Abstract
Egyptian henbane (Hyoscyamus muticus L.) plants are rich sources of alkaloids used in pharmaceutical products. Recently, rising efforts have been devoted to reducing mineral fertilizer supply, production cost, and environmental pollution via decreasing the doses of nitrogenous fertilizers and adopting biofertilizer farming systems. Two field experiments were conducted to examine the potential role of N fixing bacteria Azotobacter spp. and Azospirillum spp. on the growth, mineral status, tropane alkaloids, leaf anatomy, and seed yield of Egyptian henbane grown with different levels of mineral nitrogen fertilizer, i.e., 25%, 50%, and 100% of the recommended dose, equal to 30, 60, and 120 kg N ha−1. N fertilizer improved growth, mineral elements, tropane alkaloids, seed yield, and yield components of Egyptian henbane, which showed a gradually rising trend as the rate of N fertilizer increased. High doses of N fertilizer presumably elicited favorable changes in the anatomical structure of Egyptian henbane leaves. The application of 50% N dose plus N fixing bacteria affected Egyptian henbane trials similarly to 100% of recommended N dose. In conclusion, the N fixing bacteria proved to be a sustainable tool for a two-fold reduction in the recommended dose of mineral N fertilizer and the sustainable management of Egyptian henbane nutrition.
1. Introduction
The family Solanaceae consists of about 85 genera and 2800 species of tropical and temperate distribution [1]. Tropane alkaloids are reported in 21 genera of the Solanaceae family, of which Hyoscyamus is one of the most important in this respect. The genus comprised of about 15 species distributed in North Africa and Western Europe to Central Asia [2]. Hyoscyamus niger L. (henbane, black henbane or European henbane) is cultivated in South-Eastern England, in Thuringia and Northern Bavaria in Germany, in Russia, and Hungary, while Hyoscyamus muticus L. (Egyptian henbane) is indigenous to desert regions in Egypt, Iran, Baluchistan, Sind, Western Punjab, and it has been introduced into Algeria and Southern California in the USA [3]. Both species are of high economic importance for their tropane alkaloids (hyoscyamine and hyoscine) which are widely used in medicine because of analgesic, anticholinergic, antispasmodic, mydriatic, and sedative activity [4]. Tropane alkaloids are industrially extracted from Hyoscyamus and the other Solanaceae belonging to Atropa, Duboisia, and Datura genera because its synthetic production is uneconomical due to the complex chemical structure and route of the biosynthesis [5].
Egyptian henbane is a wild herbaceous plant about 60 cm in height, richly branched, with dense flower-spikes at the end of the stems; the leaves are pubescent, pale green, rhomboidal or broadly elliptical, up to 15 cm long [6]. The plant is an annual herb when cultivated, reaching more than 80 cm in height. Egyptian henbane contains 0.7–1.5% of total alkaloids, most of which is hyoscyamine [7]. It is reported that total alkaloids in leaves of Egyptian henbane reached its maximum level in the 14th week of cultivation, being 45.6 mg g−1 dry weight (DW) and then showed a slight decline towards the end of the growing season, reaching 42.2 mg g−1 DW in 16 weeks, at full blooming stage [6].
Mineral fertilizers, particularly those containing nitrogen (N), phosphorus (P), and potassium (K), are important factors for vigorous plant growth and hence a higher yield of all crops [8,9,10,11]. Repeated application of mineral fertilizers, however, causes environmental risk, since N fertilizers are the main source of accumulating nitrates in soils and plants. Recently, great attention has been devoted to reduce the amounts of N fertilizers through adopting biofertilizers in the farming systems [12,13,14]. Nitrogen nutrition plays a crucial role in the biosynthesis of nitrogenous secondary metabolites, including the tropane alkaloids of Egyptian henbane, so the appropriate N management is critical for the pharmacological quality of this medicinal plant.
Biological fertilization of non-legume crops by N2-fixing bacteria has gained great importance in recent years because of its’ effectiveness in accelerating crop growth and yield. Nitrogen fixers such as Azotobacter spp. and Azospirillum spp. were found to have not only the ability to fix nitrogen but also to release certain phytohormones such as gibberellins and indolic substances, which could stimulate plant growth, nutrients availability and photosynthetic processes [15]. Plants can synthesize auxins, including the major one, indole-3-acetic-acid (IAA) themselves, however, the rhizospheric bacteria, especially of the genera Aeromonas, Azotobacter, Bacillus, Burkholderia, Enterobacter, Pseudomonas, and Rhizobium significantly contribute to the plant’s auxin pool [16]. The direct effect of bacterial auxins secretion is an accelerated length of root hairs, root growth, biomass, and surface, and modified architecture of the root system. Bacteria of Achromobacter, Azospirillum, Bacillus, Enterobacter, Pseudomonas, and Rhizobium genera possess 1-Aminocyclopropane-l-carboxylate (ACC)-deaminase activity. ACC-deaminase transforms ACC into alpha-keto-butyrate and ammonia thereby lower the level of ethylene in the plant and turn to stimulate plant growth [17,18]. Biological nitrogen fixation in no-leguminous crops and bacterial interactions has been widely studied for their agronomic importance during the past five decades. For example, in paddy rice (Oryza sativa L.) [19], sweet potato (Ipomoea batatas L.) [20], sugarcane (Saccharum officinarum L.) [21], and associative nitrogen fixation are agronomically important. In this regard, Boddey et al. [22] reported that some sugarcane varieties can acquire more than 60 percent of their nitrogen from the biological nitrogen fixation process associated with the plant. Moreover, a strong nitrogen balance has been measured, indicating a substantial atmospheric intake of nitrogen in paddy rice fields [19]. Generally, the application of nitrogen-fixing bacteria provides an integrated approach for growth promotion, disease management, secondary metabolites accumulation, and maintenance of the fertility of soil [23].
There are no data on the effect of nitrogen-fixing rhizobacteria on henbane, however considering henbane as the species demanding about N nutrition, the research in this field can bring promising results. The present study was designed to analyze the potential role of nitrogen-fixing bacteria Azotobacter spp. and Azospirillum spp. on tropane alkaloids, leaf anatomy, and seed yield of Egyptian henbane fertilized with different levels of mineral nitrogen. We hypothesized that there is a possibility to reduce mineral N fertilization without a decrease in raw herb quality. We also aimed to highlight the mechanisms of plant growth-promoting rhizobacteria action through the analysis of plant vegetative growth, morphology, leaf anatomy, and mineral status of Egyptian henbane.
