Abstract
This paper aims to elucidate the effects of exogenous brassinolide (BL) on maize germination and seedling growth under chilling stress. The cold-resistant maize hybrid Tiannong 9 and the cold-sensitive hybrid Tianhe 1 were soaked at the germination stage (6 °C) and leaves were sprayed at seedling stage (4 °C), with BL at concentrations of 0, 0.01, 0.1, and 1 mg/L. The germination rate of the maize seeds and the changes in seedling biomass, antioxidant, photosynthetic, and plant endogenous hormone systems and chloroplast ultrastructures were determined. The results showed that the optimum concentration of BL to alleviate chilling stress in maize seedlings was 0.1 mg/L. This rate effectively increased the germination rate and plant biomass of maize and significantly increased the superoxide dismutase (SOD) peroxidase (POD) and catalase (CAT) activities, the net photosynthetic rate (Pn), stomatal conductance (gs) and transpiration rate (Tr), and seedling auxin (IAA), gibberellin (GA3) and trans zeatin nucleoside (t-ZR) contents under chilling stress. In addition, BL significantly reduced the malondialdehyde (MDA) content, abscisic acid (ABA) content, and intercellular carbon dioxide concentration (Ci). In the comparison of mesophyll cells, the chloroplast membrane of the treatment group was tightly attached to the stroma, and some of the plasma membranes were dissolved, but the overall structure of the chloroplast was relatively complete, and the osmiophilic granules were relatively few. The exogenous application of BL can effectively alleviate the damage caused by a low temperature in maize, maintain the normal characteristics of seedlings in chilling environments, and ensure the development and growth of plant tissue in the later stage.
1. Introduction
Maize (Zea mays L.) is one of the world’s most cultivated cereal crops and is also widely grown for food, feed and industrial purposes [1]. As the world’s second largest maize producer, according to the FAO’s 2017 data, maize production of China accounts for 21% of the global total, accounting for more than one-third of cereal production in China. Maize originated in the tropics and is a C4 thermophilic crop that is highly sensitive to low temperatures at all stages of growth and development [2]. Low temperature usually refers to the temperature range of 5–15 °C [3] and is one of the main abiotic stresses that limits plant growth and geographical distributions [4]. When the temperature is below 15 °C, maize growth is strongly restricted [5], in a manner affecting seed germination and possibly resulting in growth inhibition, chlorosis, wilting, necrosis and even death of the seedlings [6]. At this temperature, in addition to damage to the photosynthetic systems of the leaves, complex physiological and biochemical changes in the plants themselves can occur [7], resulting in a decline in biomass and yield, which affects maize yield [8].
Brassinosteroids (BLs) are an important of polyhydroxylated sterols found in the plant kingdom and are widely used in agriculture to increase production and enhance plant resistance to environmental stress [9]. Since Hamada [10] first reported that BL has an ameliorating effect on plant abiotic stress, many researchers have conducted extensive research on the role of BL [11,12]. BL can induce plant tolerance to abiotic stresses such as high temperature, low temperature [13,14], drought [15], salinity [16] and heavy metals [17]. It has a positive contribution to the cellular biochemical, physiological and morphological processes of higher plants, especially affecting cell division, antioxidant metabolism, gas exchange and the plant growth rate [18].
Interestingly, there are limited studies on the effects of the exogenous application of BL on maize germination and the physiological characteristics of seedlings under chilling stress. Recent studies have focused on the effects of exogenous application of BL on the physiological characteristics of horticultural crops such as tomato [19], grapes [20] and cucumber [21] under chilling stress. Therefore, the objectives of this study were to (1) investigate the effect of different concentrations of BL on the germination and seedling growth of two different maize hybrids under chilling stress and (2) determine the optimal concentration of BL for mitigating chilling stress in maize seedlings.
2. Materials and Methods
2.1. Experimental Materials
Two maize hybrids, the cold-resistant variety Tiannong 9 and the cold-sensitive variety Tianhe 1, were used in the experiments. These two hybrids were identified through early seed germination and seedling experiments in our previous study [22]. (Tiannong9, T106 × W08, maturity 126d, active accumulated temperature 2570 °C Tianhe1, THA5W × TH21A, maturity 111d, active accumulated temperature 2115 °C). Brassinolide was purchased from Hefei Bomei Biotechnology Co., Ltd., China.
2.2. Experimental Design
Seed germination experiment: according to the previous research conclusions of our laboratory, We soaked seeds of the two hybrids in four concentrations of BL in solution, 0 (T0), 0.01 (T1), 0.1 (T2), and 1 mg/L (T3) at 6 °C and distilled water at 25 °C (CK), and each treatment had 3 replicates. First, completely filled seeds of uniform size were selected and disinfected with 0.5% sodium hypochlorite (NaClO) solution for 5 min and then rinsed with water 3 times. The seeds were soaked in the different BL solutions for 24 h. Subsequently, the seeds were transferred to a Petri dish (9 mm) with wet filter paper and a water depth of 1 mm, for germination under dark conditions in an incubator (Percival, Perry, IA USA). The seeds were incubated at 70% relative humidity (seedlings grow best at 70% relative humidity).
Seedling experiment: the selection and disinfection of seeds was performed as described above for the germination experiment. Then seeds were soaked in distilled water for 24 h before sowing. Seeding was carried out using a plastic pot (90 mm × 100 mm), and each treatment was carried out with 200 plants and 6 replicates. Maize seedlings, at the three-leaf and one-heart stage, were used for foliar spray treatment, which was carried out once at the same time in the morning with BL concentrations of 0 (T0), 0.01 (T1), 0.1 (T2), and 1 mg/L (T3). The spray amount per plant was the same, approximately 10 mL, and distilled water at 25 °C (CK) was used as the normal temperature control. At 24 h after the leaves were sprayed, the low temperature treatment group was placed in a Percival light incubator at 4 °C for low temperature treatment. The daytime light intensity was 100––550 μm/(m2·s), the illumination time was 12 h, and the relative humidity was 70% for 2 days. The normal temperature control treatment (CK) group was transferred to the 25 °C incubator for normal growth, and the other settings were the same as those of the low temperature treatment group. After the low temperature treatment, representative plants were sampled for determination of physiological and biochemical indexes.
2.3. Germination Percentage of Seeds
The number of germinated seeds of the different varieties of maize treated above was investigated daily, and the germination rate was calculated.
2.4. Plant Biomass, Height and Root Length
After the end of the treatment, collocated fresh leaves of the maize seedlings were selected for further measurements. The fresh weight (FW) of the aboveground and underground parts of the plants was measured by a JA2603B electronic balance (Shanghai Precision Scientific Instrument Co., Ltd., China), with an accuracy of 0.001. After initial drying at 105 °C for 30 min, plant parts were then dried at 80 °C to a constant weight, to determine the dry weight (DW). Plant height refers to the height from the base to the apical growth point, the root length refers to the length from the base to the apical growth point, and the plant height and the root length were measured by a ruler with a scale of 1 mm from randomly selected positions, with 3 repetitions per treatment.
2.5. Photosynthetic Parameters
A portable photosynthetic measurement apparatus GFS-3000 (WALZ, Germany) was used to determine the net photosynthetic rate (Pn) and other parameters on the second fully expanded leaves of maize seedlings at 9:00–12:00 (stomatal conductance (gs), transpiration rate (Tr) and intercellular CO2 concentration (Ci)), with measurements taken six times per treatment. The gas flow rate was set at 750, the CO2 concentration was 380 ppm, relative humidity was 85%, and the leaf chamber temperature was the corresponding treatment temperature. The illumination intensity was the same as that used in the treatment.
