Legume Ecotypes and Commercial Cultivars Differ in Performance and Potential Suitability for Use as Permanent Living Mulch in Mediterranean Vegetable Systems




Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Site
2.2. Experiment 1: Evaluation of Commercial Cultivars of Perennial and Annual Self-Seeding Legumes
2.3. Experiment 2: Evaluation of Medicago polymorpha Ecotypes
2.4. Statistical Analysis
3. Results
3.1. Evaluation of Commercial Cultivars of Annual SELF-Seeding and Perennial Legumes
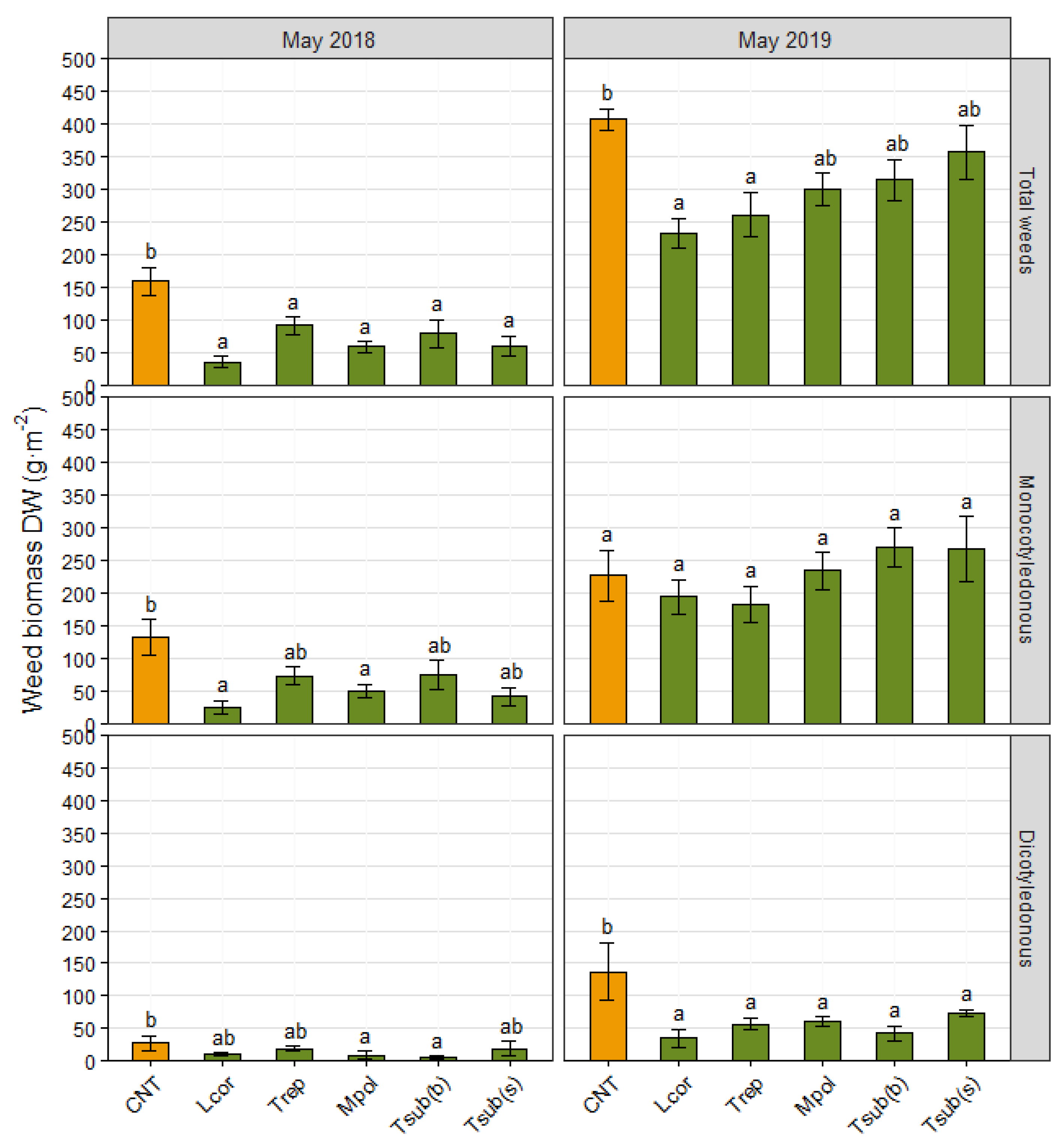
3.1.1. Weed Biomass
3.1.2. Legume Biomass
3.1.3. Canopy Ground Cover
3.1.4. Canopy Height
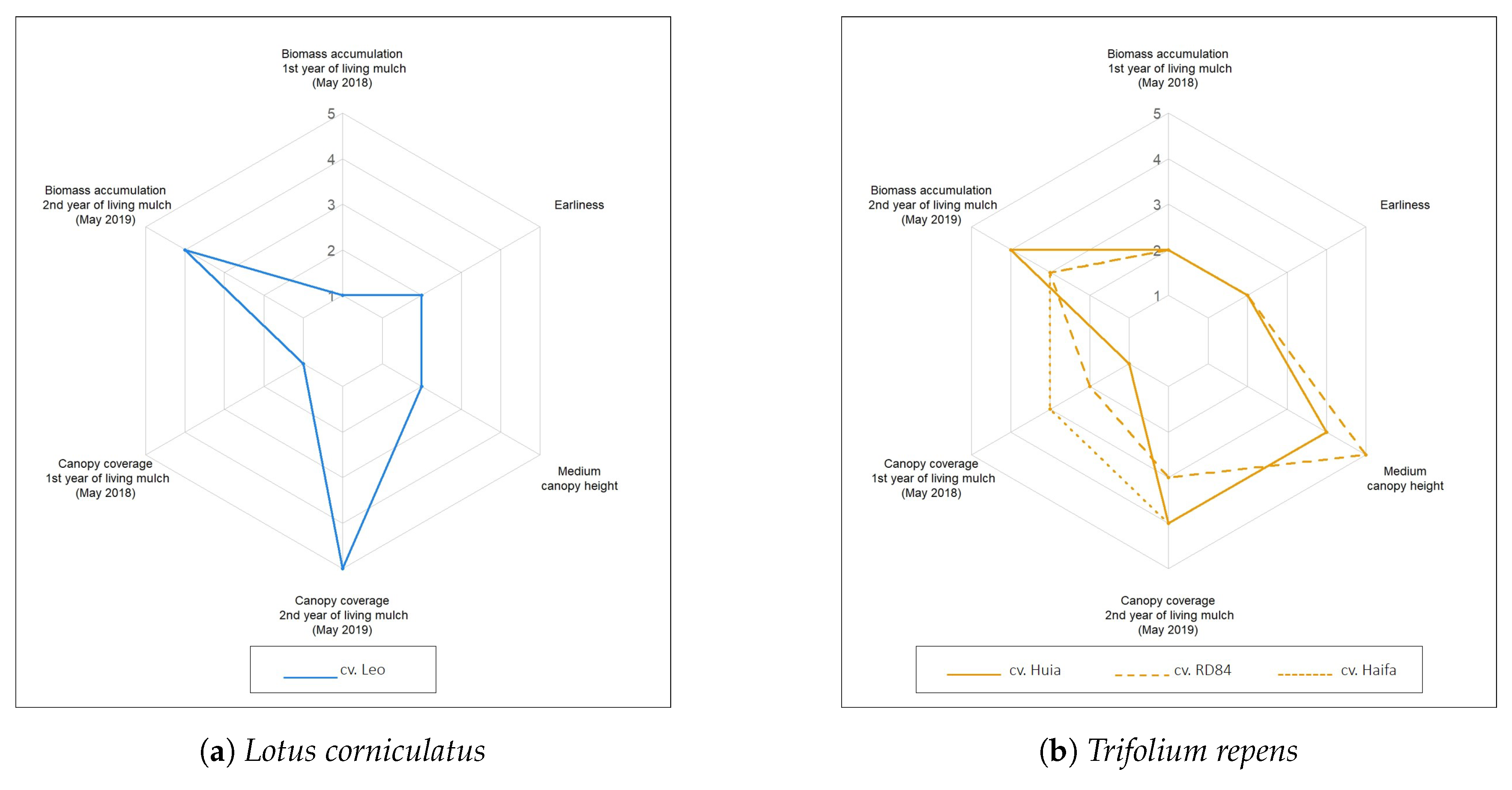
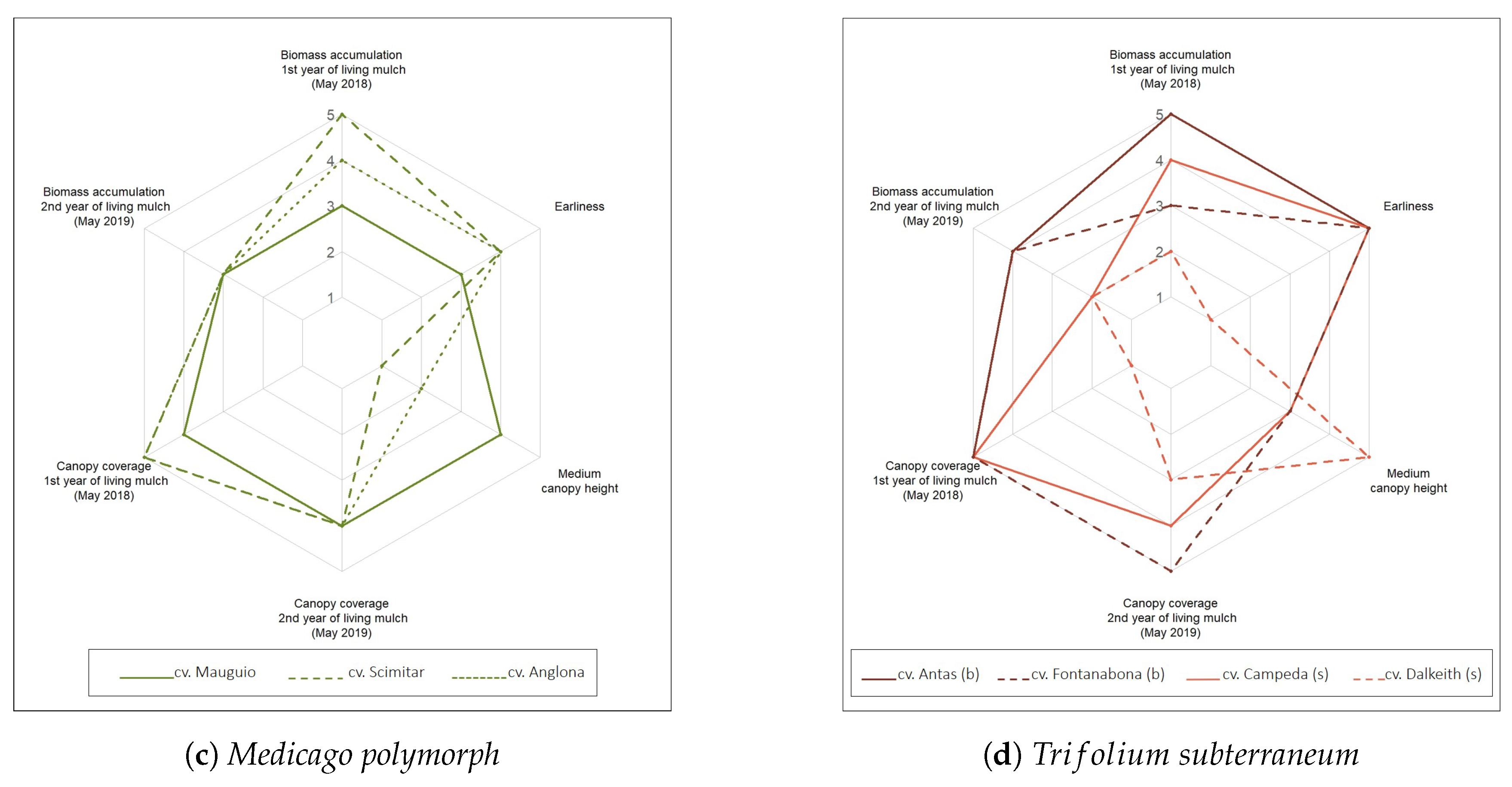
3.1.5. Legume Growth Traits as Indicators for Their Potential Use as Permanent Living Mulch
3.2. Evaluation of Medicago Polymorph Ecotypes
3.2.1. Weed Biomass
3.2.2. Legume Biomass
3.2.3. Self-Seeding Capacity (SSC)
4. Discussion
5. Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Hartwig, N.L. Crownvetch—A perennial legume “living mulch” for no-tillage crop production. Proc. Northeast. Weed Sci. Soc. 1983, 37, 28–38. [Google Scholar]
- Hartwig, N.L.; Ammon, H.U. Cover crops and living mulches. Weed Sci. 2002, 50, 688–699. [Google Scholar] [CrossRef]
- Bosco, S.; Volpi, I.; Antichi, D.; Ragaglini, G.; Frasconi, C. Greenhouse Gas Emissions from Soil Cultivated with Vegetables in Crop Rotation under Integrated, Organic and Organic Conservation Management in a Mediterranean Environment. Agronomy 2019, 9, 446. [Google Scholar] [CrossRef] [Green Version]
- Antichi, D.; Sbrana, M.; Martelloni, L.; Abou Chehade, L.; Fontanelli, M.; Raffaelli, M.; Mazzoncini, M.; Peruzzi, A.; Frasconi, C. Agronomic Performances of Organic Field Vegetables Managed with Conservation Agriculture Techniques: A Study from Central Italy. Agronomy 2019, 9, 810. [Google Scholar] [CrossRef] [Green Version]
