Root and Agro-Morphological Traits Performance in Cowpea under Drought Stress


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
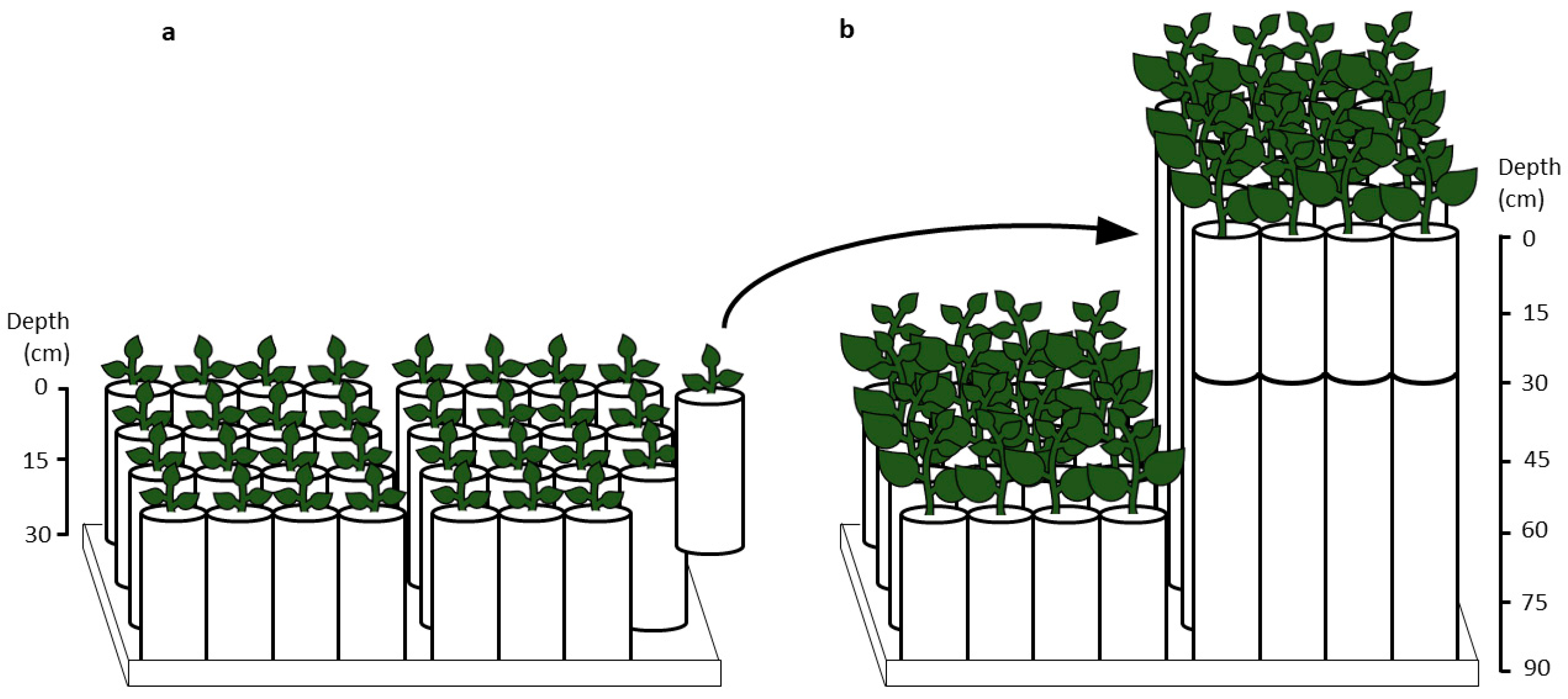
2.1. Plant Material and Experimental Design
2.2. Climate Data and Soil Water Content
2.3. Data Collection
2.4. Data Analysis
3. Results and Discussion
3.1. Soil Water Conditions
3.2. Evaluation of Agro-Morphological Parameters
3.2.1. Shoot-Related Parameters
3.2.2. Root-Related Parameters
3.3. Principal Component Analysis
3.4. Screening for Drought Tolerant Genotypes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huang, S.; Li, P.; Huang, Q.; Leng, G.; Hou, B.; Ma, L. The propagation from meteorological to hydrological drought and its potential influence factors. J. Hydrol. 2017, 547, 184–195. [Google Scholar] [CrossRef]
- Leng, G.; Hall, J. Crop yield sensitivity of global major agricultural countries to droughts and the projected changes in the future. Sci. Total Environ. 2019, 654, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Singh, H.; Raghubanshi, A.S. Challenges and opportunities for agricultural sustainability in changing climate scenarios: A perspective on Indian agriculture. Trop. Ecol. 2019, 60, 167–185. [Google Scholar] [CrossRef]
- FAOSTAT Food and Agriculture Organization of the United Nations—Statistics Division. Available online: http://faostat3.fao.org/browse/Q/QC/E (accessed on 18 February 2020).