2.6. MDA Content
The MDA content was determined by the thiobarbituric acid (TBA) method [23]. First, 0.5 g of the leaves were weighed, and 5 mL of 10% TCA was added to homogenate samples in an ice bath. Samples were centrifuged at 4000× g for 10 min, and the supernatant was taken. Then, 2 mL of 0.67% TBA was added, and the mixture was boiled in a boiling water bath for 30 min, cooled to room temperature and then centrifuged once. The supernatant was measured for OD values at 450 nm, 532 nm, and 600 nm, using a spectrophotometer (PERSEE, Beijing, China), and the control tube was filled with 2 mL of distilled water.
2.7. Antioxidant Enzyme Activity
The maize leaf samples of 0.5 g were ground in 0.05 M PBS in an ice bath and centrifuged at 10000× g for 4 min at 4 °C. The supernatant was used for enzyme activity assays. SOD activity was determined by the NBT method [24]. The reaction mixture included 3 mL phosphate buffer (pH 7.8), 0.6 mL methionine, 0.6 mL NBT, 0.6 mL EDTA-2Na, and 100 μL enzyme extract under 4000 Lux illumination for 30 min; a blank was used for adjustment to zero, and absorbance was measured at 560 nm. POD activity was determined by the Fu method [25], and the guaiacol mixture was measured at 470 nm. Catalase (CAT) activity was determined by the methods of Aebi [26].
2.8. Phytohormones
Refer to the methods of previous studies [27,28]. A total of 0.1 g of the plant sample was weighed, ground in a mortar, and 1 mL of precooled 80% methanol was added and leached overnight at 4 °C. After centrifugation at 8000× g for 10 min, the supernatant was removed. The residue was re-extracted with 0.5 mL of 80% methanol for 2 h, and the supernatant was combined twice. Nitrogen was added to 0.3 mL, the volume was adjusted to 0.5 mL with water, and 0.5 mL of petroleum ether was added for extraction. The mixture was decolonized three times. The aqueous phase was adjusted to pH 2.8 with a 0.1 mol/L citric acid solution and extracted three times with 0.5 mL of ethyl acetate. The upper organic phase was combined and dried by a nitrogen blowing apparatus. The mobile phase was brought to a volume of 0.5 mL and collected with a syringe filter into a vial with a liner to be tested.
A Rigol L3000 (Rigol, Suzhou China) high-performance liquid chromatograph with a Kromasil C18 (AKZO NOBEL, Stockholm, Sweden) reversed-phase column (250 mm × 4.6 mm, 5 μm) was used for high-performance liquid chromatography. The mobile phase preparation was as follows: A: methanol, B: 1% aqueous acetic acid. The software set the injection volume to 10 μL in the test group, the flow rate was 0.8 mL/min, the column temperature was 30 °C, the sample time was 35 min, and the UV detection wavelength was 254 nm. Four hormones were used as standard samples (Sigma, Burlington MA USA). The IAA, GA3, t-ZR, and ABA concentrations were calculated based on calibration curves generated from internal standards.
2.9. Chloroplast Ultrastructure
Maize-treated leaf samples were cut into 1 × 3 mm rectangular pieces, placed in 2.5% glutaraldehyde (pH = 7.2), and allowed to stand in a 4 °C refrigerator. The cells were washed 3 times with 0.1 M phosphate buffer (pH = 6.8), fixed with 1% citrate fixative for 2 h, then washed three times with the same phosphate buffer, and samples were dehydrated and soaked in an ethanol gradient. The samples were embedded in epoxy resin for 5 days and then placed in a constant temperature incubator for 48 h. The samples were sliced (50 to 60 nm) with an ultramicrotome stained with uranyl acetate and lead citrate and observed by transmission electron microscopy (H-7650 Hitachi, Ltd., Tokyo Japan).
2.10. Statistical Analysis
All statistical analyses were performed using analysis of variance (ANOVA) in SPSS 22 (IBM, USA). Significance was determined at p < 0.05. Figures were drawn using Origin 2018 (OriginLab, USA).
3. Results
3.1. Seed Germination
Chilling stress significantly reduced the germination rate of the two maize varieties (Figure 1). Under chilling stress, the germination rate of the two varieties increased first and then decreased with the increase in BL concentration (T0–T3). Tiannong 9 germinated on the third day with CK treatment. The seeds began to germinate on the 10th day and germination ended on the 24th day with the T0, T1 and T3 treatments. The germination rates at the beginning and end of each treatment were 7.78% and 13.33%, 10.00% and 93.33%, 96.67%, and 98.89%, respectively. The germination started on the 10th day after the T2 treatment, and the germination was completed on the 18th day, and the germination rates at the beginning and the end were 26.67% and 100.00%, respectively. Tianhe 1 began to germinate on the 3rd day under the CK treatment, and all seeds had sprouted by the 6th day; germination started on the 10th day in T0, T1 and T3, and germination ended on the 24th day; the germination rates at the beginning and end of each treatment were 8.89% and 12.22%, 14.44% and 66.67%, 60.00% and 80.00%, respectively. Germination began on the 10th day after the T2 treatment, and the germination was completed on the 24th day, and the germination rates at the beginning and end were 20.00% and 85.56%, respectively.
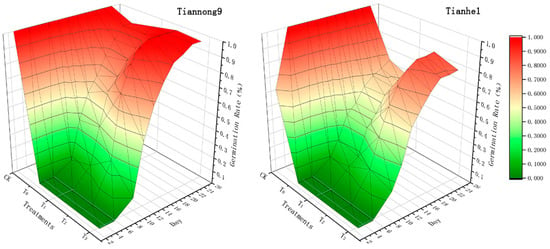
Figure 1.
The effects of soaking in different concentrations of BL on the maize seed germination rate.
BL soaking treatment significantly shortened the germination time of maize under chilling stress, especially the T2 concentration. Under T2, the initial and final germination rate of Tiannong 9 was 73.33 and 0 percentage points lower than that of the normal temperature CK treatment, and 18.89 and 6.67 percentage points higher than that of the low temperature T0 treatment. At the T2 concentration, the initial and final germination rates of Tianhe 1 were 80.00 and 14.44 percentage points lower than that of the normal temperature CK treatment, respectively, and 11.10 and 18.89 percentage points higher than the low temperature T0 treatment. So, T2 treatment was the most suitable concentration to alleviate the negative effects on the germination of seeds caused by low temperature. Among the varieties, Tiannong 9 had higher resistance to low temperature than Tianhe 1, and the effects of BL on Tiannong 9 were higher than Tianhe 1 under low temperature.
3.2. Seedling Biomass
As shown in Table 1, chilling stress can significantly reduce the plant height, root length, FW and DW of maize seedlings. The T1, T2 and T3 leaf spray treatments significantly alleviated the growth inhibition of seedlings under chilling stress. With the increase in the concentration of BL (T0–T3), the plant height and root length of the seedlings first increased and then decreased. The plant height and root length of the two varieties were the highest with the T2 treatment, and the plant heights were 32.50 and 34.76 cm, which was 20.07% and 18.93% higher than that of the T0 treatment, respectively. The root lengths were 26.30 and 32.83 cm, which increased by 41.40% and 64.15% compared with the T0 treatment, respectively. The same trend was also observed for the FW and DW of the plants. When Tiannong 9 and Tianhe 1 were treated by T2, the FW and DW of the aboveground parts increased by 44.15% and 55.92% and 57.65% and 49.15% compared with those of T0, respectively. The FW and DW of the underground parts increased by 32.04% and 38.79% and 49.87% and 14.26% compared with those of T0, respectively. The results showed that the T2 concentration was optimal for alleviating the damage to various agronomic traits of seedlings among the concentrations tested, and the difference among different varieties was significant. The T2 treatment better mitigated the negative effects on Tiannong 9 aboveground parts than on those of Tianhe 1. In contrast, the opposite was true for the underground parts.

