Leaf Response to Different Light Spectrum Compositions during Micropropagation of Gerbera Axillary Shoots



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Data Collection and Analysis
2.3. Statistical Analysis
3. Results
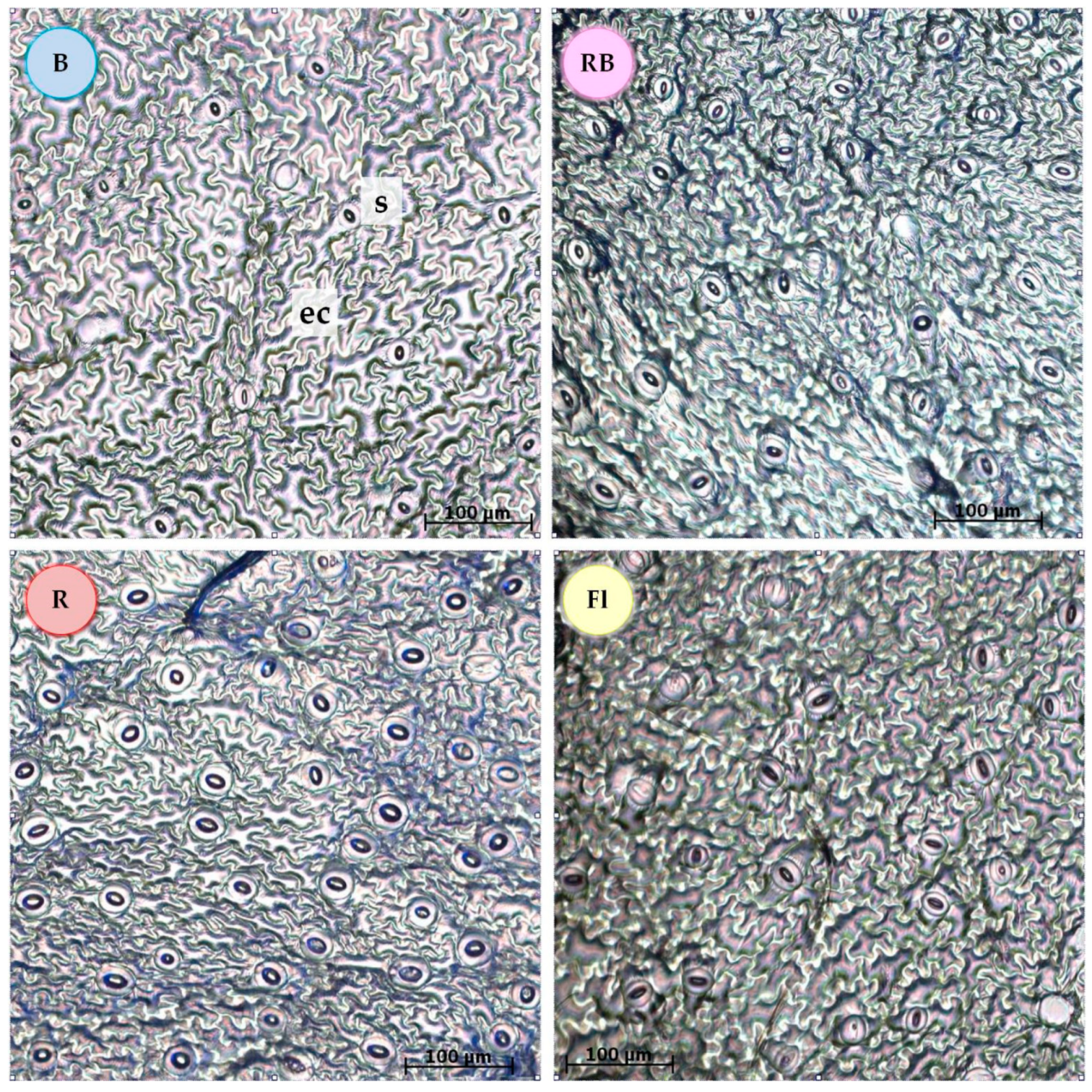
3.1. Stomata Features
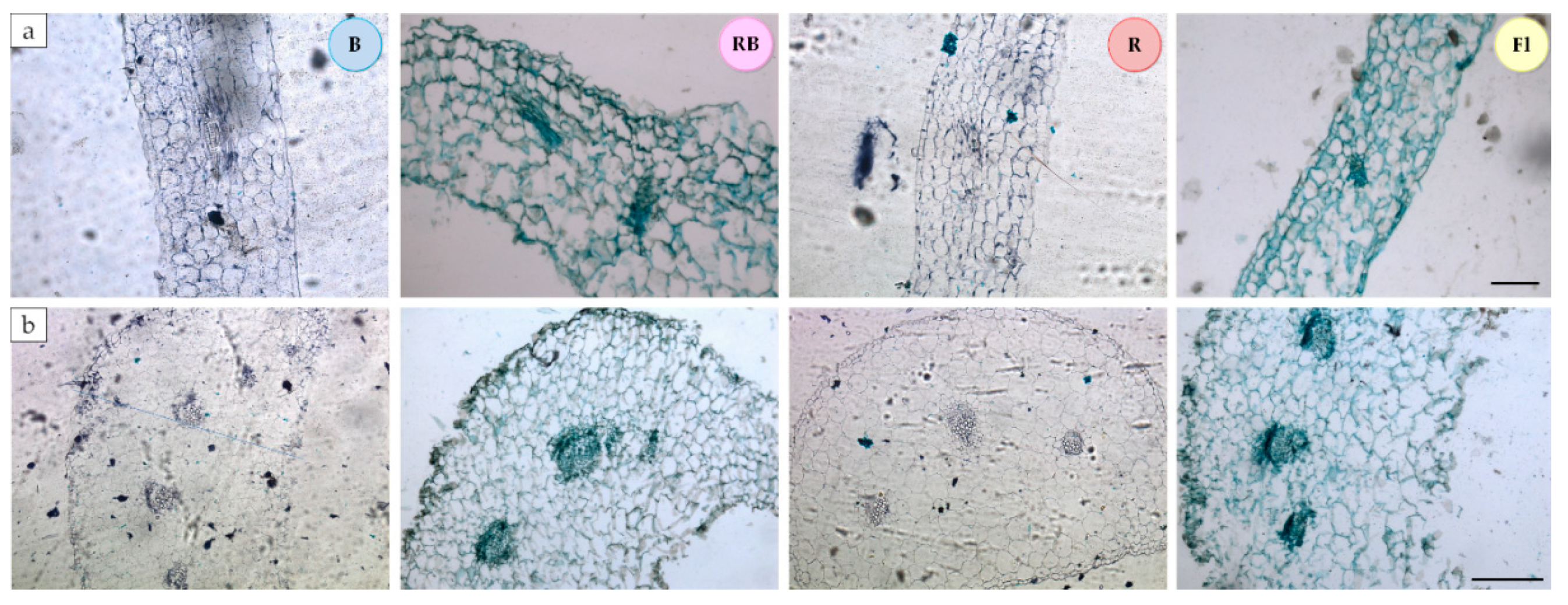
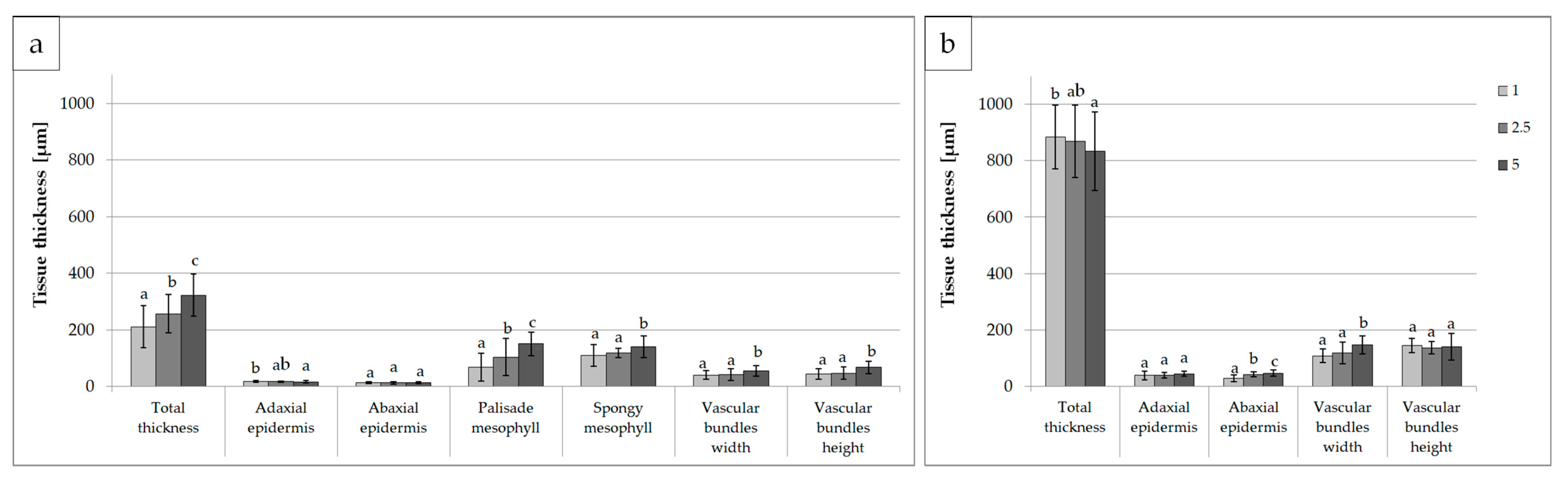
3.2. Leaf Anatomical Structure
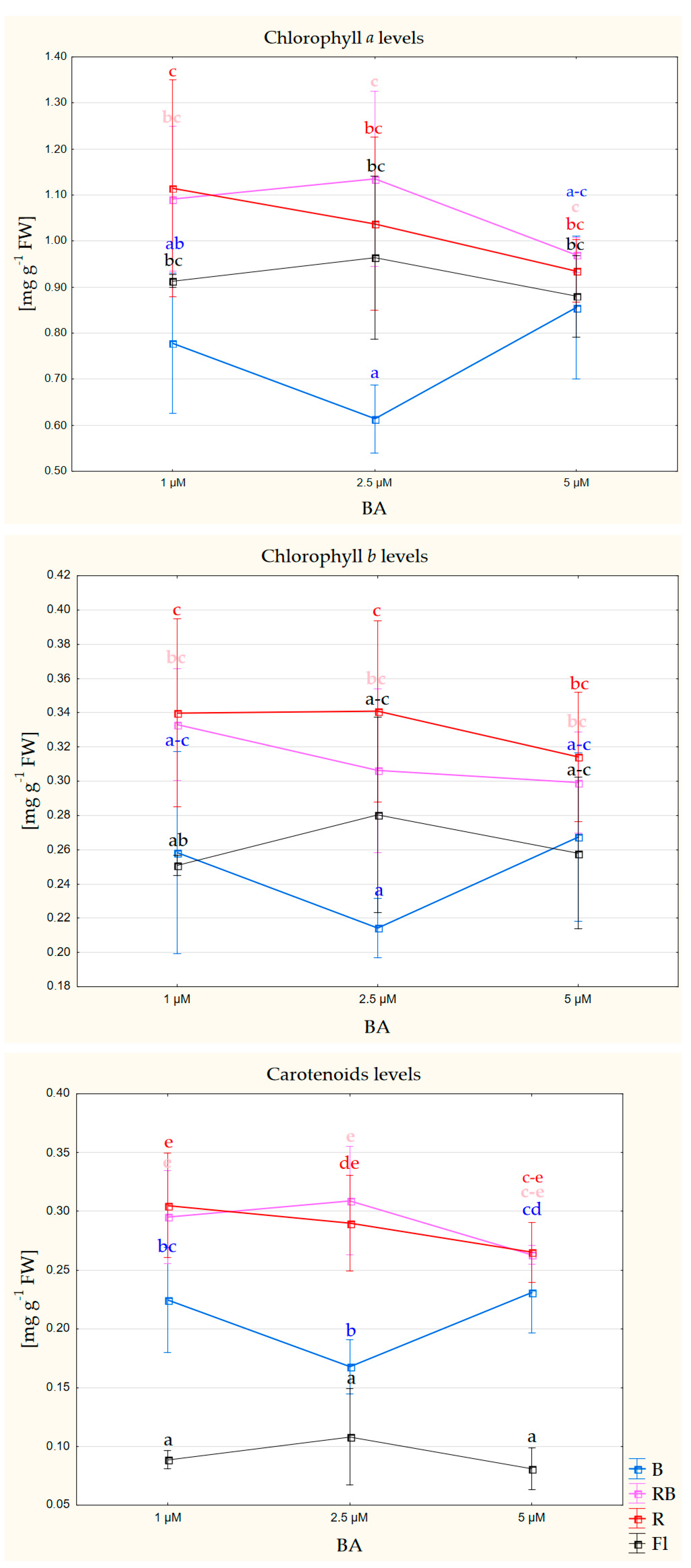
3.3. Photosynthetic Pigment Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nevo, E.; Pavlíek, T.; Beharv, A.; Bolshakova, M.A.; Martyn, G.I.; Musatenko, L.I.; Sytnikl, K.M. Drought and light anatomical adaptive leaf strategies in three woody species caused by microclimatic selection at “Evolution Canyon”, Israel. Isr. J. Plant Sci. 2000, 48, 33–46. [Google Scholar] [CrossRef]
- Johnson, D.M.; Smith, W.K.; Vogelmann, T.C.; Brodersen, C.R. Leaf architecture and direction of incident light influence mesophyll fluorescence profiles. Am. J. Bot. 2005, 92, 1425–1431. [Google Scholar] [CrossRef] [Green Version]
- Rezende, R.K.S.; Paiva, L.V.; Paiva, R.; Chalfun Júnior, A.; Torga, P.P.; Mauro de Castro, E. Capitulum organogenesis and anatomical characterization of Gerbera jamesonii Adlan leaves. Ciência e Agrotecnologia 2008, 32, 821–827. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Tholen, D.; Zhu, X.G. The influence of leaf anatomy on the internal light environment and photosynthetic electron transport rate: Exploration with a new leaf ray tracing model. J. Exp. Bot. 2016, 67, 6021–6035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeiger, E. Blue Light and stomatal function. In Blue Light Effects in Biological Systems; Springer: Berlin/Heidelberg, Germany, 1984; pp. 484–494. ISBN 978-3-642-69767-8. [Google Scholar]
