Response of Ancient and Modern Wheat Varieties to Biochar Application: Effect on Hormone and Gene Expression Involved in Germination and Growth


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description and Analysis of Samples
2.2. Seed Germination
2.3. Hormone Extraction and Analysis
2.4. Data Analysis
2.5. RNA Extraction, cDNAs Synthesis and Reverse Transcription-Quantitative PCR (RT-qPCR)
3. Results and Discussion
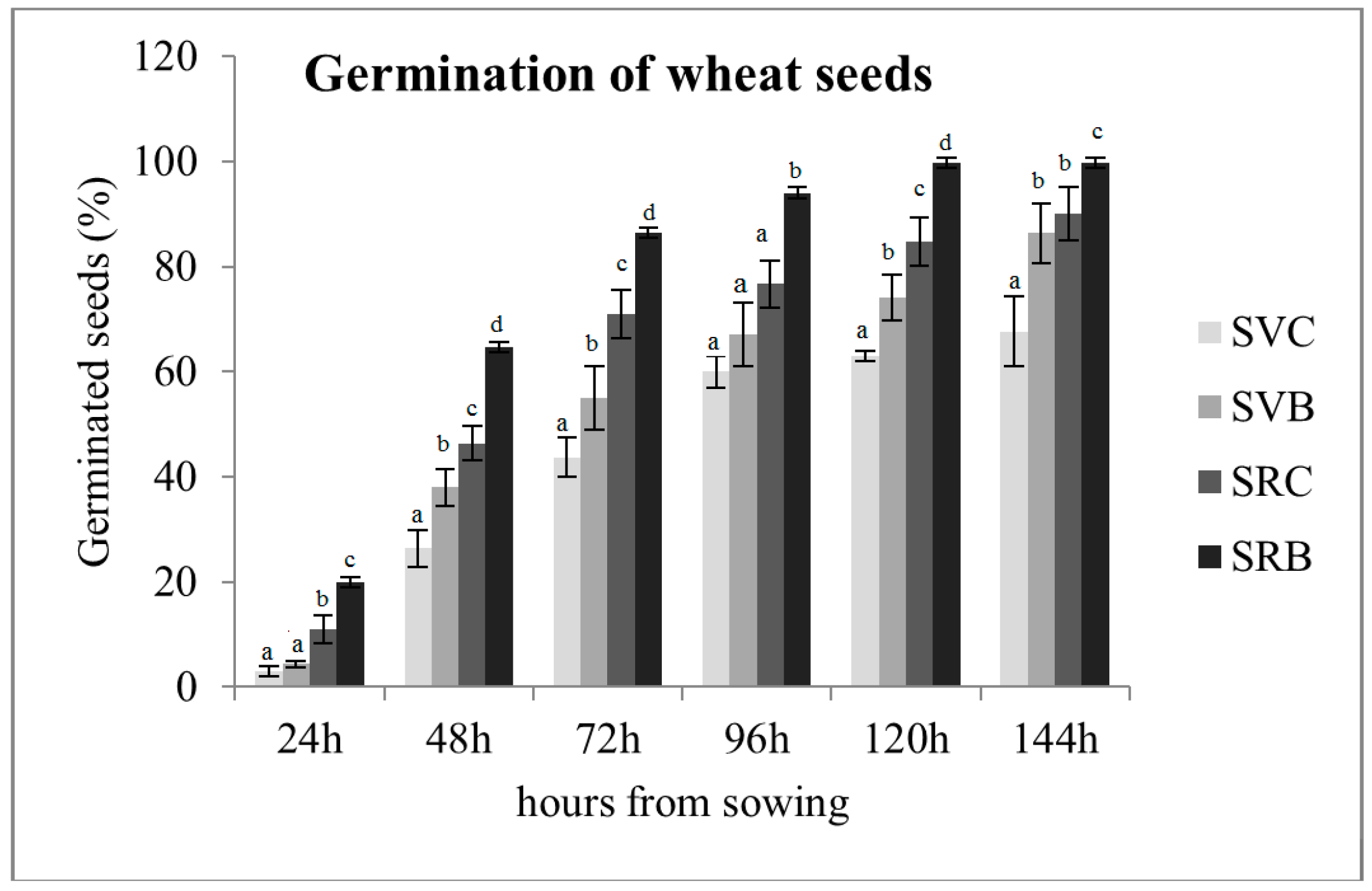
3.1. Effect of Biochar on the Germination of Svevo and Saragolla Seeds
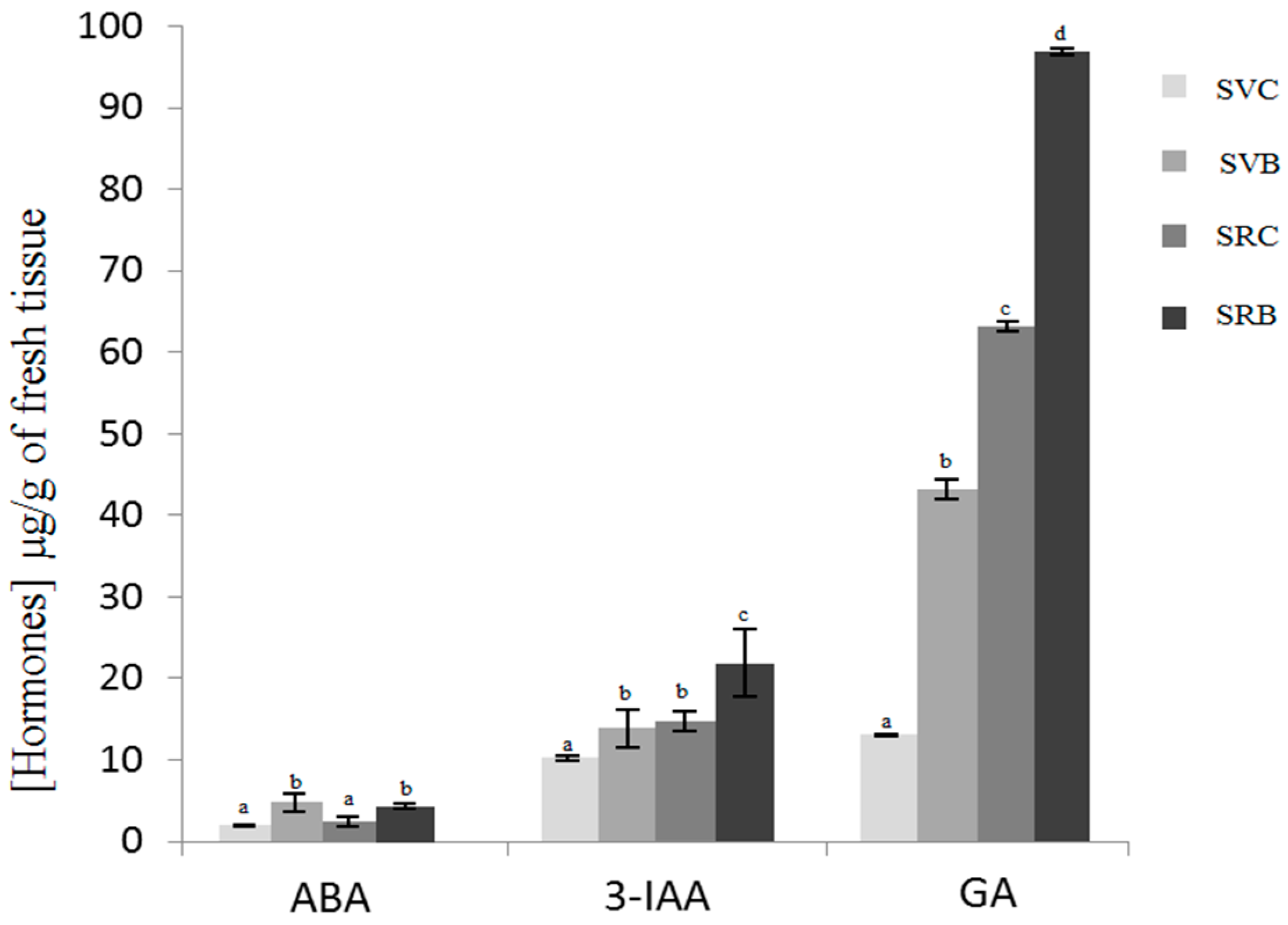
3.2. Effect of Biochar on the Hormone Content of Svevo and Saragolla Seedlings
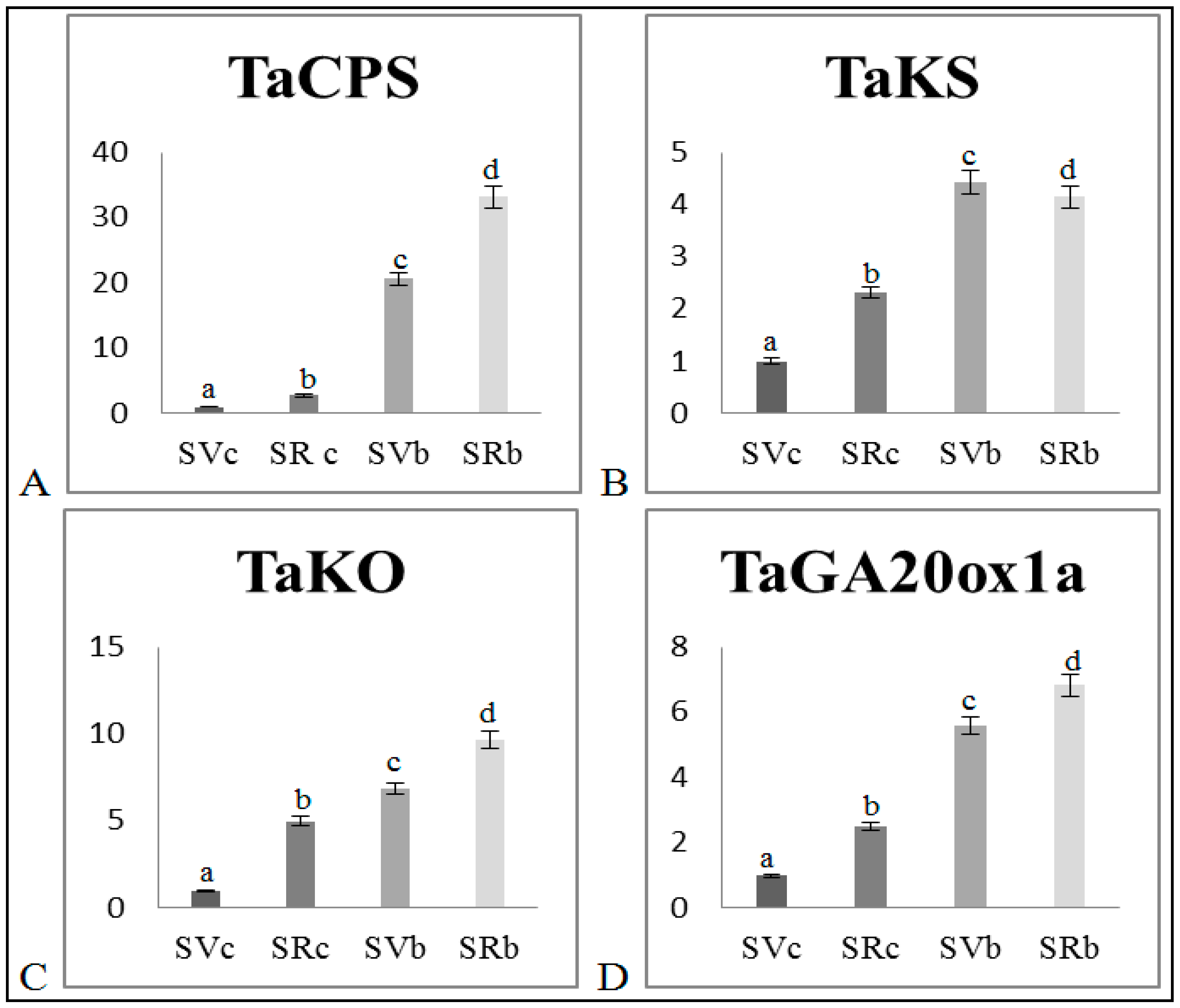
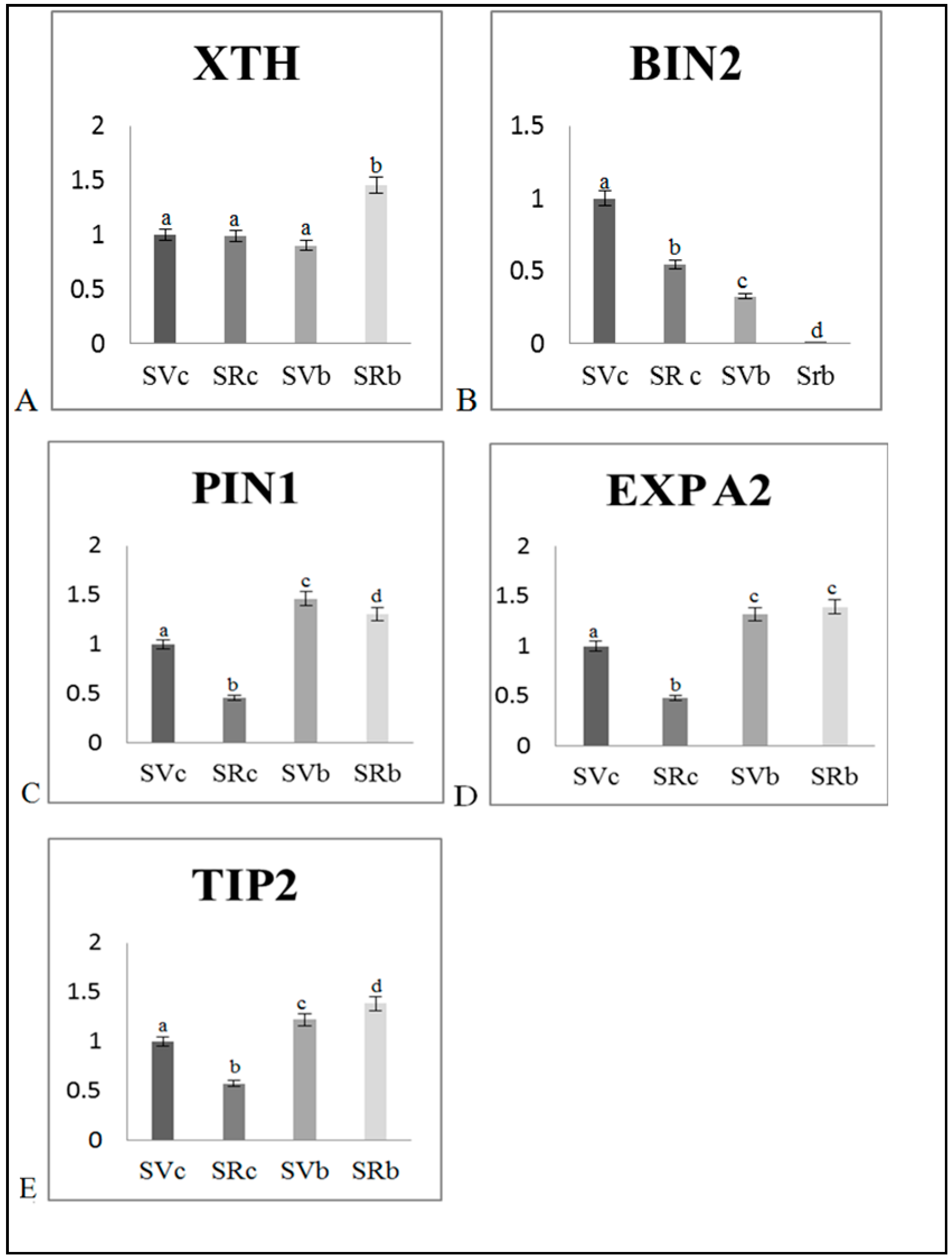
3.3. Effect of Biochar on Gene Transcription
GA Biosynthesis Genes
Author Contributions
Funding
Conflicts of Interest
References
- Călinoiu, L.F.; Vodnar, D. Whole grains and phenolic acids: A review on bioactivity, functionality, health benefits and bioavailability. Nutrients 2018, 10, 1615. [Google Scholar] [CrossRef] [Green Version]
- Hamdi, L.; Suleiman, A.; Hoogenboom, G.; Shelia, V. Response of the Durum Wheat Cultivar Um Qais (Triticum turgidum subsp. durum) to Salinity. Agriculture 2019, 9, 135. [Google Scholar] [CrossRef] [Green Version]
- Guarda, G.; Padovan, S.; Delogu, G. Grain yield, nitrogen-use efficiency and baking quality of old and modern Italian bread-wheat cultivars grown at different nitrogen levels. Eur. J. Agron. 2004, 21, 181–192. [Google Scholar] [CrossRef]
