1. Introduction
Legumes have a prominent role in sustaining agricultural productivity [
1,
2]. Pulses, legumes, and grasslands are grown because of their ability to interact with nitrogen-fixing bacteria [
3], and they can also be used in rotation systems to manage pests, diseases, and weeds [
4]. Small quantities of arable land and intensive agro-industrialization have led to the cultivation of crops on the same land without crop rotation. Such exhaustive peanut production is particularly widespread in the subtropical regions of Asia. The consecutive monoculture may, however, have caused a steady decline in productivity and performance, and increased disease susceptibility [
5].
Castro et al. [
6] reported increased yield when peanut crops were inoculated with rhizobium, but Walker et al. [
7] and Huang et al. [
8] noted that continuous cropping of peanut on the same piece of land resulted in reduced yield even when there was evidence of rhizobium inoculation. Daimon et al. [
9] showed that fertilization with nitrate prevents root hair infection and initiation, development, and formation of nodules. Molecular analysis of peanut nodule formation suggests that suppression of root nodules is related to feedback regulation (autoregulation) by nitrates [
10,
11]. Moreover, peanut crop roots secrete a variety of primary metabolites of carbon, such as sugars, amino acids or phenolic acids, and more complex secondary compounds involved in plant defense and stimulatory or inhibitory interactions with other soil microorganisms [
12,
13,
14].
The efficient development of peanut depends on its association with rhizobia which nodulate it, to enhance its nitrogen-fixing ability.
Bradyrhizobium spp. is an agronomically significant gram-negative bacterium capable of forming root nodules on peanut roots and fixing atmospheric nitrogen [
15]. Biological nitrogen-fixing (BNF) bacteria, such as
Rhizobium and
Bradyrhizobium species, produce auxins, cytokinins, abscisic acid, vitamins, riboflavin, lipochitooligosaccharides, and lumichrome that stimulate plant growth and nitrogen-fixation in legumes [
16,
17]. Bacteria that fix soil nitrogen are important contributors to primary productivity and their population is significantly affected by agrochemical applications. They fix large quantities of nitrogen biologically converting approximately 200 million tons of nitrogen to ammonia globally every year [
18]. However, since the introduction of seed treatments for the prevention of fungal diseases, inoculation of peanuts with beneficial microorganisms has not been feasible especially in maintaining the viability of seeds and inoculants [
19].
Plants use endophytic fungal products to overcome various abiotic and biotic stress conditions [
20]. Endophytes colonizing and residing in internal plant parts are efficient in suppressing a broad variety of diseases in plants and they also promote plant growth [
21].
Trichoderma species in several crops, including peanut [
22], are efficient in controlling soil- and seed-borne fungal diseases [
23]. The potential of
T. harzianum as biocontrol agent in peanuts has been widely studied for reducing aflatoxin contamination by
Aspergillus flavus [
24], crown rot by
A. niger [
25], stem rot by
Sclerotium rolfsii and
Rhizoctonia solani [
26], and brown root rot by
Fusarium solani [
27].
Trichoderma’s existence within the roots of treated crops has rendered it a mycorrhizal organism [
28]. Specializing factors for the biocontrol behavior of
Trichoderma include space and nutrient competitiveness, and the development of diffusible and/or reactive antibiotics and hydrolytic enzymes such as chitinase and β-1,3-glucanase [
23]. Several mechanisms adopted by
Trichoderma, such as the solubility of insoluble minor soil nutrients [
29], synthesis of growth hormones [
30], and increased ingestion and translocation of less available soil minerals [
31,
32,
33], have been suggested to influence the development of plants. Some studies also reported the antagonistic properties of these secondary metabolites which act as growth promoting factors. Horace et al. [
34] documented the isolation, recognition, and biological activity of 6-pentyl-apyrone—a secondary metabolite produced by
T. harzianum that has been involved as regulators of plant development. It is believed that secondary metabolites of
Trichoderma can serve as auxin-like compounds that usually have the optimum activity between 10
−5 and 10
−6 M, but can have inhibitory effect at higher concentrations [
35,
36,
37].