- Hetherington, A.M.; Woodward, F.I. The role of stomata in sensing and driving environmental change. Nature 2003, 424, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Inoue, S.I.; Kinoshita, T. Blue light regulation of stomatal opening and the plasma membrane H+-ATPase. Plant Physiol. 2017, 174, 531–538. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lu, W.; Tong, Y.; Yang, Q. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (Lactuca sativa L.) exposed to different ratios of red light to blue light. Front. Plant Sci. 2016, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hronková, M.; Wiesnerová, D.; Šimková, M.; Skůpa, P.; Dewitte, W.; Vráblová, M.; Zažímalová, E.; Šantrůček, J. Light-induced STOMAGEN-mediated stomatal development in Arabidopsis leaves. J. Exp. Bot. 2015, 66, 4621–4630. [Google Scholar] [CrossRef] [Green Version]
- Stefanova, M.A.; Koleva, D.P.; Ganeva, T.G.; Dimitrova, M.A. Effect of plant growth regulators on the regeneration of in vitro-propagated Lamium album L. plants. J. Pharm. 2011, 4, 1982–1985. [Google Scholar]
- Pawłowska, B.; Żupnik, M.; Szewczyk-Taranek, B.; Cioć, M. Impact of LED light sources on morphogenesis and levels of photosynthetic pigments in Gerbera jamesonii grown in vitro. Hortic. Environ. Biotechnol. 2018, 59, 115–123. [Google Scholar] [CrossRef]
- Pawłowska, B.; Cioć, M.; Prokopiuk, B. How LED light rooting in vitro affected Gerbera acclimatization efficiency. Acta Hortic. 2018, 583–590. [Google Scholar] [CrossRef]
- Cioć, M.; Kalisz, A.; Żupnik, M.; Pawłowska, B. Different LED light intensities and 6-benzyladenine concentrations in relation to shoot development, leaf architecture, and photosynthetic pigments of Gerbera jamesonii Bolus in vitro. Agronomy 2019, 9, 358. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tabacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Li, H.; Xu, Z.; Tang, C. Effect of light-emitting diodes on growth and morphogenesis of upland cotton (Gossypium hirsutum L.) plantlets in vitro. Plant Cell Tissue Organ Cult. 2010, 103, 155–163. [Google Scholar] [CrossRef]