- Feil, B.; Liedgens, M. Crop production in living mulches—A review. Pflanzenbauwissenschaften 2001, 5, 15–23. [Google Scholar]
- Muller-Scharer, H.; Potter, C. Cover plants in field grown vegetables: Prospects and limitations. In Proceedings of the International Conference Organised by the British Crop Protection Council Held at Brighton Centre and Brighton Metropole, Brighton, UK, 18–21 November 1991; pp. 599–604. [Google Scholar]
- Wezel, A.; Casagrande, M.; Celette, F.; jean françois, V.; Ferrer, A.; Peigné, J. Agroecological practices for sustainable agriculture. A review. Agron. Sustain. Dev. 2014, 34, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, G. Living Mulch as a Tool to Control Weeds in Agroecosystems: A Review. In Weed Control; Price, A., Ed.; InTech: London, UK, 2012. [Google Scholar] [CrossRef] [Green Version]
- Teasdale, J.R. Interaction of Light, Soil Moisture, and Temperature with Weed Suppression by Hairy Vetch Residue. Weed Sci. 1993, 41, 46–51. [Google Scholar] [CrossRef]
- Carlsen, S.C.K.; Pedersen, H.A.; Spliid, N.H.; Fomsgaard, I.S. Fate in Soil of Flavonoids Released from White Clover (Trifolium repens L.). Appl. Environ. Soil Sci. 2012, 2012, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ko, E. Living mulch in vegetable crop production: Perspectives and limitations (a review). Acta Sci. Pol.-Hortorum Cultus 2013, 12, 127–142. [Google Scholar]
- Masiunas, J.B. Production of Vegetables Using Cover Crop and Living Mulches. A Review. J. Veg. Crop. Prod. 1998, 11–31. [Google Scholar] [CrossRef]
- Bond, W.; Grundy, A.C. Non-chemical weed management in organic farming systems. Weed Res. 2001, 41, 383–405. [Google Scholar] [CrossRef]
- Kruidhof, H.M.; Bastiaans, L.; Kropff, M.J. Ecological weed management by cover cropping: Effects on weed growth in autumn and weed establishment in spring. Weed Res. 2008, 48, 492–502. [Google Scholar] [CrossRef]