- Watson, C.A.; Reckling, M.; Preissel, S.; Bachinger, J.; Bergkvist, G.; Kuhlman, T.; Lindström, K.; Nemecek, T.; Topp, C.F.; Vanhatalo, A.; et al. Grain Legume Production and Use in European Agricultural Systems. In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 2017; Volume 144, pp. 235–303. [Google Scholar]
- Gonçalves, A.; Goufo, P.; Barros, A.; Domínguez-Perles, R.; Trindade, H.; Rosa, E.A.; Ferreira, L.; Rodrigues, M. Cowpea (Vigna unguiculata L. Walp), a renewed multipurpose crop for a more sustainable agri-food system: Nutritional advantages and constraints. J. Sci. Food Agric. 2016, 96, 2941–2951. [Google Scholar] [CrossRef] [PubMed]
- Ofuya, Z.; Akhidue, V. The role of pulses in human nutrition: A review. J. Appl. Sci. Environ. Manag. 2006, 9, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Carsky, R.J.; Vanlauwe, B.; Lyasse, O. Cowpea rotation as a resource management technology for cereal-based systems in the savannas of West Africa. In Challenges and Opportunities for Enhancing Sustainable Cowpea Production; Fatokun, C.A., Tarawali, S.A., Singh, B.B., Kormawa, P.M., Tamò, M., Eds.; International Institute of Tropical Agriculture: Ibadan, Nigeria, 2002; pp. 251–266. [Google Scholar]
- Hall, A.E. Breeding for adaptation to drought and heat in cowpea. Eur. J. Agron. 2004, 21, 447–454. [Google Scholar] [CrossRef]
- Carvalho, M.; Lino-Neto, T.; Rosa, E.; Carnide, V. Cowpea: A legume crop for a challenging environment. J. Sci. Food Agric. 2017, 97, 4273–4284. [Google Scholar] [CrossRef]
- Herniter, I.; Muñoz-Amatriaín, M.; Close, T. Genetic, textual, and archaeological evidence of the historical global spread of cowpea (Vigna unguiculata [L.] Walp). Legume Sci. 2020, e57. [Google Scholar] [CrossRef]
- Agbicodo, E.M.; Fatokun, C.A.; Muranaka, S.; Visser, R.G.F.; Van Der, C.G.L. Breeding drought tolerant cowpea: Constraints, accomplishments, and future prospects. Euphytica 2009, 167, 353–370. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, M.; Matos, M.; Castro, I.; Monteiro, E.; Rosa, E.; Lino-Neto, T.; Carnide, V. Screening of worldwide cowpea collection to drought tolerant at a germination stage. Sci. Hortic. 2019, 247, 107–115. [Google Scholar] [CrossRef]
- Muchero, W.; Ehlers, J.D.; A Roberts, P. Seedling Stage Drought-Induced Phenotypes and Drought-Responsive Genes in Diverse Cowpea Genotypes. Crop. Sci. 2008, 48, 541–552. [Google Scholar] [CrossRef]
- Iseki, K.; Takahashi, Y.; Muto, C.; Naito, K.; Tomooka, N. Diversity of Drought Tolerance in the Genus Vigna. Front. Plant Sci. 2018, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Singh, B.B. Root Characteristics in Cowpea Related to Drought Tolerance at the Seedling Stage. Exp. Agric. 2003, 39, 29–38. [Google Scholar] [CrossRef]
- Wasaya, A.; Zhang, X.; Fang, Q.; Yan, Z. Root Phenotyping for Drought Tolerance: A Review. Agronomy 2018, 8, 241. [Google Scholar] [CrossRef] [Green Version]
- Ramamoorthy, P.; Lakshmanan, K.; Upadhyaya, H.D.; Vadez, V.; Varshney, R.K. Root traits confer grain yield advantages under terminal drought in chickpea (Cicer arietinum L.). Field Crop. Res. 2017, 201, 146–161. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, M.; Bebeli, P.J.; Pereira, G.; Castro, I.; Egea-Gilabert, C.; Matos, M.; Lazaridi, E.; Duarte, I.; Lino-Neto, T.; Ntatsi, G.; et al. European cowpea landraces for a more sustainable agriculture system and novel foods. J. Sci. Food Agric. 2017, 97, 4399–4407. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.; Castro, I.; Matos, M.; Lino-Neto, T.; Silva, V.; Rosa, E.; Carnide, V. Caracterização agro-morfológica de acessos de feijão-frade (Vigna unguiculata): Bases para o melhoramento. Rev. Ciências Agrárias 2016, 39, 506–517. [Google Scholar] [CrossRef] [Green Version]