- Martini, D.; Taddei, F.; Ciccoritti, R.; Pasquini, M.; Nicoletti, I.; Corradini, D.; D’Egidio, M.G. Variation of Total Antioxidant Activity and of Phenolic Acid, Total Phenolics and Yellow Coloured Pigments in Durum Wheat (Triticum turgidum L. Var. durum) as a Function of Genotype, Crop Year and Growing Area. J. Cereal Sci. 2015, 65, 175–185. [Google Scholar] [CrossRef]
- Fernández, J.M.; Hernández, D.; Plaza, C.; Polo, A. Organic matter in degraded agricultural soil amended with composted and thermally-dried sewage sludge. Sci. Total Environ. 2008, 378, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Paneque, M.; De la Rosa, J.M.; Franco-Navarro, J.; Colmenero-Flores, J.M.; Knicker, H. Effect of biochar amendment on morphology, productivity and water relations of sun-flower plants under non-irrigation conditions. Catena 2016, 147, 280–287. [Google Scholar] [CrossRef] [Green Version]
- Howden, S.M.; Soussana, J.-F.; Tubiello, F.N.; Chhetri, N.; Dunlop, M.; Meinke, H. Adapting agriculture to climate change. Proc. Natl. Acad. Sci. USA 2007, 104, 19691–19696. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Zhang, Z.; Zhu, C.; Zhai, Y.; Lu, P. Effect of biochar on sweet corn and soil salinity under conjunctive irrigation with brackish water in coastal saline soil. Sci. Hortic. 2019, 250, 405–413. [Google Scholar] [CrossRef]
- Álvarez, J.M.; Pasian, C.; Lala, R.; López, R.; Díaz, M.J. Morpho-physiological plant quality when biochar and vermicompost are used as growing media replacement in urban horticulture. Urban For. Urban Green. 2018, 34, 175–180. [Google Scholar] [CrossRef]
