Potential Fertilization Capacity of Two Grapevine Varieties: Effects on Agricultural Production in Designation of Origin Areas in the Northwestern Iberian Peninsula




Abstract
1. Introduction
2. Materials and Methods
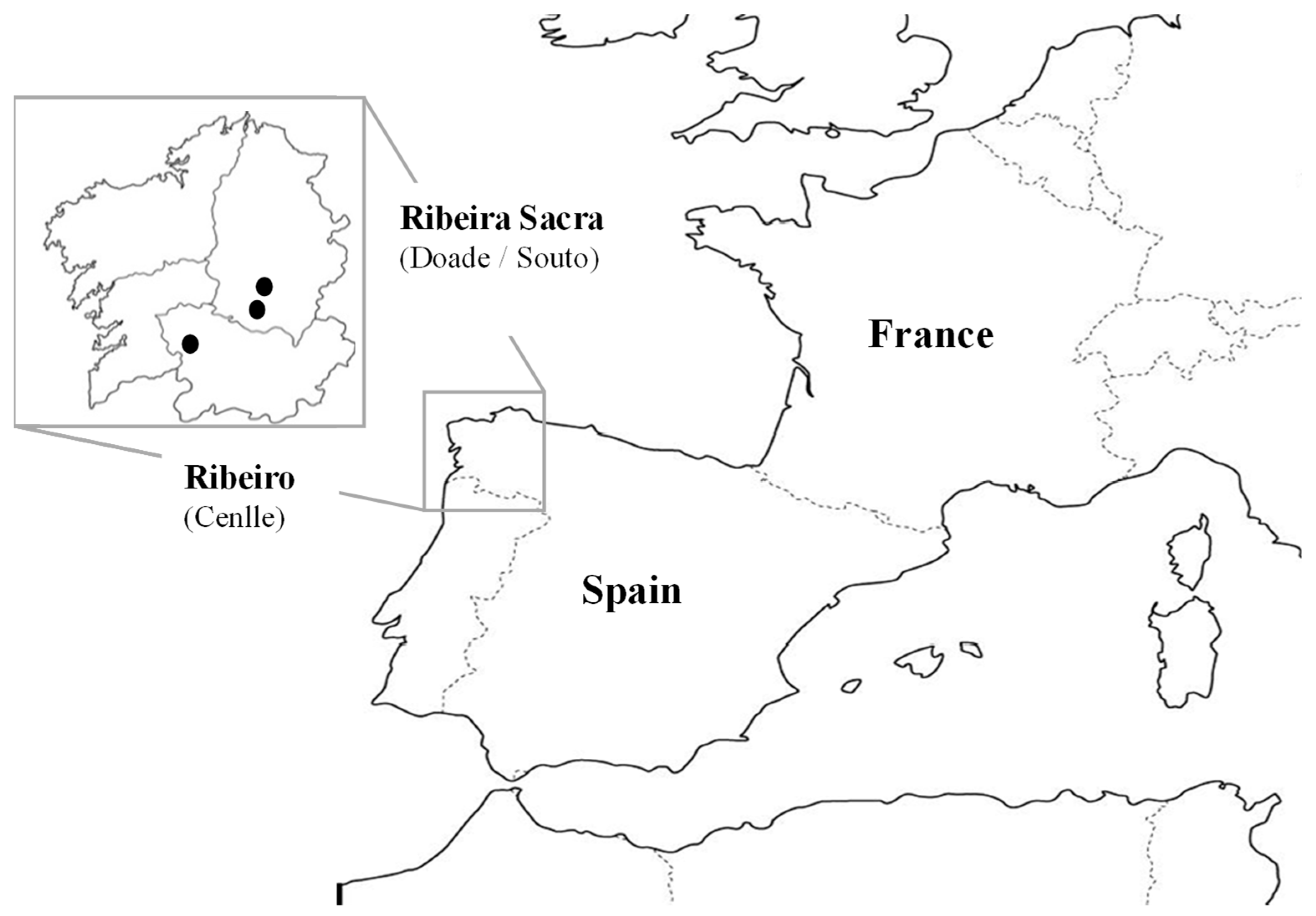
2.1. Situation of the Study Area and Climatic Characteristics
2.2. Field Experimental Tasks
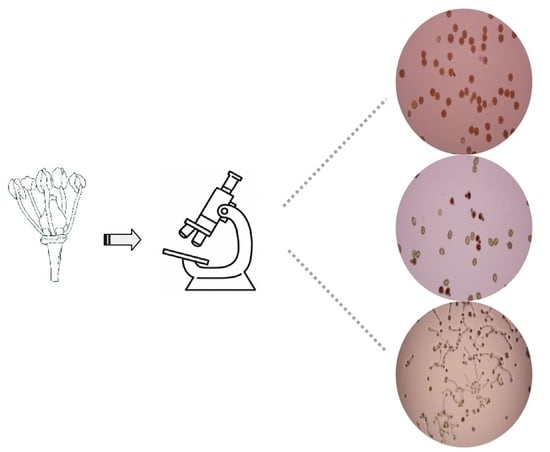
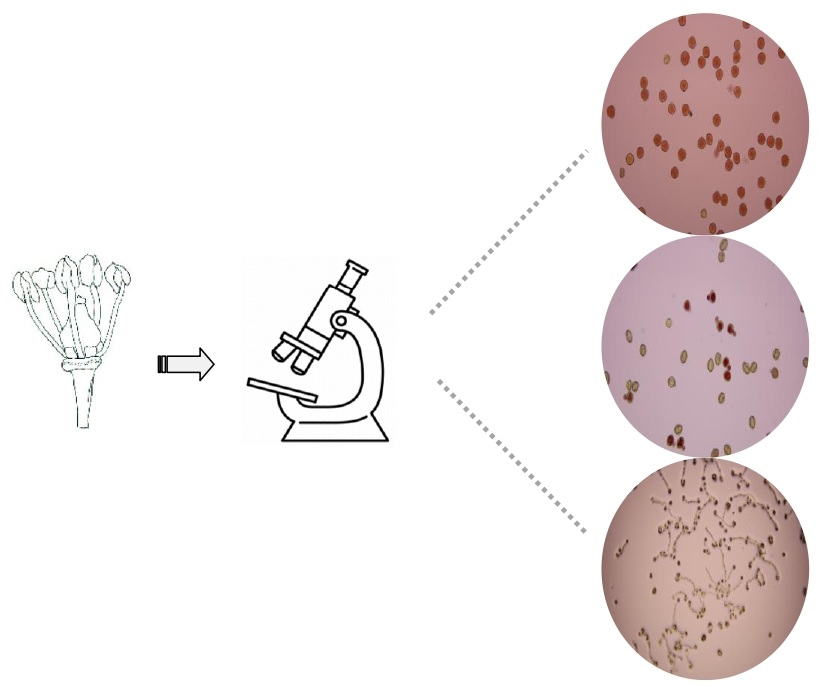
2.3. In Vitro Assays and Laboratory Analysis
2.4. Statistical Analysis
3. Results
3.1. Climatic Characterization
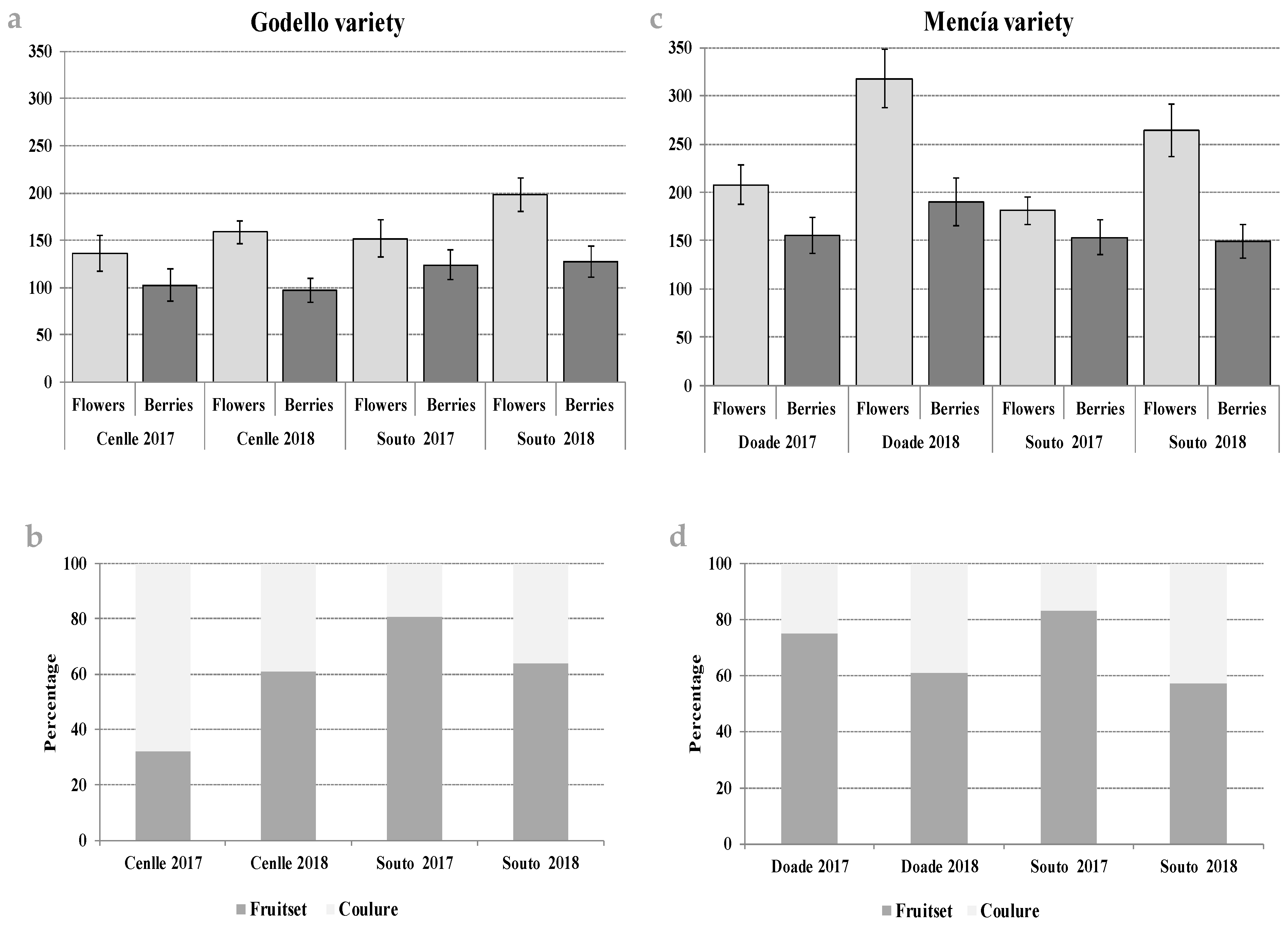
3.2. Production of Flowers and Berries: Fruitset and Coulure Indices
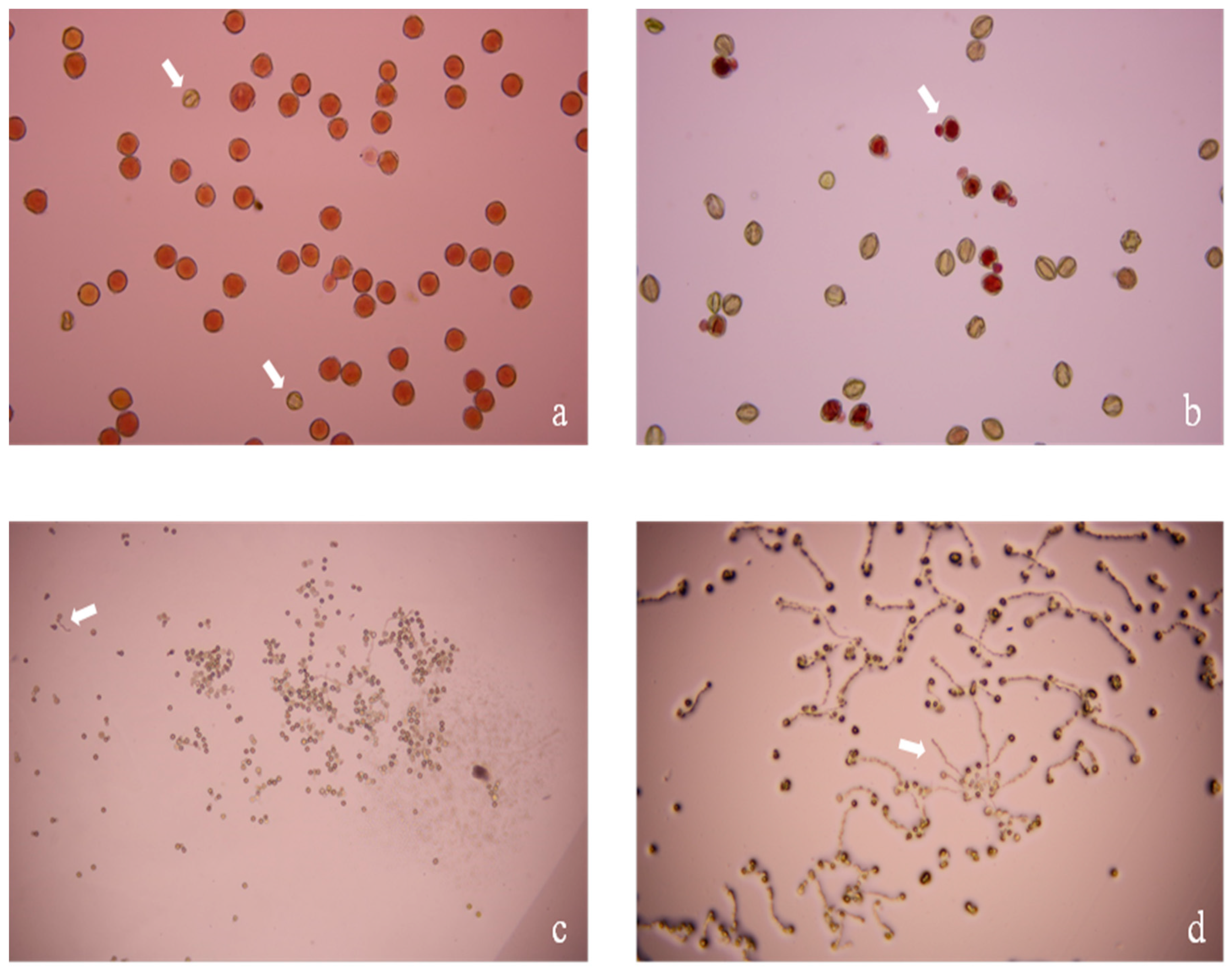
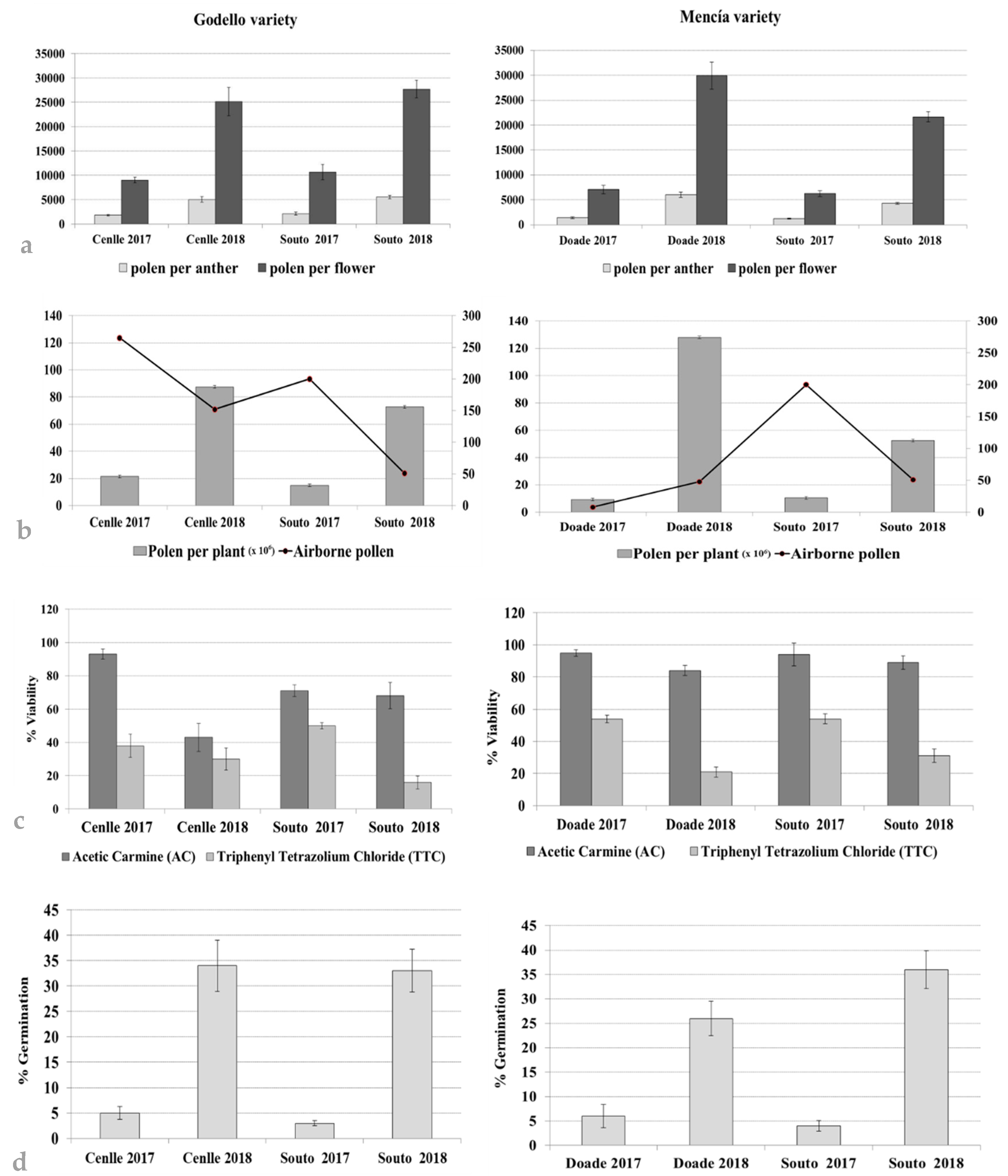
3.3. Pollen Production, Viability, and Germination Ability
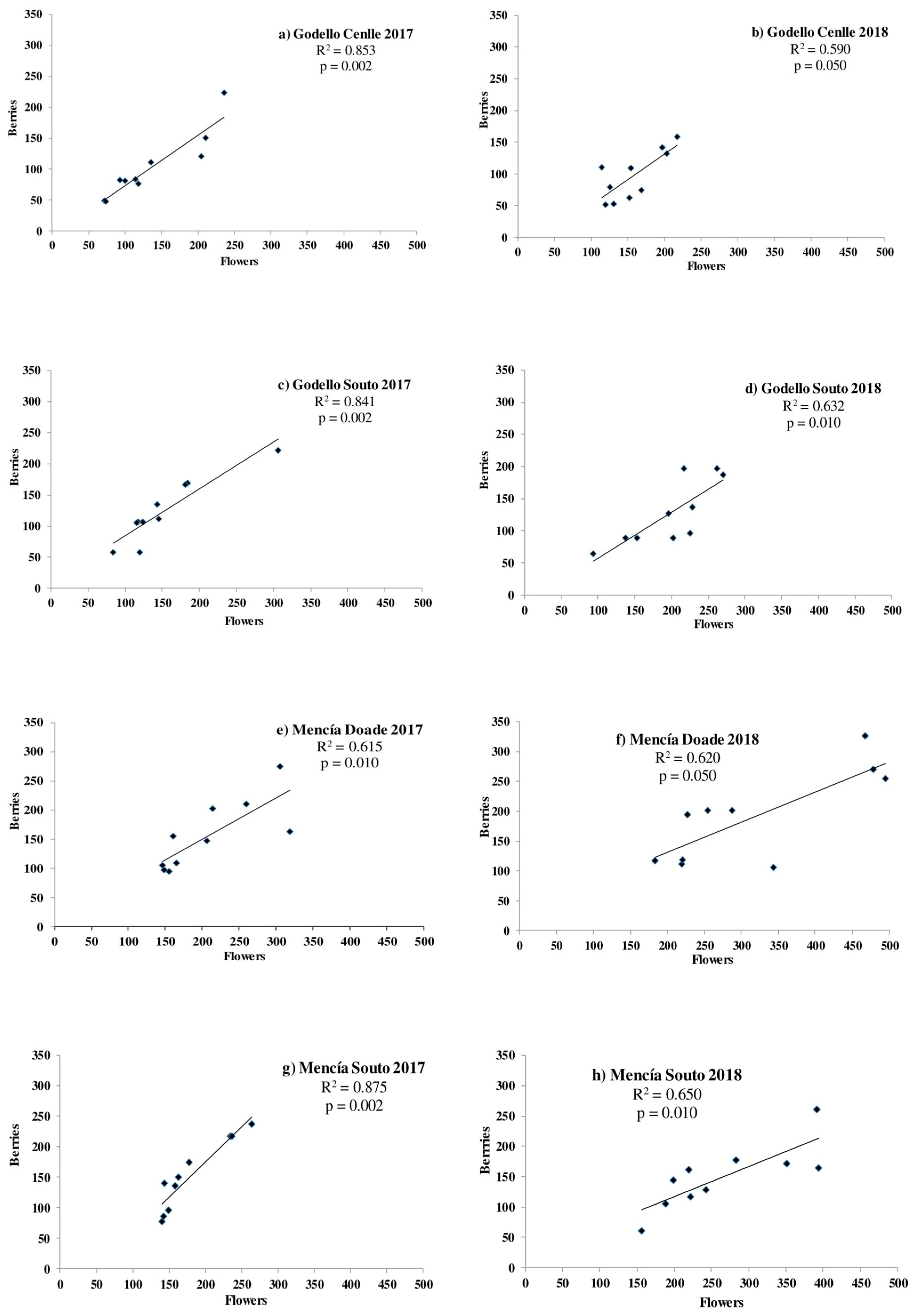
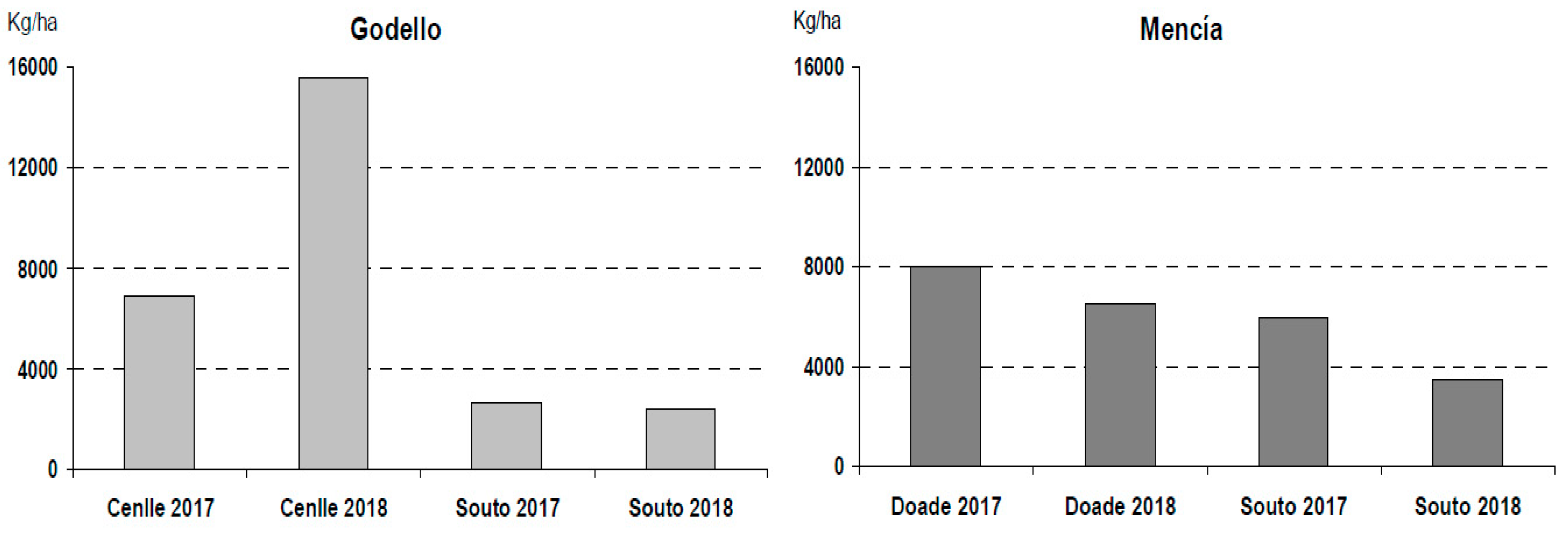
3.4. Grape Production and Its Relation with Pollen Production, Viability, Germination Rate, and Fruitset
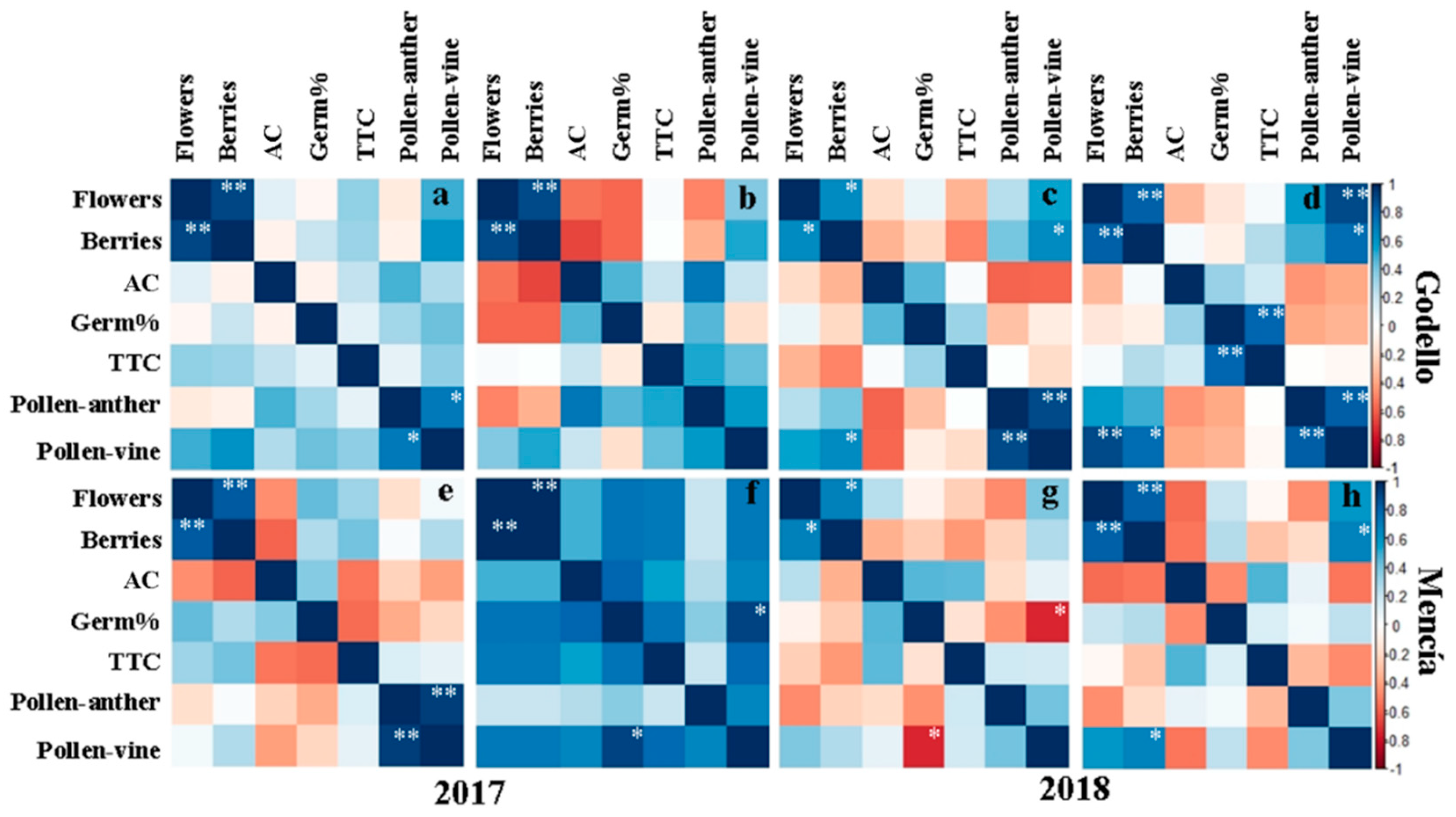
3.5. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Araujo, J.A.; Abiodun, B.J.; Crespo, O. Impacts of drought on grape yields in Western Cape, South Africa. Theor. Appl. Climatol. 2016, 123, 117–130. [Google Scholar] [CrossRef]
- Wolfe, D.W.; DeGaetano, A.; Peck, G.; Carey, M.; Ziska, L.; Lea-Cox, J.; Kemanian, A.; Hoffmann, M.; Hollinger, D. Unique challenges and opportunities for Northeastern, U.S. crop production in a changing climate. Clim. Chang. 2018, 146, 231–245. [Google Scholar] [CrossRef]