- Filutowicz, A.; Kużdowicz, A. Mikrotechnika Roślinna; PWRiL: Warszawa, Poland, 1951. [Google Scholar]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 171–178. [Google Scholar] [CrossRef]
- Sæbø, A.; Krekling, T.; Appelgren, M. Light quality affects photosynthesis and leaf anatomy of birch plantlets in vitro. Plant Cell Tissue Organ Cult. 1995, 41, 177–185. [Google Scholar] [CrossRef]
- Bello-Bello, J.J.; Martínez-Estrada, E.; Caamal-Velázquez, J.H.; Morales-Ramos, V. Effect of LED light quality on in vitro shoot proliferation and growth of vanilla (Vanilla planifolia Andrews). Afr. J. Biotechnol. 2016, 15, 272–277. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Mosqueda, M.A.; Iglesias-Andreu, L.G.; Bautista-Aguilar, J.R. The effect of light quality on growth and development of in vitro plantlet of Stevia rebaudiana Bertoni. Sugar Tech 2017, 19, 331–336. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, K.; Gong, X.; Wang, H.; Gao, Y.; Wang, X.; Zeng, Z.; Hu, Y. Effects of different LEDs light spectrum on the growth, leaf anatomy, and chloroplast ultrastructure of potato plantlets in vitro and minituber production after transplanting in the greenhouse. J. Integr. Agric. 2020, 19, 108–119. [Google Scholar] [CrossRef]
- Zobayed, S.M.A.; Afreen-Zobayed, F.; Kubota, C.; Kozai, T. Stomatal characteristics and leaf anatomy of potato plantlets cultured in vitro under photoautotrophic and photomixotrophic conditions. Vitr. Cell. Dev. Biol. Plant 1999, 35, 183–188. [Google Scholar] [CrossRef]
- Tichá, I.; Radochová, B.; Kadleček, P. Stomatal morphology during acclimatization of tobacco plantlets to ex vitro conditions. Biol. Plant. 1999, 42, 469–474. [Google Scholar] [CrossRef]
- Camargo, M.A.B.; Marenco, R.A. Density, size and distribution of stomata in 35 rainforest tree species in Central Amazonia. Acta Amaz. 2011, 41, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Hahn, E.J.; Heo, J.W.; Paek, K.Y. Effects of LEDs on net photosynthetic rate, growth and leaf stomata of chrysanthemum plantlets in vitro. Sci. Hortic. (Amst.) 2004, 101, 143–151. [Google Scholar] [CrossRef]
