Manipulation of Fruit Dry Matter via Seasonal Pruning and Its Relationship to d’Anjou Pear Yield and Fruit Quality




Abstract
:1. Introduction
2. Materials and Methods
2.1. Pruning and Vegetative Growth
2.2. Flowering, Fruit Set, and Yield
2.3. Fruit Size and Dry Matter Distribution
2.4. Fruit Quality
2.5. Statistical Methods
3. Results
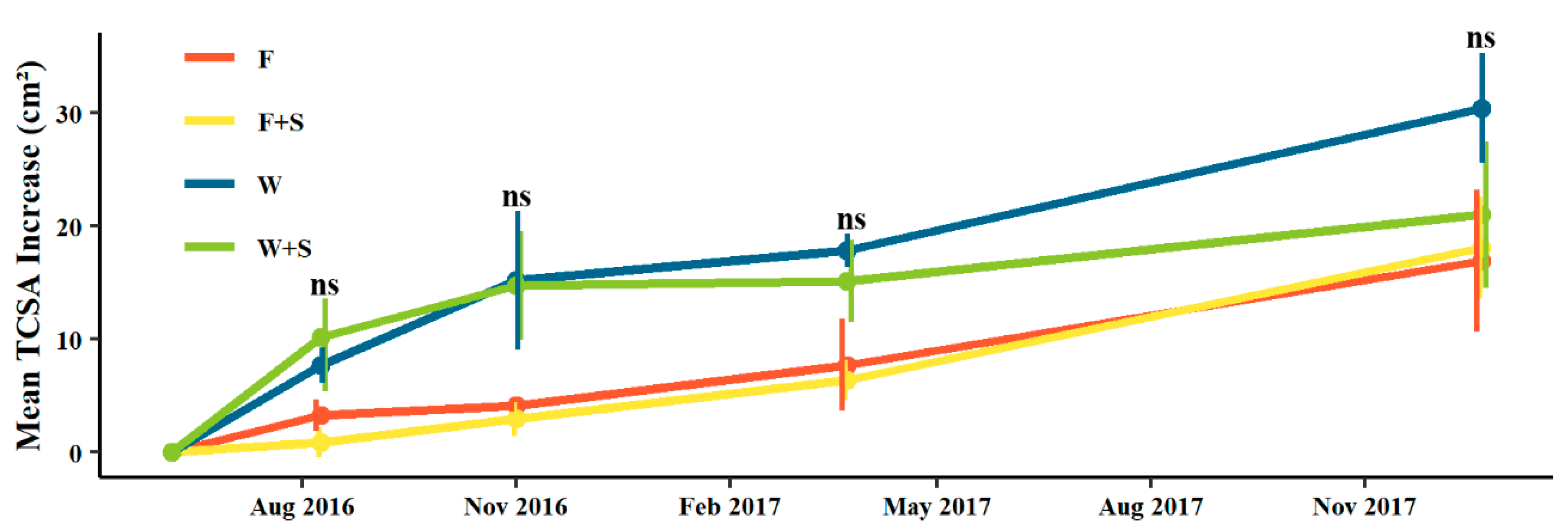
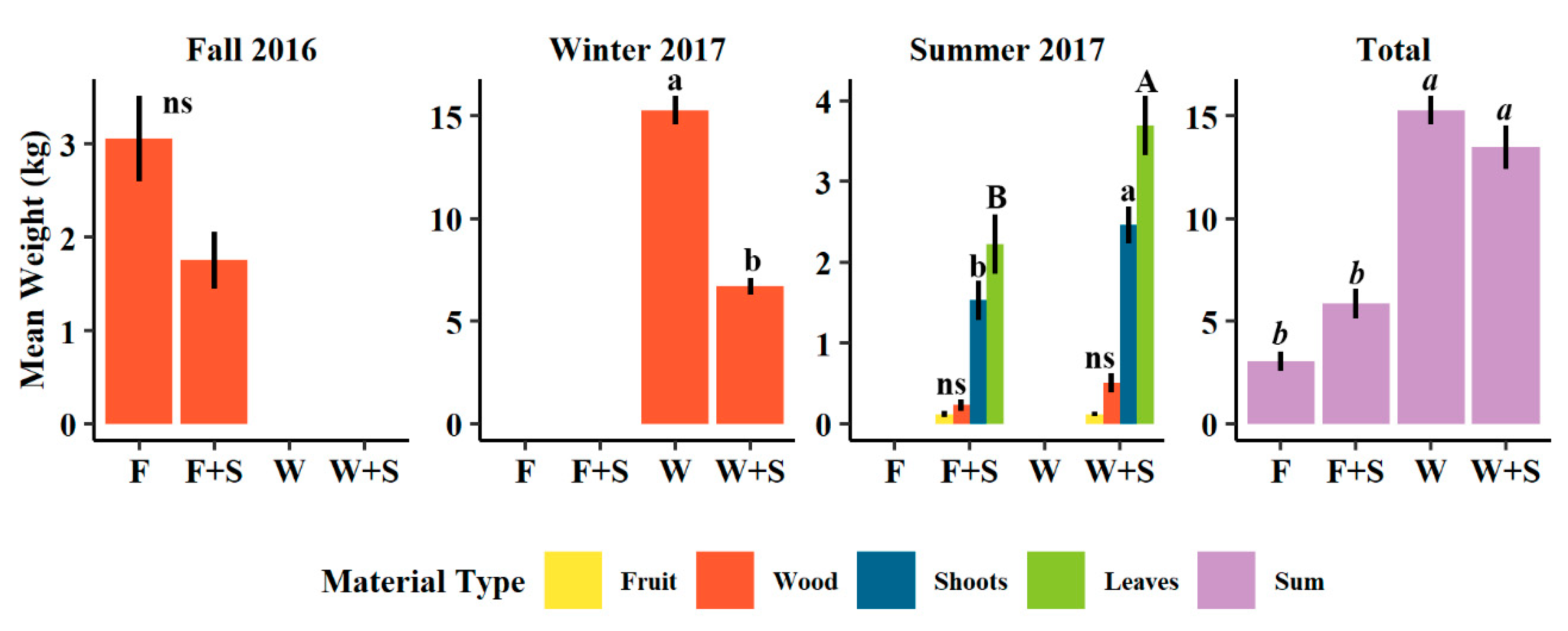
3.1. Pruning and Vegetative Growth
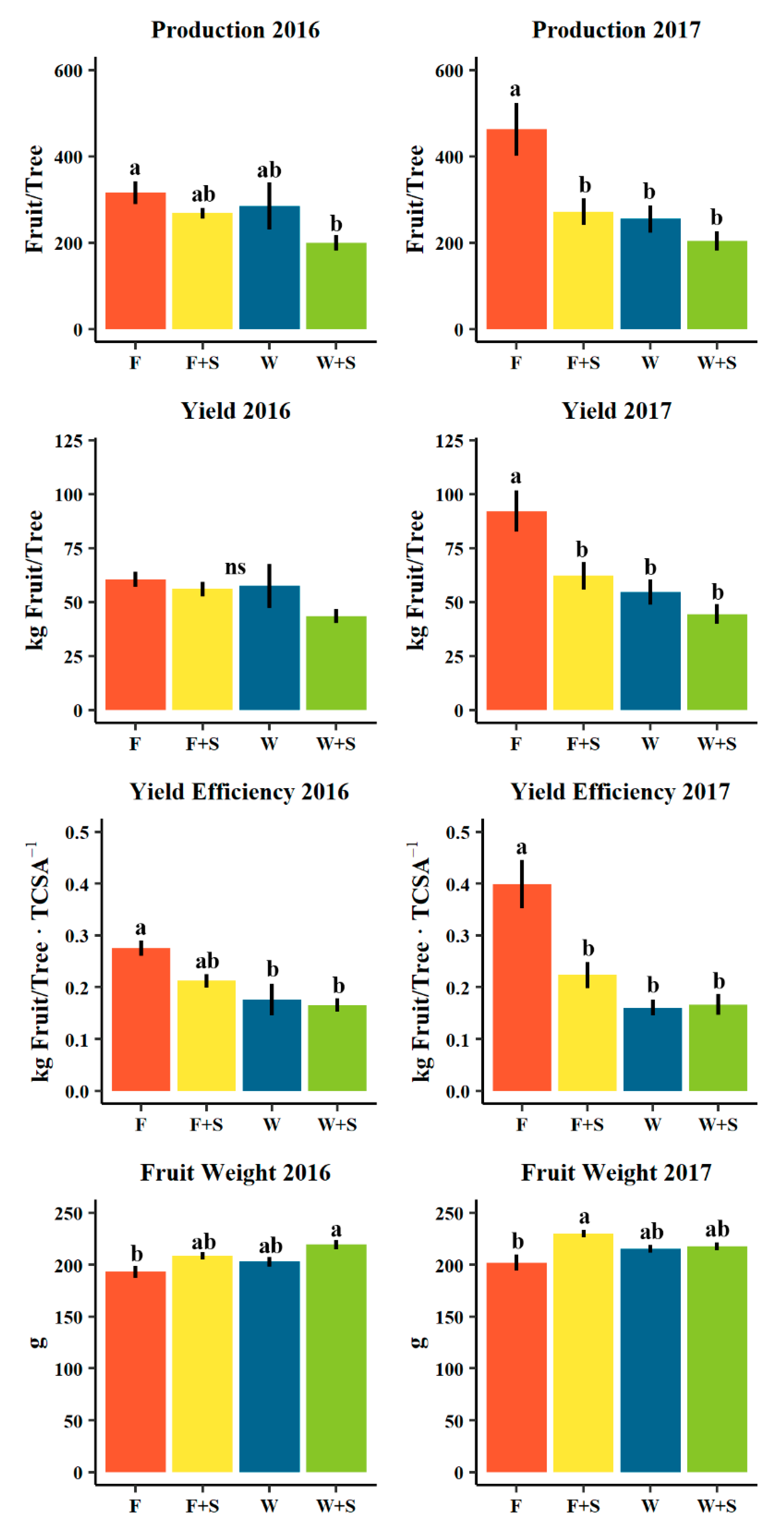
3.2. Flowering, Fruit Set, and Yield
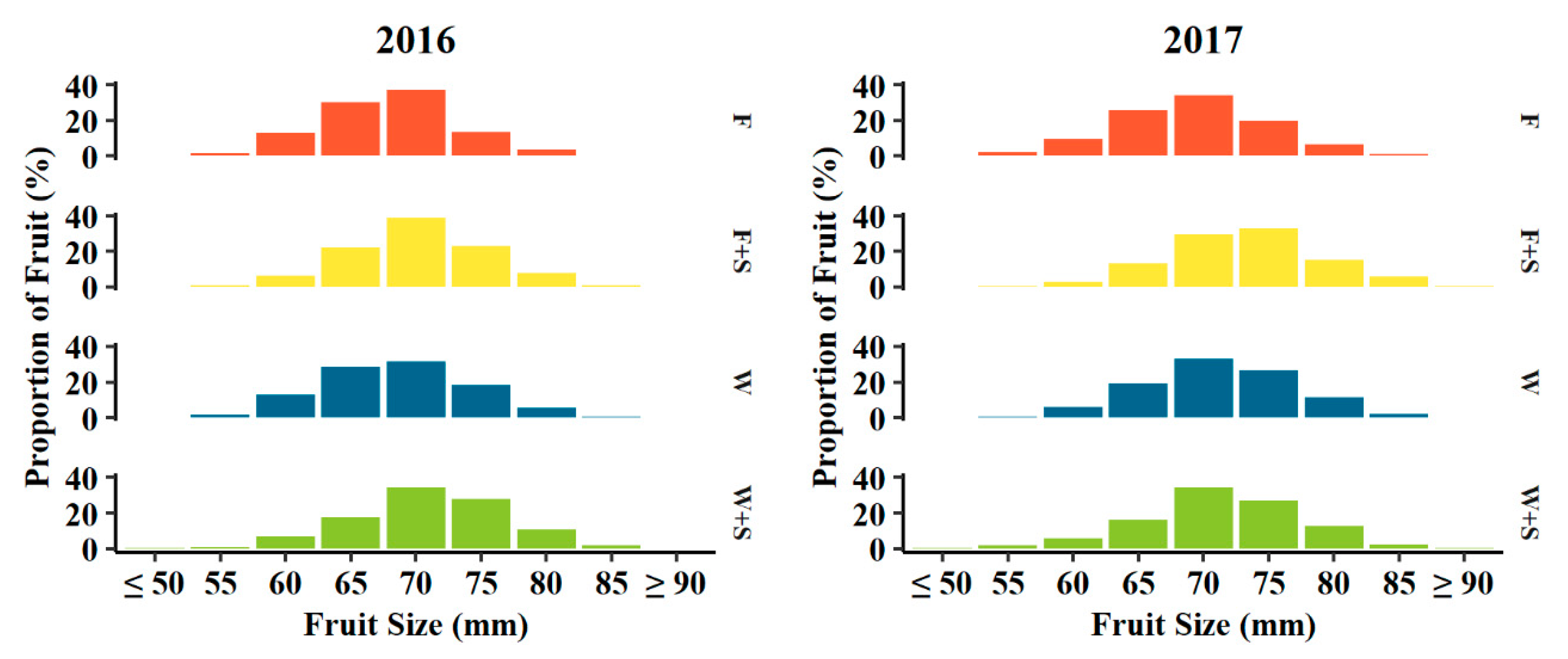
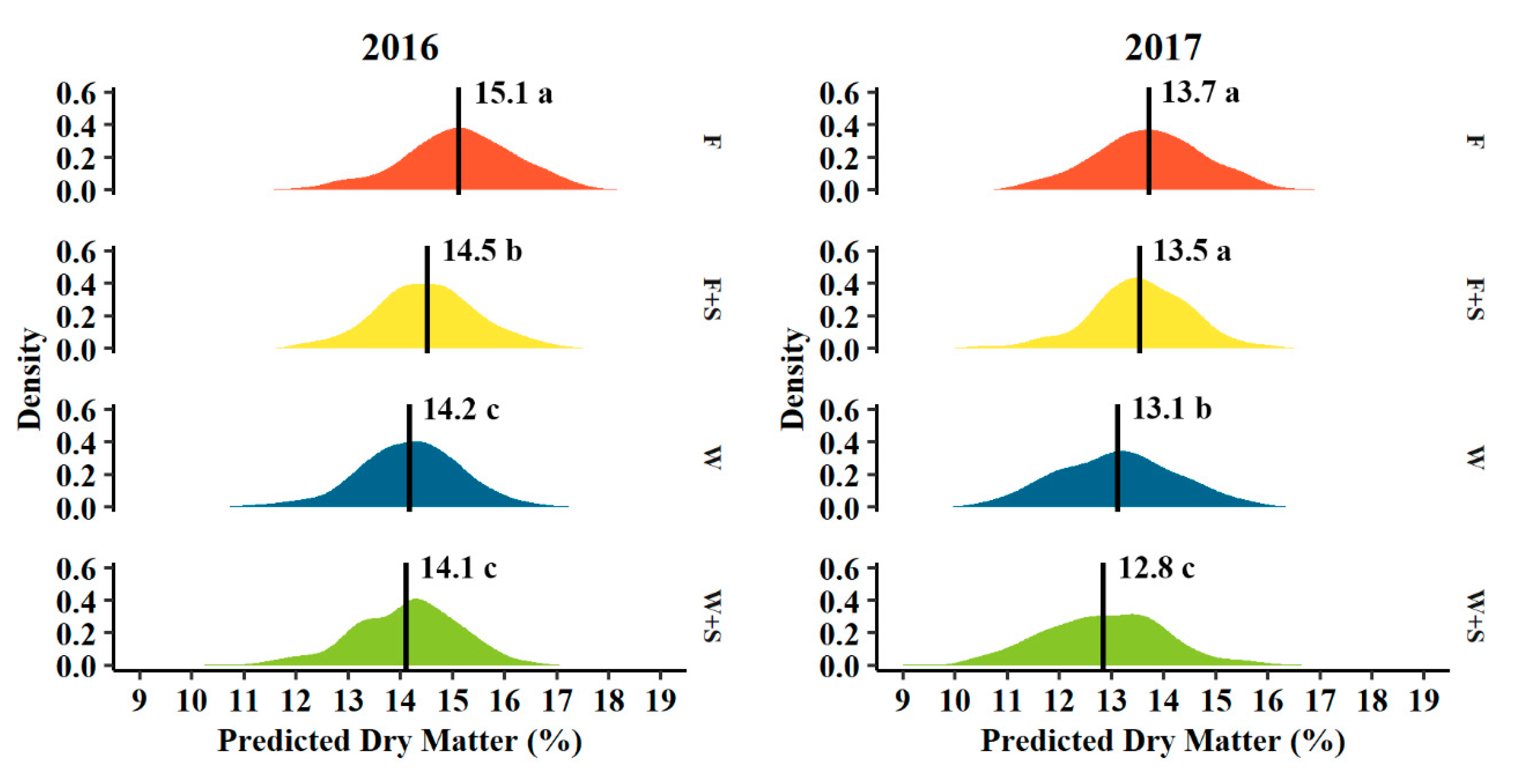
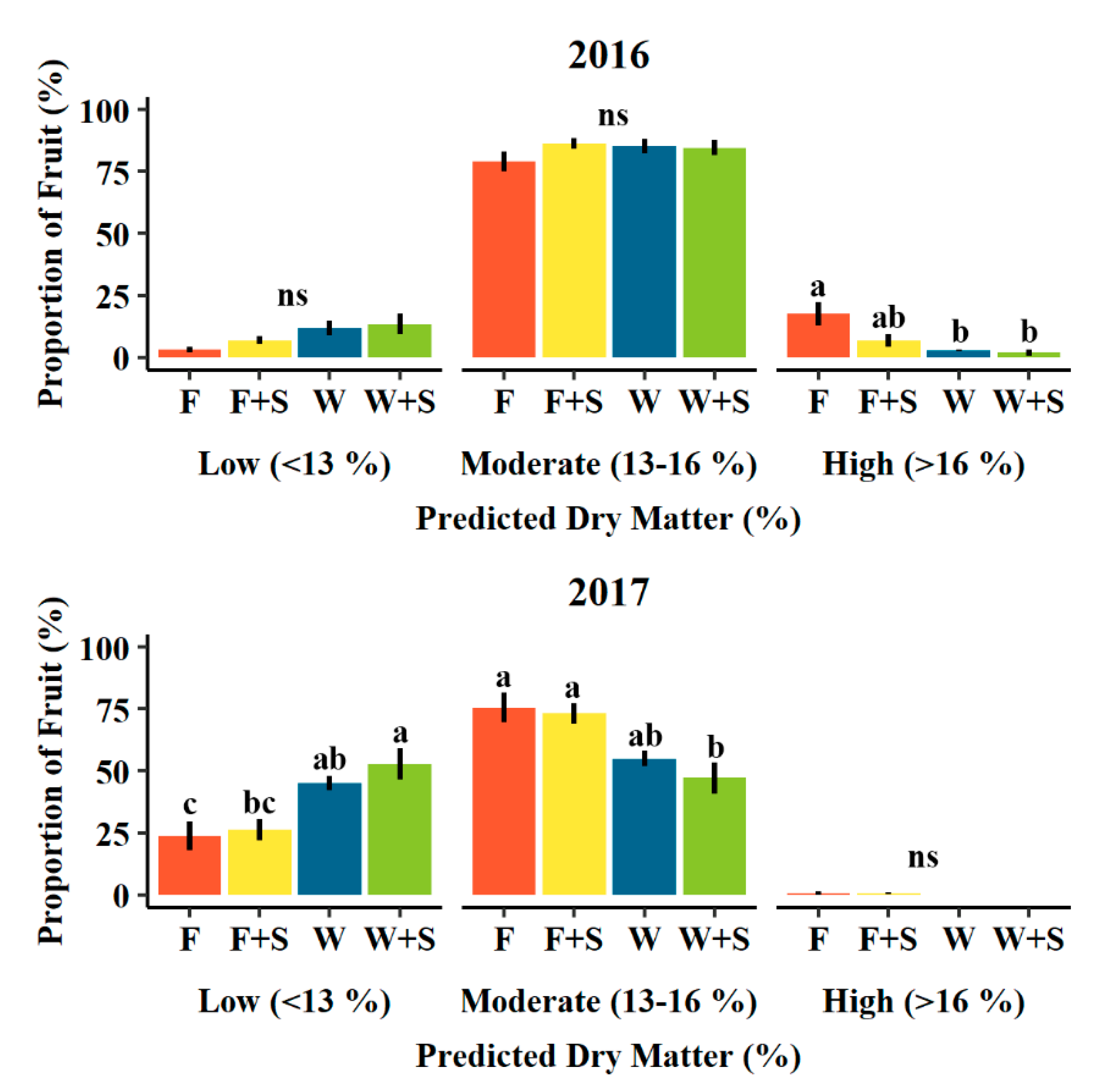
3.3. Fruit Size and Dry Matter Distribution
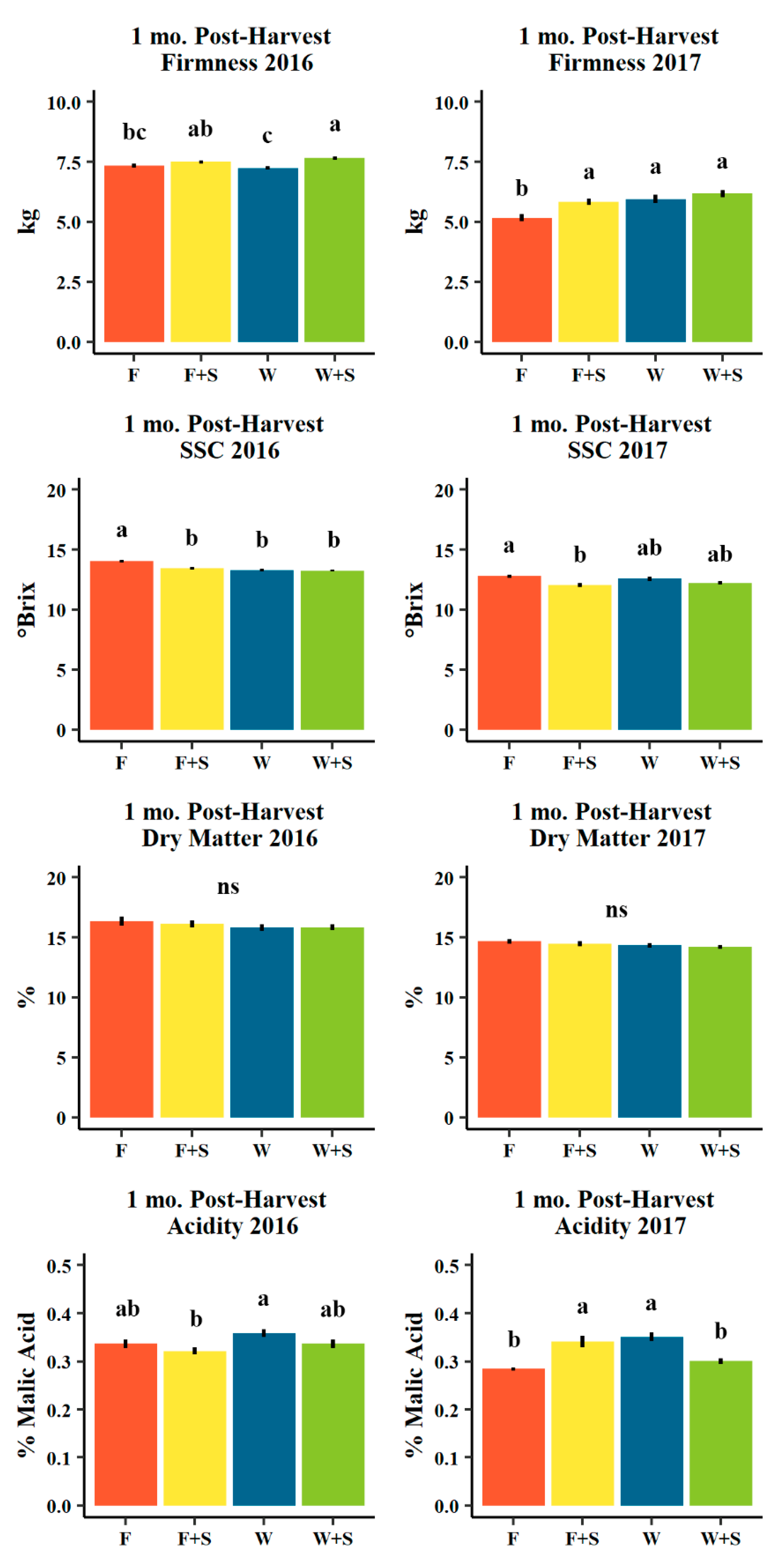
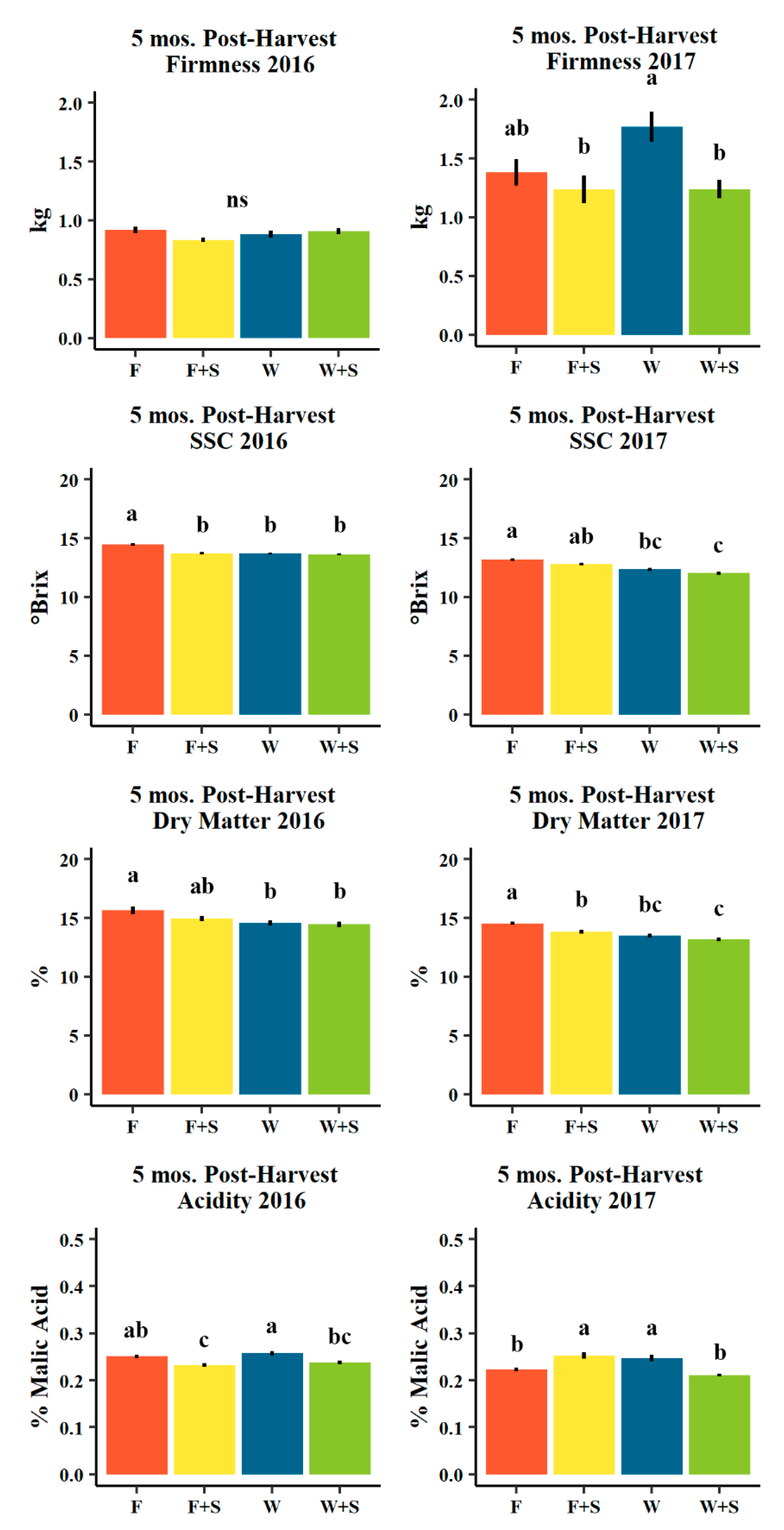
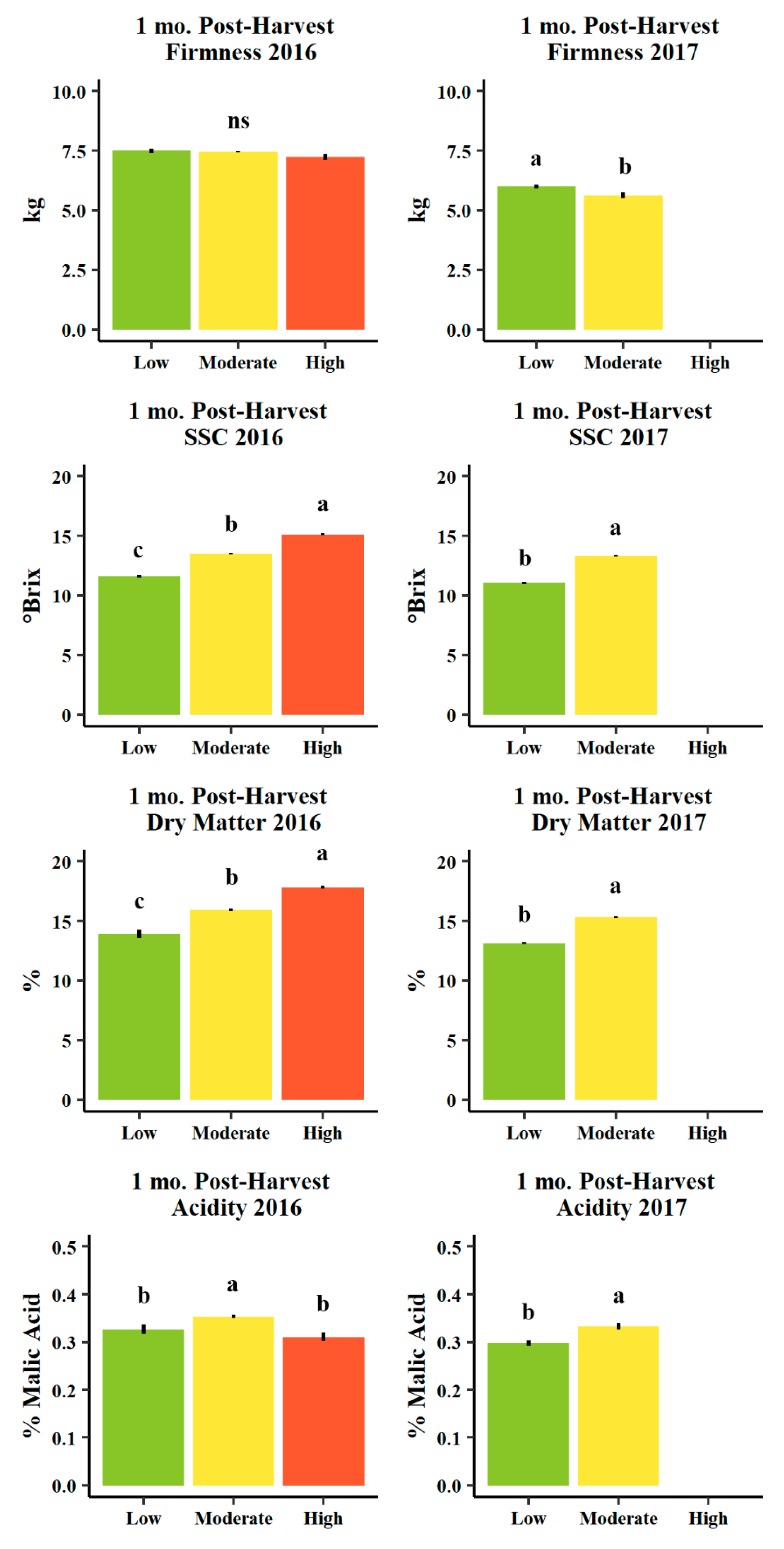
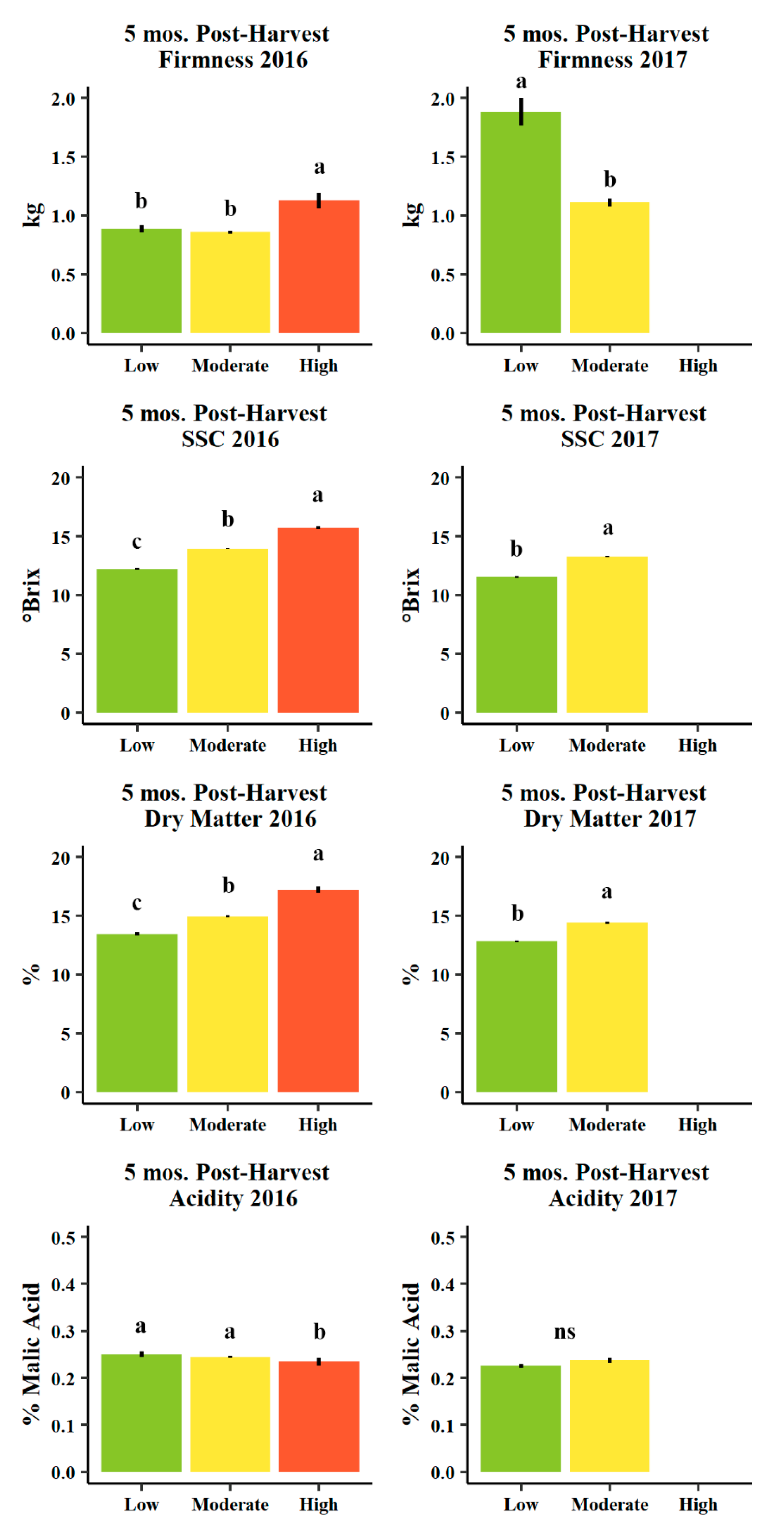
3.4. Fruit Quality
4. Discussion
4.1. Vegetative Growth
4.2. Flowering, Fruit Set, and Yield
4.3. Fruit Size and Dry Matter
4.4. Fruit Quality
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Einhorn, T.C.; Turner, J.; Gibeaut, D.; Postman, J.D. Characterization of Cold Hardiness in Quince: Potential Pear Rootstock Candidates for Northern Pear Production Regions. Acta Hortic. 2011, 909, 137–143. [Google Scholar] [CrossRef]
- Elkins, R.; Bell, R.; Einhorn, T. Needs assessment for future US pear rootstock research directions based on the current state of pear production and rootstock research. J. Am. Pomol. Soc. 2012, 66, 153–163. [Google Scholar]
- Wertheim, S.J. Rootstocks for European pear: A review. Acta Hortic. 2000, 596, 299–309. [Google Scholar] [CrossRef]
- Sansavini, S.; Castagnoli, M.; Musacchi, S. New rootstocks for William and Abate Fetel cultivars of pear tree. Comparison between progenies of quince tree and clonal stocks [Pyrus communis]. Riv. Frutticolt. Ortofloric. 1997, 59, 31–40. [Google Scholar]
- Elfving, D.C.; Lombardini, L.; McFerson, J.R.; Drake, S.R. Effects of directed applications of prohexadione-calcium to tops of mature pear trees on shoot growth, light penetration, pruning and fruit quality. J. Am. Pomol. Soc. 2003, 57, 45–57. [Google Scholar]
- Einhorn, T.C.; Turner, J.; Laraway, D. Effect of reflective fabric on yield of mature ‘d’Anjou’ pear trees. HortScience 2012, 47, 1580–1585. [Google Scholar] [CrossRef]
- Zhang, J.; Serra, S.; Leisso, R.S.; Musacchi, S. Effect of light microclimate on the quality of ‘d’Anjou’ pears in mature open-centre tree architecture. Biosyst. Eng. 2016, 141, 1–11. [Google Scholar] [CrossRef]
