Abstract
Basin wildrye [Leymus cinereus (Scribn. and Merr.) Á. Löve] and creeping wildrye [Leymus triticoides (Buckley) Pilg.] are native perennial grasses cultivated for seed used for fire rehabilitation and revegetation in western North America. Although L. cinereus produces large spike inflorescences with many seeds, it is prone to seed shattering. Seed can be harvested before shattering, but often displays poor germination and seedling vigor. Conversely, L. triticoides has fewer seeds per spike, but relatively strong seed retention. Both species are allotetraploid (2n = 4x = 28) and form fertile hybrids used for breeding and genetic research. A dominant, major-effect seed-shattering gene (SH6) from L. cinereus was previously identified in an L. triticoides backcross population. In this study, a DNA marker was used to select the recessive L. triticoides seed-retention allele (sh6) in cycle six (C6) of a L. cinereus × L. triticoides breeding population and evaluate gene × harvest date effects on seed yield and germination characteristics in a full-sib family derived from homozygous (sh6/sh6) and heterozygous (SH6/sh6) C6 parents. Although seed yields of shattering genotypes were 19.4% greater than non-shattering genotypes on the first harvest dates, yields of non-shattering genotypes were 167% greater on the last harvest dates. Seed harvested on the last harvest date reached 50% germination 4.2 days (26.4%) earlier and displayed 20.5% higher upper percentage germination limits than seed harvested on the first harvest date. Results indicate that the sh6 seed-retention gene will improve basin wildrye seed retention and indirectly improve seed germination by enabling later harvest dates.
1. Introduction
Native perennial grass seed has been used for revegetation treatments across millions of hectares of arid and semiarid rangelands of the Great Basin and other regions of western North America [1], but many plantings fail to establish and provide desired outcomes [2,3]. Variations in seed germination and seedling emergence characteristics have been identified as a primary limitation to the success of grassland restoration on these rangelands [4,5,6]. Perennial grasses with early seedling emergence are more likely to survive the first growing season on semiarid rangelands infested with the invasive annual grass Bromus tectorum L. [6]. The success of rangeland seedings may be enhanced by planting techniques [7], seed-enhancement technologies [8], and the choice of plant materials [6,9,10]. Plant breeding has potential as a cost-effective approach to improve ecosystem services of perennial plants in managed landscapes [11,12,13].
Basin wildrye [Leymus cinereus (Scribn. and Merr.) Á. Löve] (2n = 4x = 28) is one of the largest native grasses in the Great Basin region and is widely distributed throughout much of western North America. This grass provides forage and habitat for both livestock and wildlife [14,15,16,17], is useful for fire rehabilitation, rangeland revegetation, mine land reclamation, and other conservation uses [17,18], and has potential as a biofuel feedstock [19,20]. Basin wildrye is often seen as localized stands in drainage areas or as dispersed individuals that stand out as tall caespitose plants, reaching heights in excess of 2 m in some environments [21]. Spikes of L. cinereus are also quite large, producing up to 2000 florets each [22], with seed yield potential up to 350 kg/ha [17]. However, spikes are prone to shattering and require close scrutiny during maturation [17,22]. In addition, seed from native stands often display poor germination characteristics [10,23]. Thus, seed germination, seedling vigor, stand establishment, and yield of viable seeds have been focal points of L. cinereus research [24,25] and cultivar development [26,27,28].
Seed shattering can result in major economic losses to grass seed producers if harvest is delayed, but germination can be compromised if seed is harvested too early in development [29]. Evaluations of four cool-season forage grasses including crested wheatgrass [Agropyron desertorum (Fisch. Ex Link) Schult.], Russian wildrye [Psaythrostachys juncea (Fisch.) Nevski’, intermediate wheatgrass [Thinopyrum intermedium (Host) Barkw. and D.R. Dewey], and western wheatgrass [Pascopyrum smithii (Rydb.) Á. Löve] demonstrated that seed mass and seed germination would not be seriously compromised and that problems associated with seed shattering could be managed if these grasses were windrowed approximately two or three days before attaining maximal seed mass [29]. However, the extent of seed shattering varies greatly among different grass species, and the timing and rate of seed development varied among years [29]. Thus, the optimum date of seed harvest can be difficult to predict, which is certainly true for L. cinereus [17,22].
Creeping wildrye or beardless wildrye [Leymus triticoides (Buckley) Pilg.] (2n = 4x = 28) is morphologically distinct from L. cinereus in many ways, but these two species have similar distributions and readily form fertile hybrids when grown together [18,19,21]. The growth habit of L. triticoides is relatively short, strongly rhizomatous, and much less conspicuous compared to L. cinereus [21]. The spikes of L. triticoides are also much smaller, producing only approximately 10% as many seeds as L. cinereus [22], yet overall seed yields of approximately 330 kg/ha [30] are comparable to L. cinereus. Another distinctive feature of L. triticoides is that it has high levels of seed dormancy, which can lessen its ability to compete with weeds in rangeland plantings [22,31]. Although viability and seedling vigor characteristics of seed harvested from native or cultivated stands of L. cinereus can also be problematic [23], it does not display strong levels of seed dormancy like L. triticoides [22].
Experimental families derived from two L. cinereus and L. triticoides hybrids have been used to identify quantitative trait loci (QTLs) controlling functionally important traits including growth habit, flowering, biomass production, heavy-metal accumulation, forage quality, and seed production [22,32,33,34]. Among many findings, a major-effect seed-shattering QTL, explaining 43% of the phenotypic variation, was identified on linkage group 6a (LG6a) in two L. triticoides backcross populations [22]. Two other minor seed-shattering QTLs were identified on LG3b and LG4Xm, but the latter may have been caused by pleiotropic effects of a QTL for number of florets per spike on the same region of LG4Xm. Sixteen expressed sequence tag (EST) markers on Leymus LG6a, including some of the best seed-shattering QTL markers, showed colinear alignments to rice (Oryza sativa L.) Chromosome 2 [22,35]. The Leymus LG6a seed-shattering QTL showed possible alignment to the seed-shattering QTL of American wild rice (Zizania palustris L.) [22,36], but no other alignments to seed-shattering QTLs or genes were identified on Chromosome 2 of domesticated rice [22]. It was deduced from these studies that the L. cinereus seed-shattering gene, on LG6a, was dominant (designated herein as SH6) and that the non-shattering L. triticoides allele was recessive (designated herein as sh6) based on comparisons of the parents, hybrids, and backcross progeny [22]. It was also observed that the L. triticoides sh6 seed-retention gene decreased the time required for imbibed seeds to germinate, which was surprising because L. triticoides displays strong seed dormancy [22]. It was postulated that the L. cinereus SH6 gene slowed germination because those seeds were harvested at earlier dates, as seeds began to naturally disarticulate, whereas the L. triticoides sh6 gene allowed seeds to after-ripen in the field [22]. However, seed from each plant was bulked over each harvest so it was impossible to distinguish effects of the SH6 gene from those of harvest date on germination. Moreover, the actual rate of natural seed disarticulation over different harvest dates was not measured [22], so the actual effect of the SH6 gene on seed yield, over time, was not documented. Discovery of the L. cinereus SH6 seed-shattering gene in a L. triticoides backcross population was fortuitous in that it was easy to identify and map the dominant shattering allele, but there was no practical reason to introgress the L. cinereus seed-shattering gene into L. triticoides. However, it was hypothesized that introgression of the recessive L. triticoides seed-retention gene into L. cinereus would decrease its shattering and possibly improve its germination rate by extended field ripening [22]. This could provide significant practical and economic benefits to seed producers and seed users, including public and private land managers.
The objective of this project was to introgress the recessive L. triticoides LG6a seed-retention gene [22] into L. cinereus and rigorously evaluate the effect of this gene on the seed yield per spike and germination rates over an extended series of harvest dates beginning approximately 22 to 24 days after pollination (DAP), shortly before the first signs of normal seed shattering in L. cinereus genotypes, and ending approximately 91 to 99 DAP over two different years.