- De la Rosa, J.M.; Rosado, M.; Paneque, M.; Miller, A.Z.; Knicker, H. Effects of aging under field conditions on biochar structure and composition: Implications for biochar stability in soils. Sci. Total Environ. 2018, 613–614, 969–976. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Voroney, R.P.; Price, G.W. Effects of temperature and processing conditions on biochar chemical properties and their influence on soil C and N transformations. Soil Biol. Biochem. 2015, 351, 263–265. [Google Scholar] [CrossRef]
- Schmidt, H.P.; Pandit, B.H.; Martinsen, V.; Cornelissen, G.; Conte, P.; Kammann, C.I. Fourfold increase in pumpkin yield in response to low-dosage root zone application of urine-enhanced biochar to a fertile tropical soil. Agriculture 2015, 5, 723–741. [Google Scholar] [CrossRef] [Green Version]
- Dari, B.; Nair, V.D.; Harris, W.G.; Nair, P.K.R.; Sollenberger, L.; Mylavarapu, R. Relative influence of soil-vs. biochar properties on soil phosphorus retention. Geoderma 2016, 280, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.; Farooq, M.; Nawaz, A.; Al-Sadi, A.M.; Solaiman, Z.M.; Alghamdi, S.S.; Ammara, U.; Ok, Y.S.; Kadambot, H.; Siddique, M. Biochar for crop production: Potential benefits and risks. J. Soils Sediments 2016, 17, 1–32. [Google Scholar] [CrossRef]
- Graziano, S.; Marando, S.; Prandi, B.; Boukid, F.; Marmiroli, N.; Francia, E.; Pecchioni, N.; Sforza, S.; Visioli, G.; Gullì, M. Technological Quality and Nutritional Value of Two Durum Wheat Varieties Depend on Both Genetic and Environmental Factors. J. Agric. Food Chem. 2019, 67, 2384–2395. [Google Scholar] [CrossRef]
