Identifying Pollen Donors and Success Rate of Individual Pollinizers in European Plum (Prunus domestica L.) Using Microsatellite Markers

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. SSR Genotyping
3. Results and Discussion
3.1. SSR Polymorphism
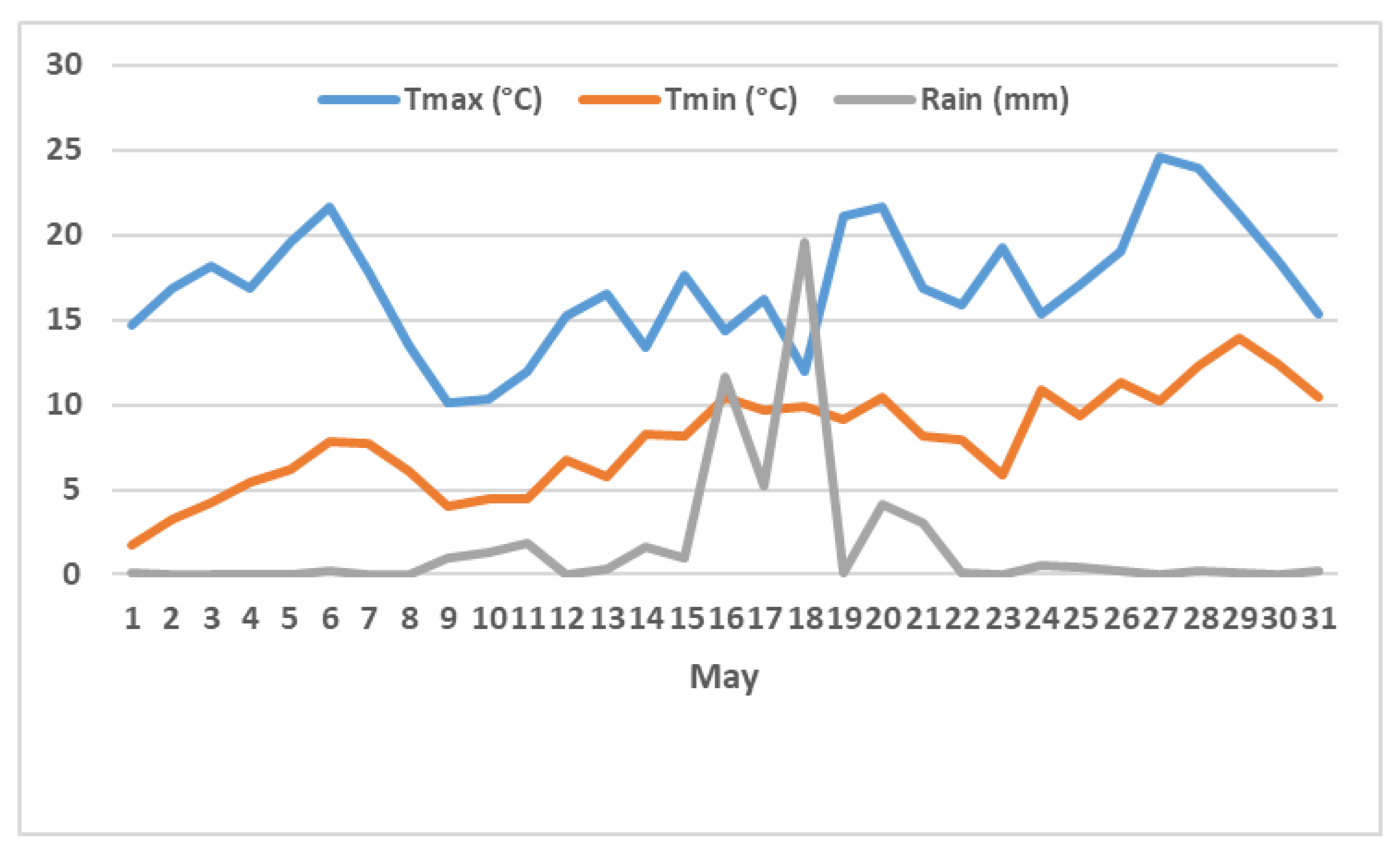
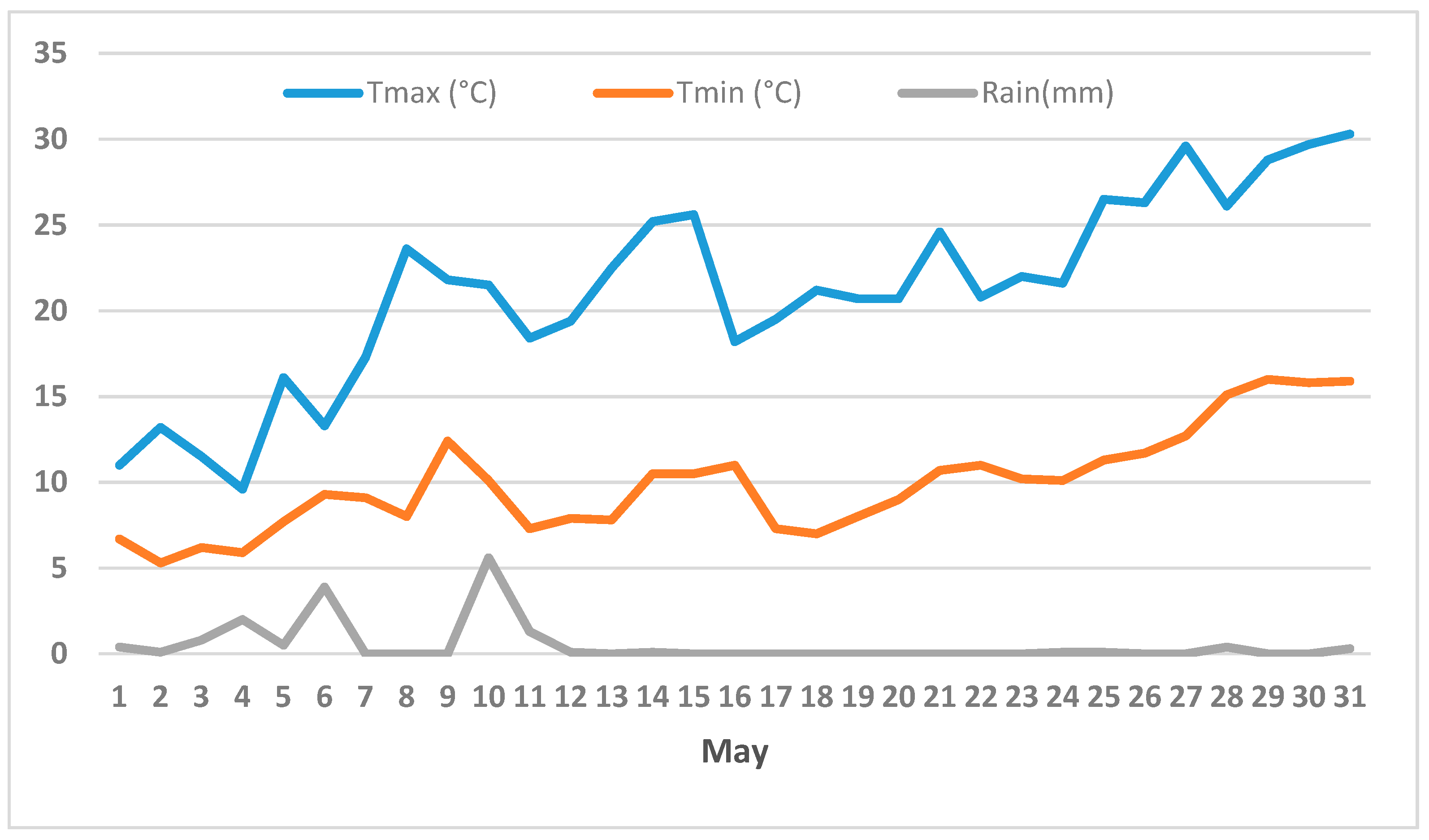
3.2. Climate and Flowering
3.3. Identifying Most Successful Pollinizers
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Koskela, E.; Kemp, H.; van Dieren, M.C.A. Flowering and pollination studies with European plum (Prunus domestica L.) cultivars. Acta Hortic. 2010, 874, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Nikolić, D.; Milatović, D. Examining self-compatibility in plum (Prunus domestica L.) by fluorescence microscopy. Genetika 2010, 42, 387–396. [Google Scholar] [CrossRef]
- Surányi, D. Comparative study of different fertile groups in plums. Int. J. Hortic. Sci. 2006, 12, 71–76. [Google Scholar] [CrossRef]
- Sharafi, Y.; Ghanbari, A.; Naji, A.M.; Kordenaeej, A.; Rezaei, A. Comparison of pollen traits of some plum cultivars of Iran. Int. J. Agron. Plant Prod. 2013, 4, 314–318. [Google Scholar]
- Nikolić, D.; Rakonjac, V.; Fotirić-Akšić, M. The effect of pollinizer on the fruitset of plum cultivar Čačanska Najbolja. J. Agric. Sci. 2012, 57, 9–18. [Google Scholar]
- Nyéki, J.; Szabó, Z. Fruit set of plum cultivars under Hungarian ecological conditions. Acta Hortic. 1996, 423, 185–192. [Google Scholar] [CrossRef]
- Kozma, P.; Nyeki, J.; Soltesz, M.; Szabo, Z. Floral Biology, Pollination and Fertilisation in Temperate Zone Fruit Species and Grape; Akadémiai Kiado: Budapest, Hungary, 2003; pp. 383–410. [Google Scholar]
- Neumüller, M. Fundamental and applied aspects of plum (Prunus domestica L.) breeding. In Methods in Temperate Fruit Breeding. Fruit, Vegetable and Cereal Science and Biotechnology; Flachowsky, H., Hanke, V.M., Eds.; GSB: Kagawa, Japan, 2010; Volume 5, pp. 139–154. [Google Scholar]
- Sutherland, B.G.; Cerović, R.; Robbins, T.P.; Tobutt, K.R. The myrobalan (Prunus cerasifera L.): A useful diploid model for studying the molecular genetics of self-incompatibility in plums. Euphytica 2009, 166, 385–398. [Google Scholar] [CrossRef]