2. Materials and Methods
2.1. Plant Materials and SH6-Linked DNA Marker
The L. cinereus shattering (SH6) and L. triticoides non-shattering (sh6) QTL alleles were associated with 168 and 158 bp amplicons, respectively, produced by the LG6a Ltc096 PCR marker [22], which was developed using an expressed sequence tag (EST) library from a L. triticoides × L. cinereus (Ltc) hybrid [35]. Sequences, map locations, and gene ontology of all EST markers used to identify and map the LG6a SH6 seed-shattering QTL [22], including the Ltc096 forward (GCGCACTACCGCCTCTTAGTT) and reverse (GTCCAGGTAGCACACCTCCG) primers, were previously described [35] in addition to two closely linked phenylalanine ammonialyase (PAL) lignin biosynthesis marker loci near the Ltc096 locus [37]. The Ltc096 marker had the highest LOD score of 1122 markers used for QTL analysis of the L. triticoides backcross populations [22]. Although several other Leymus EST markers were linked to the SH6 gene, the Ltc096 has proven to be the most useful marker in terms of PCR amplification, codominance, genome specificity, ease of scoring, and linkage to the SH6 gene. Hereafter, the 168 bp L. cinereus Ltc096 marker allele is designated “c” and the 158 bp L. triticoides Ltc096 marker allele is designated “t”, with three genotypic classes—homozygous c/c, heterozygous c/t, or homozygous t/t.
This study utilized a full-sib family of 187 F1 progeny from the cross of two parental genotypes, L57S-491 × L57S-577. The L57S-491 and L57S-577 parents arose from six cycles of selection beginning with seed obtained from a population of approximately 100 L. cinereus × L. triticoides F1 hybrids and 100 L. triticoides × L. cinereus F1 hybrids, which included one hybrid plant used to identify the SH6 locus [22]. The original L. triticoides × L. cinereus crosses were made in 1990 at the Utah Agricultural Experiment Station (UAES) Greenville Research Farm (North Logan, UT, USA), between multiple plants of M10 L. cinereus (herein Acc:636) received from Sylvester Smoliak (Agriculture Canada, Lethbridge, AB, Canada) and A30 L. triticoides (herein Acc:641) originally collected by Kay Asay near Jamieson, OR. In the sixth cycle (C6) of selection, the L57S population (including the L57S-491 and L57S-577 parents) was developed by screening 715 seedlings from cycle 5, designated L57X, for the presence of the L. triticoides Ltc096.158 (t) seed-retention marker alleles in the winter of 2007–2008, as described above and in the below. The L57S population was planted in the spring of 2008 and controlled pollinations were made among seven L57S individuals including L57S-491 (t/t) × L57S-577 (c/t) in 2010. A set of 468 full-sib F1 progeny from the cross of L57S-491 (t/t) × L57S-577 (c/t) was transplanted into a field at the Utah State University Millville Farm (approximately 41°39′23.9″ N and 111°48′50.7″ W) in 2011 in rows spaced 1 m apart with 0.75 m within each row. A subset of 187 F1 progeny were selected for evaluation based in part on the number of useable spikes per plant in 2013.
2.2. Genotyping the Ltc096 Marker
Whatman® FTA® Cards provided a fast and cost-effective approach to capture DNA and screen one PCR marker, Ltc096, for the L-57X population (715 samples). However, for genetic data analysis of the L57S-491 × L57S-577 family (including parents and progeny), total cellular DNA samples were extracted using DNeasy 96 kits (Qiagen, Valencia, CA, USA) according to the supplier’s instructions, which provided higher-quality DNA and better PCR amplifications compared to the FTA® Cards. For the DNeasy extractions, approximately 10 mg of lyophilized tissue from each plant was milled using a MM 300 mixer mill (Qiagen, Valencia, CA, USA) in collection boxes containing 96 1.2 mL microtiter tubes with caps. The Ltc096 marker was amplified using the polymerase chain reaction (PCR) in 10 µL volumes using 1X PCR buffer, 0.2 mM each of four deoxynucleotide triphosphate bases, 0.5 µM of each primer, 1.25 units jump start Taq DNA polymerase, 0.1 µmol Rhodamine-X-5-propargylamino-2′-deoxycytidine-5′-triphosphate, and 20–50 ng of DNA for each plant. The PCR amplifications were performed using the following profile: (1) 95 °C for 90 s (2) 35 cycles of 95 °C for 30 s, 60 °C for 30 s, 72 °C for 1 min and (3) a final extension of 72 °C for 6 min. Amplicons from each PCR sample were fractionated using Applied Biosystems (Foster City, CA, USA) ABI 3730 capillary electrophoresis instruments with internal GeneScan-500 LIZ size standards in each sample to estimate the number of DNA base pairs (bp) for each amplicon based on relative mobilities. The resulting trace files from each sample were compared and scored along with trace files from the L. triticoides and L. triticoides × L. cinereus F1-hybrid parental genotypes originally used to identify the SH6 gene [22], using Genographer 1.6.0 [38] graphical software with bin widths of approximately 1.0 bp and a minimum threshold of 600 fluorescence units to ensure homology of the inferred marker alleles. Using this procedure, the average estimated lengths of the Ltc096.158 (t) and Ltc096.168 (c) alleles were 157.91 and 168.61, with standard deviations of 0.14 and 0.16, respectively, in this study.
2.3. Measuring Effects of SH6 Seed-Shattering QTL Marker on Seed Yield and Seed Germination Traits
Although it would be difficult to directly measure the natural shattering or loss of seed in the field, seven or eight repeated harvests of two spikes from each plant over the course of time (24 to 91 DAP in 2015 and 22 to 99 DAP in 2017) were used to indirectly test effects of the SH6 seed-shattering QTL marker on seed yield traits and also test possible effects harvest date on seed germination. Spikes were harvested from each of the 187 L57S-491 × L57S-577 plants approximately 24, 31, 39, 49, 57, 67, 81, and 91 DAP in 2015 and approximately 22, 29, 43, 57, 71, 88, and 99 DAP in 2017. In some cases, it appeared that there would not be enough spikes to complete all seven or eight harvests, so priority was generally given to the first and last harvests. Dates of anthesis for basin wildrye typically vary as much as five to seven days, among different plants and spikes, but as a point of reference the dates of pollination were estimated to be on June 22 in both 2015 and 2017 based on recorded dates of controlled pollinations made successfully each year. Thus, corresponding dates of harvest ranged from July 16 (24 DAP) to September 21 (91 DAP) in 2015 and from July 14 (22 DAP) to September 29 (99 DAP) in 2017. Plants were not harvested in 2016 due to insufficient seed production presumably caused by drought or other unfavorable environmental conditions. Spikes from each harvest were stored in a dry greenhouse chamber until all harvests were completed. Spike length (cm) was measured from the basal spikelet branching point to the tip of the top of the uppermost spikelet for each of two spikes collected from each plant on each harvest date. The average seed yield per spike (mg seed per spike) from each harvest–plant combination was measured by manually threshing, cleaning, and weighing seeds from each pair of spikes together as one sample. Cleaned seeds were bulked into 30 seed lots corresponding to each of two marker genotypes (t/t or c/t) from each of eight harvest dates in 2015 or seven harvest dates in 2017. The resulting seed lots were then stored in a long-term seed storage facility maintained at 2 °C and 5% humidity. All seed yield data are provided (Supplementary Data 1).
