
Microbial Biosynthesis of Medium-Chain-Length Polyhydroxyalkanoate (mcl-PHA) from Waste Cooking Oil
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Strains and Culture Conditions
2.2. Screening of mcl-PHA-Producing Bacterial Isolates
2.3. Characterization and Identification of Test Isolates
2.4. Extraction, Characterization, and Quantification of mcl-PHA
- -
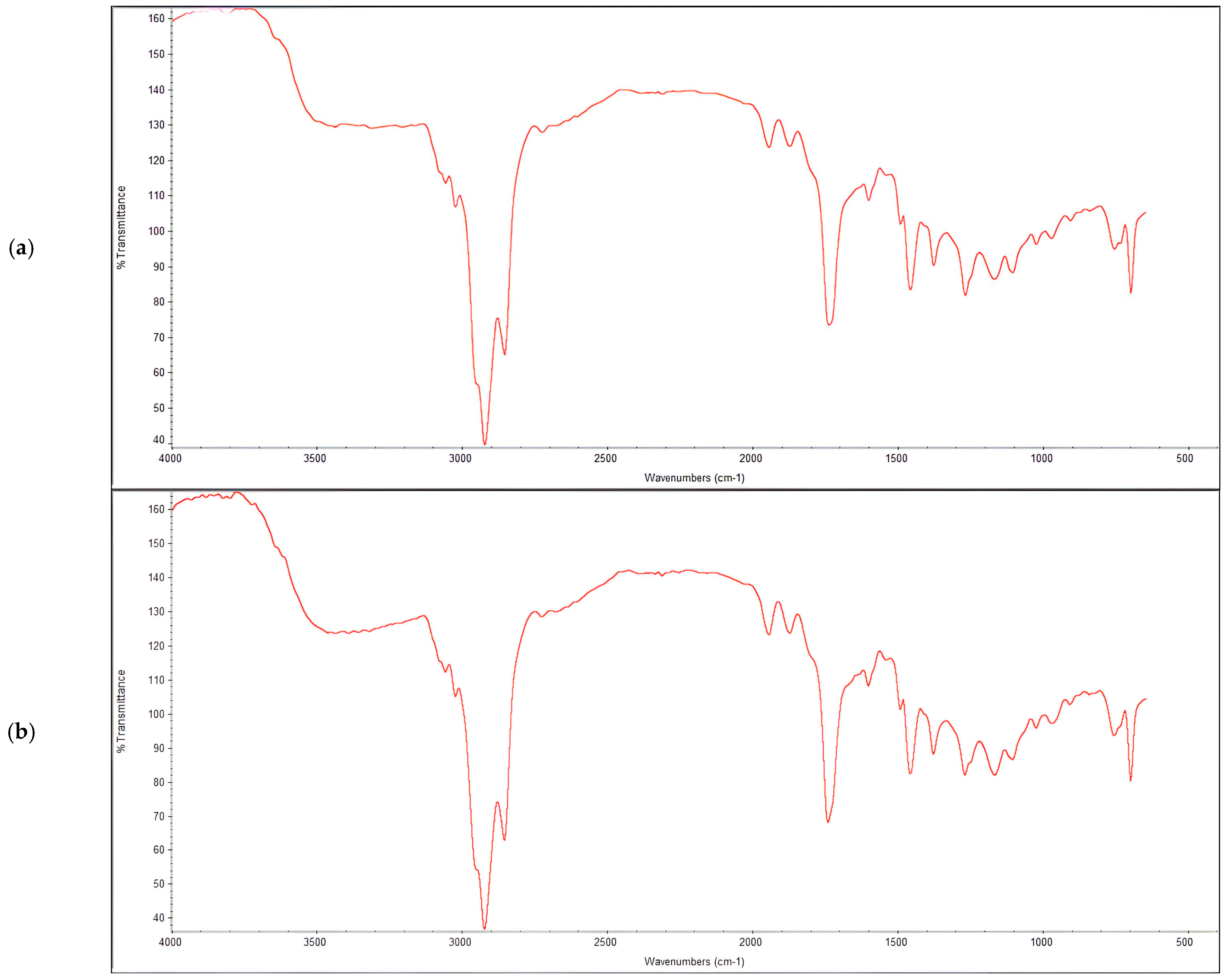
- Fourier Transform Infrared Spectroscopy (FTIR): The functional groups present in the extracted PHA were evaluated using an FT-IR spectrometer (PerkinElmer, Cambridge, UK). Briefly, 1 mg of PHA was diluted in 7 mL of chloroform and a drop of the mixture was overlaid on an FT-IR KBr disk. The infrared (IR) spectra were recorded in the range of 400–4000 cm−1, at a spectral resolution of 4 cm−1, under vacuum pressure.