- Vincent-Caboud, L.; Peigné, J.; Casagrande, M.; Silva, E. Overview of Organic Cover Crop-Based No-Tillage Technique in Europe: Farmers’ Practices and Research Challenges. Agriculture 2017, 7, 42. [Google Scholar] [CrossRef] [Green Version]
- Ciaccia, C.; Kristensen, H.L.; Campanelli, G.; Xie, Y.; Testani, E.; Leteo, F.; Canali, S. Living mulch for weed management in organic vegetable cropping systems under Mediterranean and North European conditions. Renew. Agric. Food Syst. 2017, 32, 248–262. [Google Scholar] [CrossRef]
- Kołodziejczyk, M. The effect of living mulches and conventional methods of weed control on weed infestation and potato yield. Sci. Hortic. 2015, 191, 127–133. [Google Scholar] [CrossRef]
- Walters, S.A.; Young, B.G. Utility of winter rye living mulch for weed management in zucchini squash production. Weed Technol. 2008, 22, 724–728. [Google Scholar] [CrossRef]
- Gibson, K.D.; Mcmillan, J.; Hallett, S.G.; Jordan, T.; Weller, S.C. Effect of a Living Mulch on Weed Seed Banks in Tomato. Weed Technol. 2011, 25, 245–251. [Google Scholar] [CrossRef]
- Den Hollander, N.; Bastiaans, L.; Kropff, M. Clover as a cover crop for weed suppression in an intercropping design. Eur. J. Agron. 2007, 26, 92–103. [Google Scholar] [CrossRef]
- Brandsæter, L.O.; Netland, J.; Meadow, R. Yields, Weeds, Pests and Soil Nitrogen in a White Cabbage-Living Mulch System. Biol. Agric. Hortic. 1998, 16, 291–309. [Google Scholar] [CrossRef]
- Chase, C.; Mbuya, O. Greater Interference from Living Mulches than Weeds in Organic Broccoli Production. Weed Technol. 2008, 22, 280–285. [Google Scholar] [CrossRef]
- Ilnicki, R.D.; Enache, A.J. Subterranean clover living mulch: An alternative method of weed control. Agric. Ecosyst. Environ. 1992, 40, 249–264. [Google Scholar] [CrossRef]