- Khemira, H.; Lombard, P.B.; Sugar, D.; Azarenko, A.N. Hedgerow Orientation Affects Canopy Exposure, Flowering, and Fruiting of Anjou’ Pear Trees. HortScience 1993, 28, 984–987. [Google Scholar] [CrossRef]
- Mika, A. Physiological responses of fruit trees to pruning. Hortic. Rev. 1986, 8, 337–378. [Google Scholar]
- Lakso, A.N.; Corelli-Grappadelli, L. Implications of pruning and training practices to carbon partitioning and fruit development in apple. Acta Hortic. 1991, 322, 231–240. [Google Scholar] [CrossRef]
- DeJong, T.M. Canopy and light management. In Pear Production and Handling Manual; UC ANR Publications: Davis, CA, USA, 2007; Volume 3483, pp. 59–62. [Google Scholar]
- Suni, M.; Nyman, M.; Eriksson, N.A.; Björk, L.; Björck, I. Carbohydrate composition and content of organic acids in fresh and stored apples. J. Sci. Food Agric. 2000, 80, 1538–1544. [Google Scholar] [CrossRef]
- Palmer, J.W.; Harker, F.R.; Tustin, D.S.; Johnston, J. Fruit dry matter concentration: A new quality metric for apples. J. Sci. Food Agric. 2010, 90, 2586–2594. [Google Scholar] [CrossRef]
- Serra, S.; Goke, A.; Diako, C.; Vixie, B.; Ross, C.; Musacchi, S. Consumer perception of d’Anjou pear classified by dry matter at harvest using near-infrared spectroscopy. Int. J. Food Sci. 2019, 54, 2256–2265. [Google Scholar] [CrossRef]
- Escribano, S.; Biasi, W.V.; Lerud, R.; Slaughter, D.C.; Mitcham, E.J. Non-destructive prediction of soluble solids and dry matter content using NIR spectroscopy and its relationship with sensory quality in sweet cherries. Postharvest Biol. Technol. 2017, 128, 112–120. [Google Scholar] [CrossRef]
- Burdon, J.; McLeod, D.; Lallu, N.; Gamble, J.; Petley, M.; Gunson, A. Consumer evaluation of “Hayward” kiwifruit of different at-harvest dry matter contents. Postharvest Biol. Technol. 2004, 34, 245–255. [Google Scholar] [CrossRef]
- Jaeger, S.R.; Harker, R.; Triggs, C.M.; Gunson, A.; Campbell, R.L.; Jackman, R.; Requejo-Jackman, C. Determining consumer purchase intentions: The importance of dry matter, size, and price of kiwifruit. J. Food Sci. 2011, 76, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Mooney, H.A. The carbon balance of plants. Annu. Rev. Ecol. Evol. Syst. 1972, 3, 315–346. [Google Scholar] [CrossRef]
- Cannell, M.G.R. Dry matter partitioning in tree crops. In Attributes of Trees as Crop Plants; Cannell, M.G.R., Jackson, J.E., Eds.; Institute of Terrestrial Ecology: Cambridgeshire, UK, 1985; pp. 160–193. [Google Scholar]
- DeJong, T.M. Developmental and environmental control of dry-matter partitioning in peach. HortScience 1999, 34, 1037–1040. [Google Scholar] [CrossRef] [Green Version]