- Fraga, H.; de Cortázar Atauri, G.; Malheiro, A.C.; Moutinho-Pereira, J.; Santos, J.A. Viticulture in Portugal: A review of recent trends and climate change projections. Oeno One 2017, 51, 61–69. [Google Scholar] [CrossRef]
- Zhu, X.; Moriondo, M.; van Ierland, E.C.; Trombi, G.; Bindi, M. A model-based assessment of adaptation options for Chianti wine production in Tuscany (Italy) under climate change. Reg. Environ. Chang. 2016, 16, 85–96. [Google Scholar] [CrossRef]
- Lanyon, D.M.; Cass, A.; Hansen, D. The Effect of Soil Properties on Vine Performance; Technical Report 34/04 for CSIRO Land and Water; CSIRO Land and Water: Adelaide, Australia, November 2004. [Google Scholar]
- Disegna, E.; Coniberti, A.; Dellacassa, E. Medición de área foliar de la vid: Una herramienta para producir vinos de calidad. Hortifruticultura 2005, 4, 18–20. [Google Scholar]
- Aleixandre, J.L.; Giner, J.F.; Aleixandre-Tudó, J.L. Evaluación del efecto terroir sobre la calidad de la uva y el vino (I). Enovinicultura 2013, 20, 1–11. [Google Scholar]
- Alsafadi, K.; Mohammed, S.; Habib, H.; Kiwan, S.; Hennawi, S.; Sharaf, M. An integration of bioclimatic, soil, and topographic indicators for viticulture suitability using multi-criteria evaluation: A case study in the Western slopes of Jabal Al Arab–Syria. Geocarto Int. 2019, 34, 1–23. [Google Scholar] [CrossRef]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186. [Google Scholar] [CrossRef]