Three sets of germination tests (GT1, GT2, and GT3) were conducted on five replicated samples of 100 seeds from each of 16 seed lots from 2015 and 14 seed lots from 2017. The first set of germination tests were initiated (imbibed) 226 DAP for seed harvested in 2015 (15_GT1) or 225 DAP for seed harvested in 2017 (17_GT1) with 14 day cold-stratification treatments at 4 °C (dark). The second set of germination tests were imbibed 266 DAP for seed harvested in 2015 (15_GT2) or 265 DAP for seed harvested in 2017 (17_GT2) without any cold-stratification treatment. The third set of germination tests was conducted after extended seed storage beginning 1025 DAP for seed harvested in 2015 (15_GT3) or 846 DAP for seed harvested in 2017 (17_GT3) without any cold-stratification treatment. For each of the three germination tests, five replicate samples of 100 seeds from each of the 16 seed lots from 2015 (total of 8000 seeds per test or 9000 seeds total) or 14 seed lots from 2017 (total of 7000 seeds per test or 21,000 total) were weighed and dusted with the fungicide, tetramethylthiuram disulfide (Sigma-Aldrich, St. Louis, MO, USA). Seed germination samples were evenly distributed in Cont 156C acrylic germination boxes (Hoffman Manufacturing Inc., Jefferson, OR, USA), approximately 11 × 11 × 3 cm, on blue germination blotter paper (Anchor Paper, St. Paul, MN, USA) that was previously moistened with distilled water to the point of saturation and then drained to remove excess water. The germination boxes were covered with tight-fitting lids, provided by the manufacturer, and enclosed in plastic bags with eight boxes per bag to maintain moisture. Cold-stratification treatments were applied after 2 days of imbibition at room temperature followed by a 2 week incubation at 8 °C in darkness. Otherwise, seeds were incubated at 25/15 °C in an 8/16 h daily schedule in darkness until each of the six germination tests was completed. Germination was assessed at intervals of one, two, or sometimes three days by counting and removing seeds that exhibited both root and shoot development, up to 36 days or after initial imbibition with a 14 day cold stratification or up to 28 days after initial imbibition without cold stratification until seed germinations tapered off to 3 or fewer (often zero) after a two- or three-day interval. These germination counts were numbered and identified based on the number of days after imbibition (DAI) including possible days of cold-stratification treatment. There was a total of 10 counts for 15_GT1, 11 counts for 15_GT2, and 9 counts for 15_GT3 with five replicate samples (100 seeds per sample) from 16 seed lots in 2015, making a total of 2400 counts (Supplementary Data 2). There was a total of 10 counts for 17_GT1, 9 counts for 17_GT2, and 10 counts for 17_GT3 with five replicate samples (100 seeds per sample) from 14 seed lots in 2017, making a total of 2030 counts (Supplementary Data 2). Water was added to germination boxes and drained, as needed, to maintain moisture of germination paper. All germination data are provided (Supplementary Data 2).
The average number of seeds per cm of spike was calculated using the average seed yield per spike, spike length, and average seed mass (mg per seed) based on measurements of seeds from each germination test.
2.4. Statistical Analyses
All seed yield (mg per spike and number of seeds per cm spike), spike length, and seed mass data were analyzed using the MIXED procedure of SAS (Cary, NC, USA) to test main effects and determine least significant differences (p < 0.05) among adjusted means for year, genotype, harvest (year), genotype × year, and genotype × harvest (year) effects, where genotype is c/t or t/t as determined by the Ltc096 marker, year is 2015 or 2017, and harvest(year) is one of eight possible harvest dates nested within 2015 or seven possible harvest dates nested within 2017, measured in days after pollination (DAP). Seed yield traits (mg per spike and number of seeds per cm of spike) were analyzed using individuals (ID) nested within genotype as random subjects (n = 187) with year and harvest (year) as repeated measures (up to 15 possible measurements per plant). Similarly, spike length was analyzed using individuals (ID) nested within genotype as random subjects with spike sample (n = 2) and harvest (year) as random and fixed repeated measures for each plant, respectively. Seed mass was analyzed using genotype as the subject with germination sample and harvest(year) as random and fixed repeated measures for each plant, respectively, where the germination sample was one of 15 possible 100-seed samples used for each germination test. Seed yield and spike length traits were analyzed using the UN@AR(1) first-order autoregressive (1) covariance structure for two repeated variables. Seed mass was analyzed using the CS compound symmetry structure for one variable with constant variance and covariance among levels. All SAS code and results of the seed yield data analysis are provided (Supplementary Data 1).
Germination data were also analyzed, by year, using the MIXED procedure of SAS (Cary, NC, USA) to test main effects and determine least significant differences (p < 0.05) among adjusted means for germination test, harvest, genotype, germination count (germination test) and the interaction of harvest × germination count (germination test), where germination test was one of six germination tests, as described above, and germination count was one of least nine germination counts, numbered by days after imbibition (DAI). Germination sample was included as a random variable and germination count (germination test) was treated as a repeated measure on each germination sample. The germination data were analyzed by year due to the complexity of having different harvest dates between years, different germination tests between years, and different germination counts between germination tests. All SAS code and results of the seed germination data analysis are provided (Supplementary Data 2). Moreover, for each of the six germination tests, the percentage germination response curves over DAI were fit for each of eight harvest dates in 2015 and for each of seven harvest dates in 2017 using SIGMAPLOT (Systat Software Inc., Version 13, San Jose, CA, USA), based on the mean number of germinated seeds in five 100-seed samples from 16 seed lots from 2015 (two possible genotypes × eight harvest dates) or 14 seed lots from 2017 (two possible genotypes × seven harvest dates). The percentage germination response () to number of days after imbibition () was fit to standard dose–response curves using a non-linear four-parameter logistic (4PL) model (Equation (1)) as shown:
where indicates the upper percentage germination limit, indicates the lower percentage germination limit, represents the number of days required to reach 50% of the upper percentage germination limit (i.e., MAX50), and is the slope of the line around the MAX50.
3. Results
3.1. Marker-Assisted Selection of L57S-491 and L57S-577 Parents and Segregation of Ltc096 Marker Alleles
A total of 715 L57X plants were screened using the Ltc096 marker in order to identify and select parental plants that may carry the L. triticoides seed-retention QTL allele. A total of 191 plants were heterozygous for the 168 bp L. cinereus (c) and 158 bp L. triticoides (t) Ltc096 marker alleles, with a designated genotype of c/t, and 10 plants were homozygous (t/t). The observed frequency the Ltc096 t allele was 0.148, which was significantly less (p < 0.001) than the expected frequency of alleles in an F1-derived population (0.5) or L. cinereus backross population (0.25) if the L. triticoides and L. cinereus parental accessions were fixed for the t and c marker alleles, respectively. In a small survey of five plants from each accession, of the L. triticoides Acc:641 and parental accessions, the 168 bp Ltc096 c marker allele was fixed in L. cinereus Acc:638 parental accession, but the 158 bp Ltc096 t allele was not completely fixed in L. triticoides Acc:641 parental accession. The observed genotypic frequency of 10 t/t homozygotes, 191 c/t heterozygotes, and 514 c/c homozygotes was not significantly different from the expected genotypic frequencies from random mating (approximately 16, 184, and 532, respectively) given the low allele frequency (0.148) of the Ltc096 t marker allele in the L57X population. However only two of the ten Ltc096 t/t homozygotes, including the L57S-491 parent, and three Ltc096 c/t heterozygotes displayed clear evidence of seed retention in the field. All ten t/t homozygotes and none of the c/t heterozygotes were expected to display seed-retention phenotype if there was perfect linkage between the Ltc096 marker and the seed-shattering gene. Nevertheless, progeny of a cross of the non-shattering L57S-491 (t/t) plant and the shattering L57S-577 (c/t) plant produced a full-sib family of plants that was visibly segregating for the shattering and non-shattering phenotypes. A total of 187 F1 progeny from a controlled cross the L57S-491 (t/t) × L57S-577 (c/t) were genotyped using the Ltc096 marker to in order to validate and examine its putative association with the SH6 gene. The frequency of 106 (56.7%) c/t homozygotes and 81 (43.4%) t/t heterozygotes was not significantly different (p > 0.05) from the 1:1 expected segregation ratio from the L57S-491 (t/t) × L57S-577 (c/t) cross.