2. Materials and Methods
2.1. Experimental Procedures
Two field experiments were carried out in 2017 and 2018 at the Agricultural Experiments and Research Station, Faculty of Agriculture, Cairo University, Giza, Egypt (30°01′38″ N, 31°11′35″ E).
Seeds of Egyptian henbane (Hyoscyamus muticus L.) were obtained from the Experimental Station of Medicinal Plants, Faculty of Pharmacy, Cairo University, Giza, Egypt. Seeds with uniform size and color were washed with distilled water, sterilized with 10% sodium hypochlorite solution for about 15 min, washed again with distilled water, and air-dried.
Rhizobacterein® biofertilizer was obtained from the Ministry of Agriculture and Land Reclamation, Egypt. Rhizobacterein® is the commercial name of nitrogen-fixing bacteria containing Azotobacter spp., Azospirillum spp., and Rhizobium spp., which can perform symbiotic and non-symbiotic N2-fixation in the most field, vegetable and fruit crops. Rhizobacterein was applied as a mixture of vermiculite:peat moss at a ratio of 9:1 (wt.:wt.), containing about 1 × 107 of these bacteria equally. For the inoculation of the seeds of Egyptian henbane with Rhizobacterein, a sticker was done by preparing a sugar solution of 1 cup sugar to 1 quart of warm water. The seeds were slightly moistened with a small amount of sticker solution and mixed with Rhizobacterein inoculant. Then, seeds and inoculants were poured between clean 5-gallon buckets. After mixing, all the seeds were evenly covered with specks of inoculant. Seeds were planted immediately in seedling trays, making sure the inoculated seeds were not exposed to sunlight. Rhizobacterein biofertilizer was used at a rate of 2.00 kg ha−1.
Seeds of Egyptian henbane either inoculated with Rhizobacterein biofertilizer or without inoculation were sown on 18 February 2017 in the first season and on 5 March 2018 in the second one in seedling trays in 40 × 60 cm. Uniform seedlings at 30 days after sowing were transplanted in the open field on the west side of 4 m long ridges, which were 60 cm apart, with a 30 cm spacing between hills with one seedling per hill. Each experimental unit consisted of 7 ridges assigned to a single treatment.
Physical and chemical analysis of the loamy-clay soil at the experimental site (Table 1) were done in each growing season before plowing for plantlet establishment [24].

Table 1.
Physical and chemical properties of the experimental soil in the two growing seasons.
The experiments were established in a split-plot design with three replicates. Each replicate contained three main plots, each assigned to one level of mineral nitrogen fertilizer. Each main plot was divided into two sub-plots, one of which was inoculated with Rhizobacterein biofertilizer and the other sub-plot served as an untreated control.
Nitrogen (N) fertilizers consisted of three levels, viz. 25%, 50%, and 100% of the recommended dose of 120 kg N ha−1 as ammonium sulfate (20.5% N), and considered as N30, N60, and N120, respectively. Biofertilizer included two treatments, viz., henbane plants inoculated with Rhizobacterein and without Rhizobacterein as control and considered as RB+ and RB−, respectively. Each replicate included six plots, viz., 3 N fertilizers (N30, N60, and N120) × 2 Rhizobacterein biofertilizer (BR+ and BR−). Therefore, each experiment consisted of six treatments as combinations of two factors (N fertilizers and Rhizobacterein biofertilizers) viz., T1, 30 kg N ha−1 without Rhizobacterein (30N × RB−); T2, 30 kg N ha−1 with Rhizobacterein (30N × RB+); T3, 60 kg N ha−1 without Rhizobacterein (60N × RB−); T4, 60 kg N ha−1 with Rhizobacterein (60N × RB+), T5, 120 kg N ha−1 without Rhizobacterein (120N × RB); and T6, 120 kg N ha−1 with Rhizobacterein (120N × RB+).
Phosphorus (P) fertilizer at the rate of 32 kg P ha−1 superphosphate (15.5% P2O5) was added as one dose before transplanting during the soil preparation for cultivation. Potassium (K) was added at the rate of 95 kg K ha−1 as potassium sulfate (48% K2O) in two equal doses, the first of which was added before transplanting and the second one a month later. All other cultural practices were carried out as recommended by the Ministry of Agriculture and Land Reclamation, Egypt. Figure 1 shows the general view for a side of the experiment at the full blooming stage carried out at the second growing season in 2018.

Figure 1.
General view from the experiment at Egyptian henbane full blooming stage carried out in the second season (2018).
2.2. Measurements
At 90 days after transplanting, equal to 120 days from sowing, ten plants from each experimental unit were sampled to determine morphological characters i.e., plant height in cm (PH), number of branches per plant (BNo), number of leaves per plant (LNo), and fresh weight of leaves per plant in g (LFW). The total leaf area per plant in dm2 (LA) was measured using a leaf area meter (LI-COR 3100; Licor, Lincoln, NB, USA).
For the anatomical study, at 60 days from transplanting (90 days from sowing), specimens of selected treatments were taken during the second season of 2018 from leaves developed in the middle part of the main stem. Specimens were fixed for at least 48 h in FAA (10 mL formalin, 5 mL glacial acetic acid, and 85 mL ethyl alcohol 70%). The selected materials were washed in 50% ethyl alcohol, dehydrated in normal butyl alcohol series, embedded in paraffin wax of 56 °C melting point, sectioned to a thickness of 20 micrometers (μm), double stained with the crystal violet erythrosine, cleared in xylene and mounted in Canada balsam [25]. Sections were examined to detect histological manifestations of the selected treatments and photomicrographed using an upright light microscope (AxioPlan, Zeiss, Jena, Germany). Measurements were done using a micrometer eyepiece and an average of five readings were calculated.