- Martos-Fuentes, M.; Cellini, A.; Ochoa, J.; Carvalho, M.; Carnide, V.; Rosa, E.; Pereira, G.; Barcelos, C.; Bebeli, P.J.; Egea-Gilabert, C. Genotype by environment interactions in cowpea (Vigna unguiculata L. Walp.) grown in the Iberian Peninsula. Crop. Pasture Sci. 2017, 68, 924–931. [Google Scholar] [CrossRef]
- Muñoz-Perea, C.G.; Allen, R.G.; Westermann, D.T.; Wright, J.L.; Singh, S.P. Water use efficiency among dry bean landraces and cultivars in drought-stressed and non-stressed environments. Euphytica 2007, 155, 393–402. [Google Scholar] [CrossRef]
- Mfeka, N.; Mulidzi, A.; Lewu, F.B. Growth and yield parameters of three cowpea (Vigna unguiculata L. Walp) lines as affected by planting date and zinc application rate. S. Afr. J. Sci. 2018, 115, 1–8. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAS: Paleontological Statistics Software Package for Education and Data Analysis. Palaentologia Electron. 2001, 4, 1–9. [Google Scholar]
- Carvalho, M.; Castro, I.; Moutinho-Pereira, J.; Correia, C.; Egea-Cortines, M.; Matos, M.; Rosa, E.A.; Carnide, V.; Lino-Neto, T. Evaluating stress responses in cowpea under drought stress. J. Plant Physiol. 2019, 241, 153001. [Google Scholar] [CrossRef] [PubMed]
- Gebre, M.G.; Earl, H.J. Effects of Growth Medium and Water Stress on Soybean [Glycine max (L.) Merr.] Growth, Soil Water Extraction and Rooting Profiles by Depth in 1-m Rooting Columns. Front. Plant Sci. 2020, 11, 11. [Google Scholar] [CrossRef] [PubMed]
- Kashiwagi, J.; Krishnamurthy, L.; Crouch, J.; Serraj, R. Variability of root length density and its contributions to seed yield in chickpea (Cicer arietinum L.) under terminal drought stress. Field Crop. Res. 2006, 95, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Zegaoui, Z.; Planchais, S.; Cabassa, C.; Djebbar, R.; Belbachir, O.A.; Carol, P. Variation in relative water content, proline accumulation and stress gene expression in two cowpea landraces under drought. J. Plant Physiol. 2017, 218, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Agele, S.O.; Ofuya, T.I.; James, P.O. Effects of watering regimes on aphid infestation and performance of selected varieties of cowpea (Vigna unguiculata L. Walp) in a humid rainforest zone of Nigeria. Crop. Prot. 2006, 25, 73–78. [Google Scholar] [CrossRef]
- Mendes, R.M.S.; Távora, J.A.; José, F.; Pitombeira, J.B.; Nogueira, J.M. Source-sink relationships in cowpea under drought stress. Rev. Ciência Agronômica 2007, 38, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Nadeem, M.; Li, J.; Yahya, M.; Sher, A.; Ma, C.; Wang, X.; Qiu, L. Research progress and perspective on drought stress in legumes: A review. Int. J. Mol. Sci. 2019, 20, 2541. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Subudhi, P.K.; Crasta, O.R.; Rosenow, D.T.; Mullet, J.E.; Nguyen, H.T. Molecular mapping of QTLs conferring stay-green in grain sorghum (Sorghum bicolor L. Moench). Genome 2000, 43, 461–469. [Google Scholar] [CrossRef]
