Optimizing Nitrogen Application for Growth and Productivity of Pomegranates

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Growth Indices
2.3. Water Consumption and Nutrient Uptake
2.4. Leaf Analyses
2.5. Nitrogen Use Efficiency
- PFP Partial factor productivity of applied N = fruit yield (of the treatment)/N application amount;
- NUpE Nitrogen uptake efficiency = N uptake (of the treatment)/N application;
- NUtE Nitrogen utilization efficiency = fruit yield (of the treatment)/N uptake;
2.6. Statistical Analyses
3. Results
3.1. Nitrogen Concentration in the Plant
3.2. Vegetative Growth Indices
3.3. Nitrogen Uptake
3.4. Reproduction Indices
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Figure | Parameter | Year or Month | Equation (Y = …) | R2 | P |
|---|---|---|---|---|---|
| 1 A | Leaf N concentration | 2016-May | 16.65X/(2.502 + X(1 + X/1.421e157)) | 0.6104 | <0.0001 |
| 2016-Sept. | 14.11X/(3.769 + X(1 + X/58853)) | 0.7716 | <0.0001 | ||
| 1 B | 2017-May | 22.3X/(6.561 + X(1 + X/9244)) | 0.8788 | <0.0001 | |
| 2017-Sept. | 18.18X/(5.233 + X(1 + X/1083)) | 0.6329 | <0.0001 | ||
| 1 C | 2018-May | 20.54X/(4.218 + X(1 + X/4.115e139)) | 0.8169 | <0.0001 | |
| 2018-Sept. | 18.47X/(4.016 + X(1 + X/48734)) | 0.7763 | <0.0001 | ||
| 2 A | Leaf chlorophyll content | 2016 | 64.44X/(4.083 + X(1 + X/1.028e104)) | 0.8656 | <0.0001 |
| 2017 | 62.33X/(5.287 + X(1 + X/6403)) | 0.9335 | <0.0001 | ||
| 2018 | 68.16X/(2.86 + X(1 + X/1.314e92)) | 0.8459 | <0.0001 | ||
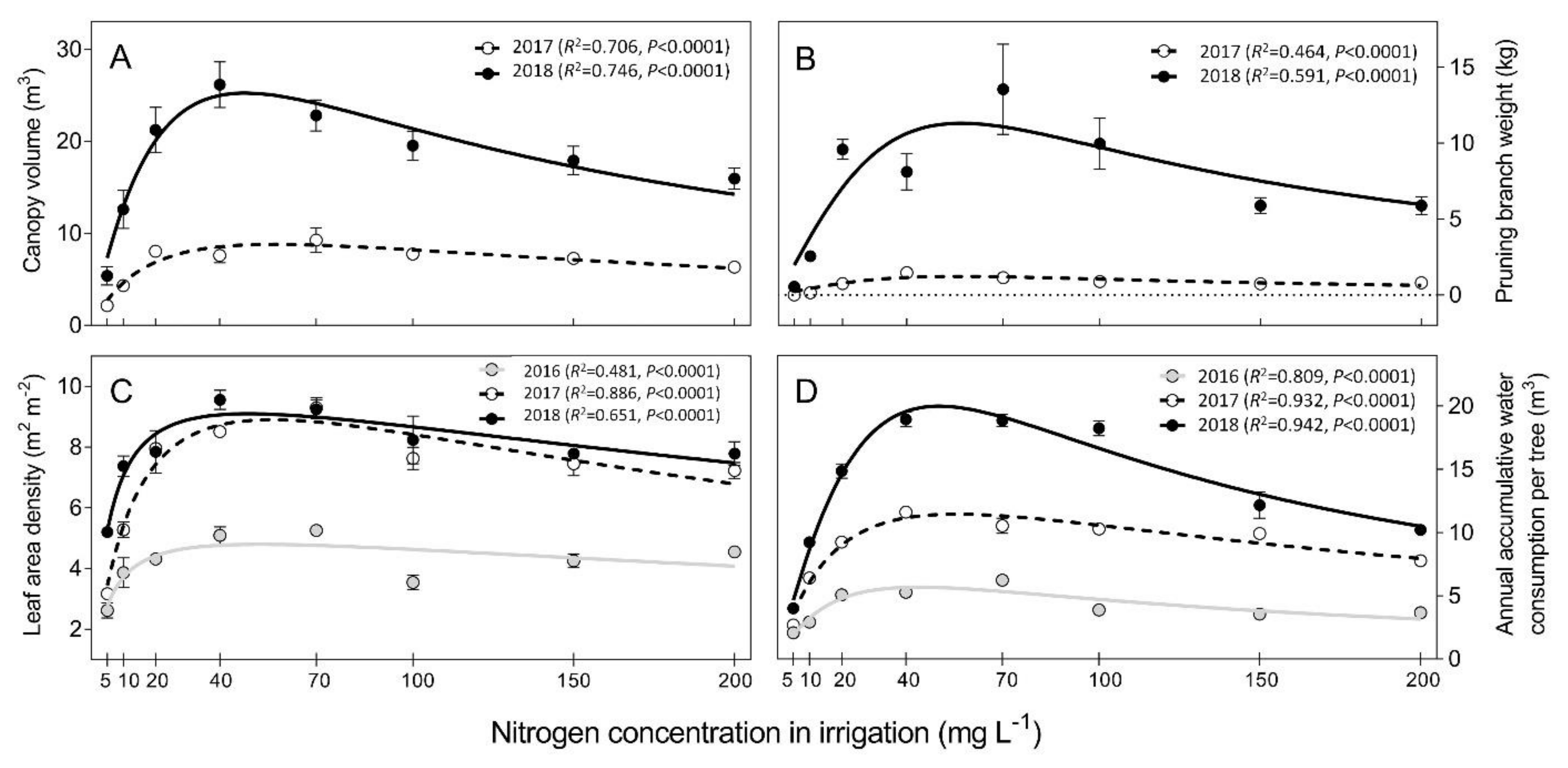
| 3 A | Canopy volume | 2017 | 16.46X/(24.93 + X(1 + X/131.7)) | 0.7059 | <0.0001 |
| 2018 | 69.74X/(42.18 + X(1 + X/54.29)) | 0.7463 | <0.0001 | ||
| 3 B | Pruning branch weight | 2017 | 1434X/(32798 + X(1 + X/0.09584)) | 0.4641 | <0.0001 |
| 2018 | 8922X/(22432 + X(1 + X/0.1446)) | 0.5911 | <0.0001 | ||
| 3 C | Leaf density | 2016 | 5.775X/(5.346 + X(1 + X/516.2)) | 0.4807 | <0.0001 |
| 2017 | 13.47X/(14.42 + X(1 + X/219.3)) | 0.8860 | <0.0001 | ||
| 2018 | 11.17X/(5.542 + X(1 + X/429.9)) | 0.6508 | <0.0001 | ||
| 3 D | Annual accumulated water consumption | 2016 | 13.33X/(29.61 + X(1 + X/65.26)) | 0.8095 | <0.0001 |
| 2017 | 21.75X/(25.09 + X(1 + X/124)) | 0.9315 | <0.0001 | ||
| 2018 | 105.2X/(107.1 + X(1 + X/23.58)) | 0.9417 | <0.0001 | ||
| 5 | Fruit yield | 2017–18 | 279X/(27.35 + X(1 + X/132.2)) | 0.7248 | <0.0001 |
| Nitrogen uptake | 2017–18 | 10002X/(293.8 + X(1 + X/74.46)) | 0.9336 | <0.0001 | |
| Nitrogen utilization efficiency | 2017–18 | 167.5X/(0.4928 + X(1 + X/27.31)) | 0.8354 | <0.0001 |
