyhydroxybutyrate (PHB) by Bradyrhizobium and Rhizobium sp. from Aeschynomene indica. Int. J. Res. Anal. Rev. 2019, 6, 823–827. [Google Scholar]
- Torres-Pedraza, A.J.; Salgado-Lugo, H.; Segura, D.; Díaz-Barrera, A.; Peña, C. Composition control of poly (3-hydroxybutyrate-co-3-hydroxyvalerate) copolymerization by oxygen transfer rate (OTR) in Azotobacter vinelandii OPNA. J. Chem. Technol. Biotechnol. 2021, 96, 2782–2791. [Google Scholar] [CrossRef]
- Hassan, M.A.; Bakhiet, E.K.; Hussein, H.R.; Ali, S.G. Statistical optimization studies for polyhydroxybutyrate (PHB) production by novel Bacillus subtilis using agricultural and industrial wastes. Int. J. Environ. Sci. Technol. 2019, 16, 3497–3512. [Google Scholar] [CrossRef]
- Kadouri, D.; Burdman, S.; Jurkevitch, E.; Okon, Y. Identification and isolation of genes involved in poly (β-hydroxybutyrate) biosynthesis in Azospirillum brasilense and characterization of a phbC mutant. Appl. Environ. Microbiol. 2002, 68, 2943–2949. [Google Scholar] [CrossRef] [Green Version]
- Kadouri, D.; Jurkevitch, E.; Okon, Y.; Castro-Sowinski, S. Ecological and Agricultural Significance of Bacterial Polyhydroxyalkanoates. Crit. Rev. Microbiol. 2005, 31, 55–67. [Google Scholar] [CrossRef]
- Senior, P.J.; Dawes, E.A. The regulation of poly-β-hydroxybutyrate metabolism in Azotobacter beijerinckii. Biochem. J. 1973, 134, 225–238. [Google Scholar] [CrossRef]
- Volova, T.G.; Kalacheva, G.S.; Gorbunova, O.V.; Zhila, N.O. Dynamics of Activity of the Key Enzymes of Polyhydroxyalkanoate Metabolism in Ralstonia eutropha. Appl. Biochem. Microbiol. 2004, 40, 170–177. [Google Scholar] [CrossRef]
- Trainer, M.A.; Charles, T.C. The role of PHB metabolism in the symbiosis of rhizobia with legumes. Appl. Microbiol. Biotechnol. 2006, 71, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Edelshtein, Z.; Kadouri, D.; Jurkevitch, E.; Vande Broek, A.; Vanderleyden, J.; Okon, Y. Characterization of genes involved in poly-β-hydroxybutyrate metabolism in Azospirillum brasilense. Symbiosis 2003, 34, 157–170. [Google Scholar]
- Kadouri, D.; Jurkevitch, E.; Okon, Y. Involvement of the reserve material poly-β-hydroxybutyrate in Azospirillum brasilense stress endurance and root colonization. Appl. Environ. Microbiol. 2003, 69, 3244–3250. [Google Scholar] [CrossRef] [Green Version]
- Müller-Santos, M.; Maltempi de Souza, E.; de Oliveira-Pedrosa, F.; Chubatsu, L.S. Polyhydroxybutyrate in Azospirillum brasilense. In Handbook for Azospirillum Technical Issues and Protocols, 1st ed.; Cassán, F.D., Okon, Y., Creus, C.M., Eds.; Springer International Publising: Cham, Switzerland, 2015; Volume 1, pp. 241–250. [Google Scholar] [CrossRef]
- Jendrossek, D. Polyhydroxyalkanoate granules are complex subcellular organelles (carbonosomes). J. Bacteriol. 2009, 191, 3195–3202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bresan, S.; Sznajder, A.; Hauf, W.; Forchhammer, K.; Pfeiffer, D.; Jendrossek, D. Polyhydroxyalkanoate (PHA) granules have no phospholipids. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Bresan, S.; Jendrossek, D. New Insights into PhaM-PhaC-Mediated Localization of Polyhydroxybutyrate Granules in Ralstonia eutropha H16. Appl. Environ. Microbiol. 2017, 83, e00505–17. [Google Scholar] [CrossRef] [Green Version]