- Bastos, E.A.; do Nascimento, S.P.; da Silva, E.M.; Filho, F.R.F.; Gomide, R.L. Identification of cowpea genotypes for drought tolerance. Rev. Cienc. Agron. 2011, 42, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, J.T.; Pedrosa, M.B.; Sobrinho, J.T. Efeito da variação de níveis de água disponível no solo sobre o crescimento e produção de feijão caupi, vagens e grãos verdes. Hortic. Bras. 2004, 22, 174–177. [Google Scholar] [CrossRef] [Green Version]
- Leite, M.D.L.; Rodrigues, J.D.; Virgens Filho, J.S. Das Efeitos do déficit hídrico sobre a sultura do caupi, cv-EMAPA-821. III-Produção. Rev. Agric. 2000, 75, 9–20. [Google Scholar] [CrossRef]
- Prince, S.J.; Murphy, M.; Mutava, R.N.; Zhang, Z.; Nguyen, N.; Kim, Y.H.; Pathan, S.M.; Shannon, G.J.; Valliyodan, B.; Nguyen, H.T. Evaluation of high yielding soybean germplasm under water limitation. J. Integr. Plant Biol. 2016, 58, 475–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.-B.; Guo, C.; Li, F.; Li, M.; He, J. High Soybean Yield and Drought Adaptation Being Associated with Canopy Architecture, Water Uptake, and Root Traits. Agronomy 2020, 10, 608. [Google Scholar] [CrossRef]
- Kashiwagi, J.; Krishnamurthy, L.; Upadhyaya, H.D.; Krishna, H.; Chandra, S.; Vadez, V.; Serraj, R. Genetic variability of drought-avoidance root traits in the mini-core germplasm collection of chickpea (Cicer arietinum L.). Euphytica 2005, 146, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Polania, J.; Rao, I.M.; Cajiao, C.; Grajales, M.; Rivera, M.; Velasquez, F.; Raatz, B.; Beebe, S.E. Shoot and Root Traits Contribute to Drought Resistance in Recombinant Inbred Lines of MD 23–24 × SEA 5 of Common Bean. Front. Plant Sci. 2017, 8, 296. [Google Scholar] [CrossRef]
- Sponchiado, B.N.; White, J.W.; Castillo, J.A.; Jones, P.G. Root Growth of Four Common Bean Cultivars in Relation to Drought Tolerance in Environments with Contrasting Soil Types. Exp. Agric. 1989, 25, 249–257. [Google Scholar] [CrossRef]
- Belachew, K.Y.; Nagel, K.A.; Fiorani, F.; Stoddard, F.L. Diversity in root growth responses to moisture deficit in young faba bean (Vicia faba L.) plants. PeerJ 2018, 6, e4401. [Google Scholar] [CrossRef] [Green Version]
- Kumar, J.; Basu, P.S.; Srivastava, E.; Chaturvedi, S.K.; Nadarajan, N.; Kumar, S. Phenotyping of traits imparting drought tolerance in lentil. Crop. Pasture Sci. 2012, 63, 547–554. [Google Scholar] [CrossRef]
- Kochian, L.V. Root architecture. J. Integr. Plant Biol. 2016, 58, 190–192. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Cui, K.; Xu, A.; Nie, L.; Huang, J.; Peng, S. Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity in rice seedlings. Acta Physiol. Plant. 2015, 37, 9. [Google Scholar] [CrossRef]
- Blum, A. Crop responses to drought and the interpretation of adaptation. Plant Growth Regul. 1996, 20, 135–148. [Google Scholar] [CrossRef]
- Maseda, P.H.; Fernández, R.J. Stay wet or else: Three ways in which plants can adjust hydraulically to their environment. J. Exp. Bot. 2006, 57, 3963–3977. [Google Scholar] [CrossRef]
- Nkomo, G.V.; Sedibe, M.M.; Mofokeng, M.A. Phenotyping cowpea accessions at the seedling stage for drought tolerance using the pot method. bioRxiv 2020. [Google Scholar] [CrossRef]