- Wertheim, S.J.; Schmidt, H. Flowering, pollination and fruit set. In Fundamentals of Temperate Zone Tree Fruit Production; Tromp, J., Webster, A.D., Wertheim, S.J., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2005; pp. 1–400. [Google Scholar]
- Meland, M. Efficacy of Chemical Bloom Thinning Agents to European Plums. Acta Agric. Scand. 2007, 57, 235–242. [Google Scholar] [CrossRef]
- Meland, M.; Maas, F.M. Regulation of fruiting in plum production. In Proceedings of the 6th Conference Innovation in Fruit Growing, Belgrade, Serbia, 14–17 November 2017; Faculty of Agriculture: Peradeniya, Srilan, 2017; pp. 51–67. [Google Scholar]
- Thompson, M.M.; Liu, L.J. Temperature, fruit set and embryo sac development in Italian prune. J. Am. Soc. Hortic. Sci. 1973, 98, 193–197. [Google Scholar]
- Williams, R.R. Factors affecting pollination in fruit trees. In Physiology of Tree Crops; Luckwill, L.C., Cutting, C.V., Eds.; Academic Press: London, UK, 1970; pp. 193–207. [Google Scholar]
- Tubiello, F.N.; Soussana, J.F.; Howden, S.M. Crop and pasture response to climate change. Proc. Natl. Acad. Sci. USA 2007, 104, 19686–19690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beppu, K.; Suehara, T.; Kataoka, I. Embryo sac development and fruit set of Satohnishiki sweet cherry as affected by temperature, GA3 and paclobutrazol. J. Jpn. Soc. Hortic. Sci. 2001, 70, 157–162. [Google Scholar] [CrossRef]
- Sanzol, J.; Herrero, M. The effective pollination period in fruit trees. Sci. Hortic. 2001, 90, 1–17. [Google Scholar] [CrossRef]
- Ðorđević, M.; Cerovic, R.; Radičević, S.; Nikolić, D.; Milošević, N.; Glišić, I.; Marić, S.; Lukić, M. Pollen tube growth and embryo sac development in ‘PoznaPlava’ plum cultivar related to fruit set. Erwerbsobsbau 2019, 61, 313–322. [Google Scholar]
- Horvath, A.; Balsemin, E.; Barbot, J.C.; Christmann, H.; Manzano, G.; Reynet, P.; Laigret, F.; Mariette, S. Phenotypic variability and genetic structure in plum (Prunus domestica L.), cherry plum (P. cerasifera Ehrh.) and sloe (P. spinosa L.). Sci. Hortic. 2011, 129, 283–293. [Google Scholar] [CrossRef]