3.2. Average Effects of the Ltc096 Marker on Spike Yield Traits in the L57S-491 × L57S-577 F1 Population
A total of 1426 seed yield measurements were obtained from the 1496 attempted harvests in 2015 (187 plants × 8 harvests) and a total of 520 seed yield measurements were obtained from 1309 attempted harvests in 2017 (187 plants × 7 harvests). At least four successful harvests were taken from 182 plants in 2015 and at least one successful harvest was taken from 107 plants in 2017. A total of 1946 seed yield measurements were obtained over both years with at least 4 harvests on 184 of the 187 plants including 104 plants with the Ltc096 t/t genotype and 80 plants with the c/t genotype. Significant (p < 0.0001) effects of year, genotype, harvest date (year), and genotype × harvest date (year) terms were detected for seed yield per spike, seed mass, and the number of seeds per spike with no significant (p > 0.05) effect for the genotype × year terms. Significant (p < 0.0001) effects of year, genotype × year, and harvest date (year) terms were detected for spike length with no significant (p > 0.05) effects for the genotype × year or genotype × harvest date (year) terms.
Seed yield per spike showed significant declines between the first and last harvests, but these yield declines were significantly more pronounced in the shattering Ltc096 c/t genotypes versus the non-shattering Ltc096 t/t genotypes (Figure 1a). Over the course of each season, the seed yield per spike of the shattering Ltc096 c/t genotypes showed significant declines from 606 to 147 mg (75.7%) in 2015 (Figure 1a; Table S1) and from 838 to 206 mg (75.4%) in 2017 (Figure 1a; Table S2), whereas the non-shattering Ltc096 t/t genotypes showed smaller declines from 527 to 454 mg (13.9%) in 2015 (Figure 1a; Table S1) and from 677 to 464 mg (31.5%) in 2017 (Figure 1a, Table S2). Compared to non-shattering Ltc096 t/t genotypes, the shattering Ltc096 c/t genotypes had 15.0% to 23.8% (average 19.4%) greater seed yield per spike on the first harvest dates of 2015 and 2017, respectively (Figure 1a; Tables S1 and S2). Conversely, the non-shattering Ltc096 t/t genotypes had 209% and 125% (average 167%) greater seed yield per spike on the last harvest dates of 2015 and 2017, respectively, compared to the shattering Ltc096 c/t genotypes (Figure 1a; Tables S1 and S2). There was a significant cross-over interaction of genotype and harvest date for seed yield per spike.
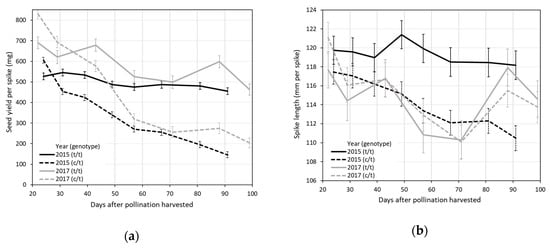
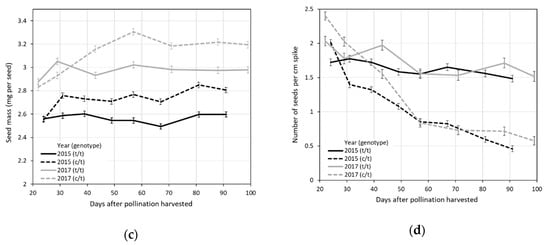
Figure 1.
Comparisons of trait averages between heterozygous (c/t) and homozygous (t/t) genotypes of the L. cinereus (c) and L. triticoides (t) Ltc096 marker alleles over eight different harvest dates in 2015 and seven different harvest dates in 2017, numbered days after pollination, for (a) seed yield per spike; (b) spike length; (c) seed mass; and (d) seed density.
The average spike length of the shattering Ltc096 c/t genotypes showed significant declines in later harvest dates of 2015 and 2017 (Figure 1b; Tables S1 and S2). The non-shattering Ltc096 t/t genotypes also showed significant declines in spike length in later harvest dates of 2017 (Figure 1b; Table S2), but not 2015 (Figure 1b; Table S1). In contrast to other seed yield traits, the average seed mass showed significant increases between the first and last harvests, especially in the shattering Ltc096 c/t genotypes (Figure 1c; Tables S1 and S2). However, the spike lengths and seed weights of shattering Ltc096 c/t genotypes were not significantly different from non-shattering Ltc096 c/t genotypes on the first harvest dates of 2015 or 2017 (Figure 1b; Tables S1 and S2).
Based on measurements of seed yield per spike, spike length, and seed mass, the calculated number of seeds per cm of spike showed significant declines between the first and last harvests (Figure 1d), like seed yield per spike (Figure 1a). However, these declines were significantly more pronounced in the shattering Ltc096 c/t genotypes than the non-shattering Ltc096 t/t genotypes. The estimated number of seeds per cm of spike of shattering Ltc096 c/t genotypes declined from 2.01 to 0.47 (76.7%) in 2015 (Figure 1d; Table S1) and from 2.43 to 0.59 (80.0%) in 2017 (Figure 1d; Table S2). In contrast, the non-shattering Ltc096 t/t genotypes showed smaller declines from 1.72 to 1.48 (14.0%) in 2015 (Figure 1d; Table S1) and from 2.04 to 1.53 (34.8%) in 2017 (Figure 1d; Table S2). Compared to non-shattering Ltc096 t/t genotypes, the shattering Ltc096 c/t genotypes had 16.9% to 19.1% (average 18.0%) more seeds per cm of spike on the first harvest dates of 2015 and 2017, respectively (Figure 1d; Tables S1 and S2). Conversely, the non-shattering Ltc096 t/t genotypes had 215% and 159% (average 188%) more seeds per cm of spike on the last harvest dates of 2015 and 2017, respectively, compared to the shattering Ltc096 c/t genotypes (Figure 1d; Tables S1 and S2). There was a significant cross-over interaction of genotype and harvest date for the number of seeds per cm of spike.
Averaged over both years, seed yield per spike declined 75.5% in shattering Ltc096 c/t genotypes versus 22.7% in non-shattering Ltc096 t/t genotypes. Similarly, the number of seeds per cm of spike declined 78.4% in shattering Ltc096 c/t genotypes versus 24.4% in non-shattering Ltc096 t/t genotypes, averaged over both years.
3.3. Effect of Harvest Date on Germination
Significant (p < 0.0001) effects of germination test, harvest date, germination count (germination test), and harvest date × germination count (germination test) for seed harvested in 2015 and 2017. Significant effects of genotype (p < 0.0001) and harvest date × germination test (p < 0.001) were also detected for seed harvested in 2017, but not for seed harvested in 2015. For seed harvested in 2015, the average percentage germination values across all germination counts and all three germination tests (Tables S3–S5), for each harvest date, were 73.4% (A) at 91 DAP, 70.2% (B) at 81 DAP, 65.3% (C) at 67 DAP, 64.2% (C) at 57 DAP, 59.3% (D) at 49 DAP, 56.3% (DE) at 39 DAP, 53.1% (E) at 31 DAP, and 39.9% (F) at 24 DAP, where different letters in parentheses indicate significant (p < 0.05) differences (Supplemental Data 2). For seed harvested in 2017, the average percentage germination values across all germination counts and all three germination tests (Tables S6–S8), for each harvest date, were 62.0% (AB) at 99 DAP, 61.8% (A) at 88 DAP, 59.8% at 71 DAP, 57.3% (DC) at 57 DAP, 53.2% (D) at 43 DAP, 45.4% (E) at 29 DAP, and 40.6% (F) at 22 DAP (Supplemental Data 2).
For seed harvested in 2017, the average percentage germination values across all germination counts and all three germination tests for the two Ltc096 genotypes were 55.6% (A) for the Ltc096 c/t genotype and 53.0% (B) for the Ltc096 c/t genotype (Supplemental Data 2). Thus, effects of genotype were relatively small (in 2017) or insignificant (in 2015) compared to the large significant effects of harvest date observed both years and the effect of genotype on germination will not be further considered.