| Figure | Parameter | Year or Month | 5 | 10 | 20 | 40 | 70 | 100 | 150 | 200 |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 A | Leaf N concentration | 2016-May | f | d | c | bc | ab | ab | ab | a |
| 2016-Sept. | c | c | b | ab | ab | ab | ab | a | ||
| 1 B | 2017-May | f | e | d | cd | ab | cd | bc | a | |
| 2017-Sept. | c | b | a | a | ab | a | a | a | ||
| 1 C | 2018-May | d | d | c | bc | ab | c | bc | a | |
| 2018-Sept. | d | c | b | b | ab | b | ab | a | ||
| 2 A | Leaf chlorophyll content | 2016 | d | c | b | b | a | b | a | a |
| 2017 | f | e | d | c | bc | abc | a | ab | ||
| 2018 | d | d | c | bc | ab | ab | ab | a | ||
| 3 A | Canopy volume | 2017 | d | c | ab | ab | a | ab | b | b |
| 2018 | e | d | abc | a | ab | bc | bc | cd | ||
| 3 B | Pruning branch weight | 2017 | d | cd | bc | a | ab | ab | bc | abc |
| 2018 | e | de | b | bc | a | ab | cd | cd | ||
| 3 C | Leaf density | 2016 | d | bc | bc | a | a | b | bc | ab |
| 2017 | e | d | bc | ab | a | c | c | c | ||
| 2018 | d | c | c | a | ab | bc | c | c | ||
| 3 D | Annual accumulated water consumption | 2016 | e | d | b | b | a | c | c | c |
| 2017 | f | e | c | a | b | b | bc | d | ||
| 2018 | e | d | b | a | a | a | c | d | ||
| 5 | Fruit yield | 2017–18 | d | c | ab | a | a | ab | ab | b |
| Nitrogen uptake | 2017–18 | e | e | d | c | b | a | a | a | |
| Nitrogen utilization efficiency | 2017–18 | a | a | b | c | cd | d | cd | d |
References
- Bernardi, A.C.D.C.; Carmello, Q.A.D.C.; Carvalho, S.A.D.; Machado, E.C.; Medina, C.L.; Gomes, M.D.M.D.A.; Lima, D.M. Nitrogen, phosphorus and potassium fertilization interactions on the photosynthesis of containerized citrus nursery trees. J. Plant. Nutr. 2015, 38, 1902–1912. [Google Scholar] [CrossRef]
- Bonner, J.; Varner, J.E. Plant Biochemistry; Academic Press: New York, NY, USA, 2012. [Google Scholar]
- Lea, P.J.; Morot-Gaudry, J.F. Plant Nitrogen; Springer Science & Business Media and INRA: Paris, France, 2013. [Google Scholar]
- Lin, Y.L.; Tsay, Y.F. Influence of differing nitrate and nitrogen availability on flowering control in Arabidopsis. J. Exp. Bot. 2017, 68, 2603–2609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huett, D. Prospects for manipulating the vegetative-reproductive balance in horticultural crops through nitrogen nutrition: A review. Aust. J. Agric. Res. 1996, 47, 47–66. [Google Scholar] [CrossRef]
- Natale, W.; Rozane, D.E.; de Medeiros Corrêa, M.C.; Parent, L.E.; de Deus, J.A.L. Diagnosis and management of nutrient constraints in guava. Fruit Crop. 2020, 711–722. [Google Scholar]