- Savary, S.; Ficke, A.; Aubertot, J.N.; Hollier, C. Crop losses due to diseases and their implications for global food production losses and food security. Food Secur. 2012, 4, 519–537. [Google Scholar] [CrossRef]
- Boso, S.; Gago, P.; Santiago, J.L.; de la Fuente, M.; Martínez, M.C. Factors affecting the vineyard populational diversity of Plasmopara viticola. Plant Pathol. J. 2019, 35, 125. [Google Scholar]
- Naegele, R.P. Evaluation of host resistance to Botrytis bunch rot in Vitis spp. and its correlation with Botrytis leaf spot. HortScience 2018, 53, 204–207. [Google Scholar] [CrossRef]
- Yildirim, Z.; Atak, A.; Akkurt, M. Determination of downy and powdery mildew resistance of some Vitis spp. Cienc. Tec. Vitivinic. 2019, 34, 15–24. [Google Scholar]
- Oliveira, M.; Arenas, M.; Lage, O.; Cunha, M.; Amorim, M.I. Epiphytic fungal community in Vitis vinifera of the Portuguese wine regions. Lett. Appl. Microbiol. 2018, 66, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Farias, S.; Riveros-Burgos, C. Modeling phenology of four grapevine cultivars (Vitis vinifera L.) in Mediterranean climate conditions. Sci. Hortic. 2019, 250, 38–44. [Google Scholar] [CrossRef]
- Cortiñas, J.A.; Fernández-González, M.; González-Fernández, E.; Vázquez-Ruiz, R.A.; Rodríguez-Rajo, F.J.; Aira, M.J. Phenological behaviour of the autochthonous godello and mencía grapevine varieties in two designation origin areas of the NW Spain. Sci. Hortic. 2020, 265, 109–221. [Google Scholar] [CrossRef]