Table 1.
The effects of different concentrations of BL on plant height, root length, shoot and root weight of maize seedlings under chilling stress.
3.3. MDA Content and Antioxidant Enzyme Activity
The effects of different concentrations of BL on the MDA content in maize seedlings under chilling stress are shown in Figure 2a. The MDA content of the two maize varieties was significantly higher than that of the CK under chilling stress. After treatment with T0–T3, the MDA content of seedlings decreased first and then increased with increasing BL concentration, and the MDA content reached the minimum level during the T2 treatment. The MDA content of Tiannong 9 and Tianhe 1 treated by T0 was increased by 234.04% and 267.07% compared with that of the CK treatment, respectively, and the increase range was Tianhe 1 > Tiannong 9. The MDA content of Tiannong 9 and Tianhe 1 decreased by 35.31% and 40.01%, respectively, following T2 compared with T0. The activities of SOD, POD and CAT in the two maize varieties were significantly higher than those of the CK under chilling stress (Figure 2b–d). After treatment with T0–T3, the activities of SOD, POD and CAT in maize seedlings significantly increased. With the increase in BL concentration, the antioxidant enzyme activity of seedlings was opposite the MDA content, and maximum antioxidant enzyme activities were realized with T2. The activities of SOD, POD and CAT of Tiannong 9 and Tianhe 1 treated by T0 were increased by 36.42%, 35.97%, 22.09% and 52.39%, 64.58%, and 19.23%, compared with those of the CK treatment, respectively. The increase range was Tianhe 1 > Tiannong 9. The SOD, POD and CAT activities of Tiannong 9 and Tianhe 1 were increased by 20.68%, 33.47%, 110.58% and 35.99%, 46.28%, 120.43%, respectively, with T2 compared with T0. The results showed that spraying BL could significantly alleviate the increase in the MDA content and increase the activities of SOD, POD and CAT in maize seedlings under chilling stress, and T2 treatment had the best mitigating effects on cold damage in maize seedlings. There were significant differences among the different varieties. The decrease in MDA contents and increase in the three antioxidant enzyme activities after BL spray treatment of maize seedlings at low temperature were as follows: Tianhe 1 > Tiannong 9.
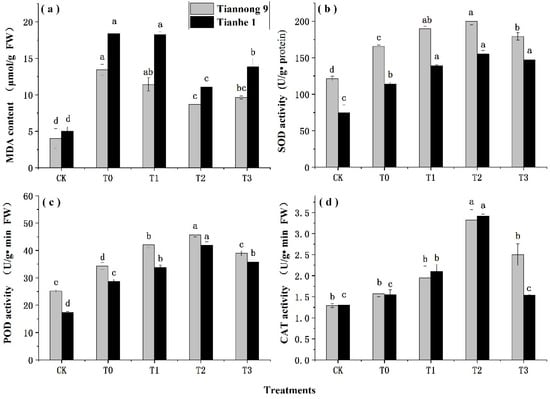
Figure 2.
Effects of different concentrations of BL on the MDA content and antioxidant enzyme activity of maize seedlings under chilling stress. CK treatment at 25 °C with distilled water, T0 treatment with BL at 0.00 mg/L, T1 treatment with BL at 0.01 mg/L, T2 treatment with BL at 0.1 mg/L, T3 treatment with BL at 1 mg/L. The data represent the means of independent measurements with three replicates. The mean values marked with different letters differ significantly at p < 0.05. (a) The variation trend of MDA content. (b) The variation trend of SOD activity. (c) The variation trend of POD activity. (d) The variation trend of CAT activity.
3.4. Photosynthesis
The Pn, gs and Tr (Figure 3a–c) of the two maize varieties were significantly lower under chilling stress than those of the normal temperature treatment, and the Ci (Figure 3d) was higher under chilling stress than under the normal temperature treatment. After treatment with T1, T2 and T3, the Pn, gs and Tr of maize seedlings under chilling stress significantly increased, and the Ci decreased. With the increase in BL concentration in T0–T3, the Pn, gs and Tr of the two varieties showed a trend of increasing and then decreasing, and the three indicators of the T2 treatment reached the maximum values. Under T2 treatment, the Pn of Tiannong 9 and Tianhe 1 increased by 25.43% and 52.75%, the gs increased by 11.86% and 11.26%, and the Tr increased by 10.78% and 5.93%, respectively, compared to those of T0. The change in Ci was the opposite. With the increase in BL concentration in T0–T3, the Ci of the two varieties decreased first and then increased, and the Ci value of T2 was lowest, but still higher than that of CK. The Ci of Tiannong 9 and Tianhe 1 decreased by 30.52% and 15.23%, respectively, compared with T0 during T2 treatment; the range in the variation of photosynthesis-related indicators was Pn > Ci > gs > Tr, and the difference among different varieties was significant. The decrease in Pn and the increase in Ci both showed the trend Tianhe 1 > Tiannong 9, and the trend for the gs and Tr was the opposite.
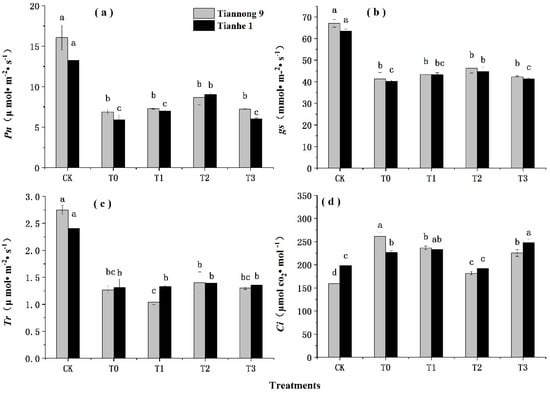
Figure 3.
Effects of different concentrations of BL on photosynthesis of maize seedlings under chilling stress. CK treatment at 25 °C with distilled water, T0 treatment with BL at 0.00 mg/L, T1 treatment with BL at 0.01 mg/L, T2 treatment with BL at 0.1 mg/L, T3 treatment with BL at 1 mg/L. The data represent the means of independent measurements with three replicates. The mean values marked with different letters differ significantly at p < 0.05. (a) The variation trend of Pn. (b) The variation trend of gs. (c) The variation trend of Tr. (d) The variation trend of Ci.
3.5. Plant Hormones
Figure 4 shows the changes in the IAA, GA3, t-ZR, and ABA content of maize seedlings under chilling stress, following treatment with different concentrations of BL. Chilling stress can significantly reduce the contents of IAA, GA3 and t-ZR (Figure 4a–c) in maize, and increase the content of ABA (Figure 4d). With the increase in BL concentration with T0–T3, IAA, GA3 and t-ZR showed a trend of increasing first and then decreasing and reached the maximum values under T2; the ABA content showed the opposite trend and reached the lowest value during T2.