- Sánchez-Reinoso, A.D.; Ligarreto-Moreno, G.A.; Restrepo-Díaz, H. Evaluation of drought indices to identify tolerant genotypes in common bean bush (Phaseolus vulgaris L.). J. Integr. Agric. 2020, 19, 99–107. [Google Scholar] [CrossRef]
- Shah, T.M.; Imran, M.; Atta, B.M.; Ashraf, M.Y.; Hameed, A.; Waqar, I.; Shafiq, M.; Hussain, K.; Naveed, M.; Aslam, M.; et al. Selection and screening of drought tolerant high yielding chickpea genotypes based on physio-biochemical indices and multi-environmental yield trials. BMC Plant Biol. 2020, 20, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Arifuzzaman, M.; Barman, S.; Hayder, S.; Azad, M.A.K.; Turin, M.T.S.; Amzad, M.A.; Masuda, M.S. Screening of bread wheat (Triticum aestivum L.) genotypes under drought stress conditions using multivariate analysis. Cereal Res. Commun. 2020, 48, 301–308. [Google Scholar] [CrossRef]
- Alidu, M.S. Evaluation of Cowpea Genotypes for Drought Tolerance Using the Pot Screening Approach. Asian Res. J. Agric. 2018, 10, 1–11. [Google Scholar] [CrossRef]
- Abdou, R.I.B.Y.; Mensah, B.; Addam, K.S.; Akromah, R. Using morpho-physiological parameters to evaluate cowpea varieties for drought tolerance. Int. J. Agric. Sci. Res. 2013, 2, 153–162. [Google Scholar]
- Ayala, C.C.; Villadiego, C.C.; Pacheco, C.P.; Tatis, H.A.; Camacho, M.M.E. Growth, biomass distribution, gas exchange and chlorophyll fluorescence in cowpea (Vigna unguiculata (L.) Walp.) under drought conditions. Aust. J. Crop. Sci. 2020, 14, 371–381. [Google Scholar] [CrossRef]
- Ravelombola, W.; Shi, A.; Qin, J.; Weng, Y.; Bhattarai, G.; Zia, B.; Zhou, W.; Mou, B. Investigation on Various Aboveground Traits to Identify Drought Tolerance in Cowpea Seedlings. HortScience 2018, 53, 1757–1765. [Google Scholar] [CrossRef] [Green Version]
- Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Rivas-Ubach, A.; Oravec, M.; Vecerova, K.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C.; et al. Opposite metabolic responses of shoots and roots to drought. Sci. Rep. 2014, 4, 6829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamidou, F.; Zombré, G.; Braconnier, S. Physiological and Biochemical Responses of Cowpea Genotypes to Water Stress Under Glasshouse and Field Conditions. J. Agron. Crop. Sci. 2007, 193, 229–237. [Google Scholar] [CrossRef]
- Ogbonnaya, C.I.; Sarr, B.; Brou, C.; Diouf, O.; Diop, N.N.; Roy-Macauley, H. Selection of Cowpea Genotypes in Hydroponics, Pots, and Field for Drought Tolerance. Crop. Sci. 2003, 43, 1114–1120. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Bank Code | Origin | Donor Institution # | Type |
|---|---|---|---|---|
| C3 | Cp 4924 | Évora, Portugal | INIAV | Landrace |
| C5 | Cp 5648 | Abrantes, Portugal | INIAV | Landrace |
| C8 | Vg 50 | Pinhel, Portugal | UTAD | Landrace |
| C11 | Vg 56 | Macedo de Cavaleiros, Portugal | UTAD | Landrace |
| C15 | Fradel | Portugal | INIAV | Variety |
| C56 | Vg 59 | Fundão, Portugal | UTAD | Landrace |
| C57 | Vg 62 | Covilhã, Portugal | UTAD | Landrace |
| C58 | Vg 72 | Mogadouro, Portugal | UTAD | Landrace |
| C18 | BGE038478 | Málaga, Spain | CRF-INIA | Landrace |
| C59 | IT97K-499-35 | Nigeria | UCR | Breeding line |
| C41 | VIG 58 | Angola | IPK | Landrace |
| C43 | VIG 90 | Egypt | IPK | Landrace |
| C45 | VIG 49 | Senegal | IPK | Landrace |
| C47 | VIG 1649 | Iran | IPK | Landrace |
| C49 | VIG 206 | Cuba | IPK | Landrace |
| C51 | VIG 10 | China | IPK | Landrace |
| C52 | NI 778 | India | BGM | Landrace |