The 4PL regression curves of percentage germination over time (DAI) from each of three germination tests on seed harvested in 2015 and 2017 (Figure 2) fit the observed percentage germination means for each count (Tables S3–S6), with corresponding regression coefficients () for each harvest date ranging from 0.791 to 0.986 in 2015 (Table 1) and from 0.858 to 0.989 in 2017 (Table 2). These 4PL regression analyses (Figure 2) include comparisons of the upper germination limits, number of days required to reach 50% germination, and rate of germination at midpoint (Table 1 and Table 2), which provide additional insights on the effects of harvest date on seed germination.
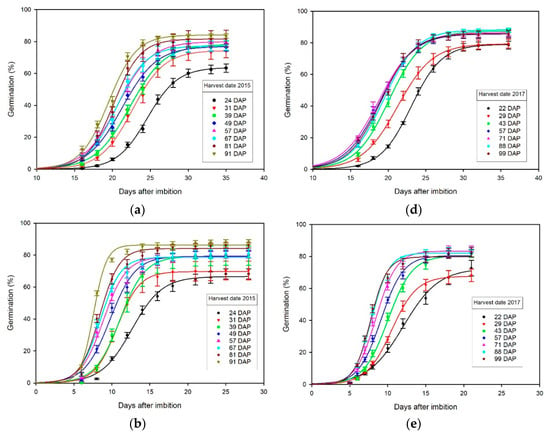
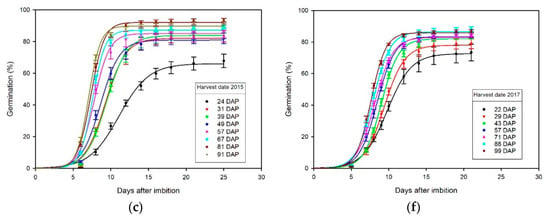
Figure 2.
Graphical comparisons of four-parameter logistic (4PL) regression curves of percentage germination over time (days after imbibition), among different harvest dates reported days after pollination (DAP) from germination tests performed: (a) 226 DAP with 14 days cold stratification for seed harvested in 2015; (b) 266 DAP for seed harvested in 2015; (c) 1025 DAP for seed harvested in 2015; (d) 225 DAP with 14 days cold stratification for seed harvested in 2017; (e) 265 DAP with 0 DPC for seed harvested in 2017; (f) 846 DAP with 0 DPC for seed harvested in 2017.

Table 1.
Four-parameter logistic (4PL) regression parameter estimates for percentage germination in response to the number of days after imbibition for seeds harvested in 2015 across eight different harvest dates numbered by days after pollination (DAP). Standard errors are shown in parentheses.
Table 2.
Four-parameter logistic (4PL) regression parameter estimates for percentage germination in response to the number of days after imbibition for seeds harvested in 2017 across eight different harvest dates numbered by days after pollination (DAP). Standard errors are shown in parentheses.
The upper percentage germination limits, , were significantly higher at later harvest dates compared to earlier harvest dates in 2015 (Table 1) and 2017 (Table 2). In 2015, the average upper germination limit of seed harvested on the last harvest date (87.1%) was approximately 26.4% higher than seed harvested on the first date (68.9%) across three different germination tests (Table 1). In 2017, the average upper percentage germination of seed harvested on the last harvest date (85.7%) was approximately 14.0% higher than seed harvested on the first date (75.2%), across three different germination tests (Table 2). Thus, the upper percentage germination limit of seed harvested on the last harvest date was approximately 20.5% higher than that for seed harvested on the first harvest date. The upper percentage germination limits of cold-stratified seed tended to be lower across different harvest dates in 2015 (15_GT1, Table 1), whereas they tended to be higher after a period of extended storage of 1025 DAP in 2015 (15_GT3, Table 1) except for the earliest harvest, 24 DAP, which was noticeably lower in all three tests (Table 1; Figure 2a–c). However, there were no consistent differences in the upper germination limits among the three different tests (17_GT1, 17_GT_2, or 17_GT3) conducted for seed harvested in 2017 (Table 2).
Estimates of the number of days, after imbibition (DAI), required to reach 50% of the upper percentage germination limit, , showed significant improvement (decrease) at later harvest dates in all germination tests of seed harvested in 2015 (Table 1) and 2017 (Table 2). In 2015, seed harvested on the last harvest date required an average of 11.4 days to reached 50% germination, across three different germination tests (Table 1), which was approximately 4.6 days (28.8%) faster than seed harvested on the first date (16.0 days average). In 2017, seed harvested on the last harvest date required an average of 11.5 days to reached 50% germination, across three different germination tests (Table 2), which was approximately 3.6 days (23.8%) faster than seed harvested on the first date (15.1 days average). Thus, seed from the last harvest date germinated approximately 4.1 days (26.3%) faster than seed harvested on the first harvest date, over both years.
The slopes of the regression lines at 50% of the upper germination limit (rates of germination at the midpoint), , were significantly higher for seeds that were cold-stratified (GT1) versus not cold-stratified (GT2 and GT3) in 2015 (Table 1) and 2017 (Table 2). This trend was especially true for seeds harvested at earlier harvest dates where the rates of germination at midpoint were approximately two times greater in GT1 with cold stratification (Table 1 and Table 2). For seeds that were not cold-stratified (GT2 and GT3), the slopes of the regression lines at the midpoint of germination tended to be significantly higher in later harvest dates compared to earlier harvest dates in 2015 (Table 1) and 2017 (Table 2), but this trend was not observed for cold-stratified seed (GT1). In fact, the rates of germination at midpoint tended to be significantly higher for earlier harvest dates compared to later harvest dates for cold-stratified seed in 2017 (17_GT1, Table 2). Thus, cold stratification showed interactions with harvest date for rates of germination at the midpoint of upper and lower percentage germination limits.
4. Discussion
4.1. Seed Yield Benefits of the sh6 Seed-Retention Gene
The SH6 seed-shattering gene was originally discovered in an experiment designed to identify the number, location, and relative effects of QTLs controlling seed production traits in a L. triticoides backcross population derived from L. cinereus × L. triticoides hybrids [22]. The previous study identified three QTLs associated with the number of florets per inflorescence, one QTL for percentage seed set (fertility), one QTL for seed mass, two QTLs for seed dormancy, and three QTLs for seed shattering, including a relatively strong seed-shattering QTL on LG6a (SH6) [22]. This study provided several insights on the reported effects of the dominant SH6 L. cinereus shattering and recessive sh6 L. triticoides seed-retention gene alleles, which were first discovered in an L. triticoides backcross population [22].
This study quantifies the rate of seed yield loss due to the SH6 seed-shattering gene over time under natural growing conditions. Previous studies reported a 45.8% difference, between SH6/sh6 and sh6/sh6 genotypes, in response to mechanical agitation in the field and laboratory [22], but there was no meaningful measurement of how fast or how much shattering occurred under natural field conditions and what this meant for commercial seed producers. This study showed that significant losses were detected as early as 31 DAP in 2015 or 29 DAP in 2017. Moreover, the average seed yield of shattering Ltc096 genotypes (c/t) declined by 75.5% over a period of 65 days (approximately 1.2% per day) out to 91 DAP in 2015 and 99 DAP in 2017. In contrast, the average seed yield of non-shattering Ltc096 genotypes (c/t) declined 22.7% loss over a period the same period (approximately 0.3% per day). Although seed yields of shattering genotypes were 19.4% greater than non-shattering genotypes on the first harvest date, yields of non-shattering genotypes were 167% greater on the last harvest date. Moreover, the Ltc096 marker may underestimate differences between SH6/sh6 and sh6/sh6 genotypes due to genetic recombination between the gene and marker since it is not easy to unambiguously distinguish the SH6/sh6 and sh6/sh6 genotypes based on phenotype and it seems as though there was evidence of recombination between the marker (Lt096) and gene (SH6) in the L57S population, which is not surprising after six generations of recombination. Although two other seed-shattering QTLs have been identified in the L. triticoides × L. cinereus crosses [22], the locations of these two other genes are much less precise than SH6 [22] and we believe that it would have been extremely difficult to select these two other QTLs in the C6 breeding population after six generation of genetic recombination.
