Abstract
Abiotic stresses such as drought, extreme temperature, and salinity can negatively impact seed germination and plant growth and have become major limitations to crop production. Most crops are vulnerable to abiotic stress factors during their early growth phase, especially during seed germination and seedling emergence. Rapid crop seed germination and seedling establishment is known to provide competitive advantages over weeds and improve yields. Seed osmopriming is defined as a pre-sowing treatment in which seeds are soaked in osmotic solutions to undergo the first stage of germination, but radicle protrusion has not occurred. The process of osmopriming involves prior exposure of seeds in low-water-potential solutions. Osmopriming can generate a series of pre-germination metabolic activities, increase the antioxidant system activities, and prepare the seed for radicle protrusion. Polyethylene glycol (PEG) is a popular osmopriming agent that can alleviate the negative impacts of abiotic stresses. This review summarizes research findings on crop responses to seed priming with PEG under abiotic stresses. The challenges, limitations, and opportunities of using PEG for crop seed priming are discussed with the goal of providing insights into future research towards effective application of seed priming in crop production.
1. Introduction
Abiotic stresses are a common cause of crop yield reduction worldwide. Yield loss may occur when crops suffer from various abiotic stresses such as cold, heat, drought, freezing, flooding, heavy metals, UV-light, and mineral deficiency [1,2,3,4,5,6]. Abiotic stresses may occur individually, sequentially, or concurrently. However, plants may suffer greater detrimental effects when they are exposed to multiple abiotic stresses, such as cold and drought, heat, and heavy metal, as well as drought and heat, than when they are exposed to each individual stress [7,8,9,10].
In recent years, a growing amount of research has been conducted to investigate the impact of multiple stress factors on crop growth and productivity [7,11,12,13,14]. Various techniques such as proper agronomic practices (e.g., pest management, soil fertility management, and seed priming), traditional breeding, and modern biotechnology have been employed to enhance crop tolerance to abiotic stresses [15,16,17]. Traditional breeding methods such as hybridization and selection, as well as mutation breeding, have contributed considerably to the generation of stress-tolerant crops [16,18,19,20,21,22]; however, the process is fairly time-consuming. The ‘omic’ biotechnologies (e.g., transcriptomics, proteomics, and genomics) can be used to identify the genes that are associated with stress tolerance in crops. The identified genes can be directly introduced into the elite crop varieties or silenced to make a transgenic plant, which is less time-consuming than traditional breeding [16,23,24,25]. Transgenic crops with improved stress tolerance have been successfully developed and cultivated in field conditions [26]. Nevertheless, transgenic technologies are not readily accepted in many countries [27,28].
Agronomic management approaches such as appropriate fertilization, irrigation, exogenous application of biostimulants, and/or selection of crop varieties have been shown to be valuable in mitigating the adverse effects of abiotic stresses [16,29,30]. Among these management approaches, seed priming is a common tactic to protect various vegetable and row crops against unfavorable environmental stresses without considerably affecting crop fitness and productivity [4,31,32,33]. Primed seeds generally show an improved germination rate, uniform germination, and early emergence [5,17,34,35]. During priming, seeds are hydrated in low water potential solutions to initiate germination, but radical protrusion through the testa has not yet happened [34,36]. When seeds are primed, the water supply is controlled at a level below that needed to complete seed germination, but is enough to activate a series of metabolic physiological reactions related to the initial germination [4,34,35,37,38,39,40,41].
Osmopriming is a commonly adopted priming technique and offers a highly attractive solution for improving seed germination performance and crop stand establishment [34,35]. A variety of chemicals, such as CaCl2, KNO3, KCl, K3PO4, NaCl, PEG, and mannitol, have been examined as potential osmopriming agents [5,35]. Among these, PEG is the most commonly used priming osmoticum. PEG is chemically inert and does not impose damaging effects on seed embryos [42]. In addition, PEG is non-damaging to proteins and does not penetrate seed tissues due to its large molecular size [42,43,44,45,46].
In most cases, PEG-primed plants exhibit positive effects on seed germination, seedling establishment, and yield, but the benefit is variable depending on several factors such as crop species and stress type [47,48,49,50]. Previous reviews on this topic have mainly focused on seed priming with various agents [4,35,51], but priming with PEG against abiotic stresses has not been well analyzed. Here, we review PEG-primed crop tolerance to drought, suboptimal temperature, salinity, or combined stresses. The effectiveness of PEG priming to improve seed germination and seedling establishment under adverse environmental conditions, and associated mechanisms are discussed by stress type with the aim of utilizing PEG priming in minimizing adverse environmental impacts on crop production and providing insights for future research.
2. Consequences of Abiotic Stresses and Their Mitigation through PEG
2.1. Drought Stress
Drought stress is a common abiotic stress affecting crop productivity [1,52]. Oxidative stress accompanies almost all abiotic stresses and occurs as a result of reactive oxygen species (ROS) in plants [53]. Under adverse conditions, plants increase the production of ROS including hydrogen peroxide (H2O2), hydroxyl radicals (-OH), superoxide radicals (O2−), and singlet oxygen (-O2) [5,54,55]. These ROS interact with cellular constituents, leading to a series of oxidative damages on carbohydrates, chlorophyll, lipids, DNA, and protein [1,54,55]. Water deficit also causes other damaging effects on plant growth including epinasty, stomatal closure, and decreased photosynthesis [1,5,56].