Macro- and micro-nutrients concentrations of N, P, K, calcium (Ca), sodium (Na) and iron (Fe), zinc (Zn), manganese (Mn), and copper (Cu) were determined in henbane plants represented all treatments grown in the second season of 2018 at 90 days after transplanting. Fresh samples of leaves were dried at 70 °C to constant moisture, and dried ground leaves (1 g) were digested in a mixture of boiling perchloric acid and hydrogen peroxide for 8 h. When the fumes were white and the solution was completely clear, it was cooled to room temperature and filled up to 10 mL with deionized water. Reagent blanks were prepared by carrying out the whole extraction procedure but in the absence of a sample. Total N was determined using the micro-Kjeldahl method. P was determined colorimetrically using stannous chloride-ammonium molybdate reagent [26], after its extraction by sodium bicarbonate [27]. K was determined using a flame photometer (ELE Flame Photometer, Leighton Buzzard, UK). Ca, Na, Fe, Zn, Mn, and Cu concentrations were determined by atomic absorption spectrophotometry [28]. N, P, K, Ca, Mg, Na, and Fe were expressed as mg g−1 DW, while Zn, Mn, and Cu were expressed as ppm.
Tropane alkaloids (TA) analysis by GC-MS and GC-FID were carried out from leaves represented all treatments and taken from plants grown in the second season of 2018 at full blooming at 90 days after transplanting equal to 120 days from sowing [29]. Samples of plant material (2.0 g) were homogenized in 5 mL EtOH and centrifuged at 4000 rpm for 10 min. This procedure was repeated three times, and the EtOH supernatants were combined and evaporated under vacuum. TA was purified from the EtOH extract by acid-base work up and analyzed by GC-MS [29]. For hyoscyamine and scopolamine quantification, GC-flame ionization detector (FID) analyses were carried out by using a Hewlett Packard 5890 GC with an FID coupled with a Hewlett Packard (HP 3396-A) electronic integrator, with SA-1 (100% polydimethylsiloxane) capillary column (30 m × 0.25 mm × 0.25 μm). The GC-FID conditions were as follows: injection temperature 250 °C; temperature program 150–300 °C, 4 °C, 1 min; split ratio 1:100; and carrier gas He 1 mL per min. An aliquot of 1 μL of the alkaloidal extracts was redissolved in 200 μL of a 2 mg mL−1 MeOH solution of stearic acid methyl ester was injected as an internal standard. Methyl stearate was used as the internal standard because of its retention time (RT), which does not overlap the RT of the TA, as a commercial alkaloid (the pyrrolizidine monocrotaline). Hyoscyamine (Hyoscy), and Scopolamine (Scopo) were measured as a component of tropane alkaloids (TA) and the sum of hyoscyamine plus scopolamine (Hyoscy+Scopo) was calculated as the total of TA and presented in mg g−1 dry weight (DW).
At 90 days after transplanting (120 days from sowing), ten plants from each treatment were assigned and labeled at the full blooming stage for recording yield characters, i.e., the number of capsules per plant (Cno), 1000-seeds weight in g (TSW), and seed yield per plant in g (SY).
2.3. Statistical Analysis
All data were subjected to an analysis of variance for a split-plot design [30], after testing for the homogeneity of error variances using the Levene test [31], and testing for normality distribution according to Shapiro and Wilk method [32]. Statistically significant differences between means were compared at p ≤ 0.05 using the least significant difference (LSD) test. The statistical analysis was carried out using GenStat 17th Edition (VSN International Ltd., Hemel Hempstead, UK). Correlation coefficient r was calculated to determine the relationship between seed yield and each of the physiological and chemical traits. Experimental data were also processed for a principal component analysis (PCA) using the Statistica 12.0 software package (StatSoft Inc., Tulsa, OK, USA), to evaluate the existing relationships with original variables.
3. Results
3.1. Vegetative Growth of Egyptian Henbane
The data reported in Table 2 reveal the effect of different levels of mineral nitrogen fertilizer alone or in combination with Rhizobacterein biofertilizer, the latter containing a mixture of nitrogen-fixing bacteria belonging to Azotbacter spp. and Azospirillium spp., on morphological and growth characteristics of Egyptian henbane plants. The increasing level of N fertilizer significantly and gradually enhanced all morphological characters in both research seasons, up to 100% of the N recommended dose. Increasing N fertilizer rate from 25% to 100% of recommended dose caused the increase of plant height by 22.4% and 31.3%, the number of branches per plant by 29.8% and 32.7%, leaf number per plant by 20.0% and 24.8%, total leaf area per plant by 20.5% and 22.4%, and leaf fresh weight per plant by 23.0% and 26.2%, in the 1st and 2nd season, respectively. The growth promotion caused by the rising level of mineral N fertilizer was similar to that induced by biofertilizer inoculation, which was generally effective in replacing half of the nitrogen fertilizer recommended dose. T4 (60N × RB+) significantly increased all growth characters with no significant difference between T4 and T5 or T6 when 120 kg N ha−1 were applied.
Table 2.
Effects of mineral nitrogen fertilizer (N) and Rrhizobacterein biofertilizer (RB) on morphological parameters of Egyptian henbane in 2017 and 2018.
3.2. Elemental Composition of Egyptian Henbane
Increasing the dose of N fertilizer significantly affected the concentrations of macronutrients (N, P, K, Ca, Mg) and micronutrients (Fe, Zn, Mn, Cu) in Egyptian henbane leaves (Table 3), except for Na which showed an opposite trend. In general, intensification of mineral salts accumulation caused by raising mineral N dose was not different from that induced by the biofertilizer. For most of the investigated elements, T4 treatment (60N × RB+) significantly increased all elements, and, in most cases, no significant differences between T4 and T5 or T6 were recorded when 120 kg N ha−1 were applied.
Table 3.
Effects of mineral nitrogen fertilizer (N) and Rhizobacterein biofertilizer (RB) on concentrations of macronutrients (N, P, K, Ca, Mg) and micronutrients (Na, Fe, Zn, Mn, Cu) in Egyptian henbane leaves.
