The Expression of ELF4-Like Genes Is Influenced by Light Quality in Petunia

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Precultivation
2.2. Experimental Growth Conditions
2.3. Gene Identification by 3′EST High Throughput Sequencing
2.4. RACE PCR for Isolation of ELF4 Like Genes
2.5. Expression Analysis of ELF4-Like Genes and Recordings of Developmental Parameters
- (a)
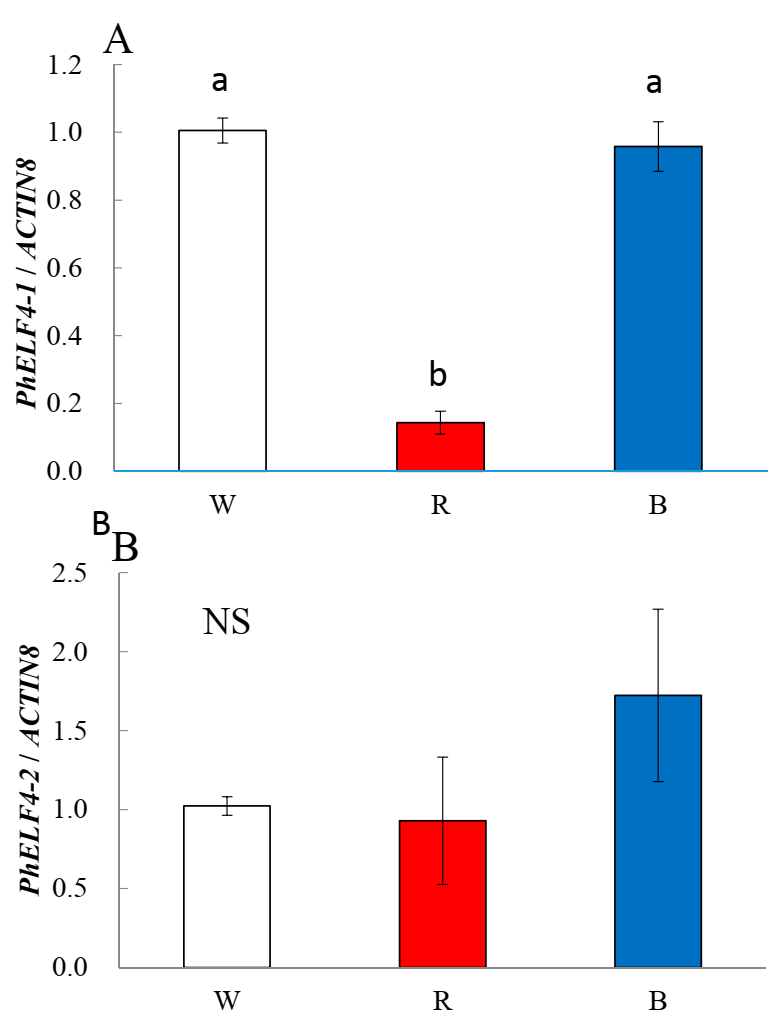
- Leaves from 3 plants per light quality treatment were sampled after 12 h of lighting the second day under the different light quality treatments of a 14 h photoperiod.
