
Structural Elucidation and Anti-Tumor Activity of a Polysaccharide (CP2-S) from Cordyceps militaris Fruit Bodies
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Regents
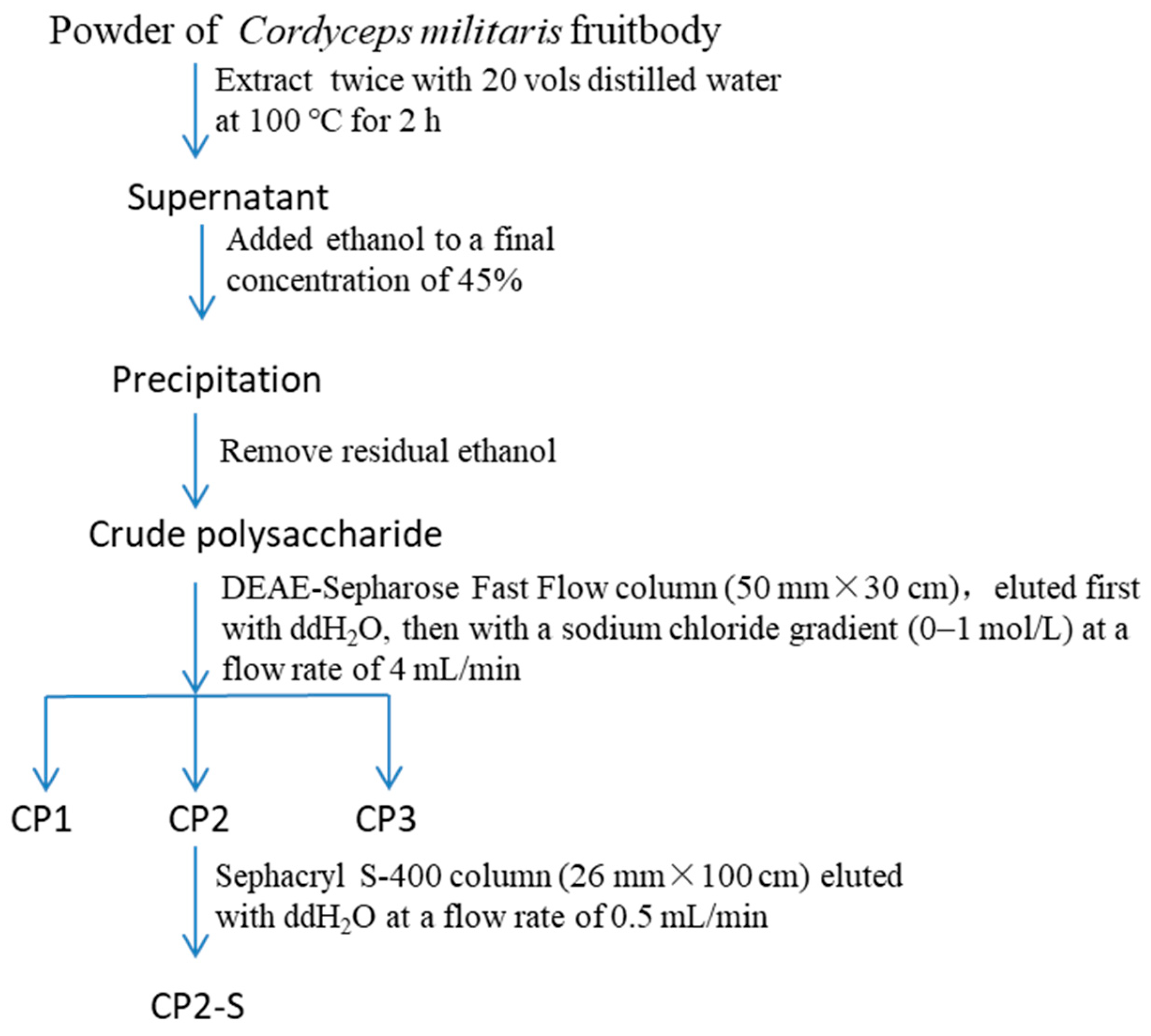
2.2. Polysaccharide Preparation
2.3. Homogeneity, Molecular Weight, and Monosaccharide Composition Analysis
2.4. Methylation and GC-MS Analysis
2.5. NMR Analysis
2.6. Effect of CP2-S on Lewis-Lung-Carcinoma-Bearing Mice In Vivo
2.6.1. Animal and Experimental Design
2.6.2. Determination of Tumor Inhibition Rate, Spleen Index, and Thymus Index
2.6.3. Determination of IgM and IgG Levels in Serum
3. Results
3.1. Structural Elucidation
3.1.1. Homogeneity, Molecular Weight, and Monosaccharide Composition Analysis
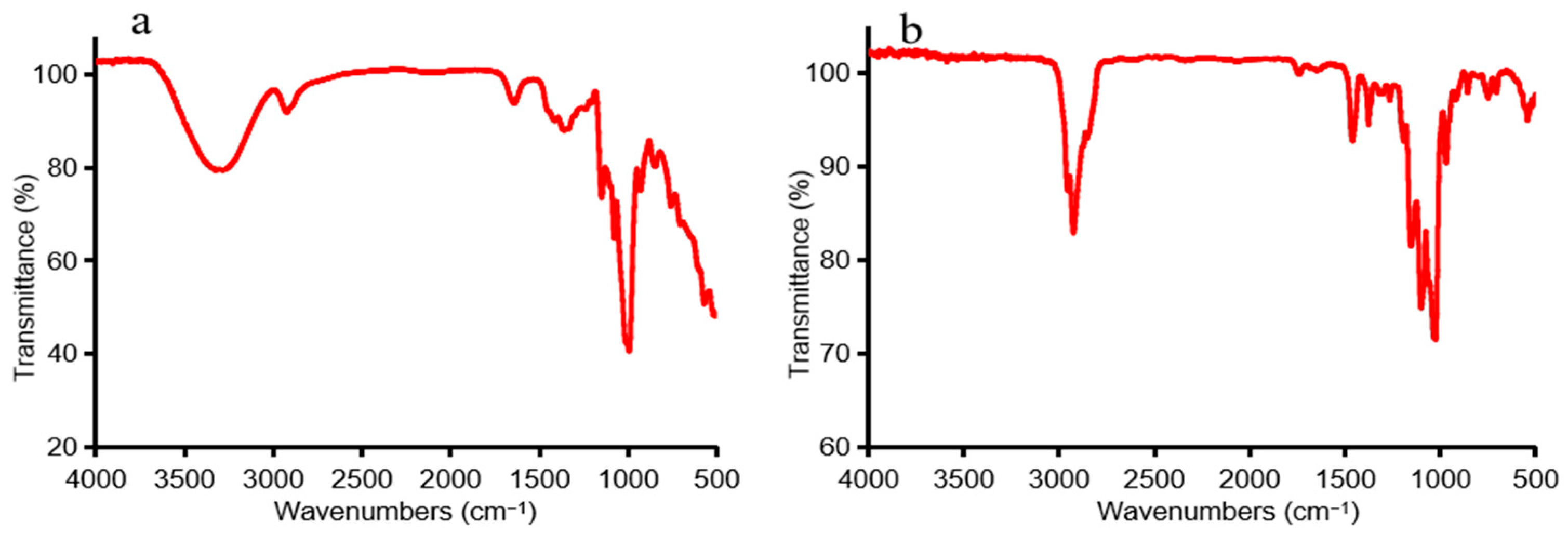
3.1.2. FTIR and Methylation Analysis
3.1.3. NMR Analysis

3.2. Effect of CP2-S on Lewis-Lung-Carcinoma-Bearing Mice In Vivo
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, S.; Wang, J.; Li, F.; Hou, P.; Yin, J.; Yang, Z.; Yang, X.; Li, T.; Xia, B.; Zhou, G.; et al. Structural characterisation and cholesterol efflux improving capacity of the novel polysaccharides from Cordyces militaris. Int. J. Biol. Macromol. 2019, 131, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Xin, X.; Weng, Y.; Gui, Z. Transcriptome-wide analysis reveals the progress of Cordyceps militaris subculture degeneration. PLoS ONE 2017, 12, e0186279. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Hong, E.K. Immunostimulating activity of the polysaccharides isolated from Cordyceps militaris. Int. Immunopharmacol. 2011, 11, 1226–1233. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Kwon, D.S.; Lee, K.R.; Park, J.M.; Ha, S.J.; Hong, E.K. Mechanism of macrophage activation induced by polysaccharide from Cordyceps militaris culture broth. Carbohydr. Polym. 2015, 120, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wen, Q.; Song, A.; Liu, Y.; Wang, F.; Jiang, B. Isolation and immune activity of a new acidic Cordyceps militaris exopolysaccharide. Int. J. Biol. Macromol. 2022, 194, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zeng, Y.; Cui, Y.S.; Liu, H.M.; Dong, C.X.; Sun, Y.X. Structural characterization, antioxidant and immunomodulatory activities of a neutral polysaccharide from Cordyceps militaris cultivated on hull-less barley. Carbohydr. Polym. 2020, 235, 115969. [Google Scholar] [CrossRef] [PubMed]