The survival and role of soil microorganisms are important to the soil ecosystems because they participate in key processes, such as the cycling of minerals, organic matter breakdown, soil structure formation, and toxins elimination [
38,
39]. Li et al. [
40] noted that the continuous secretion of exudates by peanut roots affected microbial diversity in the soil but did not contribute to the accumulation of phenolic acids levels which could be phytotoxic. Continuously secreted substances from peanut roots into the soil can stimulate the population of fungal pathogens and reduce the population of beneficial fungi, which could lead to aggravated disease of monocultural peanuts [
41]. The use of nitrogen-fixing bacteria as biofertilizer should, therefore, be embraced to leverage various positive effects on land [
42] whereas endophytes living and colonizing in inner plant habitats have been shown to be efficient in stimulating plant growth and depressing disease in a wide range of plants. This greenhouse study to understand the compatibility of
Bradyrhizobium and
Trichoderma on growth, development and yield of
Arachis hypogaea L. was therefore based on the concept of enhancing agro-ecosystem with minimal dependence on agrochemical and energy inputs, in which synergetic microbial interactions provide the system with the mechanism to support soil fertility and crop production functions.
2. Materials and Methods
2.1. Peanut Cultivar and Microbial Strains Used
Peanut (Arachis hypogaea) seeds of variety Valencia C were obtained from the College of Agricultural, Consumer and Environmental Sciences, New Mexico State University. Commercially available endophytic fungal spores of “Trichoderma harzianum” ExceedHSD (1 × 108 spores/mL) was obtained from Vision Biologicals. The potting mixture, 2:2:1 topsoil: perlite: peat moss, was inoculated with 20 mL of 1% of the endophyte inoculum diluted with deionized water per pot. The potting mixture was subjected to a temperature of 100 °C under tarp cover for three weeks to ensure a complete elimination of soil borne pathogens. Treatment with the endophyte inoculum was performed after the potting medium conditioned to the normal greenhouse temperature of 25 °C.
Commercially available nitrogen-fixing rhizobium species Bradyrhizobium spp. (America’s Best Inoculant®) was obtained from Advanced Biological marketing. The peanut seeds were inoculated with 10% of the inoculum diluted with deionized water.
2.2. Viability of Inoculated Seeds In Vitro
The ability of the microorganisms to exhibit any kind of antagonistic properties during germination was evaluated in vitro by inoculating the sterilized peanut seeds with the above-specified concentrations of the inoculum. This included four different treatments in two trials with 10 seeds in each treatment and allowed to germinate in moist towel paper. The untreated seeds were considered the control (S), while the other treatments were seeds treated with Bradyrhizobium only (S + B), seeds with Trichoderma only (S + T), and seeds treated with both Bradyrhizobium and Trichoderma (S + B + T). To study the effects of these treatments on the germination of peanuts 10 days after inoculation (DAI), all the seeds with emerged radicle and open cotyledons were considered as germinated and their fresh biomass was recorded.
2.3. Greenhouse Studies to Assess the Microorganism Effect on Growth and Development
Peanut seeds were planted in 28 cm diameter by 23 cm height pots, filled with the soil mixture specified above. All the treatments were arranged in randomized complete block design (RCBD) to compensate and/or account for light gradient within the greenhouse as the pots were placed on one side of the greenhouse only. The greenhouse was maintained under natural light conditions and 60–70% RH. The temperatures varied from 28–35 °C during the day and 20–24°C in the night. There were four treatments with 36 pots per treatment in three replications for a total of 432 pots, with three seeds sown in each pot. Each individual treatment was placed in nine rows of pots with four pots per row. The four treatments included: seed only (S); seeds inoculated with Bradyrhizobium and Trichoderma (S + B + T); seeds inoculated with Bradyrhizobium only (S + B); seeds inoculated with Trichoderma only (S + T).
Bradyrhizobium inoculum was sprayed on the seeds and then sowed immediately after drying, whereas Trichoderma inoculum was added directly to the potting mixture after sowing. Plants were irrigated with deionized water so that no extraneous nutrients were added to the growing medium. The crops were harvested after 125 days of sowing and the dry biomass of plant parts along with number of survived plants and mature pods were recorded.
2.4. Data Analysis
The following parameters were measured for statistical analyses.
Germination percentage (%).
Seedling and plant heights (cm).
Chlorophyll concentration using SPAD-502 Plus chlorophyll meter manufactured by Konica Minolta Inc., Japan (SPAD Units).
Dry biomass of the harvested plant parts (g).
Number of matured pods per pot.
Number of harvested plants per pot.
Seed biomass per treatment and 100-seed weight per treatment.
In addition, data collected from these parameters were used to calculate the following indices:
The data were analyzed by Analysis of Variance (ANOVA) in SAS version 9.4, and treatment means were compared using the Student–Newman–Keuls (SNK) method at 5% significance level.