3.3. Tropane Alkaloids in Egyptian Henbane
The increasing level of N fertilizer significantly risen hyoscyamine (Hyoscy), scopolamine (Scopo), out of tropane alkaloids (TA) in Egyptian henbane leaves at a gradually increasing rate up to 100% of the N fertilizer recommended dose (Table 4). Notably, raising the level of N fertilizer from 25 to 100% of the recommended dose induced a 50.4% increase of hyoscyamine concentration (the main tropane alkaloid), 59.1% in hyoscine concentration (the second major one), and 51.9% of total alkaloids. The increase in the concentration of tropane alkaloids due to biofertilizer treatment was 21.1% for hyoscyamine, 25.3% for hyoscine, and 21.9% for total alkaloids. Increasing N fertilizer dose combined with Rhizobacterein biofertilizer significantly induced favorable changes in Hyoscy, Scopo, and TA status in Egyptian henbane leaves, thus verifying that the increase induced by raising the level of mineral N fertilizer was similar to that induced by Rhizobacterein biofertilizer treatment. T4 (60N × RB+) significantly increased Hyoscy, Scopo, and TA concentrations with no significant difference between T4 and T5 or T6 when 120 kg N ha−1 were applied.
Table 4.
Effects of mineral nitrogen fertilizer (N) and Rhizobacterein biofertilizer (RB) on hyoscyamine, scopolamine, of total tropane alkaloids content in leaves of Egyptian henbane.
The maximum leaf alkaloids concentration in Egyptian henbane, as much as 37.57 mg g−1 DW for hyoscyamine, 8.94 mg g−1 DW for hyoscine, and 46.51 mg g−1 DW for total alkaloids was recorded at the 100% mineral N fertilizer in the combination with Rhizobacterein biofertilizer.
3.4. Leaf Anatomy of Egyptian Henbane
Microscopic measurements of some histological characters in transverse sections on the leaf blade developed in the middle part of the main stem of Egyptian henbane plant as affected by different levels of mineral N fertilizer with presence or absence of biofertilizer are reported in Table 5. Besides, microphotographs illustrating the effects of selected treatments are shown in Figure 2. Egyptian henbane plants receiving 50% of the recommended dose of mineral N fertilizer (60N) showed a prominent reduction in thickness of both midvein (−16.6%) and lamina (−12.1%) of the examined leaf, compared with the control plants which received 100% of the recommended N mineral dose (N120). The decrements referred to control were −3.4%, −40.5% and 40.1% for mesophyll thickness, midvein bundle length, and midvein bundle width, respectively. The results in Table 5 reveal that leaves of Egyptian henbane from bio-fertilized plants receiving half of the recommended dose of mineral N fertilizer (60N × BR+) showed positive changes in their structure, compared to the control plant leaves. Such a promotion effect reflected in increasing the thickness of both midvein and lamina by 9.3% concerning control. Likewise, compared to control, increases were recorded for mesophyll thickness (+8.2%) and midvein bundle length (+5.6%), an 8.8% decrease of midvein bundle width.
Table 5.
Measurements (μm) of histological characters in transverse sections of leaf blade taken from the middle part of Egyptian henbane main stem as affected by mineral nitrogen fertilizer (N) and Rhizobacterein biofertilizer (RB).
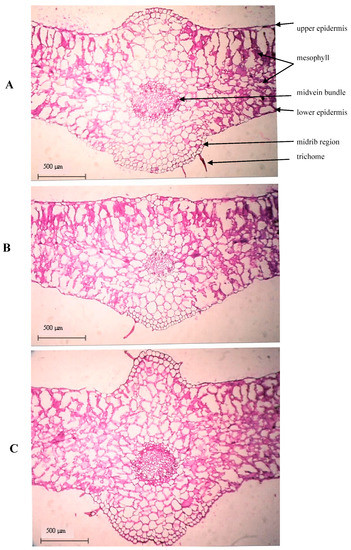
Figure 2.
Transverse sections of leaf blade taken from the middle part of Egyptian henbane main stem 90 days after sowing, as affected by N fertilizer and Rhizobacterein. (A) control plant received 120 kg N ha−1; (B) plant received 60 kg N ha−1; (C) plant received 60 kg N ha−1 plus Rhizobacterein®.
3.5. Seed Yield and Yield Components of Egyptian Henbane
Table 6 shows the effect of different levels of mineral N fertilizer alone or in combination with a Rhizobacterein on seed yield and yield components of Egyptian henbane. N fertilizer significantly increased the number of capsules per plant (CNo) and seed yield per plant (SY), with a nonsignificant increase of 1000-seeds weight (TSW) of Egyptian henbane, at a gradually rising rate up 100% of the recommended dose in both growing seasons (Table 6). Interestingly, raising the level of mineral N fertilizer from 25% to 100% of the recommended dose resulted in a significant increase of capsules number per plant by 30.2% and 31.2% and of seed yield per plant by 32.6% and 36.1%, in the first and second season respectively.
Table 6.
Effects of mineral nitrogen fertilizer (N) and Rhizobacterein biofertilizer (RB) on Egyptian henbane seed yield parameters in the growing seasons 2017 and 2018.
Rhizobacterein treatment of Egyptian henbane significantly increased capsules number per plant and seed yield per plant in both research seasons. However, bio fertilization did not significantly affect 1000-seeds weight (Table 6). The increments in seed yield characteristics due to biofertilizer inoculation were 11.5% and 13.9% for capsules number per plant and 15.3% and 15.0% for seed yield per plant in the first and second season, respectively. The promotion induced by raising the level of mineral N fertilizer was similar to that induced by biofertilizer. T4 treatment (60N × RB+) significantly increased the number of capsules per plant and seed yield per plant, with no significant differences between T4 and T5 or T6 when 120 kg N ha−1 were applied in both growing seasons.
3.6. Correlation Matrix
Pearson’s correlation coefficients (below diagonal) among all studied attributes of Egyptian henbane plants fertilized with three mineral N levels and Rhizobacterein inoculation are shown in Table 7. There is a high significance between seed yield and all of the studied traits, i.e., plant height, number of branches per plant, total leaf area per plant, fresh weight of leaves per plant, N, P, K, Ca, Fe, Zn, Mn, Cu, hyoscyamine, scopolamine, capsules number per plant, and 1000-seed weight. However, seed yield and leaf Na concentration showed a highly significant negative correlation.
Table 7.
Pearson’s correlation coefficients (below diagonal) among all studied attributes of Egyptian henbane grown with three mineral nitrogen fertilizer doses and Rhizobacterein inoculation.