- Génard, M.; Dauzat, J.; Franck, N.; Lescourret, F.; Moitrier, N.; Vaast, P.; Vercambre, G. Carbon allocation in fruit trees: From theory to modelling. Trees 2008, 22, 269–282. [Google Scholar] [CrossRef]
- Grappadelli, L.C.; Lakso, A.N.; Flore, J.A. Early season patterns of carbohydrate partitioning in exposed and shaded apple branches. J. Am. Soc. Hortic. Sci. 1994, 119, 596–603. [Google Scholar] [CrossRef] [Green Version]
- Mesa, K.; Serra, S.; Masia, A.; Gagliardi, F.; Bucci, D.; Musacchi, S. Preliminary Study on Effect of Early Defoliation on Dry Matter Accumulation and Storage of Reserves on ‘Abbé Fétel’ Pear Trees. HortScience 2019, 54, 2169–2177. [Google Scholar] [CrossRef] [Green Version]
- Palmer, J.; Diack, R.; Johnston, J.; Boldingh, H. Manipulation of fruit dry matter accumulation and fruit size in ‘Scifresh’ apple through alteration of the carbon supply, and its relationship with apoplastic sugar composition. J. Hortic. Sci. Biotechnol. 2013, 88, 483–489. [Google Scholar] [CrossRef]
- Link, H. Significance of flower and fruit thinning on fruit quality. Plant Growth Regul. 2000, 31, 17–26. [Google Scholar] [CrossRef]
- Anthony, B.; Serra, S.; Musacchi, S. Optimizing Crop Load for New Apple Cultivar: “WA38”. Agronomy 2019, 9, 107. [Google Scholar] [CrossRef] [Green Version]
- Kappel, F.; Neilsen, G.H. Relationship between light microclimate, fruit growth, fruit quality, specific leaf weight and N and P content of spur leaves of ‘Bartlett’ and ‘Anjou’ pear. Sci. Hortic. (Amst.) 1994, 59, 187–196. [Google Scholar] [CrossRef]
- Serra, S.; Sullivan, N.; Mattheis, J.P.; Musacchi, S.; Rudell, D.R. Canopy attachment position influences metabolism and peel constituency of European pear fruit. BMC. Plant Biol. 2018, 18, 364. [Google Scholar] [CrossRef] [PubMed]
- Nicolai, B.M.; Beullens, K.; Bobelyn, E.; Peirs, A.; Saeys, W.; Theron, K.I.; Lammertyn, J. Nondestructive measurement of fruit and vegetable quality by means of NIR spectroscopy: A review. Postharvest Biol. Technol. 2007, 46, 99–118. [Google Scholar] [CrossRef]
- Lin, H.; Ying, Y. Theory and application of near infrared spectroscopy in assessment of fruit quality: A review. Sens. Instrum. Food Qual. Saf. 2009, 3, 130–141. [Google Scholar] [CrossRef]
- Walsh, K.B.; Golic, M.; Greensill, C.V. Sorting of fruit using near infrared spectroscopy: Application to a range of fruit and vegetables for soluble solids and dry matter content. J. Near Infrared Spectrosc. 2004, 12, 141–148. [Google Scholar] [CrossRef]