- Xuan, H.; Ding, Y.; Spann, D.; Möller, O.; Büchele, M.; Neumüller, M. Microsatellite markers (SSR) as a tool to assist in identification of European plum (Prunus domestica L.). Acta Hortic. 2011, 918, 689–692. [Google Scholar] [CrossRef]
- Halapija Kazija, D.; Jelacic, T.; Vujevic, P.; Milinovic, B.; Cicek, D.; Bisko, A.; Pejic, I.; Simon, S.; Zulj Mihaljevic, M.; Pecina, M.; et al. Plum germplasm in Croatia and neighboring countries assessed by microsatellites and DUS descriptors. Tree Genet. Genom. 2014, 10, 761–778. [Google Scholar] [CrossRef]
- Sehic, J.; Nybom, H.; Hjeltnes, S.H.; Gaši, F. Genetic diversity and structure of Nordic plum germplasm preserved ex situ and on-farm. Sci. Hortic. 2015, 160, 195–202. [Google Scholar] [CrossRef]
- Merkouropoulos, G.; Ganopoulos, I.; Tsaftaris, A.; Papadopoulos, I.; Drogoudi, P. Combination of high resolution melting (HRM) analysis and SSR molecular markers speeds up plum genotyping: Case study genotyping the Greek plum GeneBank collection. Plant Genet. Resour. 2017, 15, 366–375. [Google Scholar] [CrossRef] [Green Version]
- Makovics-Zsohár, N.; Tóth, M.; Surányi, D.; Kovács, S.; Hegedűs, A.; Halász, J. Simple sequence repeat markers reveal hungarian plum (Prunus domestica L.) germplasm as a valuable gene resource. HortScience 2017, 52, 1655–1660. [Google Scholar] [CrossRef]
- Urrestarazu, J.; Errea, P.; Miranda, C.; Santesteban, L.G.; Pina, A. Genetic diversity of Spanish Prunus domestica L. germplasm reveals a complex genetic structure underlying. PLoS ONE 2018, 13, e0195591. [Google Scholar] [CrossRef] [Green Version]
- Pop, R.; Harta, M.; Szabo, K.; Zânescu, M.; Sisea, C.R.; Catana, C.; Pamfil, D. Genetic diversity and population structure of plum accessions from a Romanian germplasm collection assessed by simple sequence repeat (SSR) markers. Not. Bot. Horti Agrobot. Cluj Napoca 2018, 46, 90–96. [Google Scholar] [CrossRef]
- Decroocq, V.; Hagen, L.S.; Favé, M.-G.; Eyquard, J.-P.; Pierronnet, A. Microsatellite markers in the hexaploid Prunus domestica species and parentage lineage of three European plum cultivars using nuclear and chloroplast simple-sequence repeats. Mol. Breed. 2004, 13, 135–142. [Google Scholar]