- -
- Gas Chromatography–Mass Spectrometry (GC-MS): The monomer composition of the mcl-PHA was determined using GC-MS (Agilent Technologies, Cheadle, UK), equipped with a gas chromatograph (7890B) and a mass spectrometer detector (5977A). Approximately 10 mg of the extracted PHA was methanolized with 2 mL of methanol, containing 15% (v/v) sulfuric acid and 2 mL of chloroform, at 100 °C for 140 min. After cooling, 1 mL of deionized water was added, and the mixture was vortexed and allowed to separate. The organic phase was analyzed by GC-MS, equipped with a capillary column (30 m × 0.25 mm × 0.25 µm).
- -
- Quantification of mcl-PHA: The mcl-PHA content in the bacterial biomass was quantified by gravimetric analysis [15]. The dried PHA granules obtained after precipitation and drying were weighed, and the PHA content was expressed as a percentage of the cell dry weight (CDW). The percentage titer of the PHA was calculated using the following formula. The intracellular PHA accumulation was assessed as the percentage of the PHA in relation to the dry cell weight and was calculated as follows:
2.5. Optimization of Cultural Parameters for Maximum mcl-PHA Production
2.5.1. Effects of Different Carbon Sources on Growth and mcl-PHA Production
2.5.2. Effects of Different Nitrogen Sources on Growth and mcl-PHA Production
2.5.3. Effect of Varied pH on Growth and mcl-PHA Production
2.6. Bioreactor Cultivation Procedure
2.7. Analytical Techniques
2.7.1. Estimation of Cell Dry Weight
2.7.2. pH Regulation
2.8. Production Kinetics
- PHA volumetric productivity (Pv):
- 2.
- Biomass volumetric productivity (Bv):
- 3.
- Yield of biomass from the substrate (Yx/s):
- 4.
- Yield of PHA from the substrate (YP/s):
- 5.
- Specific growth rate (µ):
2.9. Data Collection and Analysis
3. Results
3.1. Strain Isolation, Screening, and Dentification
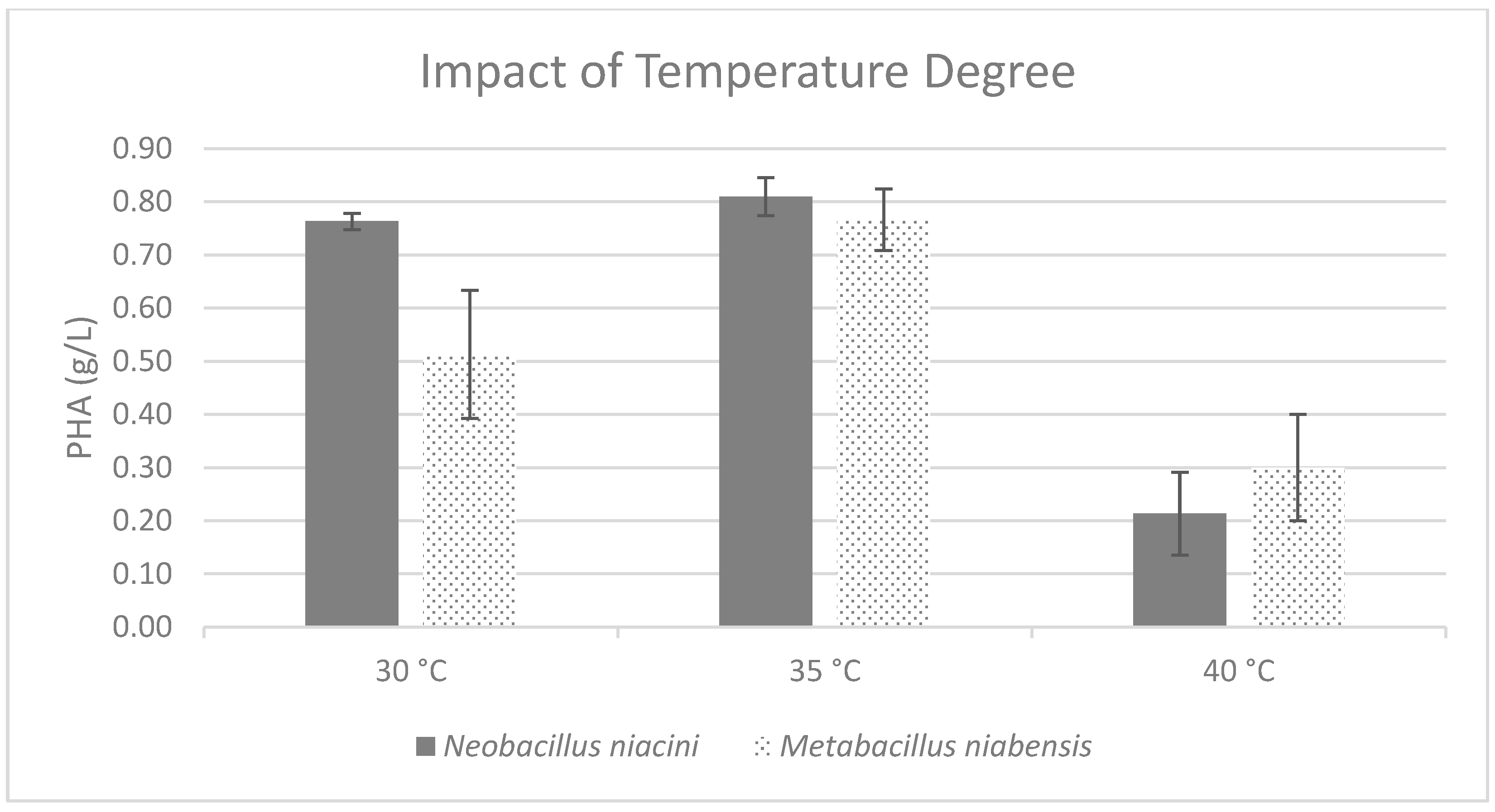
3.2. Cultivation Conditions Optimization
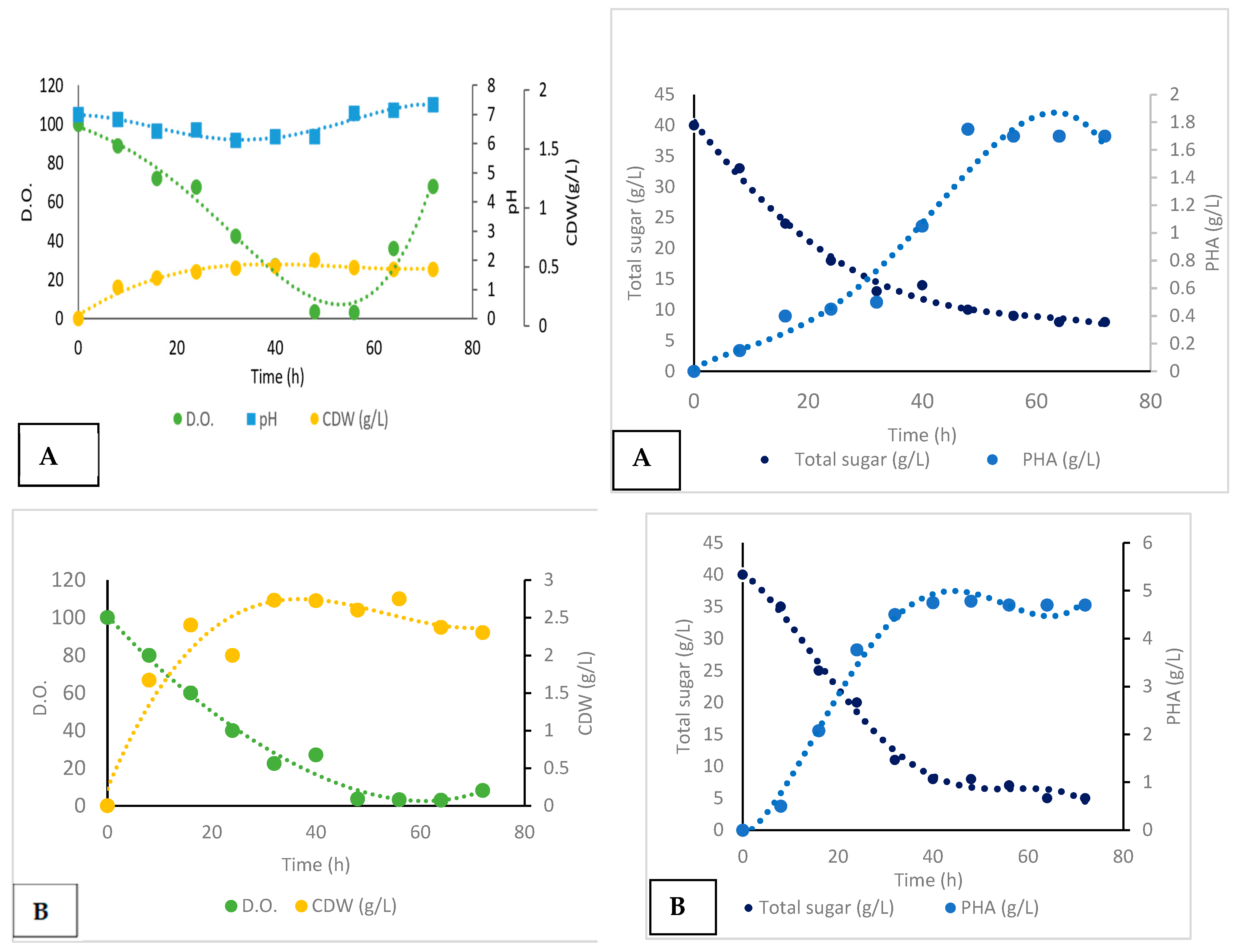
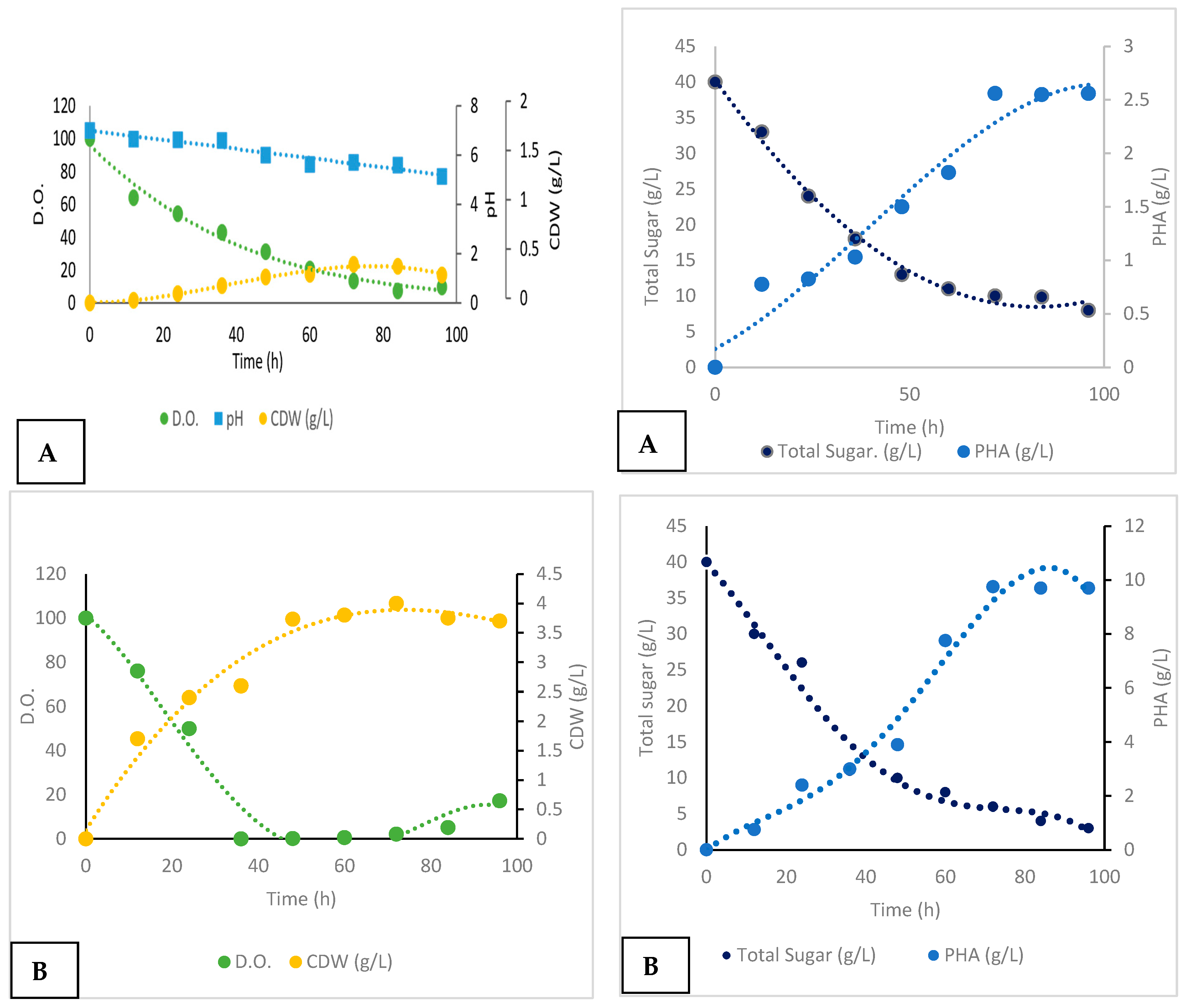
Batch Cultivation and Production