- Kelen, M.; Demirtas, I. Fertilization Biology of Some Grape Varieties (Vitis vinifera L.). Pak. J. Biol. Sci. 2003, 6, 766–769. [Google Scholar]
- Kelen, M.; Demirtas, I. Pollen viability, germination capability and pollen production level of some grape varieties (Vitis vinifera L.). Acta Physiol. Plant. 2003, 25, 229–233. [Google Scholar] [CrossRef]
- Abreu, I.; Costa, I.; Oliveira, M.; Cunha, R.; de Castro, M. Ultrastructure and germination of Vitis vinifera cv. Loureiro pollen. Protoplasma 2006, 228, 131–135. [Google Scholar] [CrossRef]
- Dzyuba, O.F.; Shurekova, O.V.; Tokarev, P.I. On the natural polymorphism of pollen grains of Acer tataricum L. Paleontol. J. (Nauka, Russia) 2006, 40, 590–594. [Google Scholar] [CrossRef]
- Gallardo, A.; Ocete, R.; Lopez, M.A.; Lara, M.; Rivera, D. Assessment of pollen dimorphism in populations of Vitis vinifera L. subsp. sylvestris (Gmelin) Hegi in Spain. Vitis 2009, 48, 59–62. [Google Scholar]
- Caporali, E.; Spada, A.; Marziani Failla, O.; Scienza, A. The arrest of development of abortive reproductive organs in the unisexual flower of Vitis vinifera ssp. sylvestris. Plant Reprod. 2003, 15, 291–300. [Google Scholar] [CrossRef]
- Ibáñez, J.; Baroja, E.; Grimplet, J.; Ibáñez, S. Cultivated Grapevine Displays a Great Diversity for Reproductive Performance Variables. Crop Breed. Genet. Genom. 2020, 2. [Google Scholar] [CrossRef]