- Brandsaeter, L.O.; Netland, J. Winter Annual Legumes for Use as Cover Crops in Row Crops in Northern Regions: I. Field Experiments. Crop Sci. 1999, 39, 1369–1379. [Google Scholar] [CrossRef]
- Ross, S.M.; King, J.R.; Izaurralde, R.; O’Donovan, J.T. Weed Suppression by Seven Clover Species. Agron. J. 2001, 93, 820–827. [Google Scholar] [CrossRef]
- Russi, L.; Lorenzetti, S. Screening germplasm of Medicago polymorpha suitable as a cover crop in Mediterranean orchards. In Lucerne and Medics for the XXI Century, Proceedings of the XIII Eucarpia Medicago spp. Group Meeting, Perugia, Italy, 13–16 September 1999; Universita di Perugia: Perugia, Italy, 2000; pp. 170–175. [Google Scholar]
- Ocumpaugh, W.; Hussey, M.; Grichar, J.; Read, J.; Bade, D.; Pinchak, W.; Smith, G.; Pitman, W.; Muir, J.; Coleman, S. Registration of ‘Armadillo’ Burr Medic. Crop Sci. 2004, 44. [Google Scholar] [CrossRef]
- Prosperi, J.; Ronfort, J.; Genier, G. Constraints to the introduction of medics in French Mediterranean farming systems. In Lucerne and Medics for the XXI Century, Proceedings of the XIII Eucarpia Medicago spp. Group Meeting, Perugia, Italy, 13–16 September 1999; Universita di Perugia: Perugia, Italy, 2000; pp. 154–161. [Google Scholar]
- Piano, E.; Pecetti, L.; Carroni, A.M. Campeda-Limbara-Losa and Antas. The first Italian varieties of subterranean clover [Sardinia]. In Sementi Elette; Istituto Sperimentale per le Colture Foraggere: Lodi, Italy, 1997. [Google Scholar]
- Porqueddu, C.; Loi, A.; Fara, G. Anglona: The first Italian variety of annual medic. In Proceedings of the IV Congresso Nazionale Biodiversità: Germoplasma Locale e SUA Valorizzazione, Alghero, Italy, 8–11 September 1998; pp. 8–11. [Google Scholar]
- Teasdale, J.R.; Beste, C.E.; Potts, W.E. Response of weeds to tillage and cover crop residue. Weed Sci. 1991, 39, 195–199. [Google Scholar] [CrossRef]
- Ross, M.; Lembi, C. Methods of weed control. In Applied Weed Science; MacMillan: New York, NY, USA, 1985; pp. 20–45. [Google Scholar]