- (b)
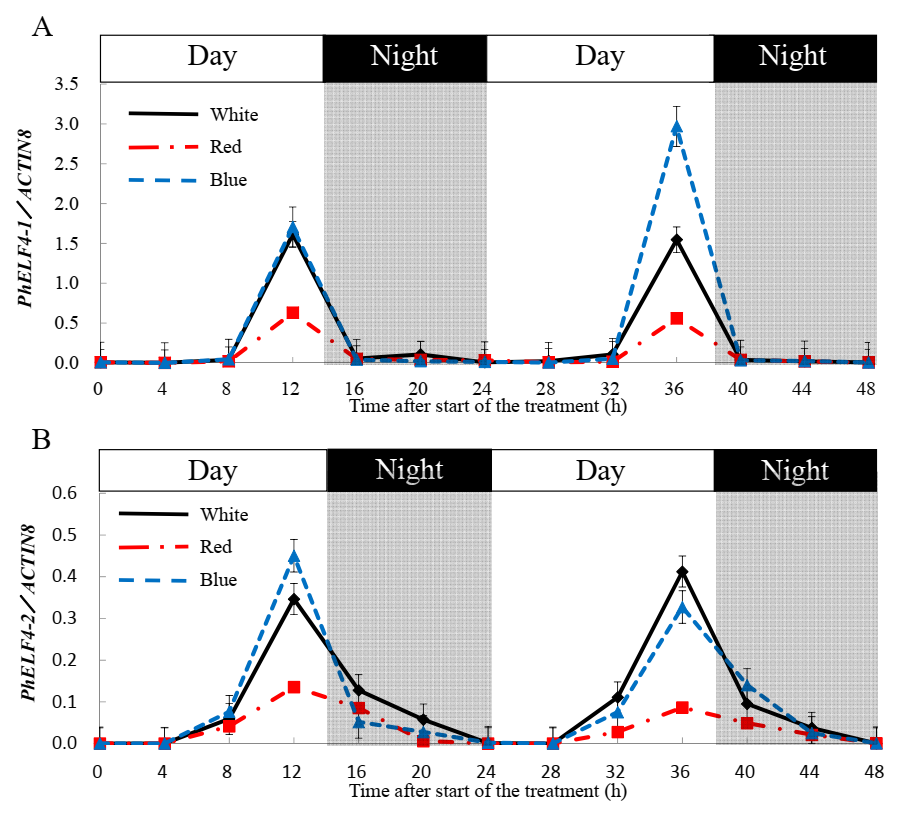
- Leaves from altogether 39 plants were sampled from each light quality treatment at 0, 4, 8, 12, 16, 20, 24, 28, 32, 36, 40, 44 and 48 h after the start of the treatments of a 14 h photoperiod.
- (c)
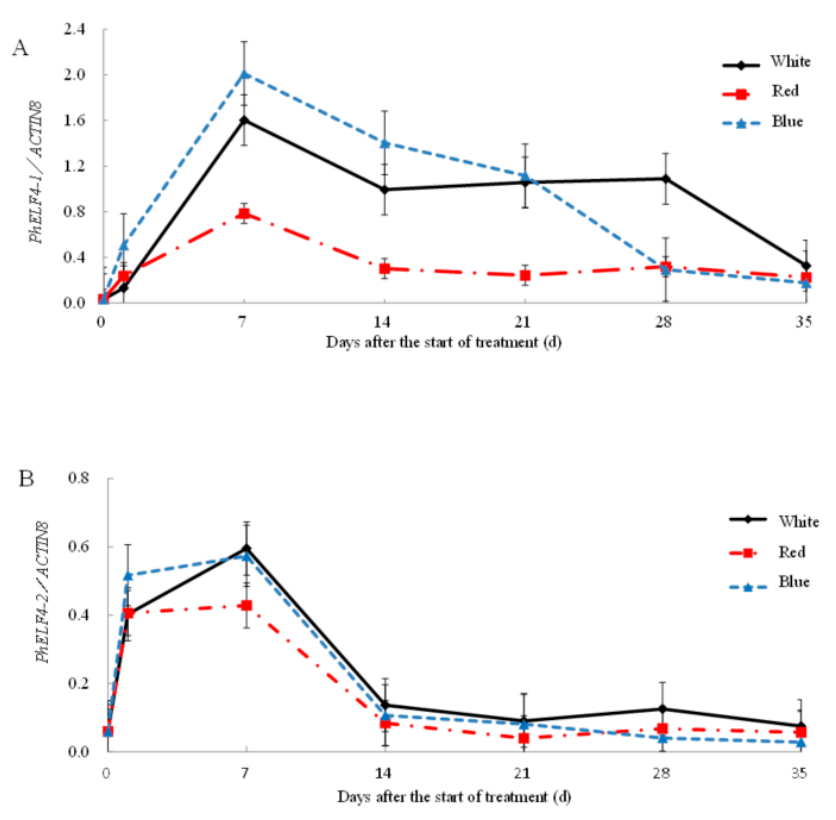
- Leaves from altogether 18 plants were sampled from each light quality treatment at 0, 7, 14, 21, 28 and 35 days after the start of the different light quality treatments, with harvest 12 h after the start of the daily lighting of a 14 h photoperiod.