One possible concern with these results is our finding that the seed yield of shattering genotypes was significantly higher than non-shattering genotypes on the first harvest date, suggesting that there could be a yield penalty associated with the L. triticoides sh6 seed-retention gene. Previous studies did not detect any significant seed yield traits associated with the SH6 seed-shattering gene in the L. triticoides backcross population [22]. However, QTLs for inflorescence (spike) length, spikelets per node, and nodes per spike per spikelet were detected on the long arm of LG6a approximately 10 to 60 centiMorgans distal from the Ltc096 marker [39]. It was shown that L. triticoides has significantly smaller spikes compared to L. cinereus [22,39], which is obvious visually, so it is quite possible that the L. triticoides sh6 seed-retention gene was linked to negative seed yield genes in the L57S-577 sh6 donor parent. Additional marker-assisted selection and backcrossing breeding efforts are need to introgress and fix the recessive L. triticoides sh6 seed-retention gene in L. cinereus germplasm and eliminate any possible linkage drag effects of other L. triticoides genes. Evidence from this study also suggests that additional research is needed to identify markers that are more closely linked to the recessive L. triticoides sh6 seed-retention gene and potentially clone this gene, which does not correspond to other seed-shattering genes that have been cloned in rice [40,41,42,43,44,45,46] or any other species [47,48,49,50,51] based on chromosome alignments and synteny with other species [22,39].
Although seed yields of shattering genotypes declined, the average seed mass showed significant increases at later harvest dates, especially in shattering genotypes. Most seed shattering in L. cinereus, and other Leymus species, occurs by breakage of the rachilla within the spikelet. Proximal florets are larger and likely to form larger seeds and distal florets are more likely (usually) empty. We speculate that increases in the average seed size at later harvest dates, especially in shattering genotypes, was cause by disarticulation (shattering) of smaller distal florets that have smaller seeds or no viable seed whatsoever. Thus, sh6 seed retention provides an effective means to retain larger seeds and potentially improve germination characteristics of the seed, which may translate to better outcomes in rangeland plantings.
The average spike length tended to decrease in later harvest dates, but these trends were not consistent (Figure 1c). These modest decreases in spike length may have three explanations. First, small decreases in spike length can be attributed to disarticulation of florets on the terminal spikelets, especially in the sh6/sh6 seed-shattering genotypes. Second, there may have been inadvertent bias to harvest taller stems with larger spikes in the early harvests even though we tried to sample spikes randomly. Third, breaks may occasionally happen at the terminal end of the spike where the rachis is thin, even though the main spike does not normally disarticulate by abscission (a trait known as “brittle rachis”) in these species. However, we do not think that there is substantial loss of seed caused by breaks in the rachis.
4.2. Germination Effects of Harvest Date
This study also demonstrated that the positive seed germination effects associated with the recessive L. triticoides sh6 seed-retention gene [22] were mainly a function of harvest date rather than the SH6 gene per se. In fact, the L. triticoides sh6 seed-retention gene was actually associated with a small but significant decrease in germination rates for seed harvested in 2017, which is contrary to what was previously reported [22]. Previous studies reported that homozygous marker genotypes (t/t), for the LG6a seed-shattering QTL, required 1.64 fewer days to germinate compared to the heterozygous (c/t) genotypes [22]. However, it was not clear whether this effect was due to negative germination effects of the L. cinereus seed-shattering gene per se, or whether it was indirectly caused by earlier harvest of seed from shattering plants, since it was known that the non-shattering parent L. triticoides generally displays slow germination resulting from seed dormancy [22,31]. This study showed that seed harvested on the last harvest date reached 50% germination 4.2 days (26.4%) faster and 20.5% higher upper percentage germination limits than seed harvested on the first harvest date. It has been shown that timing of grass seed harvest is critical to the germination and quality characteristics of grass seed [29,52]. Germination differences between early and late harvest dates could result from at least two or three different factors, including harvest of physiologically immature seed, damage to seed during storage, or release of seed dormancy as a result of thermal after-ripening in the field.
Extended seed storage seemed to improve seed germination of seed harvested at early harvest dates, reducing differences between harvest dates (Figure 2), suggesting a response to after-ripening in earlier-harvested seeds with greater dormancy. One possible exception was that seed from the first harvest date, in 2015, did not show any improvement in germination, comparing non-stratified germination tests conducted at 266 DAP (15_GT2) and 1025 DAP (15_GT3) (Table 1, Figure 2). Relatively poor performance of seed harvested on the first harvest date of 2015, even after extended storage and after-ripening (Figure 2c), suggests that some of the seeds may not be physiologically mature on the first harvest date even though some of the seeds began to disarticulate shortly after this harvest (Figure 1). Comparisons of seed yield (Figure 1) and seed germination (Figure 2) across early harvest dates suggest that some seeds or plants may not be physiologically mature while other seeds or plants begin to shatter, especially for genetically heterogeneous outcrossing plants such L. cinereus. Thus, seed quality may be a concern if seed growers harvest seed before it begins to shatter.
It is unlikely that seed from early harvest dates was damaged in a way that would explain steady improvements in seed germination associated with later harvest dates (Table 1 and Table 2; Figure 2). Our method of drying spikes in loose paper bags in a ventilated greenhouse is considered the best practical method by grass breeders in our laboratory for decades, and it is likely as good or better than many commercial seed-storage methods. In fact, release of seed dormancy is facilitated by increasing thermal temperature up to at least 50° C in Lolium ridgidum [53], and it is well documented that after-ripening is accelerated by warmer temperatures in Hordeum vulgare L., Avena fatua L. and Bromus tectorum L. [54,55,56] and other winter annuals [57,58]. Moreover, this germination effect has been reproduced over two years in this study and two years in a previous study [22].
Regardless of what caused differences in germination between earlier and later harvest dates, in this study, the most practical question is whether or not the benefits of later harvest dates, made possible by the sh6 seed-retention gene, can translate to benefits in the quality of seed produced by commercial seed growers? More specifically, what if any benefits can be gained in terms of seedling establishment in fire rehabilitation and other large-scale revegetation plantings? Further research is needed to introgress and fix the recessive L. triticoides sh6 seed-retention gene into L. cinereus germplasm, eliminating any potentially negative seed dormancy or low-seed yield genes from L. triticoides that are not directly associated with its sh6 seed-retention allele. It is possible that complete elimination of L. triticoides seed dormancy genes could mitigate some the germination problems associated with early harvest dates observed in this study but not if some of the seeds were harvested before physiological maturity. In any case, development of a non-shattering L. cinereus germplasm or variety will allow testing in rangeland planting experiments [9,10], commercial seed production, and large-scale revegetation projects.
5. Conclusions
The Ltc096 EST marker and sh6/sh6 genotypes selected from cycle six of a L. triticoides × L. cinereous breeding population in this project can be used to further introgress and fix the sh6 seed-retention gene in basin wildrye to effectively eliminate seed shattering and possibly improve seed germination by helping prevent harvest of immature seed, enabling extended seed ripening in the field, and enabling seed producers to harvest seed during optimal weather conditions. Elimination of seed shattering will have benefits to basin wildrye seed growers by eliminating the need to windrow seed fields and eliminating worries about interactions between their ability to harvest seed during inclement weather, seed losses due to shattering, or weather damage to windrowed seed. Improvements in seed germination and seedling vigor will have substantial benefits to land managers that have limited resources and need successful native plant stand establishment for fire rehabilitation and other large-scale revegetation projects in arid and semiarid environments of western North America.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4395/10/11/1740/s1, Supplemental Data 1: Seed yield data and analyses; Supplemental Data 2: Germination data and analyses, Table S1: Yield trait means 2015; Table S2: Yield trait means 2017; Table S3: Germination means from 15_GT1; Table S4: Germination means from 15_GT2; Table S5: Germination means from 15_GT3; Table S6: Germination means from 17_GT1; Table S7: Germination means from 17_GT2; Table S8: Germination means from 17_GT3.