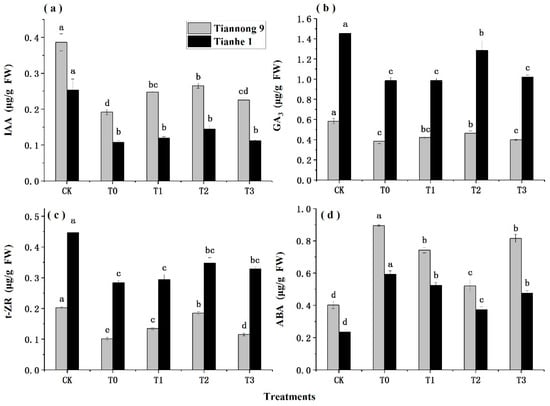
Figure 4.
Effects of different concentrations of BL on plant hormones of maize seedlings under chilling stress. CK treatment at 25 °C with distilled water, T0 treatment with BL at 0.00 mg/L, T1 treatment with BL at 0.01 mg/L, T2 treatment with BL at 0.1 mg/L, T3 treatment with BL at 1 mg/L. The data represent the means of independent measurements with three replicates. The mean values marked with different letters differ significantly at p < 0.05. (a) The variation trend of IAA content. (b) The variation trend of GA3 content. (c) The variation trend of t-ZR content. (d) The variation trend of ABA content.
The IAA content of Tiannong 9 and Tianhe 1 decreased differently under T2 and T0. The IAA content of the two varieties increased by 19.08% and 14.47% under T2 and CK, respectively, compared with the T0 and CK treatments. The same trend was also seen in the GA3 and t-ZR content changes, which increased by 13.53% and 20.61%, respectively. The t-ZR content increased by 41.14% and 14.26%, respectively. ABA also plays an important role in the abiotic stress response. The ABA content of Tiannong 9 and Tianhe 1 decreased by 41.80% and 58.96% under T2, respectively, compared with T0. The ABA content of the two varieties were 92.94% and 93.26% lower under T2 and CK than under T0 and CK, respectively.
3.6. Chloroplast Ultrastructure
Chloroplasts play a major role in photosynthesis. The ultrastructural characteristics of mesophyll cells are shown in Figure 5C, which shows the distribution of chloroplasts in the mesophyll cells of Tiannong 9 and Tianhe 1 at normal temperature. The chloroplasts are oval in shape and tightly distributed in the cells. The thylakoid structure is intact, and the lamellae are parallel and arranged horizontally. Compared with CK, under the chilling stress of T0 (Figure 5C,D), the shape of the chloroplast changed from elliptical to nearly circular. The plasma membrane gradually disassembled, and there was a clear separation between the chloroplast and plasma membrane. In severe cases, the chloroplast membrane disintegrated, small vesicles formed in the vacuoles, the intercellular gap enlarged, the thylakoid cavity inflated, the lamellae of the stroma and grana became disordered, and the osmiophilic granules increased significantly. Compared with T0, the chloroplast structure changed significantly with T2 (Figure 5E,F), although the shape of the chloroplast was still elliptical. The plasma membrane and the stroma were closely attached, and some plasma membranes were dissolved, but the overall structure of the chloroplast was relatively complete. Only some small vesicles of the stroma lamellae appeared, and the osmiophilic granules were relatively small. Under the low temperature T2 treatment, the plasma membrane was partially enlarged, as shown in Figure 5 G,H. The chloroplast bilayer membrane structure of maize mesophyll cells was evident, and some fractures appeared on the surface of the plasma membrane. The lamellae of the stroma and grana were slightly curved, and the structure of the grana lamellae was relatively good.
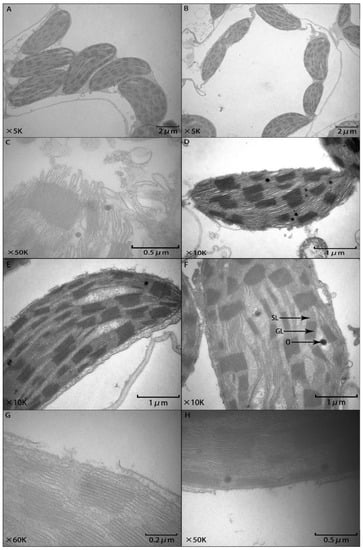
Figure 5.
Effects of different concentrations of BL on chloroplast ultrastructures of maize seedlings under chilling stress. (A,B): CK; (C,D): Chilling stress T0 treatment; (E,F): Chilling stress T2 treatment; (G,H): A partial magnification of the chloroplast under T2 treatment; GL: Grana lamellae; SL: Stroma lamella; O: Osmiophilic granule.
4. Discussion
4.1. Impacts of Chilling Stress on Maize
The optimum temperature for maize growth is 25–28 °C, which is suitable for seed germination, shoot elongation, leaf expansion and dry matter accumulation [29]; [30]. Seed germination and early seedling growth are the stages at which maize is most sensitive to environmental stress [31]. Low temperature is not conducive to the germination and early development of maize seeds and significantly prolongs the average time of seed germination and emergence, thus affecting crop growth and yield [32,33]. In this study, the germination time of the two maize cultivars was significantly prolonged by chilling stress, the germination rate was significantly reduced, and the extension of the germination time was different between varieties, which is similar to previous research results [34,35].
The effects of low temperature on plants are multifaceted. At the seedling growth stage, low temperatures can lead to complex symptoms of cell dysfunction, such as wilting, growth inhibition, yellowing and even death. Different tissues respond differently to low temperatures at different stages of growth and development. The shoot tip is very sensitive to low temperatures, which can have a negative impact on the leaf mass and growth rate [36,37]. However, in other experiments, chilling stress of the shoot tip caused delays in leaf development, but did not significantly affect the efficiency of photosynthesis [38]. Under chilling stress, leaf growth is inhibited due to an increase in cell cycle time and a decrease in the number of cells in the seedlings [39]. In this study, it was found that low temperatures significantly reduced the plant height, root length and biomass of seedlings of the two maize hybrids.
Chilling stress has been recognized as an environmental factor limiting plant growth and agricultural production [40]. Symptoms of hypothermic injury are caused by the production of large amounts of reactive oxygen species (ROS), such as superoxide radicals, singlet oxygen, hydrogen peroxide and hydroxyl radicals in plants, which cause damage to proteins, nucleic acids and cell membrane structures [41]. The accumulation of intracellular ROS depends on the balance between various ROS scavenging enzymes and ROS production. Wang [42] observed that maize seedlings showed a significantly enhanced MDA content and cell membrane permeability under chilling stress, which means that the membrane structure of the maize seedlings was damaged, solutes and water are easily lost from the cells, and the water content of the cells is reduced, leading to the young leaves of the seedlings wilting. To reduce the accumulation of ROS and cellular oxidative damage, plants must stimulate their antioxidant defense system to produce antioxidants, for example, enzymes such as SOD, POD, CAT, APX and some nonenzymatic antioxidants, such as ascorbic acid and glutathione [19,43,44]. This study found that the MDA content and antioxidant enzyme activities (SOD, POD and CAT) of the two maize varieties were significantly increased after treatment with chilling stress (Figure 2), which is similar to the results of previous studies.