- He, B.L.; Zheng, Q.W.; Guo, L.Q.; Huang, J.Y.; Lin, J.F. Structural characterization and immune-enhancing activity of a novel high-molecular-weight polysaccharide from Cordyceps militaris. Int. J. Biol. Macromol. 2020, 145, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Kim, J.Y.; Kang, J.S.; Kim, H.M.; Kim, Y.O.; Hong, I.P.; Lee, M.K.; Hong, J.T.; Kim, Y.; Han, S.B. Cordlan polysaccharide isolated from mushroom Cordyceps militaris induces dendritic cell maturation through toll-like receptor 4 signalings. Food Chem. Toxicol. 2010, 48, 1926–1933. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.L.; Lu, J.H.; Zhang, N.; Zhang, G.R.; Teng, L.R. Extraction, purification and anti-tumor activity of polysaccharide from mycelium of mutant Cordyceps militaris. Chem. Res. Chin. Univ. 2010, 26, 798–802. [Google Scholar]
- Yu, R.M.; Yang, W.; Song, L.Y.; Yan, C.Y.; Zhang, Z.; Zhao, Y. Structural characterization and antioxidant activity of a polysaccharide from the fruit bodies of cultured Cordyceps militaris. Carbohydr. Polym. 2007, 70, 430–436. [Google Scholar] [CrossRef]
- Ohta, Y.; Lee, J.B.; Hayashi, K.; Fujita, A.; Park, D.K.; Hayashi, T. In vivo anti-influenza virus activity of an immunomodulatory acidic polysaccharide isolated from Cordyceps militaris grown on germinated soybeans. J. Agr. Food Chem. 2007, 55, 10194–10199. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.L.; Yue, J.; Hui, N.; Zhi, Y.; Kashif Hayat, K.; Yang, X.J.; Zhang, D.; Chu, S.H.; Zhou, P. Anti-hyperlipidemia and gut microbiota community regulation effects of selenium-rich Cordyceps militaris polysaccharides on the high-fat diet-fed mice mode. Foods 2021, 10, 2252. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.L.; Pan, L.C.; Tang, Y.; Luo, Y.; Zhu, Z.Y.; Sun, H.Q.; Meng, M.; Zhang, Y.M. 1H NMR-based metabonomics of the hypoglycemic effect of polysaccharides from Cordyceps militaris on streptozotocin-induced diabetes in mice. Nat. Prod. Res. 2020, 34, 1366–1372. [Google Scholar] [CrossRef] [PubMed]
- Smiderle, F.; Baggio, C.; Borato, D.; Santana-Filho, A.; Sassaki, G.; Lacomini, M.; Van Griensven, L. Anti-inflammatory properties of the medicinal mushroom Cordyceps militaris might be related to its linear(1→3)- β-D-glucan. PLoS ONE 2014, 9, e110266. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Q.; Lin, P.; Wang, J.; Liu, N.; Yin, F.; Shen, N.; Guo, S.D. Purification, characterization and anti-atherosclerotic effects of the polysaccharides from the fruiting body of Cordyceps militaris. Int. J. Biol. Macromol. 2021, 181, 890–904. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.X.; Wen, C.T.; Duan, Y.Q.; Zhang, H.H.; Ma, H.L. Advance in Cordyceps militaris (Linn) Link polysaccharides: Isolation, structure, and bioactivities: A review. Int. J. Biol. Macromol. 