- Jurasek, L.; Marchessault, R.H. The role of phasins in the morphogenesis of poly (3-hydroxybutyrate) granules. Biomacromolecules 2002, 3, 256–261. [Google Scholar] [CrossRef]
- Pötter, M.; Steinbüchel, A. Poly (3-hydroxybutyrate) granule-associated proteins: Impacts on poly (3-hydroxybutyrate) synthesis and degradation. Biomacromolecules 2005, 6, 552–560. [Google Scholar] [CrossRef]
- Tirapelle, E.F.; Müller-Santos, M.; Tadra-Sfeir, M.Z.; Kadowaki, M.A.S.; Steffens, M.B.R.; Monteiro, R.A.; Souza, E.M.; Pedrosa, F.O.; Chubatsu, L.S. Identification of proteins associated with polyhydroxybutyrate granules from Herbaspirillum seropedicae SmR1-old partners, new players. PLoS ONE 2013, 8, e75066. [Google Scholar] [CrossRef] [Green Version]
- Maehara, A.; Doi, Y.; Nishiyama, T.; Takagi, Y.; Ueda, S.; Nakano, H.; Yamane, T. PhaR, a protein of unknown function conserved among short-chain-length polyhydroxyalkanoic acids producing bacteria, is a DNA-binding protein and represses Paracoccus denitrificans phaP expression in vitro. FEMS Microbiol. Lett. 2001, 200, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, J.M.; Amelia, T.S.M.; Mouriya, G.K.; Bhubalan, K.; Amirul, A.A.A.; Vigneswari, S.; Ramakrishna, S. Surface-modified highly biocompatible bacterial-poly (3-hydroxybutyrate-co-4-hydroxybutyrate): A review on the promising next-generation biomaterial. Polymers 2020, 13, 51. [Google Scholar] [CrossRef] [PubMed]
- Aneja, P.; Charles, T.C. Poly-3-hydroxybutyrate degradation in Rhizobium (Sinorhizobium) meliloti: Isolation and characterization of a gene encoding 3-hydroxybutyrate dehydrogenase. J. Bacteriol. 1999, 181, 849–857. [Google Scholar] [CrossRef] [Green Version]
- Handrick, R.; Reinhardt, S.; Jendrossek, D. Mobilization of poly (3-hydroxybutyrate) in Ralstonia eutropha. J. Bacteriol. 2000, 182, 5916–5918. [Google Scholar] [CrossRef] [Green Version]
- Bashan, Y.; Holguin, G.; De-Bashan, L.E. Azospirillum-plant relationships: Physiological, molecular, agricultural, and environmental advances (1997–2003). Can. J. Microbiol. 2004, 50, 521–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieille, M.; Elmerich, C. Characterization of an Azospirillum brasilense Sp7 gene homologous to Alcaligenes eutrophus phbB and Rhizobium meliloti nodG. Mol. Gen. Genet. 1992, 231, 375–384. [Google Scholar] [CrossRef]
- Aguilar, G.G. Análisis del Efecto de la Co-Transcripción de los Genes phbABC Sobre la Producción de PHB de Azospirillum brasilense Sp245. Master’s Thesis, Benemérita Universidad Autónoma de Puebla, Puebla, México, 2016. [Google Scholar]
- Peralta-Gil, M.; Segura, D.; Guzman, J.; Servin-Gonzalez, L.; Espin, G. Expression of the Azotobacter vinelandii poly-3-hydroxybutyrate biosynthetic phbBAC operon is driven by two overlapping promoters and is dependent on the transcriptional activator PhbR. J. Bacteriol. 2002, 184, 5672–5677. [Google Scholar] [CrossRef] [Green Version]