3.3. Characterization of Produced PHAs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dong, R.; Gao, A.; Zhu, Y.; Xu, B.; Du, J.; Ping, S. The Development of a New Thermoplastic Elastomer (TPE)-Modified Asphalt. Buildings 2023, 13, 1451. [Google Scholar] [CrossRef]
- Basak, S. Thermoplastic Elastomers in Biomedical Industry—Evolution and Current Trends. J. Macromol. Sci. Part A 2021, 58, 579–593. [Google Scholar] [CrossRef]
- Akinmulewo, A.B.; Nwinyi, O.C. Polyhydroxyalkanoate: A Biodegradable Polymer (A Mini Review). J. Phys. Conf. Ser. 2019, 1378, 042007. [Google Scholar] [CrossRef]
- Alweera, A.; Anjum, Z.; Younis, K. Natural pigments-based biodegradable food packaging films. In Renewable Dyes and Pigments; Ul Islam, S., Ed.; Elsevier: Amsterdam, The Netherlands, 2024; pp. 77–92. ISBN 9780443152139. [Google Scholar] [CrossRef]
- Mukherjee, A.; Koller, M. Microbial PolyHydroxyAlkanoate (PHA) Biopolymers—Intrinsically Natural. Bioengineering 2023, 10, 855. [Google Scholar] [CrossRef] [PubMed]
- Tortajada, M.; da Silva, L.F.; Prieto, M.A. Second-Generation Functionalized Medium-Chain-Length Polyhydroxyalkanoates: The Gateway to High-Value Bioplastic Applications. Int. Microbiol. 2013, 16, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Dartiailh, C.; Blunt, W.; Sharma, P.K.; Liu, S.; Cicek, N.; Levin, D.B. The Thermal and Mechanical Properties of Medium Chain-Length Polyhydroxyalkanoates Produced by Pseudomonas putida LS46 on Various Substrates. Front. Bioeng. Biotechnol. 2021, 8, 617489. [Google Scholar] [CrossRef] [PubMed]
- Che Ab Aziz, N.I.; Zakaria, Y.; Md Muslim, N.Z.; Nik Hassan, N.F. Emerging and Advanced Technologies in Biodegradable Plastics for Sustainability. In Handbook of Biodegradable Materials; Ali, G.A.M., Makhlouf, A.S.H., Eds.; Springer: Cham, Germany, 2022. [Google Scholar] [CrossRef]