Author Contributions
Conceptualization, S.R.L. and T.A.J.; methodology, S.R.L. and T.A.J.; formal analysis, S.R.L. and B.L.W.; investigation, S.R.L. and T.A.J.; resources, L.M.J., S.R.L., and T.A.J.; data curation, L.M.J. and S.R.L.; writing—original draft preparation, S.R.L.; writing—review and editing, B.L.W., S.R.L. and T.A.J.; visualization, S.R.L. and B.L.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors wish to thank Kevin B. Jensen for administrative support and manuscript review.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pilliod, S.D.; Welty, J.L.; Toevs, G.R. Seventy-Five Years of Vegetation Treatments on Public Rangelands in the Great Basin of North. America. Rangelands 2017, 39, 1–9. [Google Scholar] [CrossRef]
- Duniway, C.M.; Palmquist, E.; Miller, M.E. Evaluating rehabilitation efforts following the Milford Flat Fire: Successes, failures, and controlling factors. Ecosphere 2015, 6, 80. [Google Scholar] [CrossRef]
- Peppin, D.; Fulé, P.Z.; Sieg, C.H.; Beyers, J.L.; Hunter, M.E. Post-wildfire seeding in forests of the western United States: An. evidence-based review. Forest Ecol. Manag. 2010, 260, 573–586. [Google Scholar] [CrossRef]
- James, J.; Svejcar, T.; Rinella, M. Demographic processes limiting seedling recruitment in arid grassland restoration. J. Appl. Ecol. 2011, 48, 961–969. [Google Scholar] [CrossRef]
- Larson, J.E.; Sheley, R.L.; Hardegree, S.P.; Doescher, P.S.; James, J.J. Seed and seedling traits affecting critical life stage transitions and recruitment outcomes in dryland grasses. J. Appl. Ecol. 2015, 52, 199–209. [Google Scholar] [CrossRef]
- Leger, E.A.; Atwater, D.Z.; James, J.J. Seed and seedling traits have strong impacts on establishment of a perennial bunchgrass in invaded semi-arid systems. J. Appl. Ecol. 2019, 56, 1343–1354. [Google Scholar] [CrossRef]
- Knutson, K.C.; Pyke, D.A.; Wirth, T.A.; Arkle, R.S.; Pilliod, D.S.; Brooks, M.L.; Chambers, J.C.; Grace, J.B. Long-term effects of seeding after wildfire on vegetation in Great Basin shrubland ecosystems. J. Appl. Ecol. 2014, 51, 1414–1424. [Google Scholar] [CrossRef]
- Madsen, M.D.; Davies, K.W.; Boyd, C.S.; Kerby, J.D.; Svejcar, T.J. Emerging seed enhancement technologies for overcoming barriers to restoration. Restor. Ecol. 2016, 24, S77–S84. [Google Scholar] [CrossRef]
- Rigby, C.W.; Jensen, K.B.; Creech, J.E.; Thacker, E.T.; Waldron, B.L.; Derner, J.D. Establishment and Trends in Persistence of Selected Perennial Cool-Season Grasses in Western United States. Rangel. Ecol. Manag. 2018, 71, 681–690. [Google Scholar] [CrossRef]
- Robins, J.G.; Jensen, K.B.; Jones, T.A.; Waldron, B.L.; Peel, M.D.; Rigby, C.W.; Vogel, K.P.; Mitchell, R.B.; Palazzo, A.J.; Cary, T.J. Stand. establishment and persistence of perennial cool-season grasses in the intermountain west and the central and northern great plains. Rangel. Ecol. Manag. 2013, 66, 181–190. [Google Scholar] [CrossRef]
- Brummer, E.C.; Barber, W.T.; Collier, S.M.; Cox, T.S.; Johnson, R.; Murray, S.C.; Olsen, R.T.; Pratt, R.C.; Thro, A.M. Plant breeding for harmony between agriculture and the environment. Front. Ecol. Environ. 2011, 9, 561–568. [Google Scholar] [CrossRef]
- DeHaan, L.R.; van Tassel, D.L.; Cox, T.S. Perennial grain crops: A synthesis of ecology and plant breeding. Renew. Agric. Food Syst. 2007, 20, 5–14. [Google Scholar] [CrossRef]
- Jones, T.A.; Monaco, T.A.; Rigby, C.W. The potential of novel native plant materials for the restoration of novel ecosystems. Elem. Sci. Anth. 2015, 3. [Google Scholar] [CrossRef]
- Cruz, R.; Ganskopp, D. Seasonal preferences of steers for prominent northern great basin grasses. J. Range Manag. 1998, 51, 557. [Google Scholar] [CrossRef]
- Ganskopp, D.; Bohnert, D. Nutritional dynamics of 7 northern great basin grasses. J. Range Manag. 2001, 54. [Google Scholar] [CrossRef]
- Ganskopp, D.; Aguilera, L.; Vavra, M. Livestock Forage Conditioning Among Six Northern Great Basin Grasses. Rangel. Ecol. Manag. 2007, 60, 71–78. [Google Scholar] [CrossRef]
- Ogle, D.G.; Tilley, D.; John, L.S. Plant Guide for Basin Wildrye (Leymus cinereus); USDA NRCS Plant Materials Center: Aberdeen, ID, USA, 2012.
- Culumber, C.M.; Larson, S.R.; Jones, T.A.; Jensen, K.B. Wide-scale population sampling identifies three phylogeographic races of basin wildrye and low-level genetic admixture with creeping wildrye. Crop Sci. 2013, 53, 996–1007. [Google Scholar] [CrossRef]
- Larson, S.R.; Pearson, C.H.; Jensen, K.B.; Jones, T.A.; Mott, I.W.; Robbins, M.D.; Staub, J.E.; Waldron, B.L. Development and testing of cool-season grass species, varieties and hybrids for biomass feedstock production in western North. America. Agronomy 2017, 7, 3. [Google Scholar] [CrossRef]
- Porensky, L.M.; Davison, J.; Leger, E.A.; Miller, W.W.; Goergen, E.M.; Espeland, E.K.; Carroll-Moore, E.M. Grasses for biofuels: A low water-use alternative for cold desert agriculture? Biomass Bioenergy 2014, 66, 133–142. [Google Scholar] [CrossRef]
- Larson, S.R.; Wu, X.; Jones, T.A.; Jensen, K.B.; Chatterton, N.J.; Waldron, B.L.; Robins, J.G.; Bushman, B.S.; Palazzo, A.J. Comparative mapping of growth habit, plant height, and flowering QTLs in two interspecific families of Leymus. Crop Sci. 2006, 46, 2526–2539. [Google Scholar] [CrossRef]
- Larson, S.R.; Kellogg, E.A. Genetic dissection of seed production traits and identification of a major-effect seed retention QTL in hybrid Leymus (Triticeae) wildryes. Crop Sci. 2009, 49, 29–40. [Google Scholar] [CrossRef]
- Young, J.A.; Evans, R.A. Germination of great basin wildrye seeds collected from native stands1. Agron. J. 1981, 73, 917–920. [Google Scholar] [CrossRef]
- Roundy, B.A. Response of basin wildrye and tall wheatgrass seedlings to salination1. Agron. J. 1983, 75, 67–71. [Google Scholar] [CrossRef]
- Roundy, B.A.; Young, J.A.; Evans, R.A. Germination of basin wildrye and tall wheatgrass in relation to osmotic and matric potential1. Agron. J. 1985, 77, 129–135. [Google Scholar] [CrossRef]
- Evans, R.A.; Young, J.A. Magnar basin wildrye: Germination in relation to temperature. J. Range Manag. 1983, 36, 395–398. [Google Scholar] [CrossRef]
- Robins, J.G.; Bushman, B.S.; West, M.S. Effects of Selection for Seedling Vigor on the Genetic Variation in Leymus cinereus. Rangel. Ecol. Manag. 2017, 70, 504–508. [Google Scholar] [CrossRef]
- Jones, T.A.; Parr, S.D.; Winslow, S.R.; Rosales, M.A. Notice of release of ‘Continental’basin wildrye. Nativ. Plants J. 2009, 10, 57–61. [Google Scholar] [CrossRef]
- Berdahl, J.D.; Frank, A.B. Seed maturity in four cool-season forage grasses. Agron. J. 1998, 90, 483–488. [Google Scholar] [CrossRef]
- USDA-NRCS. Conservation Plant Release Brochure for ’Rio’ Beardless Wild Rye (Leymus Triticoides Buckley); USDA NRCS Plant Materials Center: Lockeford, CA, USA, 2014.