- Aneja, P.; Dai, M.; Lacorre, D.A.; Pillon, B.; Charles, T.C. Heterologous complementation of the exopolysaccharide synthesis and carbon utilization phenotypes of Sinorhizobium meliloti Rm1021 polyhydroxyalkanoate synthesis mutants. FEMS Microbiol. Lett. 2004, 239, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; McMillan, M.; Coumans, J.V.; Poljak, A.; Raftery, M.J.; Pereg, L. Cellular responses during morphological transformation in Azospirillum brasilense and its flcA knockout mutant. PLoS ONE 2014, 9, e114435. [Google Scholar] [CrossRef] [Green Version]
- Akhlaq, S.; Singh, D.; Mittal, N.; Srivastava, G.; Siddiqui, S.; Faridi, S.A.; Siddiqui, M.H. Polyhydroxybutyrate biosynthesis from different waste materials, degradation, and analytic methods: A short review. Polym. Bull. 2023, 80, 5965–5997. [Google Scholar] [CrossRef]
- Paul, E.; Mulard, D.; Blanc, P.; Fages, J.; Goma, G.; Pareilleux, A. Effects of partial O2 pressure, partial CO2 pressure, and agitation on growth kinetics of Azospirillum lipoferum under fermentor conditions. Appl. Environ. Microbiol. 1990, 56, 3235–3239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Eugenio, L.I.; Escapa, I.F.; Morales, V.; Dinjaski, N.; Galán, B.; García, J.L.; Prieto, M.A. The turno-ver of medium-chain-length polyhydroxyalkanoates in Pseudomonas putida KT2442 and the fundamental role of PhaZ depolymerase for the metabolic balance. Environ. Microbiol. 2010, 12, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Hauf, W.; Schlebusch, M.; Hüge, J.; Kopka, J.; Hagemann, M.; Forchhammer, K. Metabolic changes in Synechocystis PCC6803 upon nitrogen starvation: Excess NADPH sustains polyhydroxybutyrate accumulation. Metabolites 2013, 3, 101–118. [Google Scholar] [CrossRef] [Green Version]
- Bible, A.N.; Khalsa-Moyers, G.K.; Mukherjee, T.; Green, C.S.; Mishra, P.; Purcell, A.; Aksenova, A.; Hurst, G.B.; Alexandre, G. Metabolic adaptations of Azospirillum brasilense to oxygen stress by cell-to-cell clumping and flocculation. Appl. Environ. Microbiol. 2015, 81, 8346–8357. [Google Scholar] [CrossRef] [Green Version]
- Kefalogianni, I.; Aggelis, G. Metabolic activities in Azospirillum lipoferum grown in the presence of NH4. Appl. Microbiol. Biotechnol. 2003, 62, 574–578. [Google Scholar] [CrossRef]
- Kefalogianni, I.; Aggelis, G. Modeling growth and biochemical activities of Azospirillum spp. Appl. Microbiol. Biotechnol. 2002, 58, 352–357. [Google Scholar] [CrossRef]
- Kamnev, A.A.; Sadovnikova, J.N.; Tarantilis, P.A.; Polissiou, M.G.; Antonyuk, L.P. Responses of Azospirillum brasilense to nitrogen deficiency and to wheat lectin: A diffuse reflectance infrared Fourier transform (DRIFT) spectroscopic study. Microb. Ecol. 2008, 56, 615–624. [Google Scholar] [CrossRef]
- Gallori, E.; Bazzicalupo, M. Effect of nitrogen compounds on nitrogenase activity in Azospirillum brasilense. FEMS Microbiol. Lett. 1985. 28, 35–38. [CrossRef]
- Sun, J.; Van Dommelen, A.; Van Impe, J.; Vanderleyden, J. Involvement of glnB, glnZ, and glnD genes in the regulation of poly-3-hydroxybutyrate biosynthesis by ammonia in Azospirillum brasilense Sp7. Appl. Environ. Microbiol. 2002, 68, 985–988. [Google Scholar] [CrossRef] [Green Version]