2019, 132, 906–914. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Du, X.; Guo, Y.B.; Chang, M.C.; Deng, B.; Liu, J.Y.; Cao, J.L. Elucidation of physicochemical properties of polysaccharides extracted from Cordyceps militaris fruiting bodies with different drying treatments and their effects on ulcerative colitis in zebrafish. Front. Nutr. 2022, 9, 980357. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.H.; Yang, X.Q.; Lin, P.; Liu, N.; Li, X.J.; Zhang, B.H.; Guo, S.D. Purification, structural characterization, and PCSK9 secretion inhibitory effect of the novel alkali-extracted polysaccharide from Cordyceps militaris. Int. J. Biol. Macromol. 2021, 179, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Zhang, J.; Zhang, T.H. Immunomodulatory activities of polysaccharides from Ganoderma on immune effector cells. Food Chem. 2021, 340, 127933. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, Z.X.; Song, K.F.; Li, L.B.; Chen, M. Medicinal value of edible mushroom polysaccharides: A review. J. Future Foods. 2023, 3, 16–23. [Google Scholar] [CrossRef]
- Maity, P.; Sen, I.; Chakraborty, I.; Mondal, S.; Bar, H.; Bhanja, S.; Mandal, S.; Maity, G.N. Biologically active polysaccharide from edible mushrooms: A review. Int. J. Biol. Macromol. 2021, 172, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.X.; Jing, Y.S.; Zhou, Q.Q.; Hu, X.J.; Zhu, J.H.; Guo, Z.Y.; Song, L.Y.; Yu, R.M. Structural elucidation and immunostimulatory activity of a new polysaccharide from Cordyceps militaris. Food Funct. 2018, 9, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.P.; Duan, Y.Q.; Yang, W.Y.; Zhang, H.H.; Li, C.Z.; Zhang, J.X. Structural elucidation and immunostimulatory activity of polysaccharide isolated by subcritical water extraction from Cordyceps militaris. Carbohydr. Polym. 2017, 157, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.S.; Cui, X.; Chen, Z.Y. Elucidation and biological activities of a new polysaccharide from cultured Cordyceps militaris. Carbohydr. Polym. 2014, 102, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Kwon, J.S.; Yun, J.S.; Pahk, J.W.; Shin, W.C.; Lee, S.Y.; Hong, E.K. Structural characterization of immunostimulating polysaccharide from cultured mycelia of Cordyceps militaris. Carbohydr. Polym. 2010, 80, 1011–1017. [Google Scholar] [CrossRef]
- Liu, Y.F.; Su, P.F.; Xu, J.F.; Chen, S.; Zhang, J.S.; Zhou, S.; Wang, Y.T.; Tang, Q.J.; Wang, Y.L. Structural characterization of a bioactive water-soluble heteropolysaccharide from Nostoc sphaeroids kütz. Carbohydr. Polym. 2018, 200, 552–559. [Google Scholar] [CrossRef]