- Keller, M. (Ed.) Botany and anatomy. In The Science of Grapevines: Anatomy and Physiology; Elsevier: Amsterdam, The Netherlands, 2010; p. 522. [Google Scholar]
- Tello, J.; Montemayor, M.I.; Forneck, A.; Ibáñez, J. A new image-based tool for the high throughput phenotyping of pollen viability: Evaluation of inter- and intra-cultivar diversity in grapevine. Plant Methods 2018, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Baby, T.; Gilliham, M.; Tyerman, S.D.; Collins, C. Differential fruitset between grapevine cultivars is related to differences in pollen viability and amine concentration in flowers. Aust. J. Grape Wine Res. 2016, 22, 149–158. [Google Scholar] [CrossRef]
- De Storme, N.; Geelen, D. The impact of environmental stress on male reproductive development in plants: Biological processes and molecular mechanisms. Plant Cell Environ. 2014, 37, 1–18. [Google Scholar] [CrossRef]
- Pereira, H.S.; Delgado, M.; Avo, A.P.; Barao, A.; Serrano, I.; Viegas, W. Pollen grain development is highly sensitive to temperature stress in Vitis vinifera. Aust. J. Grape Wine Res. 2014, 20, 474–484. [Google Scholar] [CrossRef]
- Cristofolini, F.; Gottardini, E. Concentration of airborne pollen of Vitis vinifera L. and yield forecast: A case study at S. Michele all’Adige, Trento, Italy. Aerobiologia 2000, 16, 125–129. [Google Scholar] [CrossRef]
- Ma, W.; Wang, J.; Hu, J.; Guan, Y.; Li, Y.; Zheng, Y. Relation between changes in polyamine, protective enzyme activity and pollen vigor of tobacco in different flowering stages. Afric. J. Agric. Res. 2012, 7, 5491–5497. [Google Scholar]