- Walsh, K.B.; McGlone, V.A.; Han, D.H. The uses of near infra-red spectroscopy in postharvest decision support: A review. Postharvest Biol. Technol. 2020, 163, 111139. [Google Scholar] [CrossRef]
- Lakso, A.; Corelli-Grappadelli, L. Le techniche di potatura e allevamento influenzano la ripartizione dei carboidrati e lo sviluppo dei frutti di melo. Frutticoltura 1992, 54, 15–20. [Google Scholar]
- Forshey, C.G.; Elfving, D.C. The relationship between vegetative growth and fruiting in apple trees. Hortic. Rev. 1989, 11, 229–287. [Google Scholar]
- Ashraf, N.; Ashraf, M. Summer pruning in fruit trees. Afr. J. Agric. Res. 2014, 9, 206–210. [Google Scholar]
- Musacchi, S.; Beers, E.; Mattheis, D. Fall and Summer Pruning to Control Vigor and Psylla in d’Anjou Pear; PR14-104; Washington Tree Fruit Research Commission: Wenatchee, WA, USA, 2017; pp. 1–10. [Google Scholar]
- Cain, J.C. Effects of mechanical pruning of apple hedgerows with a slotting saw on light penetration and fruiting. J. Am. Soc. Hortic. Sci. 1971, 96, 664–667. [Google Scholar]
- McArtney, S.; Greene, D.; Schmidt, T.; Yuan, R. Naphthaleneacetic acid and ethephon are florigenic in the biennial apple cultivars Golden Delicious and York imperial. HortScience 2013, 48, 742–746. [Google Scholar] [CrossRef] [Green Version]
- Wilkie, J.D.; Sedgley, M.; Olesen, T. Regulation of floral initiation in horticultural trees. J. Exp. Bot. 2008, 59, 3215–3228. [Google Scholar] [CrossRef] [Green Version]
- Bartolini, S.; Viti, R.; Andreini, L. The effect of summer shading on flower bud morphogenesis in apricot (Prunus americana L.). Open Life Sci. 2013, 8, 54–63. [Google Scholar] [CrossRef]
- Elkins, R.; van den Ende, B.; Stebbins, R.; Micke, W. Training young trees. In Pear Production and Handling Manual; UC ANR Publications: Davis, CA, USA, 2007; Volume 3483, pp. 63–76. [Google Scholar]
- Sansavini, S.; Neri, D.; Ancarani, V. Innovazioni tecniche per migliorare l’efficienza produttiva e la qualità dei frutti nelle coltivazioni intensive. Riv. Frutticolt. Ortofloric. 2008, 70, 10–26. [Google Scholar]
- Lakso, A.N.; Robinson, T.L. Principles of orchard systems management optimizing supply, demand and partitioning in apple trees. Acta Hortic. 1997, 451, 405–416. [Google Scholar] [CrossRef]
- Beers, E.H.; Brunner, J.F.; Willett, M.J.; Warner, G.M. Orchard Pest Management: A Resource Book for the Pacific Northwest; Good Fruit Grower: Yakima, WA, USA, 1993; p. 276. [Google Scholar]
- Horton, D.R. Monitoring of pear psylla for pest management decisions and research. Integr. Pest Manag. Rev. 1999, 4, 1–20. [Google Scholar] [CrossRef]