- Gasi, F.; Pojskić, N.; Kurtovic, M.; Kaiser, C.; Hjeltnes, S.H.; Fotiric-Aksic, M.; Meland, M. Pollinizer efficacy of several ‘Ingeborg’ pear pollinizers in Hardanger, Norway, examined using microsatellite markers. HortScience 2017, 52, 1722–1727. [Google Scholar] [CrossRef]
- Nishio, S.; Takada, N.; Terakami, S.; Kato, H.; Inoue, H.; Takeuchi, Y.; Saito, T. Estimation of effective pollen dispersal distance for cross-pollination in chestnut orchards by microsatellite-based paternity analyses. Sci. Hortic. 2019, 250, 89–93. [Google Scholar] [CrossRef]
- Scorza, R.; Kriss, A.B.; Callahan, A.M.; Webb, K.; Demuth, M.; Gottwald., T. Spatial and temporal assessment of pollen- and seed- mediated gene flow from genetically engineered plum Prunus domestica. PLoS ONE 2013, 8, e75291. [Google Scholar] [CrossRef]
- Dirlewanger, E.; Cosson, P.; Tavud, M.; Aranzana, M.J.; Poizat, C.; Zanetto, A.; Arús, P.; Laigret, F. Development of microsatellite markers in peach [Prunus persica (L.) Batsch] and their use in genetic diversity analysis in peach and sweet cherry (Prunus avium L.). Theor. Appl. Genet. 2002, 105, 127–138. [Google Scholar] [CrossRef]
- Decroocq, V.; Favé, M.G.; Hagen, L.; Bordenave, L.; Decroocq, S. Development and transferability of apricot and grape EST microsatellite markers across taxa. Theor. Appl. Genet. 2003, 106, 912–922. [Google Scholar] [CrossRef]
- Cipriani, G.; Lot, G.; Huang, W.G.; Marrazzo, M.T.; Peterlunger, E.; Testolin, R. AC/GT and AG/CT microsatellite repeats in peach [Prunus persica (L) Batsch]: Isolation, characterization and cross-species amplification in Prunus. Theor. Appl. Genet. 1999, 99, 65–72. [Google Scholar] [CrossRef]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1978; pp. 1–512. [Google Scholar]
- Hardy, O.J.; Vekemans, X. A versatile computer program to analyse spatial genetic structure at the individual or population level. Mol. Ecol. Notes 2002, 2, 618–620. [Google Scholar] [CrossRef] [Green Version]
- Meirmans, P.G.; Van Tienderen, P.H. Genotype and Genodive: Two programs for the analysis of genetic diversity of asexual organisms. Mol. Ecol. Notes 2004, 4, 792–794. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Ogašanović, D. The most suitable pollinators for new plum cultivars. J. Yugosl. Pomol. 1985, 19, 109–115. [Google Scholar]