4. Discussion
Legume plants are closely linked to microbes that affect the plant features related to the provision of nutrients, plant growth, defenses, and abiotic stress [
47]. It has been suggested that the thorough use of soil microbial potential is essential for better nutrient management [
4]. The current research evaluated the effect of co-inoculations of nitrogen-fixing rhizobia and plant growth-promoting (PGP) endophyte in the enhancement of peanut germination, growth, development and yield under greenhouse conditions. The data show that the co-inoculation of
Bradyrhizobium and
Trichoderma had enhanced effect on growth, development, and yield of peanut.
The results from the in vitro germination tests were consistent with the observations of Yedidia et al. [
48] on cucumber. They observed that
Trichoderma enhanced seedling emergence and dry weight of cucumber five days after inoculation. The effect of
Trichoderma on peanut germination was statistically significant from the control and, in fact, the co-inoculation of
Trichoderma and
Bradyrhizobium may be antagonistic to peanut germination (
Table 6). This result makes sense since seed germination requires primarily abiotic conditions of conducive temperature for the species, moisture and oxygen. These conditions also favor the growth of some microbes which could explain the germination enhancement observed in this study.
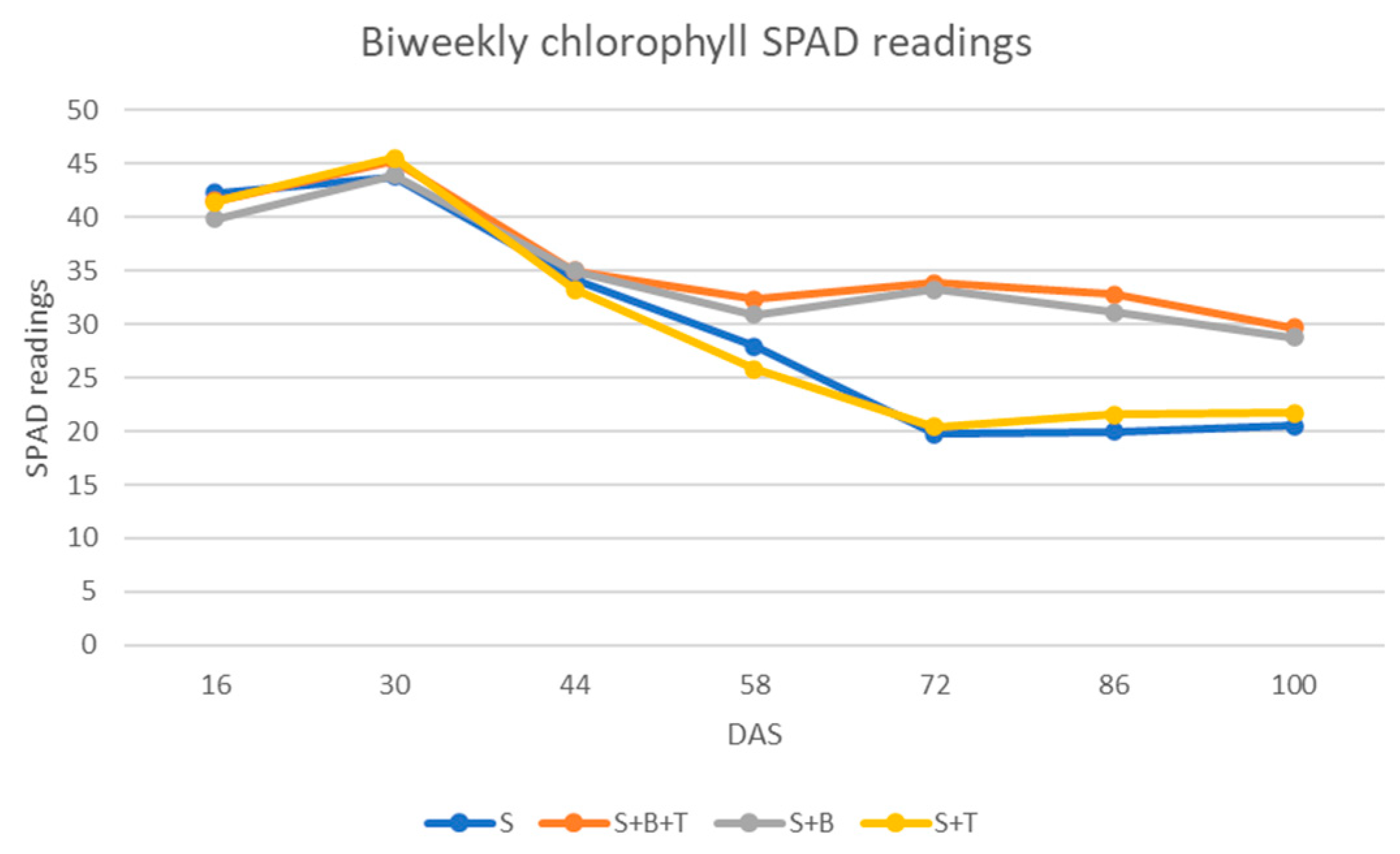
From the greenhouse studies the trends in relative plant height (RPH) synchronized with the physiological maturity of the crop and the highest RPH was recorded 76 DAS and thereafter, height growth declined gradually indicating crop reproductive maturity (
Figure 3). The efficiency of nitrogen fixation in peanuts resulted in the accumulation of nitrogen in plant tissues which in turn reflected the synthesis of chlorophyll [
49]. SPAD readings for the
Bradyrhizobium inoculated treatments are higher compared to others and this might be because of the
Bradyrhizobium’s fundamental function of biological nitrogen fixation [
50] (
Table 3). Treatments co-inoculated with
Bradyrhizobium enhanced the synthesis of chlorophyll and height growth, resulting in significant increases in plant dry weight and yield [
51]. Although the co-inoculated treatment had similar statistical effects as the treatment with
Bradyrhizobium alone in all measured growth and development parameters of peanut except seed HI, but the interactive test showed that co-inoculation had an advantageous effect on plant biomass which can be correlated to the PGP- characteristics of
Trichoderma such as stimulating substantial and vigorous plant roots and synthesis of plant hormones (31,32) along with biological nitrogen fixation by
Bradyrhizobium (see
Table 6). This postulation supports the highest mean biomass of the shoot, root, pod, seed, and shell in the co-inoculated treatment compared to the other treatments. Although the results of the 100 seed weight, shelling percentage, seed HI and pod HI showed no statistical difference between the treatments, the interactive effects of co-inoculation with
Bradyrhizobium and