Photosynthesis is one of the most sensitive plant processes to chilling stress, and low temperature can damage the structure and function of the photosynthetic system [45] and affect the photosynthetic pigments, chloroplast ultrastructure, net photosynthetic rate and gas exchange. Some researchers have found that chilling stress directly affects photosynthesis by disrupting the main components of photosynthesis, such as thylakoid membranes and chlorophyll [46,47]. Chloroplast expansion is the first apparent feature of plants suffering from chilling stress, including the deformation of thylakoids, the decrease in size and number of starch granules, and the formation of small vesicles in chloroplasts. This phenomenon was also observed in this study. However, persistent chilling damage can also cause matrix darkening and chloroplast membrane dissolution, dissolution of the matrix matter and cytoplasm, and inhibit chlorophyll biosynthesis by up to 90% in seedlings [48]. Therefore, under chilling stress, plants exchange gas through stomatal regulation, to alleviate the negative effects on photosynthetic processes [49]. Zhou [50] found that the Pn, gs, and Tr of cucumber seedlings decreased under low temperature (day/night temperature 8 °C/5 °C), while Ci increased, and the same results were obtained in this study. Flexas [51] showed that both Pn and Ci had a strong correlation with gs, whether in the field or in potted plants, but a relationship between gs and leaf water potential was not observed. When the temperature is below an optimum temperature, the absorption of CO2 and Pn drops significantly, or even [52] stops. Therefore, Pn can be invoked as a reference indicator for plant low temperature tolerance.
The optimal level of endogenous hormones in plants is an important indicator of the normal growth and development of plants, and it is necessary to maintain the dynamic balance between the synthesis and decomposition of hormones. IAA has an important regulatory role in plant growth, controlling cell elongation, cell tissue development, and apical dominance [53]. Ribaut [54] showed that under stress conditions, the change in the IAA content is similar to that of ABA, and the degree of plant growth inhibition is also related to the change in IAA, while ABA can increase the activity of antioxidant enzymes and enhance the ROS scavenging ability. Increasing IAA and t-ZR levels mitigate damage from oxidative stress [55,56]. This study found that under chilling stress, the contents of IAA, GA3 and t-ZR decreased, and the content of ABA increased (Figure 4). Therefore, under stress conditions, the reduced plant growth may be the result of changes in the hormonal balance.
4.2. Protective Effects of BL
Soaking in different concentrations of BL can significantly alleviate the inhibitory effect of chilling stress on the germination of two maize varieties, which was also confirmed in Ma Jinhu’s [57] study. In this study, the optimum BL concentration for the low temperature germination of maize was that of T2 (0.1 mg/L), and the low temperature had the least inhibitory effect on maize germination with this treatment. The T3 concentration was better than the T1 concentration. With the T2 concentration, Tiannong 9 germinated 6 days earlier under chilling stress. Tianhe 1 did not undergo germination early, but showed a significantly improved germination rate overall, indicating that Tiannong 9 had a stronger ability to resist low temperature germination than Tianhe 1.
Xu [58] and others have shown that BL can effectively alleviate the decrease in cucumber biomass caused by low temperature and can enhance the ability to resist chilling stress, by regulating the physiological metabolism of cucumber, thereby promoting the accumulation of dry matter [59]. The same conclusions have been corroborated in crops such as maize, rice and wheat [11,60,61,62]. Within the experimental range, the foliar spraying of three concentrations (T1, T2, T3) of BL solution significantly reduced the inhibitory effects maize seedlings caused by low temperature, and the T2 treatment effect was the most significant. Ma Jinhu (Ma 2015) also screened the optimal BL concentration for maize growth under chilling stress in the experimental range. When lower or higher than the T2 concentration, different tissues of different varieties react differently. Compared with the T3 concentration, when the T1 concentration was used, the aboveground parts of Tiannong 9 grew better, while the underground parts grew better at the T3 concentration; the performance of Tianhe 1 was different, both aboveground and underground growth was better with the T3 concentration. This phenomenon indicates that different varieties have different adaptability to chilling stress and BL. For cold-sensitive varieties, it is suitable to spray BL at greater than or equal to the optimum concentration, and for varieties with better cold resistance, it is appropriate to spray BL at less than or equal to the optimum concentration.
Liu [63] reported that BL treatment promoted the activity of antioxidant enzymes in Chorispora bungeana and increased the content of proline and glutathione under chilling stress, thereby eliminating excess ROS. More studies have shown that appropriate concentrations of BL can significantly increase antioxidant enzyme activity and increase antioxidant defense [20,64,65,66]. However, after spraying three concentrations (T1, T2, T3) of BL solution onto two maize hybrids under chilling stress, the MDA content of the seedlings decreased significantly, and the antioxidant enzyme activity increased significantly. The T2 treatment effect was the most significant. This is also consistent with Wu [67], who found that BL spraying can reduce the MDA content of plant leaves and cell membrane peroxidation. In this study, it was also found that T2 increased the activity of CAT more than the other two antioxidant enzymes, which can confirm the viewpoint that CAT is a highly efficient protective enzyme.
The regulation of photosynthesis by the exogenous application of phytohormones is a widely accepted phenomenon [68]. Huang [69] showed that chilling stress caused a decrease in the chlorophyll content and Pn of the herbaceous plant Dendrobium officinale, resulting in a decrease in maximum photochemical efficiency, which was significantly reduced by exogenous salicylic acid (SA) treatment. Chilling stress can significantly reduce maize Pn, and BLs can increase Pn under stress conditions, by increasing the chlorophyll content [70]; Fariduddin [59] used 28-homobrassinolide (HBL) to treat cucumber seedlings and found that the Pn, gs and Tr were significantly increased under low temperature and normal temperature conditions. This study showed that the application of exogenous BL at different concentrations could significantly alleviate the inhibitory effect of chilling stress on the Pn of maize seedlings, increase the gs and Tr, and significantly reduce the Ci. Among the treatments, T2 had the best effect, and there were differences between the different varieties, with various degrees of cold resistance. This is similar to previous studies [59,62]. In this study, it was found that the photosynthesis-related indicators of the maize variety with stronger cold resistance were less changed. Ashraf [49] also showed that in maize, varieties with a better cold tolerance during the low temperature recovery phase grow faster. This indicates that the cold-resistant maize varieties experience relatively mild temperature damage under chilling stress, and the plants have strong recovery abilities; thus, such maize varieties can be used as raw materials for cold-resistance gene mining.
Exogenous application of BL can effectively alleviate the changes in hormone content in maize seedlings under chilling stress, and the T2 treatment was the best. At the same time, it was found that the changes in the hormone contents of the two maize varieties were significantly different, and the change range of IAA was similar to that of ABA, which confirmed the conclusions of previous studies. In addition, this study found that the ABA content in Tiannong 9 plants was much higher than that in Tianhe 1 plants. At the same time, this finding was consistent with the conclusion that Wang [71] and others found, that ABA was accumulated in large amounts in plants with strong cold tolerance. There are three basic relationships among various hormones: the synchronism effect, synergistic effect and antagonistic effect. At the same time, the relationship between these hormones is closely related to the stress resistance of plants, and several different hormones in plants also have sequential regulatory and chain reactions under stress. The changes of four endogenous hormones after BL application in this study indicated that when the maize seedlings were subjected to chilling stress, various hormones regulate the stomatal movement of plant cells, reduce water loss via transpiration, and drive various hormones to affect each other and reconstruct a new dynamic balance to regulate cold resistance.