- Lawson, D.S.; Reissig, W.H.; Agnello, A.M. Effects of summer pruning and hand fruit thinning on oblique-banded leafroller (Lepidoptera: Tortricidae) fruit damage in New York state apple orchards. J. Agric. Entomol. 1998, 15, 113–123. [Google Scholar]
- Simon, S.; Sauphanor, B.; Lauri, P.E. Control of fruit tree pests through manipulation of tree architecture. Pest. Technol. 2007, 1, 33–37. [Google Scholar]
- Chandler, W.H. Results of Some Experiments in Pruning Fruit Trees; Cornell University Agricultural Experiment Station: Ithaca, NY, USA, 1923; Volume 415. [Google Scholar]
- Ashraf, N.; Ashraf, M.; Bhat, M.Y.; Sharma, M.K. Paclobutrazol and summer pruning influences fruit quality of red delicious apple. Int. J. Agric. Environ. Biotechnol. 2017, 10, 349. [Google Scholar] [CrossRef]
- Demirtas, M.N.; Bolat, I.; Ercisli, S.; Ikinci, A.; Olmez, H.A.; Sahin, M.; Altindag, M.; Celik, B. The effects of different pruning treatments on the growth, fruit quality and yield of ‘Hacihaliloglu’ apricot. Acta Sci. Pol. Hortorum Cultus 2010, 9, 183–192. [Google Scholar]
- Pramanick, K.K.; Kashyap, P.; Kishore, D.K.; Shajtma, Y.P. Effect of summer pruning and CPPU on yield and quality of kiwi fruit (Actinidia deliciosa). J. Environ. Biol. 2015, 36, 351. [Google Scholar]
- Miller, S.S. Regrowth, flowering, and fruit quality of ’Delicious’ apple trees as influenced by summer pruning. J. Am. Soc. Hortic. Sci. 1982, 107, 975–978. [Google Scholar]
- Li, K.T.; Lakso, A.N.; Piccioni, R.; Robinson, T. Summer pruning effects on fruit size, fruit quality, return bloom and fine root survival in apple trees. J. Hortic. Sci. Biotechnol. 2003, 78, 755–761. [Google Scholar] [CrossRef]
- Morgan, D.C.; Stanley, C.J.; Volz, R.; Warrington, I.J. Summer pruning of Gala apple: The relationships between pruning time, radiation penetration, and fruit quality. J. Am. Soc. Hortic. Sci. 1984, 109, 637–642. [Google Scholar]
- Ystaas, J. Effects of summer pruning on yield, fruit size, and fruit quality of the apple cultivar ‘Summer red’. Acta Hortic. 1991, 322, 277–282. [Google Scholar]
- Ferree, D.C.; Myers, S.C.; Rom, C.R.; Taylor, B.H. Physiological aspects of summer pruning. Acta Hortic. 1984, 146, 243–252. [Google Scholar] [CrossRef]
- Saure, M.C. Summer pruning effects in apple—A review. Sci. Hortic. 1987, 30, 253–282. [Google Scholar] [CrossRef]
- DeLong, W.A. Variations in the chief ash constituents of apples affected with blotchy cork. Plant Physiol. 1936, 11, 453. [Google Scholar] [CrossRef]
- Bangerth, F. Calcium-related physiological disorders of plants. Annu. Rev. Phytopathol. 1979, 17, 97–122. [Google Scholar] [CrossRef]