- Agnew, D.E.; Pfleger, B.F. Synthetic Biology Strategies for Synthesizing Polyhydroxyalkanoates from Unrelated Carbon Sources. Chem. Eng. Sci. 2013, 103, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Cánovas, V.; Garcia-Chumillas, S.; Monzó, F.; Simó-Cabrera, L.; Fernández-Ayuso, C.; Pire, C.; Martínez-Espinosa, R.M. Analysis of Polyhydroxyalkanoates Granules in Haloferax mediterranei by Double-Fluorescence Staining with Nile Red and SYBR Green by Confocal Fluorescence Microscopy. Polymers 2021, 13, 1582. [Google Scholar] [CrossRef] [PubMed]
- Moore, E.; Arnscheidt, A.; Krüger, A.; Strömpl, C.; Mau, M. Simplified Protocols for the Preparation of Genomic DNA from Bacterial Cultures. In Molecular Microbial Ecology Manual, 2nd ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 3–18. [Google Scholar]
- Azcárate-Peril, M.A.; Raya, R.R. Methods for Plasmid and Genomic DNA Isolation from Lactobacilli. In Food Microbiology Protocols, Methods in Biotechnology, 1st ed.; Spencer, J.F.T., de Ragout Spencer, A.L., Eds.; Humana Press: Totowa, NJ, USA, 2001; Volume 14, pp. 135–137. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, W.; Wang, H.; Geng, A. Polyhydroxybutyrate Production from Oil Palm Empty Fruit Bunch Using Bacillus megaterium R11. Bioresour. Technol. 2013, 147, 307–314. [Google Scholar] [CrossRef]
- Singh, G.; Kumari, A.; Mittal, A.; Yadav, A.; Aggarwal, N.K. Poly β-Hydroxybutyrate Production by Bacillus subtilis NG220 Using Sugar Industry Waste Water. Biomed Res. Int. 2013, 2013, 952641. [Google Scholar] [CrossRef]
- Hahn, S.K.; Chang, Y.K.; Lee, S.Y. Recovery and Characterization of Poly(3-Hydroxybutyric Acid) Synthesized in Alcaligenus eutrophus and Recombinant Escherichia coli. Appl. Environ. Microbiol. 1995, 61, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Sikkema, W.D.; Cal, A.J.; Hathwaik, U.I.; Orts, W.J.; Lee, C.C. Polyhydroxyalkanoate Production in Pseudomonas putida from Alkanoic Acids of Varying Lengths. PLoS ONE 2023, 18, e0284377. [Google Scholar] [CrossRef]
- Gouda, M.K.; Swellam, A.E.; Omar, S.H. Production of PHB by a Bacillus megaterium Strain Using Sugarcane Molasses and Corn Steep Liquor as Sole Carbon and Nitrogen Sources. Microbiol. Res. 2001, 156, 201–207. [Google Scholar] [CrossRef]