- Gupta, S.; Jatothu, B. Fundamentals and applications of light-emitting diodes (LEDs) in in vitro plant growth and morphogenesis. Plant Biotechnol. Rep. 2013, 7, 211–220. [Google Scholar] [CrossRef]
- Macedo, A.F.; Leal-Costa, M.V.; Tavares, E.S.; Lage, C.L.S.; Esquibel, M.A. The effect of light quality on leaf production and development of in vitro-cultured plants of Alternanthera brasiliana Kuntze. Environ. Exp. Bot. 2011, 70, 43–50. [Google Scholar] [CrossRef]
- Heo, J.; Lee, C.; Chakrabarty, D.; Paek, K. Growth responses of marigold and salvia bedding plants as affected by monochromic or mixture radiation provided by a light-emitting diode (LED). Plant Growth Regul. 2002, 38, 225–230. [Google Scholar] [CrossRef]
- Jordan, G.J.; Carpenter, R.J.; Koutoulis, A.; Price, A.; Brodribb, T.J. Environmental adaptation in stomatal size independent of the effects of genome size. New Phytol. 2015, 205, 608–617. [Google Scholar] [CrossRef] [Green Version]
- Aranda, I.; Pardo, F.; Gil, L.; Pardos, J.A. Anatomical basis of the change in leaf mass per area and nitrogen investment with relative irradiance within the canopy of eight temperate tree species. Acta Oecologica 2004, 25, 187–195. [Google Scholar] [CrossRef]
- Gonçalves, B.; Correia, C.M.; Silva, A.P.; Bacelar, E.A.; Santos, A.; Moutinho-Pereira, J.M. Leaf structure and function of sweet cherry tree (Prunus avium L.) cultivars with open and dense canopies. Sci. Hortic. (Amst.) 2008, 116, 381–387. [Google Scholar] [CrossRef]
- Fryer, M.J.; Oxborough, K.; Mullineaux, P.M.; Baker, N.R. Imaging of photo-oxidative stress responses in leaves. J. Exp. Bot. 2002, 53, 1249–1254. [Google Scholar] [CrossRef] [Green Version]