- Cunha, M.; Ribeiro, H.; Abreu, I. Pollen-based predictive modelling of wine production: Application to an arid region. Eur. J. Agron. 2016, 73, 42–54. [Google Scholar] [CrossRef]
- Padureanu, S.; Patras, A. Palynological characterization, germination potential and pollen tube growth of direct producer hybrids Noah and Othello (Vitis genus). Flora 2018, 240, 58–67. [Google Scholar] [CrossRef]
- Rodríguez de la Cruz, D.; Martín Baz, A.; Sánchez Reyes, E.; Sánchez, J. Aplicaciones de la aerobiología en el cultivo de la vid. Dossier Tierras 2014, 217, 92–96. [Google Scholar]
- Sánchez Durán, S.; García Rogado, M.R.; Sánchez Reyes, E.; Rodríguez de la Cruz, D.; Sánchez Sánchez, J. Estudio aeropalinológico de Vitis vinifera L. en la zona de Toro (España) y su relación con los parámetros meteorológicos. In Proceedings of the XVII International Symposium APLE, Ourense, Spain, 7–10 July 2010; Jato, V., Aira-Rodríguez, M.J., Rodríguez-Rajo, F.J., Eds.; Palinología Fundamental y Aplicada. Universidade de Vigo: Ourense, Spain; p. 139. [Google Scholar]
- Martínez de Toda, F. Claves de la Viticultura de Calidad: Nuevas Técnicas de Estimulación y Control de la Calidad de la uva en el Viñedo; Mundi-Prensa: Madrid, Spain, 2011; p. 253. [Google Scholar]
- MAPAMA Ministerio de Agricultura y Pesca, Alimentación y Medio Ambiente. Registro de Variedades Comerciales. Available online: www.mapama.gob.es (accessed on 15 April 2020).