- Pernicova, I.; Novackova, I.; Sedlacek, P.; Kourilova, X.; Kalina, M.; Kovalcik, A.; Koller, M.; Nebesarova, J.; Krzyzanek, V.; Hrubanova, K.; et al. Introducing the Newly Isolated Bacterium Aneurinibacillus sp. H1 as an Auspicious Thermophilic Producer of Various Polyhydroxyalkanoates (PHA) Copolymers–1. Isolation and Characterization of the Bacteria. Polymers 2020, 12, 1235. [Google Scholar] [CrossRef] [PubMed]
- Muigano, M.N.; Anami, S.E.; Onguso, J.M.; Omare, G.M. The Isolation, Screening, and Characterization of Polyhydroxyalkanoate-Producing Bacteria from Hypersaline Lakes in Kenya. Bacteria 2023, 2, 81–97. [Google Scholar] [CrossRef]
- Mizuno, K.; Ohta, A.; Hyakutake, M.; Ichinomiya, Y.; Tsuge, T. Isolation of Polyhydroxyalkanoate-Producing Bacteria from a Polluted Soil and Characterization of the Isolated Strain Bacillus cereus YB-4. Polym. Degrad. Stab. 2010, 95, 1335–1339. [Google Scholar] [CrossRef]
- Sehgal, R.; Gupta, R. Polyhydroxyalkanoate and Its Efficient Production: An Eco-Friendly Approach toward Development. 3 Biotech 2020, 10, 549. [Google Scholar] [CrossRef]
- Sriyapai, T.; Chuarung, T.; Kimbara, K.; Samosorn, S.; Sriyapai, P. Production and Optimization of Polyhydroxyalkanoates (PHAs) from Paraburkholderia sp. PFN 29 under Submerged Fermentation. Electron. J. Biotechnol. 2022, 56, 1–11. [Google Scholar] [CrossRef]
- Koller, M.; Salerno, A.; Muhr, A.; Reiterer, A.; Schober, S.; Braunegg, G. Modern Biotechnological Polymer Synthesis: A Review. Food Technol. Biotechnol. 2013, 51, 12–25. [Google Scholar]
- Kourmentza, C.; Plácido, J.; Venetsaneas, N.; Burniol-Figols, A.; Varrone, C.; Gavala, H.N.; Reis, M.A. Recent Advances and Challenges toward Sustainable Polyhydroxyalkanoate (PHA) Production. Bioengineering 2017, 4, 55. [Google Scholar] [CrossRef]
- Jiang, G.; Hill, D.J.; Kowalczuk, M.; Johnston, B.; Adamus, G.; Irorere, V.; Radecka, I. Carbon Sources for Polyhydroxyalkanoates and an Integrated Biorefinery. Int. J. Mol. Sci. 2016, 17, 1157. [Google Scholar] [CrossRef]