- Fryer, M.J.; Ball, L.; Oxborough, K.; Karpinski, S.; Mullineaux, P.M.; Baker, N.R. Control of Ascorbate Peroxidase 2 expression by hydrogen peroxide and leaf water status during excess light stress reveals a functional organisation of Arabidopsis leaves. Plant J. 2003, 33, 691–705. [Google Scholar] [CrossRef] [PubMed]
- Schuerger, A.C.; Brown, C.S.; Stryjewski, E.C. Anatomical features of pepper plants (Capsicum annuum L.) grown under red light-emitting diodes supplemented with blue or far-red light. Ann. Bot. 1997, 79, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Thompson, W.A.; Kriedemann, P.E.; Craig, I.E. Photosynthetic response to light and nutrients in sun-tolerant and shade-tolerant rainforest trees. I. Growth, leaf anatomy and nutrient content. Aust. J. Plant Physiol. 1992, 19, 1–18. [Google Scholar] [CrossRef]
- Strauss-Debenedetti, S.; Berlyn, G.P. Leaf anatomical responses to light in five tropical Moraceae of different successional status. Am. J. Bot. 1994, 81, 1582–1591. [Google Scholar] [CrossRef]
- Terashima, I.; Miyazawa, S.I.; Hanba, Y.T. Why are sun leaves thicker than shade leaves?—Consideration based on analyses of CO2 diffusion in the leaf. J. Plant Res. 2001, 114, 93–105. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, S.; Hu, H.; Li, D. Photosynthetic performance along a light gradient as related to leaf characteristics of a naturally occurring Cypripedium flavum. J. Plant Res. 2008, 121, 559–569. [Google Scholar] [CrossRef]
- Kasperbauer, M.J.; Peaslee, D.E. Morphology and photosynthetic efficiency of tobacco leaves that received end-of-day red and far red light during development. Plant Physiol. 1973, 52, 440–442. [Google Scholar] [CrossRef] [Green Version]
- Boardman, N.K. Comparative photosynthesis of sun and shade plants. Annu. Rev. Plant Physiol. 1977, 28, 355–377. [Google Scholar] [CrossRef]
- Barreiro, R.; Guiamét, J.J.; Beltrano, J.; Montaldi, E.R. Regulation of the photosynthetic capacity of primary bean leaves by the red: Far-red ratio and photosynthetic photon flux density of incident light. Physiol. Plant. 1992, 85, 97–101. [Google Scholar] [CrossRef]