3.7. Biplot Graph
Investigated treatments (green color) and measured traits (blue color) are shown in Figure 3, which illustrates a polygon view of the ordination of treatment from main component analysis outputs by trait biplots. The biplot explained 99.02% of the total variation of the standardized data. The first and second principal components (PC1 and PC2) explained 97.11% and 1.91%, respectively. This comparatively high percentage reveals the complication of the relations among the treatments and the evaluated variables. T4, including 60 kg N ha−1 with Rhizobacterein inoculation (60N × RB+), was the most efficient in most studied traits with no significant difference with either T5 or T6 when 120 kg N ha−1 was applied, either without or with biofertilizer inoculation, respectively. T1 and T2 scored the least in most of the studied variables under 30 kg N ha−1 application.
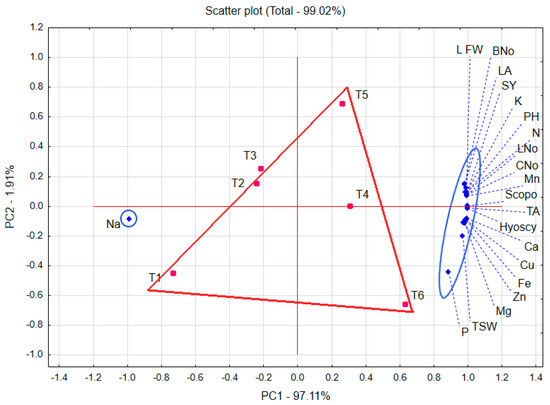
Figure 3.
Treatment ordination by trait biplots of principal component analysis outputs of Egyptian henbane research trials. T1, 30 kg N ha−1 without Rhizobacterein (30N × RB−); T2, 30 kg N ha−1 with Rhizobacterein (30N × RB+); T3, 60 kg N ha−1 without Rhizobacterein (60N × RB−); T4, 60 kg N ha−1 with Rhizobacterein (60N × RB+); T5, 120 kg N ha−1 without Rhizobacterein (120N × RB−); T6, 120 kg N ha−1 with Rhizobacterein (120N × RB+). PH, plant height; BNo, number of branches per plant; LA, total leaf area per plant; LFW, fresh weight of leaves per plant; N, nitrogen; P, phosphorus; K, potassium; Ca, calcium; Na, sodium; Fe, iron; Zn, zinc; Mn, manganese; Cu, copper; Hyoscy, hyoscyamine; Scopo, scopolamine; CNo, capsule number per plant; TSW, 1000-seed weight; SY, seed yield per plant.
4. Discussion
Nitrogen is an essential macro element for higher plants, playing a major function in their growth and metabolism, which directly affects plant productivity [11,33,34,35]. Thus, the positive effect of N on the vegetative growth characters of Egyptian henbane was due to its role as a component of amino acids, nucleic acids, many enzymes, energy-transfer compounds, and secondary metabolites, including the tropane alkaloids [36]. Similarly, in Hyoscyamus areticulatus and Hyoscyamus niger, morphological parameters were found to increase linearly with increasing mineral N doses, from 0 to 200 kg ha−1 [37]. Plant growth-promoting rhizobacteria inoculation increased root and shoot growth indexes in Hyoscyamus niger such as leaf number, area, and greenness [38]. In the present research, the increase of all vegetative growth characteristics induced by raising the level of mineral N fertilizer was similar to that induced by Rhizobacterein, which in general, substituted half of the mineral N recommended dose. Biofertilizer increased significantly all investigated morphological characteristics of Egyptian henbane in both studied seasons. The beneficial effect of inoculation with nitrogen fixers Azotobacter spp. and Azospirillum spp. was related to improving the fixation of atmospheric N and the wide spectrum of growth-promoting substances, including indole-3-acetic acid, gibberellic acid, and cytokinins, produced by applied rhizobacteria [39,40,41]. The activation of cell division, cell enlargement, and finally increased root and shoot growth potential was the result of exposure henbane root system too mentioned rhizobacteria exudates, an action described also for some other crops [15,18,42,43].
Great demand for N in different cropping systems is met through the use of N fertilizers, but production costs are increasing, and environmental concerns are also raised. This has led to increased interest in exploring other N sources such as biological nitrogen fixation. Non-symbiotic N2 fixation in soils or associated with the rhizosphere by free-living bacteria has the potential to meet some of this need particularly in the worldwide lower input cropping systems [44]. The potential N inputs from non-symbiotic N2 fixation through improved management practices, identifying better performing microbial strains and their successful field inoculation, and plant-based solutions were reviewed [44]. Plant growth-promoting rhizobacteria facilitate the plants in the uptake of essential nutrients from the soil [45] and this phenomenon was reflected in an enhanced mineral composition of bio fertilized henbane plants. In the present research, Rhizobacterein increased significantly all determined macroelements and microelements in leaves of Egyptian henbane except for sodium which showed a significant decrease in this respect. Plants inoculated with growth-promoting rhizobacteria have a well-developed root system, more branched and spread, with a high number of long hair roots increasing the surface area and minerals absorption. Changes in the root system morphology and functionality are followed by a systemic response of all plant organisms, which gains increased growth potential through better mineral nutrition [45,46]. Moreover, N-fixing bacteria can modify soil chemistry, for example, Azospirillum brasilense altered pH of the rhizosphere, increasing bioavailability of mineral elements [47]. One of the most noteworthy results of the present research is Na antagonism with the other elements, compounds, and morpho-anatomical characteristics of Egyptian henbane. Plant species are highly differentiated in their ability to accumulate or exclude Na from their tissues [48]. Na antagonistically interferes in the nitrate and ammonium uptake from the soil, thereby impairing nitrogen metabolism, for example, N mineral fertilization reduced Na toxicity in tomato [49], and this phenomenon was confirmed by the presented results.
Hyoscyamus muticus is a very important source of tropane alkaloids, which consist of up to 6% of the dry weight in the leaves of mature plants. In the present research, the mineral N fertilizer induced a significant increase in the concentration of tropane alkaloids in leaves of Egyptian henbane and the rate of elevation was proportional to the dose of mineral N fertilizer. The synthesis of tropane alkaloids, as nitrogenous compounds, depends on N availability, and this relation was confirmed for some medicinal plants, including deadly nightshade (Atropa belladonna) [50], Common periwinkle (Littorina littorea) [51], and black henbane (Hyoscyamus niger) [52]. The total alkaloid content in Hyoscyamus reticulatus ranged from 0.148% to 0.191%, and for H. niger varied from 0.14% to 0.24%, and increased proportionally to N mineral dose in research [37]. Pseudomonas putida and Pseudomonas fluorescens were proven to enhance tropane alkaloids production in Hyoscyamus niger under water deficit [38]. The mechanism of this phenomenon is rhizobacteria’s ability to produce growth regulators such as IAA, which act as elicitors on tropane alkaloids biosynthesis.