- (d)
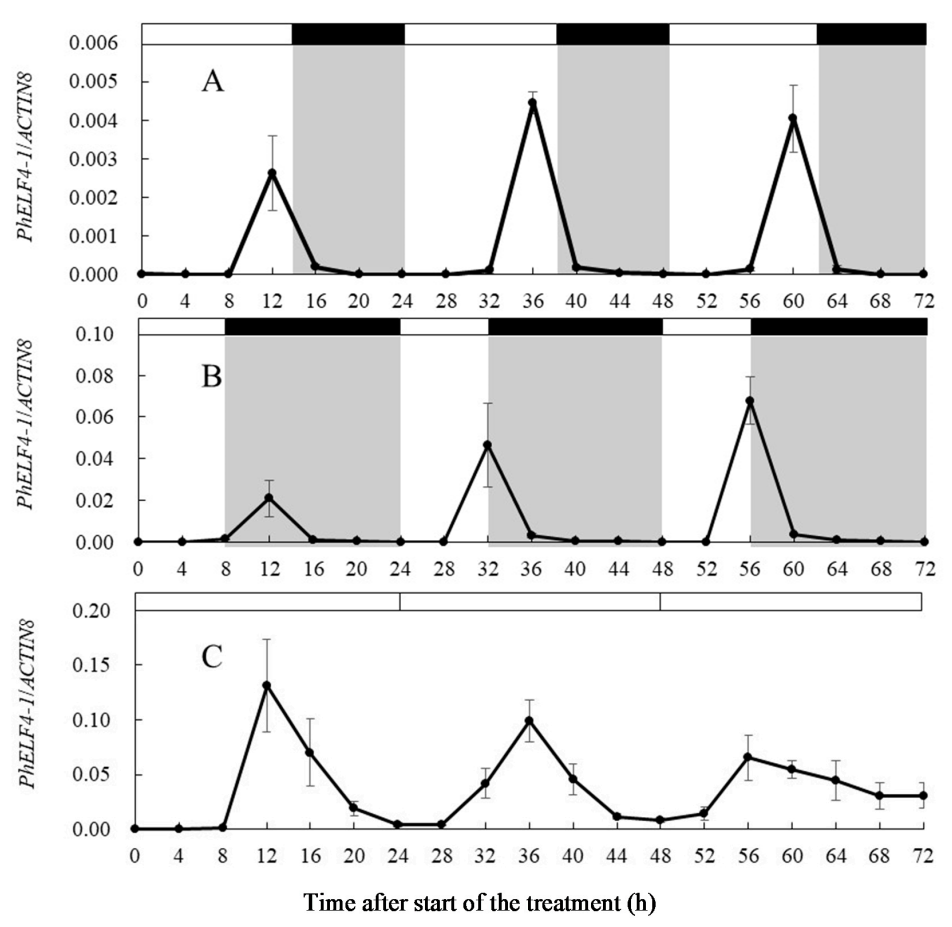
- Leaves from altogether 57 plants were sampled under each of three photoperiods of 8, 14 and 24 h provided by B LEDs (day/night of 14 h/10 h, 8 h/16 h and 24 h/ 0 h) at 0, 4, 8, 12, 16, 20, 24, 28, 32, 36, 40, 44, 48, 52, 56, 60, 64, 68 and 72 h after the start of the B LED treatment. In addition, after 4 weeks under these photoperiods with B light, plant height, number of leaves per plant, number of lateral shoots per plant and percentage of plants that had at least one fully opened flower, were recorded for 6 plants per photoperiod.