- Kukolj, C.; Pedrosa, F.O.; de Souza, G.A.; Sumner, L.W.; Lei, Z.; Sumner, B.; Lei, Z.; Summer, B.; do Amaral, F.P.; Juexin, W.; et al. Proteomic and metabolomic analysis of Azospirillum brasilense ntrC mutant under high and low nitrogen conditions. J. Proteome Res. 2020, 19, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Peng, X.; Van Impe, J.; Vanderleyden, J. The ntrB and ntrC genes are involved in the regulation of poly-3-hydroxybutyrate biosynthesis by ammonia in Azospirillum brasilense Sp7. Appl. Environ. Microbiol. 2000, 66, 113–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacomboio, E.N.M.; Kim, E.Y.S.; Ruchaud-Correa, H.L.; Bonato, P.; de Oliveira-Pedrosa, F.; de Souza, E.M.; Chubatsu, L.S.; Müller-Santos, M. The transcriptional regulator NtrC controls glucose-6-phosphate dehydrogenase expression and polyhydroxybutyrate synthesis through NADPH availability in Herbaspirillum seropedicae. Sci. Rep. 2017, 7, 13546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdman, S.; Jurkevitch, E.; Schwartsburd, B.; Hampel, M.; Okon, Y. Aggregation in Azospirillum brasilense: Effects of chemical and physical factors and involvement of extracellular components. Microbiology 1998, 144, 1989–1999. [Google Scholar] [CrossRef] [Green Version]
- Burdman, S.; Jurketvitch, E.; Soria-Diaz, M.E.; Gil-Serrano, A.M.; Okon, Y. Extracellular polysaccharide composition of Azospirillum brasilense and its relation with cell aggregation. FEMS Microbiol. Lett. 2000, 489, 259–264. [Google Scholar] [CrossRef] [Green Version]
- Fischer, S.E.; Marioli, J.M.; Mori, G. Effect of root exudates on the exopolysaccharide composition and the lipopolysaccharide profile of Azospirillum brasilense Cd under saline stress. FEMS Microbiol. Lett. 2003, 219, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.L.; Santos, O.J.; Marcelino, P.R.; Milani, K.M.; Zuluaga, M.Y.; Zucareli, C.; Gonçalves, L.S. Maize inoculation with Azospirillum brasilense Ab-V5 cells enriched with exopolysaccharides and polyhydroxybutyrate results in high productivity under low N fertilizer input. Front. Microbiol. 2017, 8, 1873. [Google Scholar] [CrossRef] [Green Version]
- Joe, M.; Karthikeyan, M.B.; Sekar, C.; Deiveekasundaram, M. Optimization of biofloc production in Azospirillum brasilense (MTCC-125) and evaluation of its adherence with the roots of certain crops. Indian J. Microbiol. 2010, 50, 21–25. [Google Scholar] [CrossRef] [Green Version]
- Zhulin, I.B.; Bespalov, V.A.; Johnson, M.S.; Taylor, B.L. Oxygen taxis and proton motive force in Azospirillum brasilense. J. Bacteriol. 1996, 178, 5199–5204. [Google Scholar] [CrossRef] [Green Version]
- Konnova, S.A.; Makarov, O.E.; Skvortsov, I.M.; Ignatov, V.V. Isolation, fractionation and some properties of polysaccharides produced in a bound form by Azospirillum brasilense and their possible involvement in Azospirillum–wheat root interactions. FEMS Microbiol. Lett. 1994, 118, 93–99. [Google Scholar] [CrossRef]
- Konnova, S.A.; Brykova, O.S.; Sachkova, O.A.; Egorenkova, I.V.; Ignatov, V.V. Protective role of the polysaccharide-containing capsular components of Azospirillum brasilense. Microbiology 2001, 70, 436–440. [Google Scholar] [CrossRef]