- Rivas-Martínez, S. Mapa de series, geoseries y geopermaseries de vegetación de España. Memoria del mapa de vegetación potencial de España, PARTE 1. Itinera Geobotanica 2005, 17, 1–436. [Google Scholar]
- Fernández-González, M.; Rodríguez-Rajo, F.J.; Escuredo, O.; Aira, M.J. Influence of thermal requirement in the aerobiological and phenological behavior of two grapevine varieties. Aerobiologia 2013, 29, 523–535. [Google Scholar] [CrossRef]
- Hirst, J.M. An automatic volumetric spore-trap. Ann. Appl. Biol. 1952, 36, 257–265. [Google Scholar] [CrossRef]
- Galán, C.; Cariñanos, P.; Alcázar, P.; Domínguez, E. Spanish Aerobiology Network (REA): Management and Quality Manual; University of Córdoba Publication Service: Córdoba, Spain, 2007. [Google Scholar]
- Galán, C.; Ariatti, A.; Bonini, M.; Clot, B.; Crouzy, B.; Dahl, A.; Fernandez-González, D.; Frenguelli, G.; Gehrig, R.; Isard, S.; et al. Recommended terminology for aerobiological studies. Aerobiologia 2017, 33, 293–295. [Google Scholar]
- Dalkiliç, Z.; Mestav, H.O. In vitro pollen quantity, viability and germination tests in quince. Afr. J. Biotechnol. 2011, 10, 16516–16520. [Google Scholar]
- De Oliveira da Silva, L.F.; Ruiz-Zambon, C.; de Oliveira, A.F.; Pio, R.; Dias-Gonçalves, E. Establishment of growth medium and quantification of pollen grains of olive cultivars in Brazil’s subtropical areas. Bragantia 2016, 75, 26–32. [Google Scholar] [CrossRef]
- Rojo, J.; Orlandi, F.; Perez-Badia, R.; Aguilera, F.; Ben Dhiab, A.; Bouziane, H.; de la Guardia, C.D.; Galan, C.; Gutierrez-Bustillo, A.M.; Moreno-Grau, S.; et al. Modeling olive pollen intensity in the Mediterranean region through analysis of emission sources. Sci. Total Environ. 2016, 551, 73–82. [Google Scholar] [CrossRef]
- Vasconcelos, M.C.; Greven, M.; Winefield, C.S.; Trought, M.C.T.; Raw, V. The Flowering Process of Vitis vinifera: A Review. Am. J. Enol. Vitic. 2009, 60, 411–434. [Google Scholar]
- Castiñeiras, P.; Vázquez-Ruiz, R.A.; Fernández-González, M.; Rodríguez-Rajo, F.J.; Aira, M.J. Production and viability of Fraxinus pollen and its relationship with aerobiological data in the northwestern Iberian Peninsula. Aerobiologia 2019, 35, 227–241. [Google Scholar] [CrossRef]
- Gaaliche, B.; Majdoub, A.; Trad, M.; Mars, M. Assessment of pollen viability, germination, and tube growth in eight Tunisian caprifigs (Ficus carica L.) cultivars. ISRN. Agronomy 2013, 2013. [Google Scholar] [CrossRef]
- Brewbaker, J.L.; Kwack, B.H. The calcium ion and substances influencing pollen growth. In Pollen Physiology and Fertilization; Linskens, H.F., Ed.; Elsevier North-Holland Publishing Co.: Amsterdam, The Netherlands, 1964; pp. 145–151. [Google Scholar]
- Koubouris, G.C.; Metzidakis, I.T.; Vasilakakis, M.D. Intraspecific variation in pollen viability, germination and ultrastructure of Olea europaea L. Afr. J. Biotechnol. 2012, 11, 13442–13446. [Google Scholar]
- Rodríguez-Riaño, T.; Dafni, A. A new procedure to asses pollen viability. Plant Reprod. 2000, 12, 241–244. [Google Scholar] [CrossRef]
- Shivanna, K.R.; Rangaswamy, N.S. Pollen Biology: A Laboratory Manual; Springer-Verlag: Berlin/Heildelberg, Germany, 1992; p. 119. [Google Scholar]
- Rejón, J.D.; Suárez, C.G.; Alché, J.D.; Castro, A.J.; Rodríguez-García, M.I. Evaluación de diferentes métodos para estimar la calidad del polen en distintos cultivares de olivo (Olea europaea L.). Polen 2010, 20, 61–72. [Google Scholar]
- May, P. Flowering and Fruitset in Grapevine; Editorial: Phylloxera and Grape Industry Board of South Australia in association with Lythrum Press; Vinehealth Australia: Adelaide, Australia, 2004; p. 119. [Google Scholar]
- Callejas, R.; Galleguillos, T.M.; Benavides, C.Z. Pérdidas de Producción Por Fallos en la Fecundación, Competencia y Anormal Desarrollo de las Bayas en vid Vinífera; CEVID Centro de Estudio de la Vid, Facultad de Ciencias Agronómicas, Universidad de Chile: La Pintana, Chile, 2004; p. 10. [Google Scholar]
- Fernández-González, M.; Escuredo, O.; Rodríguez-Rajo, F.J.; Aira, M.J.; Jato, V. Prediction of grape production by grapevine cultivar Godello in north-west Spain. J. Agric. Sci. 2011, 149, 725–736. [Google Scholar] [CrossRef]
- Pagay, V.; Collins, C. Effects of timing and intensity of elevated temperatures on reproductive development of field-grown Shiraz grapevines. Oeno One 2017, 51, 409–421. [Google Scholar] [CrossRef]
- Cortiñas, J.A.; Aira, M.J.; Fernández-González, M.; Rodríguez-Rajo, F.J.; Vázquez-Ruiz, R.A. Potential sustainable wine-growing model in the Ribeira Sacra, D.O. (NW Spain). Cienc. Tec. Vitivinic. 2018, 33, 114–145. [Google Scholar]
- Naab, O.A.; Caccavari, M.A.; Caramuti, V.E. Producción y biología del polen en algunas variedades de Vitis vinifera L. cultivadas en la Argentina. Rev. Museo Arg. Cienc. Nat. 2003, 5, 145–150. [Google Scholar] [CrossRef]
- Estévez, P.; Gil de la Peña, M.L.; Falqué, E. Potencialidade da variedade Torrontés para elaborar viños monovarietais. Cienc. Tecnol. Alim. 2002, 3, 223–228. [Google Scholar]
- Frenguelli, G.; Bricchi, E. The use of the pheno-climatic model for forecasting the pollination of some arboreal taxa. Aerobiologia 1998, 14, 39–44. [Google Scholar] [CrossRef]
- Calderón-Ezquerro, M.C.; Guerrero-Guerra, C.; Martínez-López, B.; Fuentes-Rojas, F.; Téllez-Unzueta, F.; López-Espinoza, E.D.; Calderón-Segura, M.E.; Martínez-Arroyo, A.; Trigo-Pérez, M.M. First airborne pollen calendar for Mexico City and its relationship with bioclimatic factors. Aerobiologia 2016, 32, 225–244. [Google Scholar] [CrossRef]