- Autio, W.R.; Greene, D.W. Summer Pruning Affects Yield and Improves Fruit Quality of ‘McIntosh’ Apples. J. Am. Soc. Hortic. Sci. 1990, 115, 356–359. [Google Scholar] [CrossRef] [Green Version]
- Maas, F.M. Shoot growth, fruit production and return bloom in ‘Conference’ and ‘Doyenné du Comice’ treated with Regalis (prohexadione-calcium). Acta Hortic. 2004, 671, 517–524. [Google Scholar]
- Asin, L.; Alegre, S.; Montserrat, R. Effect of paclobutrazol, prohexadione-Ca, deficit irrigation, summer pruning and root pruning on shoot growth, yield, and return bloom, in a ‘Blanquilla’ pear orchard. Sci. Hortic. 2007, 113, 142–148. [Google Scholar] [CrossRef]
- Pasa, M.S.; Einhorn, T.C. Heading cuts and prohexadione-calcium affect the growth and development of ‘d’Anjou’ pear shoots in a high-density orchard. Sci. Hortic. 2014, 168, 267–271. [Google Scholar] [CrossRef]
- Sansavini, S.; Musacchi, S. Canopy architecture, training and pruning in the modern European pear orchards: An overview. Acta Hortic. 1993, 367, 152–172. [Google Scholar] [CrossRef]
- Sansavini, S. Pear fruiting-branch models related to yield control and pruning. Acta Hortic. 2000, 596, 627–633. [Google Scholar] [CrossRef]
- Mitcham, E.J.; Elkins, R.B. Pruning mature trees. In Pear Production and Handling Manual; UC ANR Publications: Davis, CA, USA, 2007; Volume 3483, pp. 77–84. [Google Scholar]
- Garriz, P.I.; Colavita, G.M.; Alvarez, H.L. Fruit and spur leaf growth and quality as influenced by low irradiance levels in pear. Sci. Hortic. 1998, 77, 195–205. [Google Scholar] [CrossRef]
- Goke, A.; Serra, S.; Musacchi, S. Postharvest Dry Matter and Soluble Solids Content Prediction in d’Anjou and Bartlett Pear Using Near-infrared Spectroscopy. HortScience 2018, 53, 669–680. [Google Scholar] [CrossRef]
- Chen, P.M.; Mellenthin, W.M. Maturity, chilling requirement, and dessert quality of ’d’Anjou’ and ’Bosc’ pears. Acta Hortic. 1981, 124, 203–210. [Google Scholar] [CrossRef]
- Argenta, L.C.; Fan, X.; Mattheis, J.P. Influence of 1-methylcyclopropene on ripening, storage life, and volatile production by d’Anjou cv. pear fruit. J. Agric. Food Chem. 2003, 51, 3858–3864. [Google Scholar] [CrossRef] [PubMed]
- Sugar, D.; Einhorn, T.C. Conditioning temperature and harvest maturity influence induction of ripening capacity in ‘d’Anjou’ pear fruit. Postharvest Biol. Technol. 2011, 60, 121–124. [Google Scholar] [CrossRef]
- Villalobos-Acuña, M.; Mitcham, E.J. Ripening of European pears: The chilling dilemma. Postharvest Biol. Technol. 2008, 49, 187–200. [Google Scholar] [CrossRef]