- Steinmaus, S.; Elmore, C.L.; Smith, R.J.; Donaldson, D.; Weber, E.A.; Roncoroni, J.A.; Miller, P.R.M. Mulched cover crops as an alternative to conventional weed management systems in vineyards. Weed Res. 2008, 48, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Sheaffer, C.C.; Gunsolus, J.L.; Grimsbo Jewett, J.; Lee, S.H. Annual Medicago as a Smother Crop in Soybean. J. Agron. Crop. Sci. 2002, 188, 408–416. [Google Scholar] [CrossRef]
- Jahn, R.; Blume, H.; Asio, V.; Spaargaren, O.; Schad, P. Guidelines for Soil Description, 4th ed.; FAO: Rome, Italy, 2006. [Google Scholar]
- Standard Operating Procedure for Soil Organic Carbon. Walkley-Black Method: Titration and Colorimetric Method. 2019. Available online: http://www.fao.org/3/ca7471en/ca7471en.pdf (accessed on 22 November 2020).
- Bremner, J.M. Determination of nitrogen in soil by the Kjeldahl method. J. Agric. Sci. 1960, 55, 11–33. [Google Scholar] [CrossRef]
- Pierzynski, G.M. Methods of Phosphorus Analysis for Soils, Sediments, Residuals, and Waters; North Carolina State University: Raleigh, NC, USA, 2000. [Google Scholar]
- Meier, U. Growth Stages of MONO- and dicotyledonous Plants: BBCH Monograph; Open Agrar Repositorium: Quedlinburg, Germany, 2018. [Google Scholar] [CrossRef]
- De Haan, R.; Wyse, D.; Ehlke, N.; Maxwell, B.; Putnam, D. Simulation of Spring-Seeded Smother Plants for Weed Control in Corn (Zea mays). Weed Sci. 1994, 42, 35–43. [Google Scholar] [CrossRef]
- RStudio, Inc. RStudio: Integrated; Development for RBoston: Boston, MA, USA, 2015. [Google Scholar]
- Bates, D.; Machler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using {lme4}. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. emmeans: Estimated Marginal Means, aka Least-Squares Means 1.4.6. 2020. Available online: https://cran.r-project.org/web/packages/emmeans/index.html (accessed on 22 November 2020).