- Pushnik, J.C.; Miller, G.W.; Jolley, V.D.; Brown, J.C.; Davis, T.D.; Barnes, A.M. Influences of ultra-violet (UV)-blue light radiation on the growth of cotton. II. Photosynthesis, leaf anatomy, and iron reduction. J. Plant Nutr. 1987, 10, 2283–2297. [Google Scholar] [CrossRef]
- Wellman, E. UV radiation in photomorphogenisis. In Encyclopedia of Plant Physiology: New Series. Photomorphogenesis; Shropshire, W., Mohr, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1983. [Google Scholar]
- Gonzalez, R.M.; Efsted, R.; Wellburn, A.R.; Paul, N.D. Non-photosynthetic mechanisms of growth reduction in pea (Pisum sativum L.) exposed to UV-B radiation. Plant Cell Environ. 1998, 21, 23–32. [Google Scholar] [CrossRef]
- Tevini, M.; Thoma, U.; Iwanzik, W. Effects of UV-B radiation on germination, seedling growth, leaf anatomy, and pigments of some crop plants. Zeitschrift für Pflanzenphysiologie 1983, 109, 435–448. [Google Scholar] [CrossRef]
- Fukuda, N.; Fujita, M.; Ohta, Y.; Sase, S.; Nishimura, S.; Ezura, H. Directional blue light irradiation triggers epidermal cell elongation of abaxial side resulting in inhibition of leaf epinasty in geranium under red light condition. Sci. Hortic. (Amst.) 2008, 115, 176–182. [Google Scholar] [CrossRef]
- Zervoudakis, G.; Salahas, G.; Kaspiris, G.; Konstantopoulou, E. Influence of light intensity on growth and physiological characteristics of common sage (Salvia officinalis L.). Braz. Arch. Biol. Technol. 2012, 55, 89–95. [Google Scholar] [CrossRef]
- Wang, Z.; Li, G.; He, S.; Teixeira da Silva, J.A.; Tanaka, M. Effect of cold cathode fluorescent lamps on growth of Gerbera jamesonii plantlets in vitro. Sci. Hortic. (Amst.) 2011, 130, 482–484. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Main Effects: | Light Quality × BA | Light Quality | BA |