2.6. Statistical Analyses
3. Results
3.1. Floral Induction and Light Signal Transduction Genes Expressed in Petunia under Different Light Qualities as Revealed by 3′EST High Throughput Sequencing
3.2. Identification of Two Full Length ELF4-Like Genes in Petunia
3.3. Gene Expression of ELF4-Like Genes in Petunia under Different Light Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lin, C. Photoreceptors and regulation of flowering time. Plant Physiol. 2000, 123, 39–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mockler, T.; Yang, H.; Yu, X.H.; Parikh, D.; Cheng, Y.; Dolan, S.; Lin, C. Regulation of photoperiodic flowering by Arabidopsis photoreceptors. Proc. Natl. Acad. Sci. USA 2003, 100, 2140–2145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerdán, P.D.; Chory, J. Regulation of flowering time by light quality. Nature 2003, 423, 881–885. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yu, X.; Li, K.; Klejnot, J.; Yang, H.; Lisiero, D.; Lin, C. Photoexcited CRY2 interacts with CIB1 to regulate transcription and floral initiation in Arabidopsis. Science 2008, 322, 1535–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.J.; Zhang, Y.C.; Li, Q.H.; Sang, Y.; Mao, J.; Lian, H.L.; Wang, L.; Yang, H.Q. COP1-mediated ubiquitination of CONSTANS is implicated in cryptochrome regulation of flowering in Arabidopsis. Plant Cell 2008, 20, 292–306. [Google Scholar] [CrossRef]
- Higuchi, Y.; Narumi, T.; Oda, A.; Nakano, Y.; Sumitomo, K.; Fukai, S.; Hisamatsu, T. The gated induction system of systemic floral inhibitor, antiflorigen, determines obligate short-day flowering in chrysanthemums. Proc. Natl. Acad. Sci. USA 2013, 110, 17137–17142. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Araki, T.; Endo, M. Integration of input signals into the gene network in the plant circadian clock. Plant Cell Physiol. 2017, 58, 977–982. [Google Scholar] [CrossRef]
- Yeom, M.; Kim, H.; Lim, J.; Shin, A.-Y.; Hong, S.; Kim, J.-I.; Nam, H.G. How do phytochromes transmit the light quality information to the circadian clock in Arabidopsis? Mol. Plant 2014, 7, 1701–1704. [Google Scholar] [CrossRef] [Green Version]
- Doyle, M.R.; Davis, S.J.; Bastow, R.M.; McWatters, H.G.; Kozma-Bognar, L.; Nagy, F.; Millar, A.J.; Amasino, R.M. The ELF4 gene controls circadian rhythms and flowering time in Arabidopsis thaliana. Nature 2002, 419, 74–77. [Google Scholar] [CrossRef]
- Kim, Y.; Yeom, M.; Kim, H.; Lim, J.; Koo, H.J.; Hwang, D.; Somers, D.; Nam, H.G. GIGANTEA and EARLY FLOWERING 4 in Arabidopsis exhibit differential phase-specific genetic influences over a diurnal cycle. Mol. Plant 2012, 5, 678–687. [Google Scholar] [CrossRef] [Green Version]
- Bernie, G.; Perilleux, C. A physiological overview of the genetics of flowering time control. Plant Biotechnol. J. 2005, 3, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Eskins, K. Light quality effects on Arabidopsis development. Red, blue and far-red regulation of flowering and morphology. Physiol. Plant. 1992, 86, 439–444. [Google Scholar] [CrossRef]
- Adams, S.; Allen, T.; Whitelam, G.C. Interaction between the light quality and flowering time pathways in Arabidopsis. Plant J. 2009, 60, 257–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haliapas, S.; Yupsanis, T.A.; Syros, T.D.; Kofidis, G.; Economou, A.S. Petunia x hybrida during transition to flowering as affected by light intensity and quality treatments. Physiol. Plant. 2008, 30, 807–815. [Google Scholar] [CrossRef]
- Fukuda, N.; Ajima, C.; Yukawa, T.; Olsen, E.J. Antagonistic action of blue and red light on shoot elongation in petunia depends on gibberellin, but the effects on flowering are not generally linked to gibberellin. Environ. Exp. Bot. 2016, 121, 102–111. [Google Scholar] [CrossRef]