- Vendan, R.T.; Thangaraju, M. Development and standardization of cyst based liquid formulation of Azospirillum bioinoculant. Acta Microbiol. Immunol. Hung. 2007, 54, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Fadel-Picheth, C.M.T.; Souza, E.M.; Rigo, L.U.; Yates, M.G.; Pedrosa, F.O. Regulation of Azospirillum brasilense nifA gene expression by ammonium and oxygen. FEMS Microbiol. Lett. 1999, 179, 281–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, A.; Burris, R.H. Regulation of nitrogenase activity by oxygen in Azospirillum brasilense and Azospirillum lipoferum. J. Bacteriol. 1987, 169, 944–948. [Google Scholar] [CrossRef] [Green Version]
- Bleakley, B.H.; Gaskins, M.H.; Hubbell, D.H.; Zam, S.G. Floc formation by Azospirillum lipoferum grown on poly-β-hydroxybutyrate. Appl. Environ. Microbiol. 1988, 54, 2986–2995. [Google Scholar] [CrossRef] [PubMed]
- Sadasivan, L.; Neyra, C.A. Cyst production and brown pigment formation in aging cultures of Azospirillum brasilense ATCC 29145. J. Bacteriol. 1987, 169, 1670–1677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Cui, Y.; Wu, L.; Tu, R.; Chen, S. cDNA-AFLP analysis of differential gene expression related to cell chemotactic and encystment of Azospirillum brasilense. Microbiol. Res. 2011, 166, 595–605. [Google Scholar] [CrossRef]
- Santos, M.S.; Hungria, M.; Nogueira, M.A. Production of polyhydroxybutyrate (PHB) and biofilm by Azospirillum brasilense aiming at the development of liquid inoculants with high performance. Afr. J. Bitechnol. 2017, 16, 1855–1862. [Google Scholar] [CrossRef] [Green Version]
- Bahat-Samet, E.; Castro-Sowinski, S.; Okon, Y. Arabinose content of extracellular polysaccharide plays a role in cell aggregation of Azospirillum brasilense. FEMS Microbiol. Lett. 2004, 237, 195–203. [Google Scholar] [CrossRef]
- Malinich, E.A.; Bauer, C.E. Transcriptome analysis of Azospirillum brasilense vegetative and cyst states reveals large-scale alterations in metabolic and replicative gene expression. Microb. Genom. 2018, 4. [Google Scholar] [CrossRef]
- Ramírez-Mata, A.; López-Lara, L.I.; Xiqui-Vázquez, M.L.; Jijón-Moreno, S.; Romero-Osorio, A.; Baca, B.E. The cyclic-di-GMP diguanylate cyclase CdgA has a role in biofilm formation and exopolysaccharide production in Azospirillum brasilense. Res. Microbiol. 2016, 167, 190–201. [Google Scholar] [CrossRef]
- Viruega-Góngora, V.I.; Acatitla-Jácome, I.S.; Zamorano-Sánchez, D.; Reyes-Carmona, S.R.; Xiqui-Vázquez, M.L.; Baca, B.E.; Ramírez-Mata, A. The GGDEF-EAL protein CdgB from Azospirillum baldaniorum Sp245, is a dual function enzyme with potential polar localization. PLoS ONE 2022, 17, e0278036. [Google Scholar] [CrossRef]
- Tugarova, A.V.; Scheludko, A.V.; Dyatlova, Y.A.; Filip’echeva, Y.A.; Kamnev, A.A. FTIR spectroscopic study of biofilms formed by the rhizobacterium Azospirillum brasilense Sp245 and its mutant Azospirillum brasilense Sp245. 1610. J. Mol. Struct. 2017, 1140, 142–147. [Google Scholar] [CrossRef]
- Shelud’ko, A.V.; Mokeev, D.I.; Evstigneeva, S.S.; Filip’echeva, Y.A.; Burov, A.M.; Petrova, L.P.; Ponomareva, E.G.; Katsy, E.I. Cell Ultrastructure in Azospirillum brasilense Biofilms. Microbiology 2020, 89, 50–63. [Google Scholar] [CrossRef]
- Sivasakthivelan, P.; Saranraj, P.; Al-Tawaha, A.R.; Amala, K.; Al Tawaha, A.R.; Thangadurai, D.; Sangeetha, J.; Rauf, A.; Khalid, S.; Alsultan, W.; et al. Adaptation of Azospirillum to stress conditions: A review. Adv. Environ. Biol. 2021, 15, 1–5. [Google Scholar] [CrossRef]