|---|---|---|---|
| Stomata features | |||
| Stomata frequency | *** 1 | *** | n.s. |
| Stomata area | *** | *** | n.s. |
| Stomata length | *** | n.s. | n.s. |
| Stomata width | *** | *** | ** |
| Pore length | *** | n.s. | ** |
| Pore width | *** | *** | ** |
| Anatomical structure of blade | |||
| Total thickness | *** | *** | *** |
| Adaxial epidermis | *** | *** | ** |
| Abaxial epidermis | n.s. | *** | n.s. |
| Palisade mesophyll | *** | *** | *** |
| Spongy mesophyll | *** | *** | *** |
| Vascular bundles width | ** | *** | *** |
| Vascular bundles height | n.s. | *** | *** |
| Anatomical structure of petiole | |||
| Total thickness | *** | *** | ** |
| Adaxial epidermis | *** | *** | n.s. |
| Abaxial epidermis | *** | *** | *** |
| Vascular tissues width | *** | n.s. | *** |
| Vascular tissues height | *** | *** | n.s. |
| Photosynthetic pigments level | |||
| Chlorophyll a | n.s. | *** | n.s. |
| Chlorophyll b | n.s. | *** | n.s. |
| Carotenoids | n.s. | *** | n.s. |
| Tested Feature | Stomata | Pore | ||||
|---|---|---|---|---|---|---|
| Mean Number | Area (µm2) | Length (µm) | Width (µm) | Length (µm) | Width (µm) | |
| Light quality | ||||||
| B 1 | 22.00 ± 4.64 a 2 | 556.27 ± 114.83 bc | 29.25 ± 3.14 ab | 24.35 ± 3.16 b | 17.08 ± 2.46 a | 9.20 ± 1.84 b |
| RB | 29.33 ± 4.12 b | 513.31 ± 129.55 ab | 28.36 ± 3.86 ab | 23.39 ± 3.43 ab | 15.92 ± 1.90 a | 8.83 ± 2.38 b |
| R | 31.33 ± 9.41 b | 588.58 ± 121.20 c | 30.10 ± 3.95 b | 24.95 ± 2.60 b | 17.21 ± 3.27 a | 10.50 ± 2.54 c |
| Fl 1 | 30.44 ± 5.83 b 2 | 474.57 ± 56.24 a | 27.88 ± 1.77 a | 21.92 ± 1.84 a | 16.02 ± 1.84 a | 7.39 ± 1.48 a |
| BA concentrations (µM) | ||||||
| 1 | 27.67 ± 5.25 a | 563.09 ± 135.72 a | 29.19 ± 3.53 a | 24.78 ± 3.53 b | 17.40 ± 2.26 b | 9.62 ± 2.20 b |
| 2.5 | 27.17 ± 6.06 a | 511.21 ± 76.70 a | 28.59 ± 2.41 a | 23.06 ± 2.42 a | 15.60 ± 1.79 a | 8.13 ± 1.50 a |
| 5 | 30.00 ± 9.62 a | 525.25 ± 123.92 a | 28.92 ± 4.00 a | 23.11 ± 2.72 a | 16.68 ± 2.94 ab | 9.19 ± 2.94 b |