- Zipori, I.; Erel, R.; Yermiyahu, U.; Ben-Gal, A.; Dag, A. Sustainable Management of Olive Orchard Nutrition: A Review. Agriculture 2020, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- De Araujo, Q.R.; De, A.; Loureiro, G.A.; Ahnert, D.; Escalona-Valdez, R.A.; Baligar, V.C. Interactions between Soil, Leaves and Beans Nutrient Status and Dry Biomass of Beans and Pod Husk of Forastero Cacao: An Exploratory Study. Commu. Soil Sci. Plant Anal. 2020, 1–15. [Google Scholar]
- Zhai, Y.; Lei, Y.; Wu, J.; Teng, Y.; Wang, J.; Zhao, X.; Pan, X. Does the groundwater nitrate pollution in China pose a risk to human health? A critical review of published data. Environ. Sci.. Pollut. Res. 2017, 24, 3640–3653. [Google Scholar] [CrossRef]
- Zhai, Y.; Zhao, X.; Teng, Y.; Li, X.; Zhang, J.; Wu, J.; Zuo, R. Groundwater nitrate pollution and human health risk assessment by using HHRA model in an agricultural area, NE China. Ecotoxicol. Environ. Saf. 2017, 137, 130–142. [Google Scholar] [CrossRef]
- Baram, S.; Couvreur, V.; Harter, T.; Read, M.; Brown, P.; Kandelous, M.; Smart, D.R.; Hopmans, J.W. Estimating nitrate leaching to groundwater from orchards: Comparing crop nitrogen excess, deep vadose zone data-driven estimates, and HYDRUS modeling. Vadose Zone J. 2016, 15. [Google Scholar] [CrossRef] [Green Version]
- Boyle, E. Nitrogen pollution knows no bounds. Science 2017, 356, 700–701. [Google Scholar] [CrossRef]
- Cameira, M.; Mota, M. Nitrogen related diffuse pollution from horticulture production—mitigation practices and assessment strategies. Horticulturae 2017, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Erel, R.; Yermiyahu, U.; Van Opstal, J.; Ben-Gal, A.; Schwartz, A.; Dag, A. The importance of olive (Olea europaea L.) tree nutritional status on its productivity. Sci. Hortic. 2013, 159, 8–18. [Google Scholar] [CrossRef]
- Fernández-Escobar, R. Use and abuse of nitrogen in olive fertilization. Acta Hortic. 2011, 888, 249–257. [Google Scholar] [CrossRef]
- Zaman, Q.; Schumann, A.; Miller, W. Variable rate nitrogen application in Florida citrus based on ultrasonically-sensed tree size. Appl. Eng. Agric. 2005, 21, 331–335. [Google Scholar] [CrossRef]
- Nava, G.; Dechen, A.R.; Nachtigall, G.R. Nitrogen and potassium fertilization affect apple fruit quality in southern Brazil. Commun. Soil Sci. Plant Anal. 2007, 39, 96–107. [Google Scholar] [CrossRef]