- Fibach-Paldi, S.; Burdman, S.; Okon, Y. Key physiological properties contributing to rhizosphere adaptation and plant growth promotion abilities of Azospirillum brasilense. FEMS Microbiol. Lett. 2012, 326, 99–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrasco-Espinosa, K.; García-Cabrera, R.I.; Bedoya-López, A.; Trujillo-Roldán, M.A.; Valdez-Cruz, N.A. Positive effect of reduced aeration rate on growth and stereospecificity of DL-malic acid consumption by Azospirillum brasilense: Improving the shelf life of a liquid inoculant formulation. J. Biotechnol. 2015, 195, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Tribelli, P.M.; Pezzoni, M.; Brito, M.G.; Montesinos, N.V.; Costa, C.S.; López, N.I. Response to lethal UVA radiation in the Antarctic bacterium Pseudomonas extremaustralis: Polyhydroxybutyrate and cold adaptation as protective factors. Extremophiles 2020, 24, 265–275. [Google Scholar] [CrossRef]
- López, J.A.; Naranjo, J.M.; Higuita, J.C.; Cubitto, M.A.; Cardona, C.A.; Villar, M.A. Biosynthesis of PHB from a new isolated Bacillus megaterium strain: Outlook on future developments with endospore forming bacteria. Biotechnol. Bioprocess Eng. 2012, 17, 250–258. [Google Scholar] [CrossRef]
- Koch, M.; Forchhammer, K. Polyhydroxybutyrate: A useful product of chlorotic cyanobacteria. Microb. Physiol. 2021, 31, 67–77. [Google Scholar] [CrossRef]
- Majerczak, K.; Wadkin-Snaith, D.; Magueijo, V.; Mulheran, P.; Liggat, J.; Johnston, K. Polyhydroxybutyrate: A review of experimental and simulation studies of the effect of fillers on crystallinity and mechanical properties. Polym. Int. 2022, 71, 1398–1408. [Google Scholar] [CrossRef]
- Raza, Z.A.; Khalil, S.; Abid, S. Recent progress in development and chemical modification of poly (hydroxybutyrate)-based blends for potential medical applications. Int. J. Biol. Macromol. 2020, 160, 77–100. [Google Scholar] [CrossRef] [PubMed]
- McAdam, B.; Brennan, F.M.; McDonald, P.; Mojicevic, M. Production of polyhydroxybutyrate (PHB) and factors impacting its chemical and mechanical characteristics. Polymer 2020, 12, 2908. [Google Scholar] [CrossRef] [PubMed]
- Verlinden, R.A.; Hill, D.J.; Kenward, M.; Williams, C.D.; Radecka, I. Bacterial synthesis of biodegradable polyhydroxyalkanoates. J. Appl. Microbiol. 2007, 102, 1437–1449. [Google Scholar] [CrossRef] [PubMed]
- Hankermeyer, C.R.; Tjeerdema, R.S. Polyhydroxybutyrate: Plastic made and degraded by microorganisms. Rev. Environ. Contam. Toxicol. 1999, 159, 1–24. [Google Scholar] [PubMed]
- Nair, L.S.; Laurencin, C.T. Biodegradable polymers as biomaterials. Prog. Polym. Sci. 2007, 32, 762–798. [Google Scholar] [CrossRef]
- Bonartsev, A.P.; Bonartseva, G.A.; Reshetov, I.V.; Kirpichnikov, M.P.; Shaitan, K.V. Application of polyhydroxyalkanoates in medicine and the biological activity of natural poly (3-hydroxybutyrate). Acta Nat. 2019, 11, 4–16. [Google Scholar] [CrossRef]
- Shishatskaya, E.I.; Voinova, O.N.; Goreva, A.V.; Mogilnaya, O.A.; Volova, T.G. Biocompatibility of polyhydroxybutyrate microspheres: In vitro and in vivo evaluation. J. Mat. Sci. 2008, 19, 2493–2502. [Google Scholar] [CrossRef] [Green Version]