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. 2020. Available online: https://cran.r-project.org/web/packages/DHARMa/index.html (accessed on 22 November 2020).
- Bettina, G.; Kosmidis, I.; Zeileis, A. Extended Beta Regression in {R}: Shaken, Stirred, Mixed, and Partitioned. J. Stat. Softw. 2011, 48, 1–25. [Google Scholar]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-Response Analysis Using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakazawa, M. Functions for Medical Statistics Book with some Demographic Data. 2019. Available online: https://cran.r-project.org/web/packages/fmsb/index.html (accessed on 22 November 2020).
- Bàrberi, P.; Bigongiali, F.; Antichi, D.; Carlesi, S.; Fontanelli, M.; Fiasconi, C.; Lulli, L. Innovative crop and weed management strategies for organic spinach: Crop yield and weed suppression. In Proceedings of the Second Scientific Conference of the International Society of Organic Agriculture Research (ISOFAR), Modena, Italy, 18–20 June 2008; Volume 1, pp. 252–255. [Google Scholar]
- Teasdale, J.R.; Daughtry, C.S.T. Weed Suppression by Live and Desiccated Hairy Vetch (Vicia villosa). Weed Sci. 1993, 41, 207–212. [Google Scholar] [CrossRef]
- Reddy, K.N.; Koger, C.H. Live and killed hairy vetch cover crop effects on weeds and yield in glyphosate-resistant corn. Weed Technol. 2004, 18, 835–840. [Google Scholar] [CrossRef]
- Hiltbrunner, J.; Jeanneret, P.; Liedgens, M.; Stamp, P.; Streit, B. Response of Weed Communities to Legume Living Mulches in Winter Wheat. J. Agron. Crop. Sci. 2007, 193, 93–102. [Google Scholar] [CrossRef]
- Brainard, D.C.; Bakker, J.; Noyes, D.C.; Myers, N. Rye Living Mulch Effects on Soil Moisture and Weeds in Asparagus. HortScience 2012, 47, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Nichols, P.; Loi, A.; Nutt, B.; Snowball, R.; Revell, C. Domestication of New Mediterranean Annual Pasture Legumes. In Sustainable use of Genetic Diversity in Forage and Turf Breeding; Huyghe, C., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 137–141. [Google Scholar] [CrossRef]
- Mariotti, A.; Dell’Aquila, A. Decadal climate variability in the Mediterranean region: Roles of large-scale forcings and regional processes. Clim. Dyn. 2012, 38, 1129–1145. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Genus and Species | Common Name | Cultivar | Cycle | Seeding Rate |
|---|---|---|---|---|---|
| Lcor | Lotus corniculatus L. | Bird’s-foot trefoil | Leo | P | 30 |
| Trep1 | Trifolium repens L. | White clover | Huia | P | 25 |
| Trep2 | Trifolium repens L. | White clover | Haifa | P | 25 |
| Trep3 | Trifolium repens L. | White clover | RD 84 | P | 25 |
| Mpol1 | Medicago polymorpha L. | Bur clover | Scimitar | ASS | 25 |
| Mpol2 | Medicago polymorpha L. | Bur clover | Anglona | ASS | 25 |
| Mpol3 | Medicago polymorpha L. | Bur clover | Mauguio | ASS | 25 |
| Tsub(s)1 | Trifolium subterraneum L.* | Subterranean clover | Dalkeith | ASS | 35 |
| Tsub(s)2 | Trifolium subterraneum L.* | Subterranean clover | Campeda | ASS | 35 |
| Tsub(b)1 | Trifolium subterraneum L.** | Subterranean clover | Antas | ASS | 35 |