- Sperling, O.; Karuanakaran, R.; Erel, R.; Yasuor, H.; Klipcan, L.; Yermiyahu, U. Excessive nitrogen impairs hydraulics, limits photosynthesis, and alters the metabolic composition of almond trees. Plant Physiol. Biochem. 2019, 143, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Haberman, A.; Dag, A.; Shtern, N.; Zipori, I.; Erel, R.; Ben-Gal, A.; Yermiyahu, U. Significance of proper nitrogen fertilization for olive productivity in intensive cultivation. Sci. Hortic. 2019, 246, 710–717. [Google Scholar] [CrossRef]
- Muñoz-Huerta, R.; Guevara-Gonzalez, R.; Contreras-Medina, L.; Torres-Pacheco, I.; Prado-Olivarez, J.; Ocampo-Velazquez, R. A review of methods for sensing the nitrogen status in plants: Advantages, disadvantages and recent advances. Sensors 2013, 13, 10823–10843. [Google Scholar]
- Babu, D. Floral biology of pomegranate (Punica granatum L.). Pomegranate 2010, 4, 45–50. [Google Scholar]
- Romano, K.R.; Finco, F.D.B.A.; Rosenthal, A.; Finco, M.V.A.; Deliza, R. Willingness to pay more for value-added pomegranate juice (Punica granatum L.): An open-ended contingent valuation. Food Res. Int. 2016, 89, 359–364. [Google Scholar] [CrossRef] [Green Version]
- Holland, D.; Hatib, K.; Bar-Ya’akov, I. Pomegranate: Botany, Horticulture, Breeding. Hortic. Rev. 2009, 35, 127–191. [Google Scholar]
- Venkataramudu, K.; Naik, S.R.; Viswanath, M.; Chandramohan, G. Packaging and storage of pomegranate fruits and arils: A review. Int. J. Chem. 2018, 6, 1964–1967. [Google Scholar]
- Rodríguez, P.; Mellisho, C.; Conejero, W.; Cruz, Z.; Ortuno, M.; Galindo, A.; Torrecillas, A. Plant water relations of leaves of pomegranate trees under different irrigation conditions. Environ. Exp. Bot. 2012, 77, 19–24. [Google Scholar] [CrossRef]
- Sulochanamma, B.; Yellamanda Reddy, T.; Subbi Reddy, G. Effect of basin and drip irrigation on growth, yield and water use efficiency in pomegranate cv. Ganesh. Acta Hortic. 2005, 696, 277–279. [Google Scholar] [CrossRef]
- Mellisho, C.; Egea, I.; Galindo, A.; Rodríguez, P.; Rodríguez, J.; Conejero, W.; Romojaro, F.; Torrecillas, A. Pomegranate (Punica granatum L.) fruit response to different deficit irrigation conditions. Agric. Water Manag. 2012, 114, 30–36. [Google Scholar] [CrossRef]
- Glozer, K.; Ferguson, L. Pomegranate Production in Afghanistan; UCDAVIS College of Agricultural and Environmental Sciences: Davis, CA, USA, 2008. [Google Scholar]
- Ayars, J.E.; Phene, C.J.; Phene, R.C.; Gao, S.; Wang, D.; Day, K.R.; Makus, D.J. Determining pomegranate water and nitrogen requirements with drip irrigation. Agric. Water Manag. 2017, 187, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Carranca, C.; Brunetto, G.; Tagliavini, M. Nitrogen nutrition of fruit trees to reconcile productivity and environmental concerns. Plants 2018, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Dhillon, W.; Gill, P.; Singh, N. Effect of nitrogen, phosphorus and potassium fertilization on growth, yield and quality of pomegranate ‘Kandhari’. Acta Hortic. 2011, 890, 327–332. [Google Scholar] [CrossRef]