- Jiang, H.T.; Ren, Y.H.; Wu, Y.L.; Zhou, Q.C.; Hua, C. Anti-fatigue effects of Cordyceps militaris polysaccharide in mice. Acta Edulis Fungi 2014, 21, 55–59, (In Chinese with English abstract). [Google Scholar]
- Liu, J.Y.; Feng, C.P.; Li, X.L.; Chang, M.C.; Meng, J.L.; Xu, L.J. Immunomodulatory and antioxidative activity of Cordyceps militaris polysaccharides in mice. Int. J. Biol. Macromol. 2016, 86, 594–598. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.D.; Xu, R.; Zhou, J.Y.; Chen, J.Q.; Wang, L.; Liu, X.S.; Liang, C.L.; Liu, B.H.; Lu, R.R.; Wu, J.B.; et al. Cordyceps militaris polysaccharides exerted protective effects on diabetic nephropathy in mice via regulation of autophagy. Food Funct. 2019, 10, 5102. [Google Scholar] [CrossRef] [PubMed]
- Li, F.L.; Ma, Y.M.; Hua, W.Y.; Liu, Y.X.; Li, L.; Lu, Z.K.; Jiang, X.K.; Liu, C.; Liu, J.X. Cordyceps militaris polysaccharide exerted anticancer effect via activating the endogenous apoptosis pathway. Pharmacogn. Mag. 2022, 18, 669–674. [Google Scholar]
- Yu, W.Q.; Wang, X.L.; Ji, H.H.; Miao, M.; Zhang, B.H.; Li, H.; Zhang, Z.Y.; Ji, C.F.; Guo, S.D. CM3-SII polysaccharide obtained from Cordyceps militaris ameliorates hyperlipidemia in heterozygous LDLR-deficient hamsters by modulating gut microbiota and NPC1L1 and PPARα levels. Int. J. Biol. Macromol. 2023, 239, 124293. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Lin, P.; Yu, W.Q.; Shen, N.; Li, Y.; Guo, S.D. The Cordyceps militaris-derived polysaccharide CM1 alleviates atherosclerosis in LDLR(-/-) mice by improving hyperlipidemia. Front. Mol. Biosci. 2021, 8, 783807. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.Q.; Yin, F.; Shen, N.; Lin, P.; Xia, B.; Li, Y.J.; Guo, S.D. Polysaccharide CM1 from Cordyceps militaris hinders adipocyte differentiation and alleviates hyperlipidemia in LDLR(+/-) hamsters. Lipids Health Dis. 2021, 20, 178. [Google Scholar] [CrossRef]
- Zhao, H.; Li, M.; Liu, L.; Li, D.; Zhao, L.J.; Wu, Z.; Zhou, M.X.; Jia, L.; Yang, F. Cordyceps militaris polysaccharide alleviates diabetic symptoms by regulating gut microbiota against TLR4/NF-κB pathway. Int. J. Biol. Macromol. 2023, 230, 123241. [Google Scholar] [CrossRef]
- Song, L.Y.; Yang, J.T.; Kong, W.H.; Liu, Y.; Liu, S.Y.; Su, L. Cordyceps militaris polysaccharide alleviates ovalbumin-induced allergic asthma through the Nrf2/HO-1 and NF-κB signaling pathways and regulates the gut microbiota. Int. J. Biol. Macromol. 2023, 238, 124333. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.Y.; Zhu, Z.Y. Using Cordyceps militaris extracellular polysaccharides to prevent Pb2+-induced liver and kidney toxicity by activating Nrf2 signals and modulating gut microbiota. Food Funct. 