- Biddinger, D.J.; Joshi, N.K.; Rajotte, E.G.; Halbrendt, N.O.; Pulig, C.; Naithani, K.J.; Vaughan, M. An immuno marking method to determine the foraging patterns of Osmia cornifrons and resulting fruit set in a cherry orchard. Apidologie 2013, 44, 738–749. [Google Scholar] [CrossRef] [Green Version]
- Hagler, J.R.; Mueller, S.; Teuber, L.R.; Machtley, S.A.; Van Deynze, A. Foraging range of honey bees, Apis mellifera, in alfalfa seed production fields. J. Insect Sci. 2011, 11, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, S.; Moritz, R.F.A. Foraging distance in Bombus terrestris L. (Hymenoptera: Apidae). Apidologie 2008, 39, 419–427. [Google Scholar] [CrossRef]
- Zurbuchen, A.; Landert, L.; Klaiber, J.; Müller, A.; Hein, S.; Dorn, S. Maximum foraging ranges in solitary bees: Only a few individuals have the capability to cover long foraging distances. Biol. Conserv. 2009, 143, 669–676. [Google Scholar] [CrossRef]
- Lankinen, Å.; Smith, H.G.; Andersson, S.; Madjidian, J.A. Selection on pollen and pistil traits during pollen competition is affected by both sexual confl ict and mixed mating in a self-compatible herb. Am. J. Bot. 2016, 103, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Rodrigo, J.; Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Sci. Hortic. 2002, 92, 125–135. [Google Scholar] [CrossRef] [Green Version]
- Meland, M.; Frøynes, O.; Coop, L.; Kaiser, C. Modeling of sweet cherry flowering based on temperature and phenology in a mesic Nordic climate. Acta Hortic. 2017, 1162, 19–22. [Google Scholar] [CrossRef]
- Kemp, H.; Wertheim, S.J. Bestuiving. In 19e Rassenlijst Voor Groot Fruitgewassen; CPRO-DLO: Wageningen, The Netherlands, 1999; pp. 1–238. (In Dutch) [Google Scholar]
- Keulemans, J. The effect of temperature on pollen tube growth and fruit set on plum trees. Acta Hortic. 1984, 149, 95–101. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cultivars | ‘Edda’ | ‘Opal’ | ‘Jubileum’ | ‘Reeves’ | ‘Mallard’ | ‘Avalon’ | ‘Cacanska Lepotica’ | ‘Valor’ | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Orchards/year | ‘17 | ‘18 | ‘17 | ‘18 | ‘17 | ‘18 | ‘17 | ‘18 | ‘17 | ‘18 | ‘17 | ‘18 | ‘17 | ‘18 | ‘17 | ‘18 |
| Orchard 1 | x | x | ||||||||||||||