- Wang, D.; Ayars, J.; Tirado-Corbala, R.; Makus, D.; Phene, C.; Phene, R. Water and nitrogen management of young and maturing pomegranate trees; III International Symposium on Pomegranate and Minor. Mediterranean Fruits. ISHS Acta Hortic. 2013, 1089, 395–401. [Google Scholar]
- Nerya, O.; Levin, A. Innovative treatment of pomegranates from harvest to market; III International Symposium on Pomegranate and Minor. Mediterranean Fruits. ISHS Acta Hortic. 2013, 1089, 489–493. [Google Scholar]
- Easlon, H.M.; Bloom, A.J. Easy Leaf Area: Automated digital image analysis for rapid and accurate measurement of leaf area. Appl. Plant Sci. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Fallahi, E.; Fallahi, B.; Kiester, M.J. Evapotranspiration-based irrigation systems and nitrogen effects on yield and fruit quality at harvest in fully mature ‘Fuji’apple trees over four years. HortScience 2018, 53, 38–43. [Google Scholar] [CrossRef]
- Nirgude, V.; Misra, K.; Singh, P.; Singh, A.; Singh, N. NPK fertigation of stone fruit crops: A review. Int. J. Chem. Stud. 2018, 6, 3134–3142. [Google Scholar]
- Zhang, L.; Gao, Y.; Zhang, Y.; Liu, J.; Yu, J. Changes in bioactive compounds and antioxidant activities in pomegranate leaves. Sci. Hortic. 2010, 123, 543–546. [Google Scholar] [CrossRef]
- Muhammad, S.; Sanden, B.L.; Lampinen, B.D.; Saa, S.; Siddiqui, M.I.; Smart, D.R.; Olivos, A.; Shackel, K.A.; DeJong, T.; Brown, P.H. Seasonal changes in nutrient content and concentrations in a mature deciduous tree species: Studies in almond (Prunus dulcis (Mill.) DA Webb). Eur. J. Agron. 2015, 5, 52–68. [Google Scholar] [CrossRef]
- Bustan, A.; Avni, A.; Yermiyahu, U.; Ben-Gal, A.; Riov, J.; Erel, R.; Zipori, I.; Dag, A. Interactions between fruit load and macroelement concentrations in fertigated olive (Olea europaea L.) trees under arid saline conditions. Sci. Hortic. 2013, 152, 44–55. [Google Scholar] [CrossRef]
- Stateras, D.C.; Moustakas, N.K. Seasonal changes of macro-and micro-nutrients concentration in olive leaves. J. Plant Nutr. 2018, 41, 186–196. [Google Scholar] [CrossRef]
- Lazare, S.; Haberman, A.; Yermiyahu, U.; Erel, R.; Simenski, E.; Dag, A. Avocado rootstock influences scion leaf mineral content. Arch. Agron. Soil Sci. 2019, 1–11. [Google Scholar] [CrossRef]