- French, E.A.; Iyer-Pascuzzi, A.S. A role for the gibberellin pathway in biochar-mediated growth promotion. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Torabian, S. Biochar Increased Plant Growth-Promoting Hormones and Helped to Alleviates Salt Stress in Common Bean Seedlings. J. Plant Growth Regul. 2018, 37, 591–601. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.; Behrendt, U.; Abd-Allah, E.F.; Berg, G. Biochar treatment resulted in a combined effect on soybean growth promotion and a shift in plant growth promoting rhizobacteria. Front. Microbiol. 2016, 7, 209. [Google Scholar] [CrossRef] [Green Version]
- Obia, A.; Cornelissen, G.; Mulder, J.; Dörsch, P. Effect of soil pH increase by biochar on NO, N2O and N2 production during denitrification in acid soils. PLoS ONE 2015, 10, e0138781. [Google Scholar] [CrossRef] [Green Version]
- De la Rosa, J.M.; Paneque, M.; Miller, A.Z.; Knicker, H. Relating physical and chemical properties of four different biochars and their application rate to biomass production of Lolium perenne on a Calcic Cambisol during a pot experiment of 79 days. Sci. Total Environ. 2014, 499, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Jeffrey, S.; Verheijen, F.; van der Velde, M.; Bastos, A.C. A quantitative review of the effects of biochar application to soils on crop productivity using meta-analysis. 2011. Agric. Ecosyst. Environ. 2011, 144, 175–187. [Google Scholar] [CrossRef]
- Bista, P.; Ghimire, R.; Machado, S.; Pritchett, L. Biochar Effects on Soil Properties and Wheat Biomass vary with Fertility Management. Agronomy 2019, 9, 623. [Google Scholar] [CrossRef] [Green Version]
- Barros, A.C.; Freund, M.T.; Villavicencio, L.; Delincee, A.L.C.H.; Arthura, H. Identification of irradiated wheat by germination test, DNA comet assay and electron spin. Radiat. Phys. Chem. 2002, 63, 423–426. [Google Scholar] [CrossRef]