2020, 11, 9226. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wang, M.L.; Jin, C.; Chen, H.J.; Li, S.H.; Li, S.Y.; Dou, X.F.; Jia, J.Q.; Gui, Z.Z. Cordyceps militaris polysaccharide triggers apoptosis and G0/G1 cell arrest in cancer cells. J. Asia-Pac. Entomol. 2015, 18, 433–438. [Google Scholar] [CrossRef]
- Jing, Y.S.; Zhu, J.H.; Liu, T.; Bi, S.X.; Hu, X.J.; Chen, Z.Y.; Song, L.Y.; Lv, Y.J.; Yu, R.M. Structural characterization and biological activities of a novel polysaccharide from cultured Cordyceps militaris and its sulfated derivative. J. Agric. Food Chem. 2015, 63, 3464–3471. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.N.; Tang, Q.J.; Zhou, S.; Liu, Y.F.; Zhang, Z.; Gao, X.H.; Wang, S.P.; Wang, Z.L. Isolation and purification of a polysaccharide from the caterpillar medicinal mushroom Cordyceps militaris (Ascomycetes) fruit bodies and its immunomodulation of RAW 264.7 macrophages. Int. J. Med. Mushrooms 2014, 16, 247–257. [Google Scholar] [CrossRef]
- Zhang, J.S.; Tang, Q.J.; Zhou, C.Y.; Jia, W.; Silva, L.D.; Nguyen, L.D.; Reutter, W.; Fan, H. GLIS a bioactive proteoglycan fraction from Ganoderma lucidum, displays anti-tumour activity by increasing both humoral and cellular immune response. Life Sci. 2010, 87, 628–637. [Google Scholar] [CrossRef]
- Corsaro, M.M.; Castro, C.D.; Naldi, T.; Parrilli, M.; Tomas, J.M.; Regue, M. 1H and 13C NMR characterization and secondary structure of the K2 polysaccharide of Klebsiella pneumoniae strain 52145. Carbohydr Res. 2005, 340, 2212–2217. [Google Scholar] [CrossRef]
- Yang, B.; Prasad, K.N.; Jiang, Y.M. Structure identification of a polysaccharide purified from litchi (Litchi chinensis Sonn.) pulp. Carbohydr. Polym. 2016, 137, 570–575. [Google Scholar] [CrossRef]
- Rong, L.; Li, G.Q.; Zhang, Y.X.; Xiao, Y.C.; Qiao, Y.J.; Yang, M.M.; Wei, L.X.; Bi, H.T.; Gao, T.T. Structure and immunomodulatory activity of a water-soluble α-glucan from Hirsutella sinensis mycelia. Int. J. Biol. Macromol. 2021, 189, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.H.; Zhang, C.Q.; Li, M.X.; Chen, X.; Ding, K. Structural elucidation of a glucan from Crataegus pinnatifida and its bioactivity on intestinal bacteria strains. Int. J. Biol. Macromol. 2019, 128, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Guo, L.; Yan, A.P.; Feng, L.; Wan, Y.Q. Fractionation, structure and conformation characterization of polysaccharides from Anoectochilus roxburghii. Carbohydr. Polym. 2020, 231, 115688. [Google Scholar] [CrossRef] [PubMed]