- Rubio-Covarrubias, O.A.; Brown, P.H.; Weinbaum, S.A.; Johnson, R.S.; Cabrera, R.I. Evaluating foliar nitrogen compounds as indicators of nitrogen status in Prunus persica trees. Sci. Hortic. 2009, 120, 27–33. [Google Scholar] [CrossRef]
- Fox, R.H.; Walthall, C.L. Crop monitoring technologies to assess nitrogen status. Nitrogen Agric. Syst. 2008, 647–674. [Google Scholar] [CrossRef]
- Di Gioia, F.; Gonnella, M.; Buono, V.; Ayala, O.; Cacchiarelli, J.; Santamaria, P. Calcium cyanamide effects on nitrogen use efficiency, yield, nitrates, and dry matter content of lettuce. Agron. J. 2017, 109, 354–362. [Google Scholar] [CrossRef]
- Neto, C.; Carranca, C.; de Varennes, A.; Oliveira, C.; Clemente, J.; Sobreiro, J. Nitrogen Use Efficiency of Drip-irrigated ‘Rocha’ Pear Trees. Acta Hortic. 2006, 721, 337. [Google Scholar] [CrossRef]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [Green Version]
- Glass, A.D. Nitrogen use efficiency of crop plants: Physiological constraints upon nitrogen absorption. Crit. Rev. Plant Sci. 2003, 22, 453–470. [Google Scholar] [CrossRef]
- Kahramanoglu, I.; Usanmaz, S. Pomegranate Production and Marketing; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Blumenfeld, A.; Shaya, F.; Hillel, R. Cultivation of pomegranate. Options Méd. Ser. 2000, 42, 143–147. [Google Scholar]
- Khattab, M.M.; Shaban, A.E.; El-Shrief, A.H.; El-Deen, M.A. Growth and productivity of pomegranate trees under different irrigation levels. I: Vegetative growth and fruiting. J. Hortic. Sci. Ornam. Plants 2011, 3, 194–198. [Google Scholar]
- Intrigliolo, D.; Nicolas, E.; Bonet, L.; Ferrer, P.; Alarcón, J.; Bartual, J. Water relations of field grown Pomegranate trees (Punica granatum) under different drip irrigation regimes. Agric. Water Manag. 2011, 98, 691–696. [Google Scholar] [CrossRef]
- Quiñones, A.; Martínez-Alcántara, B.; Primo-Millo, E.; Legaz, F. Fertigation: Concept and application in citrus. In Advances in Citrus Nutrition; Springer: Dordrecht, The Netherlands, 2012; pp. 281–301. [Google Scholar]
- Gabriel, J.L.; Quemada, M. Water Management for Enhancing Crop Nutrient Use Efficiency and Reducing Losses. In Advances in Research on Fertilization Management of Vegetable Crops; Springer: Dordrecht, The Netherlands, 2017; pp. 247–265. [Google Scholar]




| Periods of Time | N Concentration in Irrigation (mg L−1) | |||||||
|---|---|---|---|---|---|---|---|---|
| 5 | 10 | 20 | 40 | 70 | 100 | 150 | 200 | |
| 2016/2017 | 211% d | 249% c | 281% bc | 286% bc | 329% a | 310% ab | 287% bc | 272% bc |
| 2017/2018 | 137% b | 152% a | 148% a | 144% ab | 148% a | 150% a | 145% ab | 144% ab |
| 2018/2019 | 125% b | 125% b | 131% ab | 140% a | 127% b | 128% b | 128% b | 125% b |