- Rocco, M.; Tartaglia, M.; Izzo, F.P.; Varricchio, E.; Arena, S.; Scaloni, A.; Marra, M. Comparative proteomic analysis of durum wheat shoots from modern and ancient cultivars. Plant Physiol. Biochem. 2019, 135, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)). Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Camoni, L.; Visconti, S.; Aducci, P.; Marra, M. 14-3-3 Proteins in Plant Hormone Signaling: Doing Several Things at Once. Front. Plant Sci. 2018, 9, 297. [Google Scholar] [CrossRef]
- Guangwu, Z.; Xuwen, J. Roles of gibberellin and Auxin in promoting seed germination and seedling Vigor in Pinus massoniana. For. Sci. 2014, 60, 367–369. [Google Scholar] [CrossRef]
- Vishal, B.; Kumar, P.P. Regulation of seed germination and abiotic stresses by gibberellins and abscisic acid. Front. Plant Sci. 2018, 9, 838. [Google Scholar] [CrossRef]
- Viger, M.; Hancock, R.D.; Miglietta, F.; Taylor, G. More plant growth but less plant defence? First global gene expression data for plants grown in soil amended with biochar. GCB Bioenergy 2015, 7, 658–672. [Google Scholar] [CrossRef]
- Zhang, Y.; Ni, Z.; Yao, Y.; Nie, X.; Sun, Q. Gibberellins and heterosis of plant height in wheat (Triticum aestivum L.). BMC Genet. 2007, 8, 40. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Campbell, P.; Vargheese, A.K.; Braam, J. The Arabidopsis XET-related gene family: Environmental and hormonal regulation of expression. Plant J. 1996, 9, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Vissenberg, K.; Oyama, M.; Osato, Y.; Yokoyama, R.; Verbelen, J.P.; Nishitani, K. Differential expression of AtXTH17, AtXTH18, AtXTH19 and AtXTH20 genes in Arabidopsis roots. Physiological roles in specification in cell wall construction. Plant Cell Physiol. 