- Ziosi, V.; Noferini, M.; Fiori, G.; Tadiello, A.; Trainotti, L.; Casadoro, G.; Costa, G. A new index based on vis spectroscopy to characterize the progression of ripening in peach fruit. Postharvest Biol. Technol. 2008, 49, 319–329. [Google Scholar] [CrossRef]
- McGuire, R.G. Reporting of objective color measurements. HortScience 1992, 27, 1254–1255. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing, v. 3.6; R Foundation for statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Utermark, H. Summer pruning to control growth and maintain fruiting in mature apple trees. Compact Fruit Trees 1977, 10, 86–90. [Google Scholar]
- Maggs, D.H. Dormant and summer pruning compared by pruning young apple trees once on a succession of dates. J. Hortic. Sci. 1965, 40, 249–265. [Google Scholar] [CrossRef]
- Marini, R.P.; Barden, J.A. Effects of summer vs. dormant pruning and NAA treatment on growth of one-and two-year-old apple trees. J. Am. Soc. Hortic. Sci. 1982, 107, 604–607. [Google Scholar]
- Lakso, A.N.; Robinson, T.L.; Pool, R.M. Canopy microclimate effects on patterns of fruiting and fruit development in apples and grapes. In Manipulation of Fruiting; Wright, C.J., Ed.; Bunerworths: London, UK, 1989; pp. 263–274. [Google Scholar]
- Theron, K.I. Size matters: Factors influencing fruit size in pear. Acta Hortic. 2011, 909, 545–555. [Google Scholar] [CrossRef]
- Tufts, W.; Morrow, E. Fruit-bud differentiation in deciduous fruits. Hilgardia 1925, 1, 1–14. [Google Scholar] [CrossRef]
- Mpelasoka, B.; Behboudian, M.; Green, S. Water use, yield and fruit quality of lysimeter-grown apple trees: Responses to deficit irrigation and to crop load. Irrig. Sci. 2001, 20, 107–113. [Google Scholar]
- Washington State University. AgWeatherNet Growing Degree Days. Available online: https://weather.wsu.edu/?p=90850 (accessed on 23 May 2020).
- Raese, J.T. Performance of ‘Anjou’ pear Trees Sprayed with Calcium or Summer Pruned. Acta Hortic. 1992, 322, 315–324. [Google Scholar] [CrossRef]
- Al-Ani, A.M. Postharvest Physiology of ‘Anjou’ Pear Fruit: Relations between Mineral Nutrition and Cork Spot, Respiration, and Ethylene Evolution. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 1978. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 mo. Post-Harvest | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Year | Pruning | IAD | Overcolor (%) | Hue (°h) | Chroma (C*) | ||||
| 2016 | F | 1.81 | b | 9.2 | ns | 112.6 | ab | 43.9 | b |
| F+S | 1.92 | a | 12.4 | 113.0 | a | 43.5 | b | ||
| W | 1.86 | ab | 10.4 | 112.1 | b | 44.8 | a | ||
| W+S | 1.88 | a | 12.2 | 112.3 | b | 44.5 | a | ||