The thinner leaves induced by the median level of the mineral N used could be attributed to the decrease in thickness of mesophyll as well as in dimensions of the midvein bundle. Egyptian henbane leaves from treatments with the use of biofertilizer and half of the recommended dose of mineral N showed beneficial changes in anatomical structure pointing the higher effectiveness of mineral salts transport from roots to aboveground organs. Increases the rate of xylem was reported as the effect of IAA increased level caused by inoculation with plant growth-promoting rhizobacteria [23]. Additionally, Solanaceus plants synthesize tropane alkaloids in roots, from which most of them are transported to shoots and accumulated in the vacuoles of various tissues, acting as plant protection agents. The root secondary growth as well as the xylem transport effectiveness are significant factors determining tropane alkaloid composition of a herb [53]. In the present study, well developed vascular bundles in plants treated with biofertilizers together with the highest concentration of tropane alkaloids in herb show the possible mechanism of growth-promoting bacteria action, who induce not only the synthesis of secondary metabolites in roots, but enhance the root–shoot transport and shoot accumulation. As far as the authors are aware, previous information about the effect of mineral and bio-fertilizers on leaf anatomy of henbane or other related species are not available.
Rhizobacteria in biofertilizer increased significantly the number of capsules per plant and seed yield per plant but did not significantly affect 1000-seeds weight of Egyptian henbane seeds in both studied seasons. The beneficial effect of biofertilizer on seed yield might be attributed to the vigorous growth of biofertilized plants and to the number of metabolites synthesized by these plants, which reflected in an increasing number of harvested capsules and consequently an increase in seed yield per plant. Nodule and seed parameters of lentil, namely nodule number, nodule dry weight, fresh biomass, grain yield, straw yield, and nitrogen content in grains increased as a result of lowering of the ethylene production via inoculation with plant growth-promoting rhizobacteria [54]. For henbane, seed yield per plant is closely associated with the number of capsules per plant and number of seeds per capsule [37]. Seed and capsules morphological parameters, and the seed yield of Hyoscyamus reticulatus and H. niger, were positively affected by increasing mineral N doses [37].
Unlike chemical fertilizers, the adoption of biofertilizer techniques has been promoted as the most feasible and sustainable approach to soil fertility re-establishment [55]. Moreover, it was suggested that nitrogen-fixing bacteria associated with the sugarcane might be the source of the agronomically significant nitrogen inputs to this crop, due to long-term growing sugarcane in Brazil uses low N fertilizer inputs without seeming exhaustion of soil N response [22]. Increasing water and mineral uptake from the soil is attributed to plant anatomical characteristics modified by the mentioned growth regulators released by bacteria in the rhizosphere, including increased root surface area, root hairs, and root elongation [46]. Finally, the ability to convert N2 to NH4 makes N available for plants, enhancing the production of biologically active fungistatic substances, and maintains the balance between harmful and beneficial microorganisms in the rhizosphere [56].
5. Conclusions
Egyptian henbane plants are sources of tropane alkaloids for the pharmaceutical industry. However, the alkaloid yield is highly N-dose dependent. With the increasing problems associated with the use of synthetic N fertilizers in agriculture, there has been an ever-increasing interest in bio fertilization within sustainable crop systems. The effectiveness of beneficial microorganisms in N-fixation and improving plant productivity through multifactorial actions was shown in the present study. Biofertilizer application enabled 50% reduction of synthetic N dose without any adverse effect on Egyptian henbane yield and quality, evaluated through an analysis of plant growth characteristics, mineral composition, tropane alkaloids content, and seed yield and yield components. Moreover, biofertilizer application modified the anatomical structure of Egyptian henbane leaves towards a more effective vascular tissue transport. The use of N-fixing bacteria has proved to be a potential tool for sustainable Egyptian henbane production.
Author Contributions
This work is a combined effort of all the authors; Conceptualization and designing the field experiment, R.M.A.N., E.A.S., and M.T.A.; Performing the field experiment, collecting the samples for analysis, and collecting yield data, R.M.A.N., E.A.S., and M.T.A.; Performing the chemical analyses and anatomical studies, R.M.A.N. and E.A.S.; Statistical analysis of data, and producing the presentation tables and figures, A.S., G.C., and M.T.A.; Writing the original draft with contributions from all of the authors, R.M.A.N., E.A.S., G.C., A.S., and M.T.A.; Funding acquisition, reviewing and editing the whole manuscript, G.C., A.S., and M.T.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Faculty of Agriculture, Cairo University, Giza, Egypt, The National Research Centre, Cairo, Egypt, University of Naples Federico II, Italy, and the University of Agriculture in Krakow, Poland.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Simpson, M.G. Plant Systematics; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Boulos, L. Flora of Egypt; Al Hadara Publishing Cairo: Cairo, Egypt, 2005; Volume 4. [Google Scholar]