2005, 46, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rodríguez, C.; Rubio-Somoza, I.; Sibout, R.; Persson, S. Phytohormones and the cell wall in Arabidopsis during seedling growth. Trends Plant Sci. 2010, 15, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Singla, B.; Tyagi, A.K.; Khurana, J.P.; Khurana, P. Analysis of expression profile of selected genes expressed during auxin-induced somatic embryogenesis in leaf base system of wheat (Triticum aestivum) and their possible interactions. Plant Mol. Biol. 2007, 65, 677–692. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.W.; Lee, S.M.; Joo, S.H.; Yun, H.S.; Lee, Y.; Kaufman, P.B.; Kirakosyan, A.; Kim, S.H.; Nam, K.H.; Lee, J.S.; et al. Elongation and gravitropic responses of Arabidopsis roots are regulated by brassinolide and IAA. Plant Cell Environ. 2007, 30, 679–689. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | pH | % C | % N | % WHC | % Ash |
|---|---|---|---|---|---|
| Soil | 5.62 | 3.1 | 0.16 | 35.9 | 88.0 |
| Biochar | 8.21 | 81.1 | 0.91 | 364.4 | 7.7 |
| Soil + Biochar (5%) | 7.10 | n.a. | n.a. | 53.4 | n.a. |
| Primers | Sequences |
|---|---|
| β-ACTIN | F: TGGACTCTGGTGATGGTGTC R: CCTCCAATCCAAACACTGTA |
| BIN2 | F: GAGATCTAAAGCCTCAAAATCTT R: TGGCTTCACCTTTAACGAGCT |
| PIN1 | F: ATCATCTGGTACACGCTCAT R: GGGAACTGCTCGGTTGAT |
| TIP2 | F: GATGACTCCTTCAGCTTGG R: GGCGAAGACGAAGATGAG |