Trichoderma had qualitative value in peanut. This is an indication of added market value to yield when peanut cropping adopts co-inoculation with these microbes in cultural practices. Furthermore, seed HI was significantly higher in the co-inoculation treatment than the treatment with
Bradyrhizobium alone. It is important to note the sensitivity of Colby’s [
46] mathematical interactive method on the qualitative value of the crop. Statistical analysis showed no significant difference on the qualitatively computed information about the species but the slight differences between the treatments were discernable by Colby [
46].
Although Badawi et al. [
51] concluded that co-inoculation resulted to increased peanut growth and development in field conditions, this study, performed in controlled pathogen free conditions, confirms their results. Additionally, this study revealed that plant growth promotion is not a common function of
Trichoderma [
52].
Nieto-Jacobo et al. [
53] demonstrated that the production of IAA was dependent on
Trichoderma strains as influenced by different external stimuli. In addition, various
Trichoderma species have been shown to affect plant growth and the growth of other pathogens in conducive soil environments [
53,
54,
55,
56,
57]. However, a range of effects by
Trichoderma secondary metabolites have been documented [
54]. These effects range from no response to sensitive response by plants.
The positive interaction of
Bradyrhizobium and
Trichoderma increased the various yield attributes when compared to the control, as indicated by the interactive effects of these microorganisms (
Table 6). Zhang et al. [
58] noted that
Trichoderma inoculation increased the secretion of flavonoids by peanut roots and this induced rhizobial nod genes leading to improved
Bradyrhizobial colonization. Crops co-inoculated with
Bradyrhizobium and
Trichoderma, and those inoculated with
Bradyrhizobium alone increased the biomass and yield of the species. Increase in biomass and yield demonstrate the compatibility between microorganisms and the synergism among them to promote and enhance crop growth, development, and yield.
5. Conclusions
This study assessed the growth promoting properties of co-inoculation of microorganisms in peanut crop production. While co-inoculations are not a usual practice, this work confirms that co-inoculation with Bradyrhizobium and Trichoderma enhanced growth, development, and yield in Arachis hypogaea. The inoculations improved growth attributes of the species, such as chlorophyll content, relative height growth, survival rate, biomass, and yield, as well as parameters such as 100 seed weight, shelling percentage, seed HI and pod HI. Moreover, the results of the study showed that the beneficial effects of co-inoculation occurred at seedling growth stage, leading us to conclude that the timing for co-inoculation of peanut with Bradyrhizobium and Trichoderma should be performed at seedling emergence. The findings of this study gave a better understanding of the effects of trilateral symbiotic association between plant–rhizobia–endophyte in leguminous crops. These associations will help in developing agro-ecosystems with minimal dependence on agrochemical inputs, and they simulate natural ecological interactions between crop plants and soil organisms.






{kind=link}
{kind=link}
{kind=link}