- Lau, N.S.; Sudesh, K. Revelation of the Ability of Burkholderia sp. USM (JCM 15050) PHA Synthase to Polymerize 4-Hydroxybutyrate Monomer. AMB Express 2012, 2, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chavan, S.; Yadav, B.; Tyagi, R.D.; Drogui, P. A Review on Production of Polyhydroxyalkanoate (PHA) Biopolyesters by Thermophilic Microbes Using Waste Feedstocks. Bioresour. Technol. 2021, 341, 125900. [Google Scholar] [CrossRef] [PubMed]
- Thu, N.T.T.; Hoang, L.H.; Cuong, P.K.; Viet-Linh, N.; Nga, T.T.H.; Kim, D.D.; Yoong, K.L.; Le, T.N. Evaluation of Polyhydroxyalkanoate (PHA) Synthesis by Pichia sp. TSLS24 Yeast Isolated in Vietnam. Sci. Rep. 2023, 13, 3137. [Google Scholar] [CrossRef]
- Ryu, H.W.; Hahn, S.K.; Chang, Y.K.; Chang, H.N. Production of Poly(3-hydroxybutyrate) by High Cell Density Fed-Batch Culture of Alcaligenes eutrophus with Phosphate Limitation. Biotechnol. Bioeng. 1997, 55, 28–32. [Google Scholar] [CrossRef]
- Karmann, S.; Panke, S.; Zinn, M. Fed-Batch Cultivations of Rhodospirillum rubrum Under Multiple Nutrient-Limited Growth Conditions on Syngas as a Novel Option to Produce Poly(3-Hydroxybutyrate) (PHB). Front. Bioeng. Biotechnol. 2019, 7, 59. [Google Scholar] [CrossRef] [PubMed]
- Yootoum, A.; Jantanasakulwong, K.; Rachtanapun, P.; Moukamnerd, C.; Chaiyaso, T.; Pumas, C.; Nuttapol, T.; Masanori, W.; Toshiaki, F.; Chayatip, I. Characterization of Newly Isolated Thermotolerant Bacterium Cupriavidus sp. CB15 from Composting and Its Ability to Produce Polyhydroxyalkanoate from Glycerol. Microb. Cell Fact. 2023, 22, 68. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, V.M.; Silva, L.F.; Gomez, J.G.C.; Fonseca, G.G. Growth of Burkholderia sacchari LFM 101 cultivated in glucose, sucrose and glycerol at different temperatures. Sci. Agric. 2016, 73, 429–433. [Google Scholar] [CrossRef]
- Brandl, H.; Gross, R.A.; Lenz, R.W.; Fuller, R.C. Plastics from bacteria and for bacteria: Poly(3-hydroxyalkanoates) as natural, biocompatible, and biodegradable polyesters. Adv. Biochem. Eng. Biotechnol. 1990, 41, 77–93. [Google Scholar] [CrossRef]