| Orchard 2 | x | |||||||||||||||
| Orchard 3 | x | x | ||||||||||||||
| Orchard 4 | x | x | x | x | x | x | x | |||||||||
| Orchard 5 | x | x | x | x | x | x | x | x | x | x | x | x | x | x | ||
| Orchard 6 | x | |||||||||||||||
| Orchard 7 | x | |||||||||||||||
| Orchard 8 | x | x | ||||||||||||||
| Orchard 9 | x | x | ||||||||||||||
| Pollinizer | Year | Orchard 1 | Orchard 2 | Orchard 3 | Orchard 4 | Orchard 5 | Orchard 6 | Orchard 7 | Orchard 8 | Orchard 9 |
|---|---|---|---|---|---|---|---|---|---|---|
| ‘Edda’ | ‘17 | 10 | 50 | |||||||
| ‘18 | 10 | 20 | 50 | |||||||
| ‘Opal’ | ‘17 | 70 | 50 | 10 | 20 | 10 | 30 | |||
| ‘18 | 70 | 50 | 50 | 10 | 20 | 10 | 30 | |||
| ‘Jubileum’ | ‘17 | 15 | 9 | 5 | ||||||
| ‘18 | 20 | 20 | 9 | |||||||
| ‘Reeves’ | ‘17 | 30 | 20 | 30 | ||||||
| ‘18 | 50 | 20 | 20 | 30 | ||||||
| ‘Mallard’ | ‘17 | 25 | 21 | 30 | ||||||
| ‘18 | 38 | 21 | 20 | 30 | ||||||
| Avalon | ‘17 | 34 | 9 | 5 | ||||||
| ‘18 | 9 | 40 | ||||||||
| ‘Valor’ | ‘17 | 10 | 30 | 5 | ||||||
| ‘18 | 10 | 5 | ||||||||
| ‘Cacanska Lepotica’ | ‘17 | 1 | 11 | 50 | ||||||
| ‘18 | 2 | 11 | 50 | |||||||
| ‘Herman’ | ‘17 | 5 | ||||||||
| ‘18 | ||||||||||
| ‘Victoria’ | ‘17 | 5 | 50 | |||||||
| ‘18 | 10 | 5 | 50 | |||||||
| ‘Excalibur’ | ‘17 | 50 | ||||||||
| ‘18 | 10 |
| Main cv. and poll. | ‘Edda’ kernels | ‘Opal’ kernels | ‘Jubileum’ kernels | ‘Reeves’ kernels | ‘Mallard’ kernels | ‘Avalon’ kernels | ‘Cacanska Lepotica’ kernels | ‘Valor’ kernels | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Na | Na ‘17 | Na ‘18 | Na ‘17 | Na ‘18 | Na ‘17 | Na ‘18 | Na ‘17 | Na ‘18 | Na ‘17 | Na ‘18 | Na ‘17 | Na ‘18 | Na ‘17 | Na ‘18 | Na ‘17 | Na ‘18 | |
| BPPCT034 | 14 | 8 | 8 | 9 | 10 | 9 | 10 | 9 | 9 | 11 | 11 | 11 | 11 | 7 | 10 | 13 | 9 |
| BPPCT039 | 13 | 9 | 8 | 8 | 6 | 9 | 8 | 7 | 10 | 10 | 8 | 8 | 8 | 4 | 6 | 9 | 8 |
| BPPCT040 | 8 | 7 | 6 | 7 | 8 | 8 | 7 | 7 | 8 | 8 | 8 | 8 | 8 | 5 | 7 | 8 | 8 |
| PacA33 | 13 | 6 | 5 | 5 | 8 | 6 | 8 | 8 | 8 | 5 | 8 | 7 | 8 | 4 | 7 | 9 | 9 |
| BPPCT014 | 13 | 8 | 6 | 8 | 10 | 9 | 11 | 11 | 12 | 12 | 10 | 9 | 10 | 7 | 7 | 10 | 9 |
| UDP96 | 11 | 7 | 8 | 8 | 9 | 8 | 10 | 9 | 10 | 9 | 9 | 9 | 8 | 3 | 8 | 7 | 5 |