| Tested Features: | r ≥ | −1 | −0.8 | −0.6 | −0.4 | −0.2 | 0 | 0.2 | 0.4 | 0.6 | 0.8 | 1 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chlorophyll a | Chlorophyll b | Carotenoids | ||||||||||
| Photosynthetic pigments | ||||||||||||
| chlorophyll a | 1.000000 | 0.888045 | 0.588219 | |||||||||
| chlorophyll b | 0.888045 | 1.000000 | 0.711361 | |||||||||
| carotenoids | 0.588219 | 0.711361 | 1.000000 | |||||||||
| Stomata features | ||||||||||||
| stomata width | −0.047408 | 0.145536 | 0.268465 | |||||||||
| pore width | 0.142891 | 0.304001 | 0.416299 | |||||||||
| Leaf anatomical structure | ||||||||||||
| blade spongy mesophyll | −0.314132 | −0.243908 | −0.007865 | |||||||||
| blade vascular tissue hight | −0.061806 | 0.026109 | 0.264615 | |||||||||
| petiole adaxial epidermis | −0.299653 | −0.325324 | −0.324174 | |||||||||
| petiole abaxial epidermis | −0.227974 | −0.323510 | −0.473888 | |||||||||
| petiole vascular bundles height | −0.039401 | −0.121789 | −0.423516 | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cioć, M.; Pawłowska, B. Leaf Response to Different Light Spectrum Compositions during Micropropagation of Gerbera Axillary Shoots. Agronomy 2020, 10, 1832. https://doi.org/10.3390/agronomy10111832
Cioć M, Pawłowska B. Leaf Response to Different Light Spectrum Compositions during Micropropagation of Gerbera Axillary Shoots. Agronomy. 2020; 10(11):1832. https://doi.org/10.3390/agronomy10111832
Chicago/Turabian StyleCioć, Monika, and Bożena Pawłowska. 2020. "Leaf Response to Different Light Spectrum Compositions during Micropropagation of Gerbera Axillary Shoots" Agronomy 10, no. 11: 1832. https://doi.org/10.3390/agronomy10111832
APA StyleCioć, M., & Pawłowska, B. (2020). Leaf Response to Different Light Spectrum Compositions during Micropropagation of Gerbera Axillary Shoots. Agronomy, 10(11), 1832. https://doi.org/10.3390/agronomy10111832