| Tsub(b)2 | Trifolium subterraneum L.** | Subterranean clover | Fontanabona | ASS | 35 |
| CNT | Spontaneous vegetation | - | Control | - | - |
| Code | Genus Species | Cultivar * |
|---|---|---|
| Commercial | ||
| Mpol1 | M. polymorpha L. | Scimitar |
| Mpol2 | M. polymorpha L. | Anglona |
| Mpol3 | M. polymorpha L. | Mauguio |
| Ecotypes | ||
| Mpol4 | M. polymorpha L. | Pitigliano (Siena, SI) |
| Mpol5 | M. polymorpha L. | Manciano (Grosseto, GR) |
| Mpol6 | M. polymorpha L. | Talamone (Grosseto, GR) |
| Mpol7 | M. polymorpha L. | Principina (Grosseto, GR) |
| Mpol8 | M. polymorpha L. | Villa Salto (Sud Sardegna, SU) |
| Mpol9 | M. polymorpha L. | San Felice Circeo (Latina, LT) |
| Mpol10 | M. polymorpha L. | Tarquinia (Viterbo, VT) |
| CNT | Spontaneous vegetation | Control |
| Legume Species | Cultivar | DW | |||||
|---|---|---|---|---|---|---|---|
| May 2018 | May 2019 | M18–M19 | |||||
| L. corniculatus L. | Leo | 41.61± 8.15 | a | 241.24 ± 39.45 | ab | +199.63±49.4 | *** |
| T. repens L. | Haifa | 58.03 ± 13.88 | a | 161.05 ± 26.87 | ab | +103.02±31.1 | *** |
| T. repens L. | RD 84 | 82.50 ± 16.11 | a | 174.73 ± 29.66 | ab | +92.23±33.4 | ** |
| T. repens L. | Huia | 57.73 ± 11.33 | a | 227.80 ± 39.36 | ab | +220.07±39.4 | *** |
| M. polymorpha L. | Mauguio | 180.16 ± 31.20 | ab | 176.52 ± 29.97 | ab | −3.64±42.7 | n.s. |
| M. polymorpha L. | Anglona | 455.85 ± 64.59 | d | 137.02 ± 23.32 | ab | −318.83±63.5 | *** |
| M. polymorpha L. | Scimitar | 275.76 ± 43.33 | bc | 129.30 ± 21.93 | a | −146.46±46.6 | ** |
| T. subterraneum L. | Dalkeith * | 74.67 ± 25.06 | ab | 88.99 ± 15.15 | a | +14.32 ± 31.6 | n.s. |
| T. subterraneum L. | Campeda * | 260.24 ± 46.48 | bc | 102.58 ± 17.54 | a | −157.66 ± 44.2 | *** |
| T. subterraneum L. | Fontanabona ** | 198.55 ± 30.31 | bc | 288.38 ± 38.59 | b | +89.83 ± 56.1 | *** |
| T. subterraneum L. | Antas ** | 335.83 ± 53.36 | cd | 216.42 ± 36.69 | ab | −119.41 ± 59.0 | * |
| Legume Species | Cultivar | 2018 | 2019 | ||||
|---|---|---|---|---|---|---|---|
| C50 | Cmax | Cmax | |||||
| L. corniculatus L. | Leo | 18.00 ± 6.02 | a | 81.61 ± 6.07 | bc | ||
| T. repens L. | Haifa | 27.32 ± 6.76 | a | 59.38 ± 8.90 | abc | ||
| T. repens L. | RD 84 | 39.38 ± 7.67 | ab | 78.99 ± 6.39 | abc | ||
| T. repens L. | Huia | 20.84 ± 6.37 | a | 87.29 ± 5.21 | c | ||
| M. polymorpha L. | Mauguio | 134.76 ± 1.06 | b | 64.80 ± 7.53 | bc | 62.49 ± 7.60 | abc |
| M. polymorpha L. | Anglona | 129.23 ± 0.55 | ab | 90.29 ± 4.62 | d | 62.67 ± 7.59 | abc |
| M. polymorpha L. | Scimitar | 129.64 ± 0.58 | ab | 84.02 ± 5.74 | cd | 57.49 ± 7.76 | ab |
| T. subterraneum L. | Dalkeith * | 18.36 ± 6.07 | a | 51.25 ± 7.85 | a | ||
| T. subterraneum L. | Campeda * | 127.89 ± 0.82 | a | 95.31 ± 3.25 | d | 63.66 ± 7.55 | abc |
| T. subterraneum L. | Fontanabona ** | 127.64 ± 0.87 | a | 94.64 ± 3.48 | d | 83,02 ± 5.88 | bc |
| T. subterraneum L. | Antas ** | 127.20 ± 0.72 | a | 95.63 ± 3.13 | d | 84.70 ± 5.21 | c |
| Legume Species | Cultivar | Canopy Height | |||
|---|---|---|---|---|---|
| May 2018 | May 2019 | ||||