Chloroplasts are considered to be the principal site of photosynthesis, because photoreactions are only carried out in chloroplasts [49]. In this study, the ultrastructures of the chloroplasts of the two maize varieties changed significantly under chilling stress. Chloroplast swelling was obvious in the T0 treatment, the loose rupture of the chloroplast membrane gradually disintegrated, the osmiophilic granules increased, and the structure became disordered. Xu [72] et al. observed that chloroplasts and some thylakoids swelled in cucumber leaves under chilling stress and formed many vesicles formed by thin slices. This feature was also observed in this study. From this, we conclude that chilling stress has a negative effect on photosynthesis in maize leaves. At the same time, it was observed the T2 treatment significantly alleviated this low temperature damage, the chloroplast maintained their original elliptical structure, the bilayer membrane structure was clear, and only some of the plasma membranes had fractures. This result indicated that treatment with exogenous BL could significantly attenuate the dissolution of the chloroplast plasma membrane and the deformation of the lamellar structure under chilling stress. The same research results were confirmed in other stress environments [18,73].
5. Conclusions
In conclusion, chilling stress significantly inhibited the germination and seedling growth of different maize varieties. Spraying BL at low temperature could alleviate the damage to maize under chilling stress. Different concentrations of BL had significant effects on the germination rate, agronomic traits, photosynthetic system, antioxidant system and endogenous hormone levels in maize, and the alleviation of chilling effects in maize seedlings was best with BL application at 0.1 mg/L. Most notably, under chilling stress, the germination rate and biomass of maize increased significantly. The SOD activity, POD activity, Pn, gs and ABA content of seedlings increased markedly, and the contents of MDA, Ci and IAA decreased significantly (Figure 6). These data show that the application of BL can alleviate the damage caused by chilling stress to varying degrees. The focus of this paper is mainly on the effects of exogenous BL on the cold resistance of maize under chilling stress. Whether the signal transduction of BL under abiotic stress is linked to its role in plant immunity and whether the application of exogenous BL will regulate the expression of cold resistance genes will be further explored in future studies.
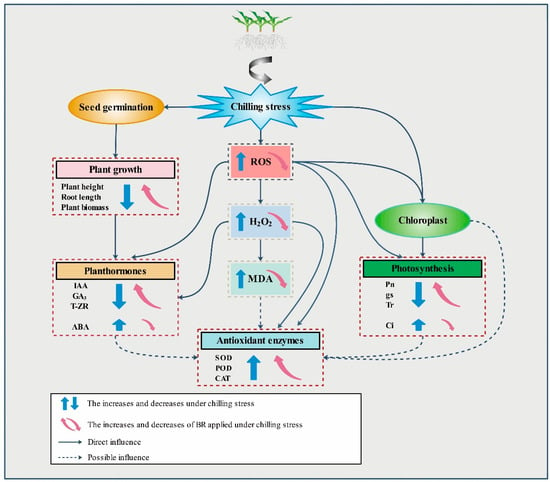
Figure 6.
Probable model for BL mode of action in the maize response to chilling stress.
Author Contributions
D.Y. designed the research and provided the experimental equipment and materials; Q.Z. conceived the experiments and edited the paper; Y.S. completed the experiment and analyzed the data to write the article. All the authors reviewed the article. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Key Research and Development (R&D) Program of China (2018YFD0300103), Agricultural research outstanding talents and innovation team (2015), the University Nursing Program for Young Scholars with Creative Talents in Heilongjiang Province (UNPYSCT-2017012) and Young Talents of Northeast Agricultural University (17QC01).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Casadei, E.; Albert, J. Food and Agriculture Organization of the United Nations. Encycl. Food Sci. Nutr. 2003, 1, 2587–2593. [Google Scholar]
- Krasensky-Wrzaczek, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [PubMed]
- Leipner, J.; Stamp, P. Chilling Stress in Maize Seedlings. In Handbook of Maize: Its Biology; Jeff, L.B., Sarah, C.H., Eds.; Springer: New York, NY, USA, 2009; pp. 291–310. [Google Scholar]
- Atıcı, Ö.; Nalbantoǧlu, B.; Atici, O.; Nalbantoglu, B. Antifreeze proteins in higher plants. Phytochem. 2003, 64, 1187–1196. [Google Scholar] [CrossRef]
- Miedema, P. The Effects of Low Temperature on Zea mays. Advances in Agronomy 1982, 35, 93–128. [Google Scholar]
- Janowiak, F.; Maas, B.; Dörffling, K. Importance of abscisic acid for chilling tolerance of maize seedlings. J. Plant Physiol. 2002, 159, 635–643. [Google Scholar] [CrossRef]
- Bajguz, A.; Hayat, S. Effects of brassinosteroids on the plant responses to environmental stresses. Plant Physiol. Biochem. 2009, 47, 1–8. [Google Scholar] [CrossRef]
- Louarn, G.; Chenu, K.; Fournier, C.; Andrieu, B.; Giauffret, C. Relative contributions of light interception and radiation use efficiency to the reduction of maize productivity under cold temperatures. Funct. Plant Boil. 2008, 35, 885–899. [Google Scholar] [CrossRef]
- Krishna, P. Brassinosteroid-Mediated Stress Responses. J. Plant Growth Regul. 2003, 22, 289–297. [Google Scholar] [CrossRef]
- Hamada, K. Brassinolide: Some Effects for Crop Cultivations. In Proceedings of the Conference Proceedings International Seminar Plant Growth Regulation, Tokyo, Japan, 15 October 1986; pp. 188–196. [Google Scholar]
- Fariduddin, Q.; Yusuf, M.; Ahmad, I.; Ahmad, A. Brassinosteroids and their role in response of plants to abiotic stresses. Boil. Plant. 2013, 58, 9–17. [Google Scholar] [CrossRef]
- Vardhini, B.V. Modifications of morphological and anatomical characteristics of plants by application of brassinosteroids under various abiotic stress conditions—A review. Plant Gene 2017, 11, 70–89. [Google Scholar] [CrossRef]
- Yu, J.; Zhou, Y.; Ye, S.F.; Huang, L.F. 24-epibrassinolide and abscisic acid protect cucumber seedlings from chilling injury. J. Hortic. Sci. Biotechnol. 2002, 77, 470–473. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Zhu, X.H.; Ding, H.D.; Yang, S.J.; Chen, Y.Y. Foliar application of 24-epibrassinolide alleviates high-temperature-induced inhibition of photosynthesis in seedlings of two melon cultivars. Photosynthetica 2013, 51, 341–349. [Google Scholar] [CrossRef]
- Tanveer, M.; Shahzad, B.; Sharma, A.; Khan, E.A. 24-Epibrassinolide application in plants: An implication for improving drought stress tolerance in plants. Plant Physiol. Biochem. 2019, 135, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.-D.; Zhu, X.-H.; Zhu, Z.-W.; Yang, S.-J.; Zha, D.-S.; Wu, X.-X. Amelioration of salt-induced oxidative stress in eggplant by application of 24-epibrassinolide. Boil. Plant. 2012, 56, 767–770. [Google Scholar] [CrossRef]