| 2017 | F | 1.76 | ns | 5.8 | ns | 111.6 | a | 44.4 | bc |
| F+S | 1.83 | 6.1 | 112.3 | a | 44.2 | c | |||
| W | 1.76 | 6.4 | 110.9 | b | 45.0 | a | |||
| W+S | 1.81 | 7.8 | 111.8 | a | 44.9 | ab | |||
| 5 Mos. Post-Harvest | |||||||||
| Year | Pruning | IAD | Overcolor (%) | Hue (°h) | Chroma (C*) | ||||
| 2016 | F | 0.84 | b | 9.2 | ns | 98.3 | b | 44.9 | b |
| F+S | 0.91 | b | 12.4 | 98.8 | b | 44.9 | b | ||
| W | 1.01 | a | 8.8 | 99.2 | b | 44.6 | b | ||
| W+S | 1.05 | a | 10.2 | 100.6 | a | 46.3 | a | ||
| 2017 | F | 1.17 | b | 9.0 | a | 102.7 | b | 45.0 | ab |
| F+S | 1.00 | c | 7.6 | ab | 102.7 | b | 45.3 | ab | |
| W | 1.37 | a | 3.3 | b | 104.3 | ab | 44.6 | b | |
| W+S | 1.26 | ab | 5.6 | ab | 104.9 | a | 45.7 | a | |
| 1 mo. Post-Harvest | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Year | Dry Matter Class | IAD | Overcolor (%) | Hue (°h) | Chroma (C*) | ||||
| 2016 | Low | 2.04 | a | 3.5 | c | 112.9 | a | 44.0 | ns |
| Moderate | 1.89 | b | 9.5 | b | 112.6 | a | 44.1 | ||
| High | 1.64 | c | 25.4 | a | 111.5 | b | 44.4 | ||
| 2017 | Low | 1.87 | a | 1.6 | b | 111.8 | 44.7 | ns | |
| Moderate | 1.73 | b | 10.1 | a | 111.6 | 44.6 | |||
| High | - | - | - | - | |||||
| 5 mos. Post-Harvest | |||||||||
| Year | Dry Matter Class | IAD | Overcolor (%) | Hue (°h) | Chroma (C*) | ||||
| 2016 | Low | 1.36 | a | 2.4 | c | 102.3 | a | 46.3 | a |
| Moderate | 0.91 | b | 9.9 | b | 98.9 | b | 45.0 | b | |
| High | 0.72 | c | 24.2 | a | 96.7 | c | 44.2 | b | |
| 2017 | Low | 1.46 | a | 2.6 | b | 105.7 | a | 45.3 | ns |
| Moderate | 1.04 | b | 8.8 | a | 102.4 | b | 45.0 | ||
| High | - | - | - | - | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goke, A.; Serra, S.; Musacchi, S. Manipulation of Fruit Dry Matter via Seasonal Pruning and Its Relationship to d’Anjou Pear Yield and Fruit Quality. Agronomy 2020, 10, 897. https://doi.org/10.3390/agronomy10060897
Goke A, Serra S, Musacchi S. Manipulation of Fruit Dry Matter via Seasonal Pruning and Its Relationship to d’Anjou Pear Yield and Fruit Quality. Agronomy. 2020; 10(6):897. https://doi.org/10.3390/agronomy10060897
Chicago/Turabian StyleGoke, Alex, Sara Serra, and Stefano Musacchi. 2020. "Manipulation of Fruit Dry Matter via Seasonal Pruning and Its Relationship to d’Anjou Pear Yield and Fruit Quality" Agronomy 10, no. 6: 897. https://doi.org/10.3390/agronomy10060897
APA StyleGoke, A., Serra, S., & Musacchi, S. (2020). Manipulation of Fruit Dry Matter via Seasonal Pruning and Its Relationship to d’Anjou Pear Yield and Fruit Quality. Agronomy, 10(6), 897. https://doi.org/10.3390/agronomy10060897