| L. corniculatus L. | Leo | 20.69 ± 2.42 | b | 26.18 ± 2.69 | c |
| T. repens L. | Haifa | 9.09 ± 1.83 | a | 13.88 ± 2.01 | ab |
| T. repens L. | RD 84 | 10.83 ± 2.04 | a | 17.43 ± 2.27 | ab |
| T. repens L. | Huia | 12.03 ± 2.08 | a | 17.74 ± 2.29 | b |
| M. polymorpha L. | Mauguio | 11.77 ± 2.07 | a | 15.96 ± 2.22 | ab |
| M. polymorpha L. | Scimitar | 24.89 ± 2.62 | b | 18.09 ± 2.30 | b |
| M. polymorpha L. | Anglona | 39.16 ± 3.43 | c | 21.12 ± 2.44 | bc |
| T. subterraneum L. | Dalkeith * | 8.82 ± 1.83 | a | 9.36 ± 2.00 | a |
| T. subterraneum L. | Campeda * | 22.49 ± 2.50 | b | 16.16 ± 2.22 | ab |
| T. subterraneum L. | Fontanabona ** | 22.45 ± 2.51 | b | 17.60 ± 2.28 | ab |
| T. subterraneum L. | Antas ** | 21.06 ± 2.43 | b | 15.18 ± 2.19 | ab |
| Legume Species | Cultivar | DW | |||||
|---|---|---|---|---|---|---|---|
| May 2018 | May 2019 | M18–M19 | |||||
| Commercial | |||||||
| M. polymorpha | Scimitar | 275.76 ± 45.82 | ab | 129.30 ± 24.68 | a | −146.46 ± 43.41 | *** |
| M. polymorpha | Anglona | 455.85 ± 41.11 | b | 137.02 ± 26.78 | a | −318.83 ± 63.20 | *** |
| M. polymorpha | Mauguio | 180.16 ± 38.50 | a | 176.52 ± 28.81 | ab | −3.64 ± 38.63 | n.s. |
| Ecotype | |||||||
| M. polymorpha | Pitigliano (SI) | 333.31 ± 75.28 | ab | 204.21 ± 28.34 | ab | −129.10 ± 35.26 | * |
| M. polymorpha | Manciano (GR) | 203.97 ± 54.15 | a | 240.00 ± 27.88 | ab | +36.03 ± 48.62 | n.s. |
| M. polymorpha | Talamone (GR) | 154.59 ± 18.50 | a | 237.94 ± 39.13 | ab | +83.35 ± 40.91 | n.s. |
| M. polymorpha | Principina (GR) | 147.42 ± 45.92 | a | 264.69 ± 37.60 | ab | +117.27 ± 43.45 | ** |
| M. polymorpha | Villa Salto (SS) | 261.95 ± 34.83 | ab | 199.97 ± 26.93 | ab | −61.98 ± 44.76 | n.s. |
| M. polymorpha | San Felice Circeo (LT) | 109.78 ± 9.56 | a | 306.66 ± 46.27 | b | +196.88 ± 48.54 | *** |
| M. polymorpha | Tarquinia (VT) | 279.27 ± 60.35 | a | 159.67 ± 30.52 | a | −119.60 ± 47.12 | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leoni, F.; Lazzaro, M.; Carlesi, S.; Moonen, A.-C. Legume Ecotypes and Commercial Cultivars Differ in Performance and Potential Suitability for Use as Permanent Living Mulch in Mediterranean Vegetable Systems. Agronomy 2020, 10, 1836. https://doi.org/10.3390/agronomy10111836
Leoni F, Lazzaro M, Carlesi S, Moonen A-C. Legume Ecotypes and Commercial Cultivars Differ in Performance and Potential Suitability for Use as Permanent Living Mulch in Mediterranean Vegetable Systems. Agronomy. 2020; 10(11):1836. https://doi.org/10.3390/agronomy10111836
Chicago/Turabian StyleLeoni, Federico, Mariateresa Lazzaro, Stefano Carlesi, and Anna-Camilla Moonen. 2020. "Legume Ecotypes and Commercial Cultivars Differ in Performance and Potential Suitability for Use as Permanent Living Mulch in Mediterranean Vegetable Systems" Agronomy 10, no. 11: 1836. https://doi.org/10.3390/agronomy10111836
APA StyleLeoni, F., Lazzaro, M., Carlesi, S., & Moonen, A.-C. (2020). Legume Ecotypes and Commercial Cultivars Differ in Performance and Potential Suitability for Use as Permanent Living Mulch in Mediterranean Vegetable Systems. Agronomy, 10(11), 1836. https://doi.org/10.3390/agronomy10111836