- Bajguz, A. An enhancing effect of exogenous brassinolide on the growth and antioxidant activity in Chlorella vulgaris cultures under heavy metals stress. Environ. Exp. Bot. 2010, 68, 175–179. [Google Scholar] [CrossRef]
- Yuan, L.; Shu, S.; Sun, J.; Guo, S.; Tezuka, T. Effects of 24-epibrassinolide on the photosynthetic characteristics, antioxidant system, and chloroplast ultrastructure in Cucumis sativus L. under Ca(NO3)2 stress. Photosynth. Res. 2012, 112, 205–214. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Yuan, H.-L.; Ogweno, J.O.; Zhou, Y.; Xia, X.; Mao, W.-H.; Shi, K.; Yu, J. Brassinosteroid alleviates phenanthrene and pyrene phytotoxicity by increasing detoxification activity and photosynthesis in tomato. Chemosphere 2012, 86, 546–555. [Google Scholar] [CrossRef]
- Xi, Z.; Wang, Z.; Fang, Y.; Hu, Z.; Hu, Y.; Deng, M.; Zhang, Z.-W. Effects of 24-epibrassinolide on antioxidation defense and osmoregulation systems of young grapevines (V. vinifera L.) under chilling stress. Plant Growth Regul. 2013, 71, 57–65. [Google Scholar] [CrossRef]
- Hu, W.H.; Wu, Y.; Zeng, J.Z.; He, L.; Zeng, Q.M. Chill-induced inhibition of photosynthesis was alleviated by 24-epibrassinolide pretreatment in cucumber during chilling and subsequent recovery. Photosynthetica 2010, 48, 537–544. [Google Scholar] [CrossRef]
- Yang, D.; Sun, Y.; Irfan, A.R.; Liu, X.; Lv, J.; Yu, J.; Gong, L.; Liu, Z.; Bai, B. Effect of Low Temperature Stress on Germination and Physiological of Maize Seedling. J. Northeast Agric. Univ. 2018, 49, 4–11. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases: I. Occurrence in Higher Plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Li, F.; Xu, T.; Cai, S.; Chu, W.; Qiu, H.; Sha, S.; Cheng, G.; Xu, Q. Bioaccumulation, subcellular, and molecular localization and damage to physiology and ultrastructure in Nymphoides peltata (Gmel.) O. Kuntze exposed to yttrium. Environ. Sci. Pollut. Res. 2013, 21, 2935–2942. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzym. 1984, 105, 121–126. [Google Scholar]
- Zhang, Y.; Li, Q.; Jiang, L.; Kai, W.; Liang, B.; Wang, J.; Du, Y.; Zhai, X.; Wang, J.; Zhang, Y.; et al. Suppressing Type 2C Protein Phosphatases Alters Fruit Ripening and the Stress Response in Tomato. Plant Cell Physiol. 2017, 59, 142–154. [Google Scholar] [CrossRef]
- Su, Q.; Lai, Z.; Ye, L.; Wu, S. The Hplc Detection of Endogrnous Hormines from Seedling in Several Species of Acacia. Chin. Agric. Sci. Bull. 2010, 26, 216–221. [Google Scholar]
- Cutforth, H.W.; Shaykewich, C.F.; Cho, C.M. Effect of Soil Water and Temperature on Corn (Zea mays L.) Root Growth during Emergence. Can. J. Soil Sci. 1986, 66, 51–58. [Google Scholar] [CrossRef]
- Muñoz, B.G.; Lekfeldt, J.D.S.; Magid, J.; Jensen, L.S.; De Neergaard, A. Seed treatment with Penicillium sp. or Mn/Zn can alleviate the negative effects of cold stress in maize grown in soils dependent on soil fertility. J. Agron. Crop. Sci. 2018, 204, 603–612. [Google Scholar] [CrossRef]
- Bano, S.; Aslam, M.; Saleem, M.; Basra, S.M.A.; Aziz, K. Evaluation of Maize Accessions under Low Temperature Stress at Early Growth Stages. J. Anim. Plant Sci. 2015, 25, 392–400. [Google Scholar]
- Huang, J.; Zhang, J.; Li, W.; Hu, W.; Duan, L.; Feng, Y.; Qiu, F.; Yue, B. Genome-wide Association Analysis of Ten Chilling Tolerance Indices at the Germination and Seedling Stages in Maize. J. Integr. Plant Boil. 2013, 55, 735–744. [Google Scholar] [CrossRef]
- Sánchez, B.; Rasmussen, A.; Porter, J.R. Temperatures and the growth and development of maize and rice: A review. Glob. Chang. Boil. 2013, 20, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Saeidnejad, A.H.; Pouramir, F.; NaghiZadeh, M. Improving Chilling Tolerance of Maize Seedlings under Cold Conditions by Spermine Application. Not. Sci. Boil. 2012, 4, 110–117. [Google Scholar] [CrossRef][Green Version]
- LIU, Y.C.; YANG, D.G.; LI, L.; CHAI, M.Z.; JU, R.; BAI, B.; LIU, Y.L. Effect of Low Temperature Stress on Maize Seed Germination and Starch Decomposition Enzyme Activity. J. Maize Sci. 2018, 26, 64–68. [Google Scholar]
- Engels, C. Effect of Root and Shoot Meristem Temperature on Shoot to Root Dry Matter Partitioning and the Internal Concentrations of Nitrogen and Carbohydrates in Maize and Wheat. Ann. Bot. 1994, 73, 211–219. [Google Scholar] [CrossRef]
- Stone, P.; Sorensen, I.; Jamieson, P. Effect of soil temperature on phenology, canopy development, biomass and yield of maize in a cool-temperate climate. Field Crop. Res. 1999, 63, 169–178. [Google Scholar] [CrossRef]
- Sowiński, P.; Rudzińska-Langwald, A.; Adamczyk, J.; Kubica, I.; Fronk, J. Recovery of maize seedling growth, development and photosynthetic efficiency after initial growth at low temperature. J. Plant Physiol. 2005, 162, 67–80. [Google Scholar] [CrossRef]
- Rymen, B.; Fiorani, F.; Kartal, F.; Vandepoele, K.; Inzé, D.; Beemster, G.T.S. Cold Nights Impair Leaf Growth and Cell Cycle Progression in Maize through Transcriptional Changes of Cell Cycle Genes1[W][OA]. Plant Physiol. 2007, 143, 1429–1438. [Google Scholar] [CrossRef]
- Mauch-Mani, B.; Mauch, F. The role of abscisic acid in plant–pathogen interactions. Curr. Opin. Plant Boil. 2005, 8, 409–414. [Google Scholar] [CrossRef]
- Wise, R.R.; Naylor, A.W. Chilling-Enhanced Photooxidation: The Peroxidative Destruction of Lipids During Chilling Injury to Photosynthesis and Ultrastructure. Plant Physiol. 1987, 83, 272–277. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, M.; Gong, X.; Liu, C.; Hong, M.; Wang, L.; Hong, F. Influence of Lanthanides on the Antioxidative Defense System in Maize Seedlings Under Cold Stress. Boil. Trace Element Res. 2010, 142, 819–830. [Google Scholar] [CrossRef]
- Gapper, C.; Dolan, L. Control of Plant Development by Reactive Oxygen Species1. Plant Physiol. 2006, 141, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Guo, Z.; Xing, J.; Huang, B. Nitric oxide is involved in abscisic acid-induced antioxidant activities in Stylosanthes guianensis. J. Exp. Bot. 2005, 56, 3223–3228. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic Response of Plants Under Different Abiotic Stresses: A Review. J. Plant Growth Regul. 2019, 1–23. [Google Scholar] [CrossRef]