- Wasser, S.P. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar]
- Miao, M.; Yu, W.Q.; Li, Y.; Sun, Y.L.; Guo, S.D. Structural elucidation and activities of Cordyceps militaris-derived polysaccharides: A review. Front. Nutr. 2022, 9, 898674. [Google Scholar] [CrossRef]
- Ji, C.F.; Zhang, Z.Y.; Zhang, B.H.; Chen, J.Y.; Liu, R.Y.; Song, D.X.; Li, W.L.; Lin, N.; Zou, X.; Wang, J.; et al. Purification, characterization, and in vitro antitumor activity of a novel glucan from the purple sweet potato Ipomoea Batatas (L.) Lam. Carbohydr. Polym. 2021, 257, 117605. [Google Scholar] [CrossRef]
- Bi, S.X.; Huang, W.J.; Chen, S.; Huang, C.H.; Li, C.L.; Guo, Z.Y.; Yang, J.N.; Zhu, J.H.; Song, L.Y.; Yu, R. Cordyceps militaris polysaccharide converts immunosuppressive macrophages into M1-like phenotype and activates T lymphocytes by inhibiting the PD-L1/PD-1 axis between TAMs and T lymphocytes. Int. J. Biol. Macromol. 2020, 150, 261–280. [Google Scholar] [CrossRef]
- Yan, J.K.; Wang, W.Q.; Li, L.; Wu, J.Y. Physiochemical properties and antitumor activities of two α-glucans isolated from hot water and alkaline extracts of Cordyceps (Cs-HK1) fungal mycelia. Carbohydr. Polym. 2011, 85, 753–758. [Google Scholar] [CrossRef]
- Zhu, Z.Y.; Liu, N.; Si, C.L.; Liu, Y.; Ding, L.N.; Jing, C.; Liu, A.J.; Zhang, Y.M. Structure and anti-tumor activity of high-molecular-weight polysaccharides from cultured mycelium of Cordyceps gunnii. Carbohydr. Polym. 2012, 88, 1072–1076. [Google Scholar] [CrossRef]
- Klis, F.M.; Ram, A.F.J.; De Groot, P.W.J. A molecular and genomic view of the fungal cell wall. In Biology of the Fungal Cell; Howard, R.J., Gow, N.A.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; Volume 8, pp. 97–120. [Google Scholar]
- Liu, Y.F.; Zhao, Y.; Yang, Y.; Tang, Q.J.; Zhou, S.; Wu, D.; Zhang, J.S. Structural characteristics and hypoglycemic activity of polysaccharides from Coprinus comatus. Bioact. Carbohydr. Dietary Fibre. 2013, 2, 164–169. [Google Scholar] [CrossRef]
- Biswas, S.K.; Mantovani, A. Macrophage plasticity and interaction with lymphocyte subsets: Cancer as a paradigm. Nat. Immunol. 2010, 11, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Epelman, S.; Lavine, K.J.; Randolph, G.J. Origin and functions of tissue macrophages. Immunity 2014, 41, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Lawrence, T.; Nizet, V. Innate immunity gone awry: Linking microbial infections to chronic inflammation and cancer. Cell 2006, 124, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Zhao, W.Y.; Xie, Q.P.; Zhan, Y.H.; Wu, B. Lentinan reduces tumor progression by enhancing gemcitabine chemotherapy in urothelial bladder cancer. Surg. Oncol. 2015, 24, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Xiao, N. Effect of lentinan combined with docetaxel and cisplatin on the proliferation and apotosis of BGC823 cells. Tumor Boil. 2013, 34, 1531. [Google Scholar] [CrossRef]