- Knapp, A.D.; Wiesner, L.E. Seed dormancy of beardless wildrye (Elymus triticoides Buckl.). J. Seed Technol. 1978, 3, 1–9. [Google Scholar]
- Larson, S.R.; Jensen, K.B.; Robins, J.G.; Waldron, B.L. Genes and quantitative trait. loci controlling biomass yield and forage quality traits in perennial wildrye. Crop Sci. 2014, 54, 111–126. [Google Scholar] [CrossRef]
- Yun, L.; Larson, S.R.; Jensen, K.B.; Staub, J.E.; Grossl, P.R. Quantitative trait loci (QTL) and candidate genes associated with trace element concentrations in perennial grasses grown on phytotoxic soil contaminated with heavy metals. Plant Soil 2015, 396, 277–296. [Google Scholar] [CrossRef]
- Yun, L.; Larson, S.R.; Mott, I.W.; Jensen, K.B.; Staub, J.E. Genetic control of rhizomes and genomic localization of a major-effect growth habit QTL in perennial wildrye. Mol. Genet. Genom. 2014, 289, 383–397. [Google Scholar] [CrossRef]
- Bushman, B.S.; Larson, S.R.; Mott, I.W.; Cliften, P.F.; Wang, R.R.C.; Chatterton, N.J.; Hernandez, A.G.; Ali, S.; Kim, R.W.; Thimmapuram, J.; et al. Development and annotation of perennial Triticeae ESTs and SSR markers. Genome 2008, 51, 779–788. [Google Scholar] [CrossRef]
- Kennard, C.; Phillips, L.; Porter, A. Genetic dissection of seed shattering, agronomic, and color traits in American wildrice (Zizania palustris var. interior L.) with a comparative map. Theor. Appl. Genet. 2002, 105, 1075–1086. [Google Scholar] [CrossRef]
- Larson, S.R.; Kishii, M.; Tsujimoto, H.; Qi, L.; Chen, P.; Lazo, G.R.; Jensen, K.B.; Wang, R.R.C. Leymus EST linkage maps identify 4NsL-5NsL reciprocal translocation, wheat-Leymus chromosome introgressions, and functionally important gene loci. Theor. Appl. Genet. 2012, 124, 189–206. [Google Scholar] [CrossRef]
- Benham, J.J. Genographer Version 1.6.0; Montana State University: Bozeman, MT, USA, 2001. [Google Scholar]
- Larson, S.R.; Kellogg, E.A.; Jensen, K.B. Genes and QTLs Controlling Inflorescence and Stem Branch Architecture in Leymus (Poaceae: Triticeae) Wildrye. J. Hered. 2013, 104, 678–691. [Google Scholar] [CrossRef]
- Konishi, S.; Izawa, T.; Lin, S.Y.; Ebana, K.; Fukuta, Y.; Sasaki, T.; Yano, M.A. SNP caused loss of seed shattering during rice domestication. Science 2006, 312, 1392–1396. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhou, A.; Sang, T. Rice domestication by reducing shattering. Science 2006, 311, 1936–1939. [Google Scholar] [CrossRef]
- Sang, T.; Ge, S. The puzzle of rice domestication (seed shatter disarticulation). J. Integr. Plant Biol. 2008, 49, 760–768. [Google Scholar] [CrossRef]
- Win, K.T.; Yamagata, Y.; Doi, K.; Uyama, K.; Nagai, Y.; Toda, Y.; Kani, T.; Ashikari, M.; Yasui, H.; Yoshimura, A. A single base change explains the independent origin of and selection for the nonshattering gene in African rice domestication. New Phytol. 2017, 213, 1925–1935. [Google Scholar] [CrossRef]
- Yoon, J.; Cho, L.H.; Antt, H.W.; Koh, H.J.; An, G. KNOX Protein OSH15 Induces Grain Shattering by Repressing Lignin Biosynthesis Genes. Plant Physiol. 2017, 174, 312–325. [Google Scholar] [CrossRef]
- Yoon, J.; Cho, L.H.; Kim, S.L.; Choi, H.; Koh, H.J.; An, G. The BEL1-type homeobox gene SH5 induces seed shattering by enhancing abscission-zone development and inhibiting lignin biosynthesis. Plant J. 2014, 79, 717–728. [Google Scholar] [CrossRef]
- Zhou, Y.; Lu, D.; Li, C.; Luo, J.; Zhu, B.F.; Zhu, J.; Shangguan, Y.; Wang, Z.; Sang, T.; Zhou, B.; et al. Genetic control of seed shattering in rice by the APETALA2 transcription factor Shattering Abortion1. Plant Cell 2012, 24, 1034–1048. [Google Scholar] [CrossRef] [PubMed]
- Doebley, J. Unfallen grains: How ancient farmers turned weeds into crops. Science 2006, 312, 1318–1319. [Google Scholar] [CrossRef]
- Li, W.; Gill, B.S. Multiple genetic pathways for seed shattering in the grasses. Funct. Integr. Genom. 2006, 6, 300–309. [Google Scholar] [CrossRef]
- Pourkheirandish, M.; Hensel, G.; Kilian, B.; Senthil, N.; Chen, G.; Sameri, M.; Azhaguvel, P.; Sakuma, S.; Dhanagond, S.; Sharma, R.; et al. Evolution of the grain dispersal system in barley. Cell 2015, 162, 527–539. [Google Scholar] [CrossRef]
- Sang, T. Genes and Mutations Underlying Domestication Transitions in Grasses. Plant Physiol. 2009, 149, 63–70. [Google Scholar] [CrossRef]
- Balanzà, V.; Roig-Villanova, I.; Di Marzo, M.; Masiero, S.; Colombo, L. Seed abscission and fruit dehiscence required for seed dispersal rely on similar genetic networks. Development 2016, 143, 3372–3381. [Google Scholar] [CrossRef]
- Hill, M.J.; Watkin, B.R. Seed production studies on perennial ryegrass, timothy and prairie grass: 2. Changes in physiological components during seed development and time and method of harvesting for maximum seed yield. Grass Forage Sci. 1975, 30, 131–140. [Google Scholar] [CrossRef]
- Steadman, K.J.; Crawford, A.D.; Gallagher, R.S. Dormancy release in Lolium rigidum seeds is a function of thermal after-ripening time and seed water content. Funct. Plant Biol. 2003, 30, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Favier, F.J.; Woods, J.L. The quantification of dormancy loss in barley (Hordeum vulgare L.). Seed Sci. Technol. 1993, 21, 653–674. [Google Scholar]
- Foley, M.E. Temperature and water status of seed affect afterripening in wild oat (Avena fatua). Weed Sci. 1994, 42, 200–204. [Google Scholar] [CrossRef]
- Bauer, M.C.; Meyer, S.E.; Allen, P.S. A simulation model to predict seed dormancy loss in the field for Bromus tectorum L. J. Exp. Bot. 1998, 49, 1235–1244. [Google Scholar]
- Baskin, J.M.; Baskin, C.C. High temperature requirement for afterripening in seeds of winter annuals. New Phytol. 1976, 77, 619–624. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Temperature requirements for after-ripening in seeds of nine winter annuals. Weed Res. 1986, 26, 375–380. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).