| XTH | F:GCCCTTCGTCGCCTCCTAC R: CGGCACAACAACAACTAGTGGTAG |
| EXP A2 | F: CCACCATGATGTGTTGTTCC R: AGTAGGAGTGGCCGTTGATG |
| TaCPS | F: GTATGCAAGCTTACCGCGTG R: ACCCCCACAAGAATGTCCTC |
| TaKS | F: CAGGCCGGGGAGAAATCTT R: TGAGACAGCTCATCTGGGGA |
| TaKO | F: CCGGCACCGAGATAGTCATC R: GAGCAAATCCAGCACCTCAT |
| TaGA20ox1a | F: CCATCCTCCACCAGGACAAC R: GAGCTCCATCCTCTGTCTGG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Racioppi, M.; Tartaglia, M.; De la Rosa, J.M.; Marra, M.; Lopez-Capel, E.; Rocco, M. Response of Ancient and Modern Wheat Varieties to Biochar Application: Effect on Hormone and Gene Expression Involved in Germination and Growth. Agronomy 2020, 10, 5. https://doi.org/10.3390/agronomy10010005
Racioppi M, Tartaglia M, De la Rosa JM, Marra M, Lopez-Capel E, Rocco M. Response of Ancient and Modern Wheat Varieties to Biochar Application: Effect on Hormone and Gene Expression Involved in Germination and Growth. Agronomy. 2020; 10(1):5. https://doi.org/10.3390/agronomy10010005
Chicago/Turabian StyleRacioppi, Marco, Maria Tartaglia, José María De la Rosa, Mauro Marra, Elisa Lopez-Capel, and Mariapina Rocco. 2020. "Response of Ancient and Modern Wheat Varieties to Biochar Application: Effect on Hormone and Gene Expression Involved in Germination and Growth" Agronomy 10, no. 1: 5. https://doi.org/10.3390/agronomy10010005
APA StyleRacioppi, M., Tartaglia, M., De la Rosa, J. M., Marra, M., Lopez-Capel, E., & Rocco, M. (2020). Response of Ancient and Modern Wheat Varieties to Biochar Application: Effect on Hormone and Gene Expression Involved in Germination and Growth. Agronomy, 10(1), 5. https://doi.org/10.3390/agronomy10010005