- Harada, K.; Itashiki, Y.T.; Ueyama, Y. Effects of lentinan alone and in combination with fluoropyrimidine anticancer agent on growth of human oral squamous cell carcinoma in vitro and in vivo. Int. J. Oncol. 2010, 37, 623–631. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PMAA | Type of Linkage | Molar Ratio | Major Mass Fragments (m/z) |
|---|---|---|---|
| 1,5-Ac2-2,3,4,6-Me4-Glcp | Terminal-Glcp | 1.00 | 87, 102, 118, 129, 145, 161, 162, 205 |
| 1,4,5-Ac3-2,3,6-Me3-Glcp | →4)-Glcp-(1→ | 10.14 | 87, 99, 102, 113, 118, 129, 131, 162, 173, 233 |
| 1,4,5,6-Ac4-2,3-Me2-Glcp | →4,6)-Glcp-(1→ | 0.97 | 85, 99, 102, 118, 127, 142, 159, 162, 187, 201, 261 |
| Residues | Proton or Carbon | |||||
|---|---|---|---|---|---|---|
| H-1/C-1 | H-2/C-2 | H-3/C-3 | H-4/C-4 | H-5/C-5 | H-6/C-6 | |
| A: →4)-α-Glcp(1→ | 5.38 | 3.67 | 3.97 | 3.64 | 3.85 | 3.83 a, 3.89 b |
| 100.62 | 72.55 | 74.22 | 78.40 * | 72.29 | 61.61 | |
| B: →4,6)-α-Glcp(1→ | 5.37 | 3.60 | 3.73 | 3.61 | 3.84 | 3.44 |
| 100.83 | 72.79 | 73.98 | 79.04 * | 72.29 | 70.52 * | |
| C: α-Glcp(1→ | 4.98 | 3.62 | 4.03 | 3.86 | 3.70 | 3.76 a, 3.86 b |
| 99.57 | 72.65 | 74.22 | 71.29 | 73.97 | 61.61 | |
| Group | Dosage (mg/kg/d) | Tumor Inhibition Rate (%) | Spleen Index | Thymus Index |
|---|---|---|---|---|
| CP2-S | 12.5 | 17.8 ± 0.6 | 0.81 ± 0.04 | 0.26 ± 0.02 |
| 50 | 24.5 ± 0.7 | 0.94 ± 0.06 | 0.26 ± 0.02 | |
| 100 | 29.5 ± 0.4 | 0.88 ± 0.05 | 0.28 ± 0.02 | |
| Cisplatin | 2 | 78.3 ± 0.3 | 0.66 ± 0.03 | 0.19 ± 0.01 |
| Control | physiological saline | 0 | 1.00 | 1.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, L.; Wang, J.; Tang, Q.; Liu, Y. Structural Elucidation and Anti-Tumor Activity of a Polysaccharide (CP2-S) from Cordyceps militaris Fruit Bodies. Polymers 2024, 16, 1972. https://doi.org/10.3390/polym16141972
Zhu L, Wang J, Tang Q, Liu Y. Structural Elucidation and Anti-Tumor Activity of a Polysaccharide (CP2-S) from Cordyceps militaris Fruit Bodies. Polymers. 2024; 16(14):1972. https://doi.org/10.3390/polym16141972
Chicago/Turabian StyleZhu, Lina, Jinyan Wang, Qingjiu Tang, and Yanfang Liu. 2024. "Structural Elucidation and Anti-Tumor Activity of a Polysaccharide (CP2-S) from Cordyceps militaris Fruit Bodies" Polymers 16, no. 14: 1972. https://doi.org/10.3390/polym16141972