- Tsukamoto, A.; Hirai, T.; Chin, D.P.; Mii, M.; Mizoguchi, T.; Mizuta, D.; Yoshida, H.; Olsen, E.J.; Ezura, H.; Fukuda, N. The FT-like gene PehFT in petunia responds to photoperiod and light quality but is not the main gene promoting light quality-associated flowering. Plant Biotechnol. 2016, 33, 297–307. [Google Scholar] [CrossRef]
- Chevreux, B. Using the miraEST assembler for reliable and automated mRNA transcript assembly and SNP detection in sequenced ESTs. Genome Res. 2004, 14, 1147–1159. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, Q.; Liu, Y.; Zhao, X.; Imaizumi, T.; Somers, D.E.; Tobin, E.M.; Lin, C. Arabidopsis CRY2 and ZTL mediate blue-light regulation of the transcription factor CIB1 by distinct mechanisms. Proc. Natl. Acad. Sci. USA 2013, 110, 17582–17587. [Google Scholar] [CrossRef] [Green Version]
- Hara, M.; Kamada, H.; Mizoguchi, T. CO-EXPRESSED WITH CLOCK GENES LHY AND CCA1 1 (CEC1) is regulated by LHY and CCA1 and plays a key role in phase setting of GI in Arabidopsis thaliana. Plant Biotechnol. 2014, 41, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Pérez-García, P.; Pokhilko, A.; Millar, A.J.; Antoshechkin, I.; Riechmann, J.L.; Mas, P. Mapping the core of the Arabidopsis circadian clock defines the network structure of the oscillator. Science 2012, 336, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Graf, A.; Coman, D.; Uhrig, R.G.; Walsh, S.; Flis, A.; Stitt, M.; Gruissem, W. Parallel analysis of Arabidopsis circadian clock mutants reveals different scales of transcriptome and proteome regulation. Open Biol. 2017, 7, 160333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McWatters, H.G.; Kolmos, E.; Hall, A.; Doyle, M.R.; Amasino, R.M.; Gyula, P.; Nagy, F.; Millar, A.J.; Davis, S.J. ELF4 is required for oscillatory properties of the circadian clock. Plant Physiol. 2007, 144, 391–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fowler, S.; Lee, K.; Onouchi, H.; Samach, A.; Richardson, K.; Morris, B.; Coupland, G.; Putterill, J. GIGANTEA: A circadian clock-controlled gene that regulates photoperiodic flowering in Arabidopsis and encodes a protein with several possible membrane-spanning domains. EMBO J. 1999, 8, 4679–4688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, J.S.; Kubota, A.; Imaizumi, T. Circadian clock and photoperiodic flowering in Arabidopsis: CONSTANS is a hub for signal integration. Plant Physiol. 2017, 173, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Rugnone, M.L.; Faigon Soverna, A.; Sanchez, S.E.; Schlaen, R.G.; Hernando, C.E.; Seymour, D.K.; Mancini, E.; Chernomoretz, A.; Weigel, D.; Mas, P.; et al. LNK genes integrate light and clock signaling networks at the core of the Arabidopsis oscillator. Proc. Natl. Acad. Sci. USA 2013, 110, 12120–12125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrales, A.; Carrillo, L.; Lasierra, P.; Nebauer, S.G.; Dominguez-figueroa, J.; Renau-morata, B. Multifaceted role of cycling DOF factor 3 (CDF3) in the regulation of flowering time and abiotic stress responses in Arabidopsis. Plant Cell 2017, 3, 748–764. [Google Scholar] [CrossRef]
- Sawa, M.; Nusinow, D.A.; Kay, S.A.; Imaizumi, T. FKF1 and GIGANTEA complex formation is required for daylength measurement in Arabidopsis. Science 2007, 318, 261–265. [Google Scholar] [CrossRef] [Green Version]