| UDP98 | 12 | 8 | 7 | 5 | 9 | 7 | 10 | 8 | 7 | 6 | 8 | 8 | 8 | 5 | 6 | 7 | 6 |
| Mean | 12.00 | 7.57 | 6.86 | 7.14 | 8.57 | 8.00 | 9.14 | 8.43 | 9.14 | 8.71 | 8.86 | 8.57 | 8.71 | 5.00 | 7.29 | 9.00 | 7.71 |
| He | He ‘17 | He ‘18 | He ‘17 | He ‘18 | He ‘17 | He ‘18 | He ‘17 | He ‘18 | He ‘17 | He ‘18 | He ‘17 | He ‘18 | He ‘17 | He ‘18 | He ‘17 | He ‘18 | |
| BPPCT034 | 0.91 | 0.85 | 0.85 | 0.71 | 0.76 | 0.81 | 0.83 | 0.83 | 0.82 | 0.86 | 0.84 | 0.82 | 0.84 | 0.84 | 0.83 | 0.88 | 0.85 |
| BPPCT039 | 0.90 | 0.87 | 0.87 | 0.84 | 0.82 | 0.85 | 0.81 | 0.80 | 0.83 | 0.82 | 0.83 | 0.80 | 0.82 | 0.75 | 0.76 | 0.85 | 0.85 |
| BPPCT040 | 0.88 | 0.82 | 0.82 | 0.77 | 0.81 | 0.83 | 0.83 | 0.85 | 0.87 | 0.87 | 0.87 | 0.86 | 0.86 | 0.80 | 0.82 | 0.85 | 0.87 |
| PacA33 | 0.88 | 0.74 | 0.72 | 0.68 | 0.73 | 0.76 | 0.82 | 0.79 | 0.81 | 0.78 | 0.82 | 0.75 | 0.82 | 0.68 | 0.70 | 0.84 | 0.86 |
| BPPCT014 | 0.90 | 0.83 | 0.80 | 0.81 | 0.83 | 0.86 | 0.85 | 0.88 | 0.88 | 0.85 | 0.85 | 0.86 | 0.86 | 0.69 | 0.72 | 0.87 | 0.87 |
| UDP96 | 0.85 | 0.83 | 0.84 | 0.78 | 0.84 | 0.86 | 0.83 | 0.84 | 0.83 | 0.78 | 0.84 | 0.83 | 0.83 | 0.63 | 0.69 | 0.70 | 0.61 |
| UDP98 | 0.89 | 0.82 | 0.82 | 0.56 | 0.64 | 0.81 | 0.76 | 0.70 | 0.71 | 0.74 | 0.78 | 0.74 | 0.77 | 0.74 | 0.72 | 0.79 | 0.73 |
| Mean | 0.89 | 0.82 | 0.82 | 0.74 | 0.78 | 0.83 | 0.82 | 0.81 | 0.82 | 0.81 | 0.83 | 0.81 | 0.83 | 0.73 | 0.75 | 0.83 | 0.81 |
| Pollinizers | Cultivar | ‘Edda’ | ‘Opal’ | ‘Jubileum’ | ‘Reeves’ | ‘Mallard’ | ‘Avalon’ | ‘Cacanska Lepotica’ | ‘Valor’ |
|---|---|---|---|---|---|---|---|---|---|
| Season | |||||||||
| ‘Edda’ | ‘17 | ||||||||
| ‘18 | |||||||||
| ‘Opal’ | ‘17 | 93.5 | 73 | 73 | 83.5 | 36.5 | 50 | 20 | |
| ‘18 | 96.5 | 50 | 43.5 | 43 | 53.5 | 60 | 26.5 | ||
| ‘Jubileum’ | ‘17 | 6.5 | 3.5 | 6.5 | 20 | 23.5 | |||
| ‘18 | 16.5 | 33.5 | 30 | 13.5 | |||||
| ‘Reeves’ | ‘17 | 3.5 | 13.5 | 3 | 3.5 | 71.5 | |||
| ‘18 | 23 | 13.5 | 10 | 3.5 | 73.5 | ||||
| ‘Mallard’ | ‘17 | 16.5 | 6.5 | ||||||
| ‘18 | 3.5 | 6.5 | 20 | 23 | |||||
| ‘Avalon’ | ‘17 | 13.5 | |||||||
| ‘18 | 3.5 | 3.5 | |||||||
| ‘Valor’ | ‘17 | 10 | 2 | ||||||
| ‘18 | 3.5 | 3.5 | |||||||
| ‘Cacanska Lepotica’ | ‘17 | 3.5 | 30 | 23.5 | 93 | 6.5 | |||
| ‘18 | 3.5 | 6.5 | 76.5 | ||||||