| C53 | Baio Cooafom | Brazil | EMBRAPA | Variety |
| C19 | BGE002195 | Orense, Spain | CRF-INIA | Landrace |
| C20 | BGE019751 | Girona, Spain | CRF-INIA | Landrace |
| C21 | BGE024703 | Baleares, Spain | CRF-INIA | Landrace |
| C22 | BGE025213 | Caceres, Spain | CRF-INIA | Landrace |
| C23 | BGE047731 | Pontevedra, Spain | CRF-INIA | Landrace |
| C24 | BGE036461 | Huelva, Spain | CRF-INIA | Landrace |
| C29 | BGE039237 | Cordoba, Spain | CRF-INIA | Landrace |
| C31 | MG 113779 | Puglia, Italy | CRF-INIA | Landrace |
| C39 | AUA 1 | Crete, Greece | AUA | Landrace |
| References | ||||
| IT93K-503-1 | IT93K-503-1 | Nigeria | UCR | Breeding line |
| Bambey21 | Bambey21 | Senegal | UCR | Cultivar |
| Parameter # | Source | DF * | Residual DF | F | Prob < F |
|---|---|---|---|---|---|
| SDW | Treatment | 1 | 116 | 293.1 | <0.001 |
| Genotype | 28 | 116 | 3.1 | <0.001 | |
| Treatment: Genotype | 28 | 116 | 2.7 | <0.001 | |
| PH | Treatment | 1 | 116 | 13.493 | <0.001 |
| Genotype | 28 | 116 | 6.6 | <0.001 | |
| Treatment: Genotype | 28 | 116 | 1.2 | 0.234 | |
| PDW | Treatment | 1 | 112 | 253.5 | <0.001 |
| Genotype | 27 | 112 | 4.6 | <0.001 | |
| Treatment: Genotype | 27 | 112 | 2.6 | <0.001 | |
| SG | Treatment | 1 | 116 | 130.5 | <0.001 |
| Genotype | 28 | 116 | 6.9 | <0.001 | |
| Treatment: Genotype | 28 | 116 | 2.3 | 0.001 | |
| NPP | Treatment | 1 | 112 | 151.5 | <0.001 |
| Genotype | 27 | 112 | 5.3 | <0.001 | |
| Treatment: Genotype | 27 | 112 | 2.1 | 0.004 | |
| NGP | Treatment | 1 | 112 | 186.8 | <0.001 |
| Genotype | 27 | 112 | 5.2 | <0.001 | |
| Treatment: Genotype | 27 | 112 | 4.4 | <0.001 | |
| RDW | Treatment | 1 | 116 | 15.5 | <0.001 |
| Genotype | 28 | 116 | 3.4 | <0.001 | |
| Treatment: Genotype | 28 | 116 | 1.1 | 0.352 | |
| RSR | Treatment | 1 | 116 | 294.6 | <0.001 |
| Genotype | 28 | 116 | 3.3 | <0.001 | |
| Treatment: Genotype | 28 | 116 | 2.7 | <0.001 | |
| DFF | Treatment | 1 | 116 | 0.0 | 0.906 |
| Genotype | 28 | 116 | 5.1 | <0.001 | |
| Treatment: Genotype | 28 | 116 | 1.3 | 0.178 |
| Parameters # | PC 1 | PC 2 | PC3 |
|---|---|---|---|
| SDW | 0.29 | −0.26 | 0.51 |
| PH | −0.05 | −0.01 | 0.67 |
| SG | −0.29 | −0.23 | 0.37 |
| NPP | 0.49 | 0.05 | 0.06 |
| PDW | 0.54 | 0.10 | −0.01 |
| RDW | 0.08 | 0.62 | 0.38 |
| NGP | 0.51 | 0.05 | −0.12 |
| RSR | −0.20 | 0.69 | 0.01 |
| Eigenvalue | 3.36 | 1.80 | 1.61 |
| % variance | 40.57 | 21.73 | 19.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, R.; Carvalho, M.; Rosa, E.; Carnide, V.; Castro, I. Root and Agro-Morphological Traits Performance in Cowpea under Drought Stress. Agronomy 2020, 10, 1604. https://doi.org/10.3390/agronomy10101604
Santos R, Carvalho M, Rosa E, Carnide V, Castro I. Root and Agro-Morphological Traits Performance in Cowpea under Drought Stress. Agronomy. 2020; 10(10):1604. https://doi.org/10.3390/agronomy10101604
Chicago/Turabian StyleSantos, Ricardo, Márcia Carvalho, Eduardo Rosa, Valdemar Carnide, and Isaura Castro. 2020. "Root and Agro-Morphological Traits Performance in Cowpea under Drought Stress" Agronomy 10, no. 10: 1604. https://doi.org/10.3390/agronomy10101604
APA StyleSantos, R., Carvalho, M., Rosa, E., Carnide, V., & Castro, I. (2020). Root and Agro-Morphological Traits Performance in Cowpea under Drought Stress. Agronomy, 10(10), 1604. https://doi.org/10.3390/agronomy10101604