- Marcolino-Gomes, J.; Nakayama, T.J.; Molinari, H.B.C.; Basso, M.F.; Henning, L.M.M.; Fuganti-Pagliarini, R.; Harmon, F.G.; Nepomuceno, A.L. Functional characterization of a putative Glycine max ELF4 in transgenic Arabidopsis and its role during flowering control. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Brandoli, C.; Petri, C.; Egea-Cortines, M.; Weiss, J. The clock gene Gigantea 1 from Petunia hybrida coordinates vegetative growth and inflorescence architecture. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Li, G.; Siddiqui, H.; Teng, Y.; Lin, R.; Wan, X.; Li, J.; Lau, O.-S.; Ouyang, X.; Dai, M.; Wan, J.; et al. Coordinated transcriptional regulation underlying the circadian clock in Arabidopsis. Nat. Cell Biol. 2011, 13, 616–622. [Google Scholar] [CrossRef]
- Khanna, R.; Kikis, E.A.; Quail, P.H. EARLY FLOWERING 4 functions in phytochrome B-regulated seedling de-etiolation. Plant Physiol. 2003, 133, 1530–1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liew, L.C.; Hecht, V.; Laurie, R.E.; Knowles, C.L.; Vander Schoor, J.K.; Macknight, R.C.; Weller, J.L. DIE NEUTRALIS and LATE BLOOMER 1 contribute to regulation of the pea circadian clock. Plant Cell 2009, 21, 3198–3211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bendix, C.; Marshall, C.M.; Harmon, F.G. Circadian clock genes universally control key agricultural traits. Mol. Plant 2015, 8, 1135–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, N.; Yoshida, T.; Olsen, J.E.; Senaha, C.; Jikumaru, Y.; Kamiya, Y. Short main shoot length and inhibition of floral bud development under red light can be recovered by application of gibberellin and cytokinin. Acta Hortic. 2012, 956, 215–222. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Light Quality Treatments | Photoperiod (h) | Harvest Time after Start of the Light Quality/Photoperiod Treatments | Number of Replicates Per Treatment Per Time Point ** | Total Number of Plants Harvested |
|---|---|---|---|---|---|
| a | W, R, B * | 14 | After 12 h light the 2nd day | 3 | 9 |
| b | W, R, B | 14 | 0, 4, 8, 12, 16, 20, 24, 28, 32, 36, 40, 44, 48 h | 3 | 39 |
| c | W, R, B | 14 | 0, 7, 14, 21, 28, 35 days (after 12 h light each day) | 3 | 18 |
| d | B | 8, 14, 24 | 0, 4, 8, 12, 16, 20, 24, 28, 32, 36, 40, 44, 48, 52, 60, 64, 68, 72 h | 3 | 57 *** |
| Molecular Function | Organism | Accession No. | BLAST | Read No. | ||
|---|---|---|---|---|---|---|
| Score | E-Value | Red | Blue | |||
| APETALA (AP) 2-like protein | Nicotiana tabacum | gb|ACY30435.1| | 193 | 1.00 × 10−47 | 4 | 1 |
| APETALA (AP) 2-like protein | Nicotiana tabacum | gb|ACY30435.1| | 69.7 | 1.00 × 10−10 | 0 | 3 |
| APETALA (AP) 2-like protein | Ipomoea nil | gb|ABN10954.2| | 85.9 | 6.00 × 10−15 | 2 | 3 |
| CONSTANS (CO) interacting protein 2a | Solanum lycopersicum | gb|AAS67369.1| | 79 | 2.00 × 10−13 | 8 | 8 |
| CONSTANS (CO) interacting protein 2b, putative | Capsicum chinense | dbj|BAG16521.1| | 238 | 5.00 × 10−61 | 12 | 13 |
| CONSTANS interacting protein 3 | Solanum lycopersicum | gb|AAS67371.1| | 192 | 1.00 × 10−47 | 50 | 36 |
| CONSTANS interacting protein 5 | Solanum lycopersicum | gb|AAS67373.1| | 290 | 1.00 × 10−76 | 15 | 10 |
| CONSTANS interacting protein 6 | Solanum lycopersicum | gb|AAS67374.1| | 202 | 2.00 × 10−50 | 1 | 3 |
| CONSTANS (CO) -like protein | Solanum tuberosum | gb|ABH09237.1| | 127 | 4.00 × 10−28 | 5 | 0 |
| CONSTANS (CO) -like protein | Solanum tuberosum | gb|ABH09237.1| | 202 | 2.00 × 10−50 | 26 | 15 |
| CONSTANS (CO) -like protein | Solanum tuberosum | gb|ABH09237.1| | 266 | 10−121 | 9 | 14 |
| CONSTANS (CO) -like protein | Allium cepa | gb|ACT22759.1| | 83.6 | 8.00 × 10−15 | 0 | 2 |
| Cryptochrome (CRY) 2 | Nicotiana sylvestris | gb|ABB36797.1| | 197 | 2.00 × 10−48 | 3 | 1 |