- Evans, W.C. Trease and Evans Pharmacognosy, International Edition E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Al-Snafi, A.E. Therapeutic importance of Hyoscyamus species grown in Iraq (Hyoscyamus albus, Hyoscyamus niger and Hyoscyamus reticulates)-A review. IOSR J. Pharm. 2018, 8, 18–32. [Google Scholar]
- Dehghan, E.; Ahmadi, F.S.; Ravandi, E.G.; Reed, D.W.; Covello, P.S.; Bahrami, A.R. An atypical pattern of accumulation of scopolamine and other tropane alkaloids and expression of alkaloid pathway genes in Hyoscyamus senecionis. Plant Physiol. Biochem. 2013, 70, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Nassar, R.M.; Azoz, S.N.; Salama, A.M. Salama Botanical Studies on Egyptian Henbane (Hyoscymus muticus L.). I- Morphology of Vegetative and Reproductive Growth and Alkaloidal Content. Curr. Sci. Int. 2016, 5, 8–25. [Google Scholar]
- Abd Elmaksood, W.; Ebad, F.; Hussein, B. In vitro Propagation of the Endangered Medicinal Plant Hyoscyamusmuticus L.(Egyptian Henbane). J. Appl. Environ. Biol. Sci. 2016, 6, 25–34. [Google Scholar]
- Attar, H.; Blavet, D.; Selim, E.; Abdelhamid, M.; Drevon, J.-J. Relationship between phosphorus status and nitrogen fixation by common beans (Phaseolus vulgaris L.) under drip irrigation. Int. J. Environ. Sci. Technol. 2012, 9, 1–13. [Google Scholar] [CrossRef]
- Dawood, M.G.; Abdelhamid, M.T.; Schmidhalter, U. Potassium fertiliser enhances the salt-tolerance of common bean (Phaseolus vulgaris L.). J. Hortic. Sci. Biotechnol. 2014, 89, 185–192. [Google Scholar] [CrossRef]
- El-Lethy, S.R.; Abdelhamid, M.T.; Reda, F. Effect of potassium application on wheat (Triticum aestivum L.) cultivars grown under salinity stress. World Appl. Sci. J. 2013, 26, 840–850. [Google Scholar]
- Rady, M.M.; Semida, W.M.; Hemida, K.A.; Abdelhamid, M.T. The effect of compost on growth and yield of Phaseolus vulgaris plants grown under saline soil. Int. J. Recycl. Org. Waste Agric. 2016, 5, 311–321. [Google Scholar] [CrossRef]
- Abdelhamid, M.T.; Selim, E.; El-Ghamry, A. Integrated effects of bio and mineral fertilizers and humic substances on growth, yield and nutrient contents of fertigated cowpea (Vigna unguiculata L.) grown on sandy soils. J. Agron. 2011, 10, 34–39. [Google Scholar] [CrossRef]
- Awad, N.; Turky, A.; Abdelhamid, M.; Attia, M. Ameliorate of environmental salt stress on the growth of Zea mays L. plants by exopolysaccharides producing bacteria. J. Appl. Sci. Res. 2012, 8, 2033–2044. [Google Scholar]
- Rady, M.M.; Mounzer, O.; Alarcón, J.; Abdelhamid, M.; Howladar, S. Growth, heavy metal status and yield of salt-stressed wheat (Triticum aestivum L.) plants as affected by the integrated application of bio-, organic and inorganic nitrogen-fertilizers. J. Appl. Bot. Food Qual. 2016, 89. [Google Scholar] [CrossRef]
- Bashan, Y.; De-Bashan, L.E. How the plant growth-promoting bacterium Azospirillum promotes plant growth—A critical assessment. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2010; Volume 108, pp. 77–136. [Google Scholar]
- Ahmad, F.; Ahmad, I.; Khan, M. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res. 2008, 163, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Müller, K.M.; Charles, T.C.; Vesely, S.; Glick, B.R. 1-aminocyclopropane-1-carboxylate (ACC) deaminase genes in rhizobia from southern Saskatchewan. Microb. Ecol. 2009, 57, 423–436. [Google Scholar] [CrossRef]
- Govindasamy, V.; Senthilkumar, M.; Gaikwad, K.; Annapurna, K. Isolation and characterization of ACC deaminase gene from two plant growth-promoting rhizobacteria. Curr. Microbiol. 2008, 57, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Ladha, J.; Tirol-Padre, A.; Reddy, C.; Cassman, K.; Verma, S.; Powlson, D.; Van Kessel, C.; Richter, D.d.B.; Chakraborty, D.; Pathak, H. Global nitrogen budgets in cereals: A 50-year assessment for maize, rice and wheat production systems. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Hill, W.A.; Bacon-Hill, P.; Crossman, S.M.; Stevens, C. Characterization of N2-fixing bacteria associated with sweet potato roots. Can. J. Microbiol. 1983, 29, 860–862. [Google Scholar] [CrossRef]
- Dobereiner, J. Nitrogen-fixing bacteria of the genusBeijerinckia Derx in the rhizosphere of sugar cane. Plant Soil 1961, 15, 211–216. [Google Scholar] [CrossRef]
- Boddey, R.M.; Urquiaga, S.; Alves, B.J.; Reis, V. Endophytic nitrogen fixation in sugarcane: Present knowledge and future applications. Plant Soil 2003, 252, 139–149. [Google Scholar] [CrossRef]
- Gupta, G.; Parihar, S.S.; Ahirwar, N.K.; Snehi, S.K.; Singh, V. Plant growth promoting rhizobacteria (PGPR): Current and future prospects for development of sustainable agriculture. J. Microb. Biochem. Technol. 2015, 7, 96–102. [Google Scholar]
- Jakson, M. Soil Chemical Analysis; Prentice Hall of India Private Limited: New Delhi, India, 1967. [Google Scholar]
- Nassar, M.; El-Sahhar, K. Botanical Preparations and Microscopy (Microtechnique); Academic Bookshop Giza: Dokki, Egypt, 1998; p. 219. [Google Scholar]
- King, E.J. Micro-Analysis in Medical Biohemistry; J. & A. Churchill: New Delhi, India, 1951. [Google Scholar]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; US Department of Agriculture: Washington, DC, USA, 1954.