| Year | N in Irrigation (mg N L−1) | Yield (kg tree−1) | Fruit Per Tree | N app. Amount (g tree−1) | N Uptake (g tree−1) | N Ratio Uptake/Application | PFP (kg kg−1) | NUtE (kg kg−1) | Volume of Irrigation (m3 tree−1) | WUE (kg m−3) | WUpE (kg m−3) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2017 | 5 | 3.4 c | 7 c | 33 | 27 f | 0.81 a | 101.2 bc | 124 b | 6.6 | 0.51 c | 1.25 c |
| 10 | 19.4 b | 34 b | 121 | 104 ef | 0.86 a | 160.2 a | 186 a | 12.1 | 1.60 b | 3.03 b | |
| 20 | 43.4 a | 75 a | 309 | 255 de | 0.82 a | 140.3 ab | 170 ab | 15.5 | 2.81 a | 4.66 a | |
| 40 | 48.2 a | 80 a | 623 | 355 d | 0.57 b | 77.4 cd | 139 ab | 15.6 | 3.09 a | 4.14 ab | |
| 70 | 43.6 a | 75 a | 1203 | 694 c | 0.58 b | 36.3 de | 63 c | 17.2 | 2.54 a | 4.25 ab | |
| 100 | 38.2 a | 65 a | 1588 | 822 bc | 0.52 b | 24.1 e | 47 c | 15.9 | 2.41 ab | 3.72 ab | |
| 150 | 44.9 a | 77 a | 2305 | 957 ab | 0.42 c | 19.5 e | 48 c | 15.4 | 2.92 a | 4.5 ab | |
| 200 | 36.8 a | 66 a | 2737 | 1037 a | 0.38 c | 13.4 e | 38 c | 13.7 | 2.69 a | 4.74 a | |
| 2018 | 5 | 15.8 d | 29 c | 48 | 48 e | 1.00 a | 333.3 a | 333 a | 9.5 | 1.67 c | 3.95 e |
| 10 | 51.2 c | 85 b | 179 | 179 e | 1.00 a | 287.7 a | 287 a | 17.8 | 2.88 b | 5.57 bcd | |
| 20 | 89.0 ab | 147 a | 502 | 468 d | 0.93 a | 177.3 b | 190 b | 25.1 | 3.55 ab | 5.96 abc | |
| 40 | 92.8 ab | 159 a | 1170 | 801 bc | 0.69 b | 79.3 c | 118 bc | 29.2 | 3.17 ab | 4.9 cde | |
| 70 | 97.3 a | 177 a | 2065 | 749 c | 0.36 d | 47.1 c | 131 bc | 29.5 | 3.30 ab | 5.18 cde | |
| 100 | 79.8 ab | 137 a | 2824 | 1294 a | 0.46 c | 28.3 c | 64 c | 28.2 | 2.83 b | 4.39 de | |
| 150 | 89.8 ab | 177 a | 3549 | 922 bc | 0.26 e | 25.3 c | 100 c | 23.7 | 3.79 a | 7.44 a | |
| 200 | 71.6 bc | 140 a | 3991 | 947 b | 0.24 e | 17.9 c | 78 c | 20.0 | 3.59 ab | 7.03 ab |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazare, S.; Lyu, Y.; Yermiyahu, U.; Heler, Y.; Ben-Gal, A.; Holland, D.; Dag, A. Optimizing Nitrogen Application for Growth and Productivity of Pomegranates. Agronomy 2020, 10, 366. https://doi.org/10.3390/agronomy10030366
Lazare S, Lyu Y, Yermiyahu U, Heler Y, Ben-Gal A, Holland D, Dag A. Optimizing Nitrogen Application for Growth and Productivity of Pomegranates. Agronomy. 2020; 10(3):366. https://doi.org/10.3390/agronomy10030366
Chicago/Turabian StyleLazare, Silit, Yang Lyu, Uri Yermiyahu, Yehuda Heler, Alon Ben-Gal, Doron Holland, and Arnon Dag. 2020. "Optimizing Nitrogen Application for Growth and Productivity of Pomegranates" Agronomy 10, no. 3: 366. https://doi.org/10.3390/agronomy10030366