| Cryptochrome (CRY) DASH | Solanum lycopersicum | sp|Q38JU2.2| | 162 | 2.00 × 10−38 | 2 | 3 |
| Cryptochrome (CRY) 1 | Nicotiana sylvestris | gb|ABB36796.1| | 90.5 | 1.00 × 10−16 | 3 | 6 |
| Cryptochrome (CRY) 1 | Solanum lycopersicum | gb|AAD44161.1| | 67.8 | 1.00 × 10−9 | 0 | 4 |
| Early flowering (ELF) 1 protein, similar | Nasonia vitripennis | ref|XP_001601451.1 | 36.2 | 1.4 | 4 | 2 |
| Early flowering (ELF) 4 protein, putative | Solanum lycopersicum | gb|AAW22881.1| | 135 | 2.00 × 10−30 | 6 | 0 |
| Early flowering (ELF) 4 protein, putative | Solanum lycopersicum | gb|AAW22881.1| | 173 | 2.00 × 10−41 | 16 | 7 |
| Early flowering (ELF) 4 protein, putative | Solanum lycopersicum | gb|AAW22881.1| | 147 | 7.00 × 10−34 | 1 | 14 |
| Early light inducible protein (ELIP) | Solanum lycopersicum | gb|AAS92268.1| | 85.5 | 3.00 × 10−15 | 4 | 0 |
| Early light inducible protein (ELIP) | Solanum lycopersicum | gb|AAS92268.1| | 99 | 2.00 × 10−19 | 1 | 1 |
| Early Tobacco Anther1 | Nicotiana tabacum | gb|AAO43000.1| | 36.2 | 6.00 × 10−5 | 2 | 4 |
| Early Tobacco Anther1 | Nicotiana tabacum | gb|AAO43000.1| | 133 | 2.00 × 10−29 | 5 | 10 |
| Embryonic flower (EMF) 2 | Solanum lycopersicum | gb|ABI99480.1| | 195 | 2.00 × 10−48 | 5 | 5 |
| Floral binding protein (FBP) 13 | Petunia x hybrida | gb|AAK21250.1| | 327 | 6.00 × 10−88 | 7 | 7 |
| Floral binding protein (FBP) 22 | Petunia x hybrida | gb|AAK21253.1| | 132 | 2.00 × 10−34 | 2 | 1 |
| Floral binding protein (FBP) 28 | Petunia x hybrida | gb|AAK21257.1| | 164 | 3.00 × 10−39 | 1 | 2 |
| Flower-specific phytochrome-associated protein phosphate 3 (ATFYPP3) | Arabidopsis thaliana | ref|NP_188632.1| | 265 | 4.00 × 10−69 | 7 | 6 |
| Phytochrome A (PHYA) | Solanum tuberosum | sp|P30733.2| | 222 | 1.00 × 10−56 | 3 | 4 |
| Phytochrome A -associated F-box protein/Empfindlicher im Dunkelroten Licht 1 (EID1), putative | Ricinus communis | ref|XP_002523077.1| | 274 | 6.00 × 10−72 | 9 | 14 |
| ZEITLUPE (ZTL) | Ipomoea nil | gb|ABC25060.2| | 102 | 4.00 × 10−20 | 5 | 4 |
| Phytochrome A (PHYA) | Solanum tuberosum | sp|P30733.2| | 222 | 1.00 × 10−56 | 3 | 4 |
| Phytochrome A -associated F-box protein/Empfindlicher im Dunkelroten Licht 1 (EID1), putative | Ricinus communis | ref|XP_002523077.1| | 274 | 6.00 × 10−72 | 9 | 14 |
| ZEITLUPE (ZTL) | Ipomoea nil | gb|ABC25060.2| | 102 | 4.00 × 10−20 | 5 | 4 |
| Photoperiod (Day/Night) | Plant Height (cm) | No. of Leaves (No. Per Plant) | No. of Lateral Shoots (No. Per Plant) | No. of Days to Flowering | Percentage of Plants with Flowers (%) |
|---|---|---|---|---|---|
| 8 h/16 h | 13.0 b | 19.8 b | 0.0 b | - | 0 |
| 14 h/10 h | 16.4 b * | 25.0 b | 0.3 b | 28.2 b | 75 |
| 24 h/0 h | 26.6 a | 31.5 a | 4.0 a | 22.8 a | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fukuda, N.; Suenaga, T.; Miura, E.; Tsukamoto, A.; Olsen, J.E. The Expression of ELF4-Like Genes Is Influenced by Light Quality in Petunia. Agronomy 2020, 10, 1800. https://doi.org/10.3390/agronomy10111800
Fukuda N, Suenaga T, Miura E, Tsukamoto A, Olsen JE. The Expression of ELF4-Like Genes Is Influenced by Light Quality in Petunia. Agronomy. 2020; 10(11):1800. https://doi.org/10.3390/agronomy10111800
Chicago/Turabian StyleFukuda, Naoya, Tomohiro Suenaga, Erika Miura, Atsuko Tsukamoto, and Jorunn E. Olsen. 2020. "The Expression of ELF4-Like Genes Is Influenced by Light Quality in Petunia" Agronomy 10, no. 11: 1800. https://doi.org/10.3390/agronomy10111800
APA StyleFukuda, N., Suenaga, T., Miura, E., Tsukamoto, A., & Olsen, J. E. (2020). The Expression of ELF4-Like Genes Is Influenced by Light Quality in Petunia. Agronomy, 10(11), 1800. https://doi.org/10.3390/agronomy10111800