- Pandian, S.R.K.; Kunjiappan, S.; Pavadai, P.; Sundarapandian, V.; Chandramohan, V.; Sundar, K. Delivery of Ursolic Acid by Polyhydroxybutyrate Nanoparticles for Cancer Therapy: In silico and in vitro Studies. Drug Res. 2022, 72, 72–81. [Google Scholar]
- Parsian, M.; Mutlu, P.; Yalcin, S.; Gunduz, U. Characterization of gemcitabine loaded polyhydroxybutyrate coated magnetic nanoparticles for targeted drug delivery. Anticancer Agents Med. Chem. 2020, 20, 1233–1240. [Google Scholar] [CrossRef]
- Lins, L.C.; Bazzo, G.C.; Barreto, P.L.; Pires, A.T. Composite PHB/chitosan microparticles obtained by spray drying: Effect of chitosan concentration and crosslinking agents on drug relesase. J. Braz. Chem. Soc. 2014, 25, 1462–1471. [Google Scholar] [CrossRef]
- Yagmurlu, M.F.; Korkusuz, F.; Gürsel, I.; Korkusuz, P.; Örs, Ü.; Hasirci, V. Sulbactam-cefoperazone polyhydroxybutyrate-co-hydroxyvalerate (PHBV) local antibiotic delivery system: In vivo effectiveness and biocompatibility in the treatment of implant-related experimental osteomyelitis. J. Biomed. Mater. Res. 1999, 46, 494–503. [Google Scholar] [CrossRef]
- Shrivastav, A.; Kim, H.Y.; Kim, Y.R. Advances in the applications of polyhydroxyalkanoate nanoparticles for novel drug delivery system. Biomed. Res. Int. 2013, 2013, 581684. [Google Scholar] [CrossRef] [Green Version]
- Barouti, G.; Jaffredo, C.G.; Guillaume, S.M. Advances in drug delivery systems based on synthetic poly (hydroxybutyrate)(co) polymers. Prog. Polym. Sci. 2017, 73, 1–31. [Google Scholar] [CrossRef]
- Prakash, P.; Lee, W.H.; Loo, C.Y.; Wong, H.S.J.; Parumasivam, T. Advances in polyhydroxyalkanoate nanocarriers for effective drug delivery: An overview and challenges. Nanomaterial 2022, 12, 175. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Lin, X.S.; Zheng, Z.; Deng, J.G.; Chen, J.C.; Ma, H.; Chen, G.Q. Poly (hydroxybutyrate-co-hydroxyhexanoate) promoted production of extracellular matrix of articular cartilage chondrocytes in vitro. Nanomaterial 2003, 24, 4273–4281. [Google Scholar] [CrossRef] [PubMed]
- Lootz, D.; Behrend, D.; Kramer, S.; Freier, T.; Haubold, A.; Benkiesser, G.; Schmitz, K.P.; Becher, B. Laser cutting: Influence on morphological and physicochemical properties of polyhydroxybutyrate. Nanomaterial 2001, 22, 2447–2452. [Google Scholar] [CrossRef]
- Sharma, N. Polyhydroxybutyrate (PHB) production by bacteria and its application as biodegradable plastic in various industries. Acad. J. Polym. Sci. 2019, 2, 001–003. [Google Scholar] [CrossRef]
- Lee, C.W.; Horiike, M.; Masutani, K.; Kimura, Y. Characteristic cell adhesion behaviors on various derivatives of poly (3-hydroxybutyrate) (PHB) and a block copolymer of poly (3-[RS]-hydroxybutyrate) and poly (oxyethylene). Polym. Degrad. Stab. 2015, 111, 194–202. [Google Scholar] [CrossRef]
- Garcia-Garcia, D.; Ferri, J.M.; Boronat, T.; López-Martínez, J.; Balart, R. Processing and characterization of binary poly (hydroxybutyrate)(PHB) and poly (caprolactone)(PCL) blends with improved impact properties. Polym. Bull. 2016, 73, 3333–3350. [Google Scholar] [CrossRef] [Green Version]