| ‘Herman’ | ‘17 | ||||||||
| ‘18 | |||||||||
| ‘Victoria’ | ‘17 | 3.5 | 3.5 | ||||||
| ‘18 | 36.5 | 16.5 | |||||||
| ‘Excalibur’ | ‘17 | 3.5 | |||||||
| ‘18 | 3.5 |
| Year | Flowering | ‘Edda’ | ‘Opal’ | ‘Jubileum’ | ‘Reeves’ | ‘Mallard’ | ‘Avalon’ | ‘Cacanska Lepotica’ | ‘Valor’ | ‘Victoria’ | ‘Excalibur’ |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2017 | First Bloom | 5/11 | 5/12 | 5/13 | 5/14 | 5/9 | 5/9 | 5/13 | 5/17 | 5/12 | 5/14 |
| Full Bloom | 5/13 | 5/14 | 5/15 | 5/16 | 5/11 | 5/11 | 5/15 | 5/22 | 5/15 | 5/16 | |
| Petal fall | 5/17 | 5/18 | 5/19 | 5/19 | 5/17 | 5/19 | 5/19 | 5/26 | 5/19 | 5/21 | |
| 2018 | First Bloom | 5/9 | 5/9 | 5/9 | 5/12 | 5/8 | 5/9 | 5/12 | 5/13 | 5/9 | 5/10 |
| Full Bloom | 5/10 | 5/11 | 5/10 | 5/14 | 5/10 | 5/10 | 5/14 | 5/15 | 5/10 | 5/12 | |
| Petal fall | 5/16 | 5/16 | 5/16 | 5/20 | 5/16 | 5/16 | 5/23 | 5/23 | 5/16 | 5/19 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meland, M.; Frøynes, O.; Fotiric Akšić, M.; Pojskić, N.; Kalamujić Stroil, B.K.; Lasic, L.; Gasi, F. Identifying Pollen Donors and Success Rate of Individual Pollinizers in European Plum (Prunus domestica L.) Using Microsatellite Markers. Agronomy 2020, 10, 264. https://doi.org/10.3390/agronomy10020264
Meland M, Frøynes O, Fotiric Akšić M, Pojskić N, Kalamujić Stroil BK, Lasic L, Gasi F. Identifying Pollen Donors and Success Rate of Individual Pollinizers in European Plum (Prunus domestica L.) Using Microsatellite Markers. Agronomy. 2020; 10(2):264. https://doi.org/10.3390/agronomy10020264
Chicago/Turabian StyleMeland, Mekjell, Oddmund Frøynes, Milica Fotiric Akšić, Naris Pojskić, Belma Kalamujić Kalamujić Stroil, Lejla Lasic, and Fuad Gasi. 2020. "Identifying Pollen Donors and Success Rate of Individual Pollinizers in European Plum (Prunus domestica L.) Using Microsatellite Markers" Agronomy 10, no. 2: 264. https://doi.org/10.3390/agronomy10020264
APA StyleMeland, M., Frøynes, O., Fotiric Akšić, M., Pojskić, N., Kalamujić Stroil, B. K., Lasic, L., & Gasi, F. (2020). Identifying Pollen Donors and Success Rate of Individual Pollinizers in European Plum (Prunus domestica L.) Using Microsatellite Markers. Agronomy, 10(2), 264. https://doi.org/10.3390/agronomy10020264