- Chapman, H. Cation-exchange capacity. In Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties; John Wiley & Sons: Hoboken, NJ, USA, 1965; Volume 9, pp. 891–901. [Google Scholar]
- Freitas, A.V.; Trigo, J.R.; Brown, K.S.; Witte, L.; Hartmann, T.; Barata, L.E. Tropane and pyrrolizidine alkaloids in the ithomiinesPlacidula euryanassa andMiraleria cymothoe (Lepidoptera: Nymphalidae). Chemoecology 1996, 7, 61–67. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley & Sons: Hoboken, NJ, USA, 1984. [Google Scholar]
- Levene, H. Robust tests for equality of variances. In Contributions to Probability and Statistics. Essays in Honor of Harold Hotelling; Stanford University Press: Redwood City, CA, USA, 1961; pp. 279–292. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Abdelhamid, M.; Horiuchi, T.; Oba, S. Evaluation of the SPAD Value in Faba Bean (Vicia faba L.) Leaves in Relation to Different Fertilizer Applications. Plant Prod. Sci. 2003, 6, 185–189. [Google Scholar] [CrossRef]
- Gad, N.; Sekara, A.; Abdelhamid, M.T. The Potential Role of Cobalt and/or Organic Fertilizers in Improving the Growth, Yield, and Nutritional Composition of Moringa oleifera. Agronomy 2019, 9, 862. [Google Scholar] [CrossRef]
- Saleh, S.; Liu, G.; Liu, M.; Liu, W.; He, H.; Abdelhamid, M.T. Do NH4: NO3 ratio and harvest time affect celery (Apium graveolens) productivity and product quality? Folia Hortic. 2019, 31, 343–353. [Google Scholar] [CrossRef]
- Gregory, P.J.; Nortcliff, S. Soil Conditions and Plant Growt; Wiley-Blackwell: Chichester, West Sussex, UK, 2013. [Google Scholar]
- Kizil, S.; Tonҫer, Ö.; Sogut, T. Effect of Different Nitrogen Doses on Some Agricultural Characteristics and Alkaloid Content of Hyoscyamus reticulatus L. and Hyoscyamus niger L. Cercet. Agron. Mold. 2017, 50, 83–90. [Google Scholar] [CrossRef]
- Ghorbanpour, M.; Hatami, M.; Khavazi, K. Role of plant growth promoting rhizobacteria on antioxidant enzyme activities and tropane alkaloid production of Hyoscyamus niger under water deficit stress. Turk. J. Biol. 2013, 37, 350–360. [Google Scholar] [CrossRef]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Gray, E.; Smith, D. Intracellular and extracellular PGPR: Commonalities and distinctions in the plant–bacterium signaling processes. Soil Biol. Biochem. 2005, 37, 395–412. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Sankar, B.; Kishorekumar, A.; Gopi, R.; Somasundaram, R.; Panneerselvam, R. Pseudomonas fluorescens enhances biomass yield and ajmalicine production in Catharanthus roseus under water deficit stress. Colloids Surf. B Biointerfaces 2007, 60, 7–11. [Google Scholar] [CrossRef]
- Arkhipova, T.; Veselov, S.; Melentiev, A.; Martynenko, E.; Kudoyarova, G. Ability of bacterium Bacillus subtilis to produce cytokinins and to influence the growth and endogenous hormone content of lettuce plants. Plant Soil 2005, 272, 201–209. [Google Scholar] [CrossRef]
- Oberson, A.; Frossard, E.; Buehlmann, C.; Mayer, J.; Maeder, P.; Luescher, A. Nitrogen fixation and transfer in grass-clover leys under organic and conventional cropping systems. Plant Soil 2013, 371, 237–255. [Google Scholar] [CrossRef]
- Roper, M.; Gupta, V. Enhancing non-symbiotic N2 fixation in agriculture. Open Agric. J. 2016, 10, 7–27. [Google Scholar] [CrossRef]
- Rana, A.; Saharan, B.; Nain, L.; Prasanna, R.; Shivay, Y.S. Enhancing micronutrient uptake and yield of wheat through bacterial PGPR consortia. Soil Sci. Plant Nutr. 2012, 58, 573–582. [Google Scholar] [CrossRef]
- Potters, G.; Pasternak, T.P.; Guisez, Y.; Palme, K.J.; Jansen, M.A. Stress-induced morphogenic responses: Growing out of trouble? Trends Plant Sci. 2007, 12, 98–105. [Google Scholar] [CrossRef]
- Carrillo, A.E.; Li, C.Y.; Bashan, Y. Increased acidification in the rhizosphere of cactus seedlings induced by Azospirillum brasilense. Naturwissenschaften 2002, 89, 428–432. [Google Scholar] [CrossRef]
- Subbarao, G.; Ito, O.; Berry, W.; Wheeler, R. Sodium—A functional plant nutrient. Crit. Rev. Plant Sci. 2003, 22, 391–416. [Google Scholar]
- Zahedifar, M.; Ronaghi, A.; Moosavi, A.; Safarzadeh Shirazi, S. Influence of nitrogen and salinity levels on the fruit yield and chemical composition of tomato in a hydroponic culture. J. Plant Nutr. 2012, 35, 2211–2221. [Google Scholar] [CrossRef]
- Bensaddek, L.; Gillet, F.; Saucedo, J.E.N.; Fliniaux, M.-A. The effect of nitrate and ammonium concentrations on growth and alkaloid accumulation of Atropa belladonna hairy roots. J. Biotechnol. 2001, 85, 35–40. [Google Scholar] [CrossRef]
- Sreevalli, Y.; Kulkarni, R.; Baskaran, K.; Chandrashekara, R. Increasing the content of leaf and root alkaloids of high alkaloid content mutants of periwinkle through nitrogen fertilization. Ind. Crops Prod. 2004, 19, 191–195. [Google Scholar] [CrossRef]
- Banani, N.; Mehrafarin, A.; Hosseini Mazinani, S. Changes of Tropane Alkaloids in Black Henbane (Hyoscyamus niger L.) in Response to Different Types of Nitrogenous Fertilizers. J. Med. Plants 2017, 1, 56–67. [Google Scholar]
- Nakanishi, F.; Sasaki, K.; Shimomura, K. Kinetics of littorine content in various developing stages of regenerates of Atropa belladonna. Plant Cell Rep. 2000, 19, 1021–1026. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.A.; Khalid, M.; Shahzad, S.M.; Ahmad, M.; Soleman, N.; Akhtar, N. Integrated use of Rhizobium leguminosarum, plant growth promoting rhizobacteria and enriched compost for improving growth, nodulation and yield of lentil (Lens culinaris Medik.). Chil. J. Agric. Res. 2012, 72, 104. [Google Scholar] [CrossRef]
- Raimi, A.; Adeleke, R.; Roopnarain, A. Soil fertility challenges and Biofertiliser as a viable alternative for increasing smallholder farmer crop productivity in sub-Saharan Africa. Cogent Food Agric. 2017, 3, 1400933. [Google Scholar] [CrossRef]
- Apte, R.; Shende, S. Studies on Azotobacter chroococcum: IV. Seed Bacterization with Strains of Azotobacter chroococcum and Their Effect on Crop Yield. Zentralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene. Zweite Naturwissenschaftliche Abteilung: Mikrobiologie der Landwirtschaft, der Technologie und des Umweltschutzes 1981, 136, 637–640. [Google Scholar]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).