- Chu, C.F.; Lu, A.; Liszkowski, M.; Sipehia, R. Enhanced growth of animal and human endothelial cells on biodegradable polymers. Biochim. Biophys. Acta 1999, 1472, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Sanhueza, C.; Acevedo, F.; Rocha, S.; Villegas, P.; Seeger, M.; Navia, R. Polyhydroxyalkanoates as biomaterial for electrospun scaffolds. Int. J. Biol. Macromol. 2019, 124, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Rhim, J.W.; Park, H.M.; Ha, C.S. Bio-nanocomposites for food packaging applications. Prog. Polym Sci. 2013, 38, 1629–1652. [Google Scholar] [CrossRef]
- Popa, M.S.; Frone, A.N.; Panaitescu, D.M. Polyhydroxybutyrate blends: A solution for biodegradable packaging? Int. J. Biol. Macromol. 2022, 207, 263–277. [Google Scholar] [CrossRef]
- Atta, O.M.; Manan, S.; Shahzad, A.; Ul-Islam, M.; Ullah, M.W.; Yang, G. Biobased materials for active food packaging: A review. Food Hydrocoll. 2022, 125, 107419. [Google Scholar] [CrossRef]
- Hosokawa, M.N.; Darros, A.B.; Moris, V.A.D.; Paiva, J.M.F.D. Polyhydroxybutyrate composites with random mats of sisal and coconut fibers. Mater. Res. 2016, 20, 279–290. [Google Scholar] [CrossRef] [Green Version]
- Rajan, K.P.; Thomas, S.P.; Gopanna, A.; Chavali, M. Polyhydroxybutyrate (PHB): A standout biopolymer for environmental sustainability. In Handbook of Ecomaterials, 1st ed.; Torres-Martínez, L.M., Vasilievna-Kharissova, O., Ildusovich-Kharisov, B., Eds.; Springer: Cham, Switzerland, 2020; pp. 2803–2825. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Strain | Carbon Source | %PHB/DCW | Reference |
|---|---|---|---|
| A. brasilense Sp7 | Malate, fructose, pyruvate | 70–88% | [2,8,9,18] |
| R. eutropha | Glucose | 80–90% | [25] |
| R. rubrum | Acetate | [20] | |
| Pseudomonas extremaustralis | Octanoate, fructose, glucose, glycerol | 70–80% | [21,26,27] |
| Methylocystis hirsuta | Methanol:ethanol, methane | 73–85% | [28] |
| Bradyrhizobium diazoefficiens | Mannitol, glucose, and glycerol | 68% | [29] |
| A. vinelandii | Sucrose | 85% | [30] |
| Bacillus subtilis | Various sources | 60% | [31] |
| Rhizobium nepotum | Pyruvate | 62% | [28] |
| Parameter | PP | PHB |
|---|---|---|
| Tensile modulus (GPa) | 1.95 | 3–3.5 |
| Tensile strength (Mpa) | 31–45 | 20–40 |
| Elongation at break (%) | 50–400 | 5–10 |
| Crystallinity (%) | 42.6–70 | 50–60 |
| Melting temperature (°C) | 160–176 | 165–180 |
| Glass transition (°C) | −20–−5 | 2–9 |
| Density (g/cm2) | 0.905 | 1.25 |
| UV resistance | Poor | Good |
| Biodegradability | No | Yes |
| Biocompatibility | No | Yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez, M.d.l.Á.M.; Urzúa, L.S.; Carrillo, Y.A.; Ramírez, M.B.; Morales, L.J.M. Polyhydroxybutyrate Metabolism in Azospirillum brasilense and Its Applications, a Review. Polymers 2023, 15, 3027. https://doi.org/10.3390/polym15143027
Martínez MdlÁM, Urzúa LS, Carrillo YA, Ramírez MB, Morales LJM. Polyhydroxybutyrate Metabolism in Azospirillum brasilense and Its Applications, a Review. Polymers. 2023; 15(14):3027. https://doi.org/10.3390/polym15143027
Chicago/Turabian StyleMartínez, María de los Ángeles Martínez, Lucía Soto Urzúa, Yovani Aguilar Carrillo, Mirian Becerril Ramírez, and Luis Javier Martínez Morales. 2023. "Polyhydroxybutyrate Metabolism in Azospirillum brasilense and Its Applications, a Review" Polymers 15, no. 14: 3027. https://doi.org/10.3390/polym15143027