- Blunt, W.; Levin, D.B.; Cicek, N. Bioreactor Operating Strategies for Improved Polyhydroxyalkanoate (PHA) Productivity. Polymers 2018, 10, 1197. [Google Scholar] [CrossRef]
- Raza, Z.A.; Tariq, M.R.; Majeed, M.I.; Banat, I.M. Recent Developments in Bioreactor Scale Production of Bacterial Polyhydroxyalkanoates. Bioprocess Biosyst. Eng. 2019, 42, 901–919. [Google Scholar] [CrossRef] [PubMed]
- Mahato, R.P.; Kumar, S.; Singh, P. Optimization of Growth Conditions to Produce Sustainable Polyhydroxyalkanoate Bioplastic by Pseudomonas aeruginosa EO1. Front. Microbiol. 2021, 12, 711588. [Google Scholar] [CrossRef] [PubMed]
- Madhusoodanan, G.; Hariharapura, R.C.; Somashekara, D. Dissolved Oxygen as a Propulsive Parameter for Polyhydroxyalkanoate Production Using Bacillus endophyticus Cultures. Environ. Dev. Sustain. 2022, 24, 4641–4658. [Google Scholar] [CrossRef]
- Możejko-Ciesielska, J.; Ray, S.; Sankhyan, S. Recent Challenges and Trends of Polyhydroxyalkanoate Production by Extremophilic Bacteria Using Renewable Feedstocks. Polymers 2023, 15, 4385. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Lee, S.M.; Park, S.L.; Choi, T.R.; Song, H.S.; Kim, H.J.; Bhatia, S.K.; Gurav, R.; Kim, Y.G.; Kim, J.H.; et al. Tung Oil-Based Production of High 3-Hydroxyhexanoate-Containing Terpolymer Poly (3-Hydroxybutyrate-co-3-Hydroxyvalerate-co-3-Hydroxyhexanoate) Using Engineered Ralstonia eutropha. Polymers 2021, 13, 1084. [Google Scholar] [CrossRef]
- Zhu, Y.; Ai, M.; Jia, X. Optimization of a Two-Species Microbial Consortium for Improved Mcl-PHA Production from Glucose–Xylose Mixtures. Front. Bioeng. Biotechnol. 2022, 9, 794331. [Google Scholar] [CrossRef]
- Huang, J.; Okoshi, T.; Mizuno, S.; Hiroe, A.; Tsuge, T. Gas Chromatography-Mass Spectrometry-Based Monomer Composition Analysis of Medium-Chain-Length Polyhydroxyalkanoates Biosynthesized by Pseudomonas spp. Biosci. Biotechnol. Biochem. 2018, 82, 1615–1623. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Neobacillus niacini PHA (g/L) | Metabacillus niabensis PHA (g/L) | |