- Holá, D.; Kočová, M.; Rothová, O.; Wilhelmova, N.; Benesova, M. Recovery of maize (Zea mays L.) inbreds and hybrids from chilling stress of various duration: Photosynthesis and antioxidant enzymes. J. Plant Physiol. 2007, 164, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Huang, L.; Zhang, Y.; Shi, K.; Yu, J.; Nogués, S. Chill-Induced Decrease in Capacity of RuBP Carboxylation and Associated H2O2 Accumulation in Cucumber Leaves are Alleviated by Grafting onto Figleaf Gourd. Ann. Bot. 2007, 100, 839–848. [Google Scholar] [CrossRef]
- Yun, J.G.; Hayashi, T.; Yazawa, S.; Katoh, T.; Yasuda, Y. Acute morphological changes of palisade cells ofSaintpaulia leaves induced by a rapid temperature drop. J. Plant Res. 1996, 109, 339–342. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Zhou, C.F.; Wu, C.T.; Li, D.D.; Zhang, X.W.; Bi, H.G.; Ai, X.Z. Hydrogen Sulfide Promotes Chilling Tolerance of Cucumber Seedlings by Alleviating Low-Temperature Photoinhibition. Plant Physiol. J. 2018, 54, 411–420. [Google Scholar]
- Flexas, J.; Bota, J.; Escalona, J.M.; Sampol, B.; Medrano, H. Effects of drought on photosynthesis in grapevines under field conditions: An evaluation of stomatal and mesophyll limitations. Funct. Plant Boil. 2002, 29, 461. [Google Scholar] [CrossRef]
- Aniszewski, T.; Drozdov, S.N.; Kholoptseva, E.S.; Kurets, V.K.; Obshatko, L.A.; Popov, E.G.; Talanov, A.V. Effects of Light and Temperature Parameters on Net Photosynthetic Carbon Dioxide Fixation by Whole Plants of Five Lupin Species ( Lupinus albus L.; Lupinus angustifolius L.; Lupinus luteus L.; Lupinus mutabilis Sweet. and Lupinus polyphyllus Lindl.). Acta Agric. Scand. Sect. B - Plant Soil Sci. 2001, 51, 17–27. [Google Scholar] [CrossRef]
- Wang, Y.; Mopper, S.; Hasenstein, K. Effects of salinity on endogenous ABA, IAA, JA, AND SA in Iris hexagona. J. Chem. Ecol. 2001, 27, 327–342. [Google Scholar] [CrossRef] [PubMed]
- Ribaut, J.M.; Pilet, P.E. Water stress and indol-3yl-acetic acid content of maize roots. Planta 1994, 193, 502–507. [Google Scholar] [CrossRef]
- Wang, F.; Tan, X.H.; Gan, L.; Wu, Y.Y.; Wang, R.Z.; & Li, Q.M. Effects of Cold Shock Treatments on Endogenous Phytohormones Contents and Proteins Variations of Nai-Plume Fruits. Food Sci. 2006, 27, 71–74. [Google Scholar]
- Lin, Y. Influence of Natural Plant Growth Regulator-Oxyenadenine against Growth and Increase Production with Tomato. AGROCHEMICALS 2007, 46, 349–350. [Google Scholar]
- Ma, J. Physiolgical Mechanism of Ebr Promotr Low Temperature Stress in Maize Seed Germination. PH.D. Thesis, Shanxi Agricultural University, Jinzhong, Shanxi, China, December 2015. [Google Scholar]
- Xu, X.Y.; Yu, J.H.; Xie, J.M.; Hu, L.L.; Li, J. Effects of Exogenous Salicylic Acid and Brassinolide on Photosynthesis of Cucumber Seedlings under Low Temperature Stress. Chin. J. Appl. Ecol. 2016, 27, 3009–3015. [Google Scholar]
- Fariduddin, Q.; Yusuf, M.; Chalkoo, S.; Hayat, S.; Ahmad, A. 28-homobrassinolide improves growth and photosynthesis in Cucumis sativus L. through an enhanced antioxidant system in the presence of chilling stress. Photosynthetica 2011, 49, 55–64. [Google Scholar] [CrossRef]
- Fujii, S.; Saka, H. The Promotive Effect of Brassinolide on Lamina Joint-Cell Elongation, Germination and Seedling Growth under Low-Temperature Stress in Rice (Oryza sativa L.). Plant Prod. Sci. 2001, 4, 210–214. [Google Scholar] [CrossRef]
- Shahbaz, M.; Ashraf, M.; Athar, H. Does exogenous application of 24-epibrassinolide ameliorate salt induced growth inhibition in wheat (Triticum aestivum L.)? Plant Growth Regul. 2008, 55, 51–64. [Google Scholar] [CrossRef]
- Singh, I.; Kumar, U.; Singh, S.K.; Gupta, C.; Singh, M.; Kushwaha, S.R. Physiological and biochemical effect of 24-epibrassinoslide on cold tolerance in maize seedlings. Physiol. Mol. Boil. Plants 2012, 18, 229–236. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, Z.; Si, J.; Di, C.; Han, J.; An, L. Brassinosteroids alleviate chilling-induced oxidative damage by enhancing antioxidant defense system in suspension cultured cells of Chorispora bungeana. Plant Growth Regul. 2009, 59, 207–214. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Bhardwaj, R.; Zheng, B. Phytohormones Regulate Accumulation of Osmolytes Under Abiotic Stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.-H.; Yan, X.-H.; Xiao, Y.-A.; Zeng, J.-J.; Qi, H.-J.; Ogweno, J.O. 24-Epibrassinosteroid alleviate drought-induced inhibition of photosynthesis in Capsicum annuum. Sci. Hortic. 2013, 150, 232–237. [Google Scholar] [CrossRef]
- Li, X.; Wei, J.-P.; Ahammed, G.J.; Zhang, L.; Li, Y.; Yan, P.; Zhang, L.-P.; Han, W.-Y. Brassinosteroids Attenuate Moderate High Temperature-Caused Decline in Tea Quality by Enhancing Theanine Biosynthesis in Camellia sinensis L. Front. Plant Sci. 2018, 9, 1016. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.X.; Ding, H.D.; Chen, J.L.; Zhu, Z.; Zha, D.S. Exogenous spray application of 24-epibrassinolide induced changes in photosynthesis and anti-oxidant defences against chilling stress in eggplant ( Solanum melongena L.) seedlings. J. Hortic. Sci. Biotechnol. 2015, 90, 217–225. [Google Scholar] [CrossRef]
- Holá, D. Brassinosteroids and Photosynthesis. In Brassinosteroids: A Class of Plant Hormone; Hayat, S., Ahmad, A., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 143–192. [Google Scholar]
- Huang, C.; Wang, D.; Sun, L.; Wei, L. Effects of exogenous salicylic acid on the physiological characteristics of Dendrobium officinale under chilling stress. Plant Growth Regul. 2016, 79, 199–208. [Google Scholar] [CrossRef]
- Alyemeni, M.; Al-Quwaiz, S.M. Effect of 28-homobrassinolide on the performance of sensitive and resistant varieties of Vigna radiata. Saudi J. Boil. Sci. 2016, 23, 698–705. [Google Scholar] [CrossRef]
- WANG, X.; YU, J.; YANG, Y.; CANG, J.; LI, Z.F. Changes of Endogenous Hormones of Winter Wheat Varieties with Different Cold-Resistances under Low Temperature. J. Triticeae Crops 2009, 5, 827–831. [Google Scholar]
- Xu, P.-L.; Guo, Y.-K.; Bai, J.-G.; Shang, L.; Wang, X.-J. Effects of long-term chilling on ultrastructure and antioxidant activity in leaves of two cucumber cultivars under low light. Physiol. Plant. 2008, 132, 467–478. [Google Scholar] [CrossRef]
- Agami, R.A. Alleviating the adverse effects of NaCl stress in maize seedlings by pretreating seeds with salicylic acid and 24-epibrassinolide. S. Afr. J. Bot. 2013, 88, 171–177. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).