- Lebon, G.; Duchêne, E.; Brun, O.; Clément, C. Phenology of flowering and starch accumulation in grape (Vitis vinifera L.) cuttings and vines. Ann. Bot. 2005, 95, 943–948. [Google Scholar] [CrossRef] [PubMed]
- González-Fernández, E.; Piña-Rey, A.; Fernández-González, M.; Aira, M.J.; Rodríguez-Rajo, F.J. Prediction of Grapevine Yield Based on Reproductive Variables and the Influence of Meteorological Conditions. Agronomy 2020, 10, 714. [Google Scholar]
- Kozma, P. Exploration of flower types in grapes. In Floral Biology, Pollination and Fertilisation in Temperate Zone Fruit Species and Grape; Kozma, P., Nyéki, J., Soltész, M., Szabó, Z., Eds.; Akademiai Kiado: Budapest, Hungary, 2003; pp. 75–226. [Google Scholar]
- Cunha, M.; Abreu, I.; Pinto, P.; de Castro, R. Airborne pollen samples for early-season estimates of wine production in a Mediterranean climate area of northern Portugal. Am. J. Enol. Vitic. 2003, 54, 189–194. [Google Scholar]
- Bronner, A.; Wagner, R. Pollen et floraison chez Vitis vinifera L. Techniques de controle du pouvoir germinatif du pollen. Prog. Agric. Vit. 1997, 114, 130–139. [Google Scholar]
- Staudt, G. Pollen germination and pollen tube growth in vivo and the dependence on temperature. Vitis 1982, 21, 205–216. [Google Scholar]
- Rajasekaran, K.; Mullins, M.G. 1985. Somatic embryo formation by cultured ovules of Cabernet Sauvignon grape: Effects of fertilization and of the male gameticide toluidine glue. Vitis 1985, 24, 151–157. [Google Scholar]
- Dokoozlian, N.K. Grape Berry Growth and Development. In Raisin Production Manual; Christensen, L.P., Ed.; University of California. Agricultural and Natural Resources, Communication Services, Publication 3393: Oakland, CA, USA, 2000; pp. 30–37. [Google Scholar]
- Kliewer, W.M. Effect of high temperatures during the bloom-set period on fruit-set, ovule fertility, and berry growth of several grape cultivars. Am. J. Enol. Vitic. 1977, 28, 215–222. [Google Scholar]
- Ebadi, A.; May, P.; Sedgley, M.; Coombe, B.G. Effect of low temperature near flowering time on ovule development and pollen tube growth in the grapevine (Vitis vinifera L.), cvs Chardonnay and Shiraz. Aust. J. Grape Wine R. 1995, 1, 11–18. [Google Scholar] [CrossRef]
- Pereira, M.A.R.; Ribeiro, H.; Cunha, M.; Abreu, I. Comparison of pollen quality in Vitis vinifera L. cultivars. Sci. Hortic. 2018, 227, 112–116. [Google Scholar] [CrossRef]
- Lallana, V.H.; Elizalde, J.H.; García, L.F. Germinación y Latencia de Semillas y Yemas. Unidad Temática 11. Cátedra de Fisiología Vegetal; Universidad de Entre Ríos: Buenos Aires, Argentina, 2005; p. 22. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ribeiro | Ribeira Sacra | ||||
|---|---|---|---|---|---|
| Year | 2017 | 2018 | 2017 | 2018 | |
| Annual Average | Tmax (°C) | 23.9 | 23.7 | 21.2 | 21.2 |
| Tmin (°C) | 6.4 | 8.2 | 7.9 | 9.2 | |
| Tmean (°C) | 13.9 | 15.2 | 13.6 | 14.2 | |
| Mean RH (%) | 75.8 | 75.9 | 74.3 | 77.5 | |
| Sunshine (hours) | 5.9 | 4.9 | 5.7 | 4.6 | |
| Annual Total | Rainfall (L/m2) | 814.5 | 775.2 | 693.9 | 804.4 |
| Maximum | Daily rainfall (L/m2) | 100.8 | 39.4 | 64.9 | 39.2 |
| Date | Dec. 10th | Feb. 28th | Dec. 10th | Mar. 11th | |
| GODELLO | ||||
| Cenlle | Cenlle | Souto | Souto | |
| 2017 | 2018 | 2017 | 2018 | |
| Flowers/Berries | 0.903 ** | 0.636 * | 0.927 ** | 0.812 ** |
| Pollen Anther/Pollen Vine | 0.718 * | 0.894 ** | 0.580 | 0.827 ** |
| Pollen Vine/Berries | 0.600 | 0.636 * | 0.522 | 0.763 * |
| Pollen Vine/Flowers | 0.491 | 0.539 | 0.371 | 0.891 ** |
| Germ%/TTC | 0.104 | 0.317 | −0.279 | 0.794 ** |
| MENCÍA | ||||
| Doade | Doade | Souto | Souto | |
| 2017 | 2018 | 2017 | 2018 | |
| Flowers/Berries | 0.842 ** | 0.681 * | 0.964 ** | 0.818 ** |
| Pollen Anther/Pollen Vine | 0.934 ** | 0.401 | 0.657 | 0.389 |
| Pollen Vine/Berries | 0.262 | 0.267 | 0.714 | 0.672 * |
| Germ%/Pollen Vine | −0.217 | −0.742 * | 0.912 * | 0.226 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortiñas Rodríguez, J.A.; Fernández-González, M.; González-Fernández, E.; Vázquez-Ruiz, R.A.; Rodríguez-Rajo, F.J.; Aira, M.J. Potential Fertilization Capacity of Two Grapevine Varieties: Effects on Agricultural Production in Designation of Origin Areas in the Northwestern Iberian Peninsula. Agronomy 2020, 10, 961. https://doi.org/10.3390/agronomy10070961
Cortiñas Rodríguez JA, Fernández-González M, González-Fernández E, Vázquez-Ruiz RA, Rodríguez-Rajo FJ, Aira MJ. Potential Fertilization Capacity of Two Grapevine Varieties: Effects on Agricultural Production in Designation of Origin Areas in the Northwestern Iberian Peninsula. Agronomy. 2020; 10(7):961. https://doi.org/10.3390/agronomy10070961
Chicago/Turabian StyleCortiñas Rodríguez, J. Antonio, María Fernández-González, Estefanía González-Fernández, Rosa A. Vázquez-Ruiz, F. Javier Rodríguez-Rajo, and María Jesús Aira. 2020. "Potential Fertilization Capacity of Two Grapevine Varieties: Effects on Agricultural Production in Designation of Origin Areas in the Northwestern Iberian Peninsula" Agronomy 10, no. 7: 961. https://doi.org/10.3390/agronomy10070961
APA StyleCortiñas Rodríguez, J. A., Fernández-González, M., González-Fernández, E., Vázquez-Ruiz, R. A., Rodríguez-Rajo, F. J., & Aira, M. J. (2020). Potential Fertilization Capacity of Two Grapevine Varieties: Effects on Agricultural Production in Designation of Origin Areas in the Northwestern Iberian Peninsula. Agronomy, 10(7), 961. https://doi.org/10.3390/agronomy10070961