|---|---|---|
| Carbon Sources | ||
| Sunflower oil | 1.1 ± 0.06 | 0.76 ± 0.01 |
| Waste olive oil | 0.13 ± 0.06 | 0.84 ± 0.05 |
| Nitrogen Sources | ||
| Yeast extract | 0.84 ± 0.04 | 0.33 ± 0.06 |
| Urea | 0.38 ± 0.02 | 0.76 ± 0.01 |
| pH | ||
| pH 7 | 0.75 ± 0.09 | 0.73 ± 0.05 |
| Temperature | ||
| 35 °C | 0.81 ± 0.04 | 0.77 ± 0.06 |
| Agitation | ||
| 100 rpm | 1.13 ± 0.12 | 0.75 ± 0.07 |
| 150 rpm | 0.5 ± 0.1 | 0.85 ± 0.07 |
| Fermentation Process | Fermentation Period (h) | Specific Growth Rate (µ) | CDW (g/L) | PHA (g/L) | YP/S (g/g) | Yx/S (g/g) | Biomass Vol. Prod. (g/L/h) | PHA Vol. Prod. (g/L/h) |
|---|---|---|---|---|---|---|---|---|
| Uncontrolled pH batch | 72 | 0.07 | 2 | 1.75 | 0.25 | 0.15 | 0.02 | 0.035 |
| Controlled pH batch | 72 | 0.08 | 4 | 9.75 | 0.6 | 1.62 | 0.05 | 0.13 |
| Fermentation Process | Fermentation Period (h) | Specific Growth Rate (µ) | CDW (g/L) | PHA (g/L) | YP/S (g/g) | Yx/S (g/g) | Biomass Vol. Prod. (g/L/h) | PHA Vol. Prod. (g/L/h) |
|---|---|---|---|---|---|---|---|---|
| Uncontrolled pH batch | 48 | 0.11 | 2 | 1.75 | 0.17 | 0.2 | 0.04 | 0.036 |
| Controlled pH batch | 48 | 0.11 | 2.6 | 4.78 | 0.6 | 0.32 | 0.054 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elazzazy, A.M.; Ali Abd, K.; Bataweel, N.M.; Mahmoud, M.M.; Baghdadi, A.M. Microbial Biosynthesis of Medium-Chain-Length Polyhydroxyalkanoate (mcl-PHA) from Waste Cooking Oil. Polymers 2024, 16, 2150. https://doi.org/10.3390/polym16152150
Elazzazy AM, Ali Abd K, Bataweel NM, Mahmoud MM, Baghdadi AM. Microbial Biosynthesis of Medium-Chain-Length Polyhydroxyalkanoate (mcl-PHA) from Waste Cooking Oil. Polymers. 2024; 16(15):2150. https://doi.org/10.3390/polym16152150
Chicago/Turabian StyleElazzazy, Ahmed M., Khawater Ali Abd, Noor M. Bataweel, Maged M. Mahmoud, and Afra M. Baghdadi. 2024. "Microbial Biosynthesis of Medium-Chain-Length Polyhydroxyalkanoate (mcl-PHA) from Waste Cooking Oil" Polymers 16, no. 15: 2150. https://doi.org/10.3390/polym16152150