As shown in Table 1, drought stress considerably decreased the germination performance of asparagus (Asparagus officinalis L.) [32], barley (Hordeum vulgare L.) [31], celery (Apium graveolens L.) [57], cumin (Cuminum syminum L.), rice (Oryza sativa L.) [58,59], sorghum (Sorghum bicolor L. Moench) [5], and wheat (Triticum aestivum L.) [2]. However, under the conditions of drought stress, the above-mentioned crops pretreated with PEG have shown improved seed germination and seedling establishment [31,57,60,61,62,63,64].

Table 1.
A summary of published reports on the effectiveness of PEG priming plants for abiotic stress tolerance in crops.
In response to abiotic stress, plants utilize a complex antioxidant defense machinery that protects them against the damaging effect caused by oxidative stress [5,7,53]. The capacity of the antioxidant defense system comprises enzymatic and non-enzymatic antioxidant systems [53]. The main free radical scavenger of enzymatic antioxidants includes ascorbate peroxidase (APX), catalase (CAT), glutathione reductase (GR), guaiacol peroxidase, glutathione reductase (GR), glutathione-S-transferase, glutathione peroxidase (GPX), monodehydroascorbate reductase, and superoxide dismutase (SOD), while the main non-enzymatic antioxidants include ascorbic acid (vitamin C), carotenoids, glutathione, and tocopherols (vitamin E) [53,65,66].
PEG priming caused a rapid enhancement of some antioxidant enzymes in seedlings exposed to drought stress, thus mitigating the detrimental effect on seed germination and stand establishment [5,58,67]. For example, PEG-primed rice exhibited higher seed germination and seedling growth rates with higher GPX activity compared to unprimed plants [58]. PEG-primed rice seedlings exhibited elevated levels of MnSOD in drought conditions [58]. Zhang et al. [67] reported that PEG-primed perilla mint (Perilla frutescens L. Britt) increased protective enzyme activities of CAT, peroxidase (POD), and SOD in plant leaves. In other research, Zhang et al. [5] found that unprimed sorghum seedlings demonstrated increased electrolyte leakage and O2− content, and decreased membrane stability under drought conditions; PEG-primed sorghum seedlings exhibited less lipid peroxidation, improved cell membrane stability, and enhanced activities of APX, CAT, POD, and SOD. Overall, previous evidences collectively indicate that enhanced drought tolerance in PEG-primed plants is likely due to elevated antioxidant activities that restricted the accumulation of ROS [5,9,53,65,66,67].
2.2. Temperature Stress
Plants display optimal germination and emergence at an ideal range of temperature to which they have adapted [68,69,70,71,72]. In contrast, under unfavorable temperature conditions, seed germination and seedling establishment are negatively affected [70,73,74]. Temperature stress can be classified into low positive temperature stress (chilling), negative temperature stress (frost), or high temperature stress (heat) [70]. A great amount of research has been performed with the objective of improving crop performance under suboptimal temperature conditions [68,69,71,72]. However, temperature stress is still one of the major abiotic factors limiting crop productivity.
Seed priming with PEG resulted in earlier and synchronized seed germination in various crops such as alfalfa, cereal, turfgrasses, and vegetable crops upon exposure to low or high temperature stress (Table 1) [33,57,62,64,75,76,77,78,79,80]. For instance, seed priming with PEG effectively promoted seed germination and seedling establishment of soybean [Glycine max (L.) Merr.] [81] and masson pine (Pinus massoniana L.) [82] under negative temperature stress, as well as carrot [78] and leek [80] under high temperature stress. Priming benefits are more pronounced under temperature stress than in optimal temperature conditions [35,51,83]. For instance, Patanè et al. [83] reported that the beneficial effect of PEG priming in sweet corn was only evident when plants were exposed to the suboptimal temperatures rather than optimal conditions.
Priming with PEG rapidly enhanced α-amylase activity and increased fructose and glucose content in rice [75]. Plants primed with PEG exhibited improved α-amylase and/or β-amylase activities, which facilitated starch degradation and sugar accumulation, leading to a greater respiration rate, seed viability, seed germination rate, and seedling establishment than unprimed plants [9,12,72,84,85,86,87,88]. Nevertheless, excessive priming, even in an optimal PEG solution, may disrupt α-amylase activity, and thus lower seed germination rate and cause abnormal plumule and radicle growth [89,90]. It has been suggested that appropriate priming regimes including priming solution concentration, priming duration, and priming temperature can all influence the effectiveness of seed priming [4,51,91,92,93,94].
Seed priming with PEG at low temperature conditions facilitated the differentiation and duplication of mitochondria [95]. The increased mitochondrial enzyme activity might help improve the enzymatic activities involved in seed reserve mobilization, such as fats, proteins, and sugars [96,97,98], enhancing the tolerance to chilling injury [99]. Priming with PEG also enhanced the enzymatic activities of acid phosphatase and esterase, as well as RNA synthesis in various crop species such as celery, onion (Allium cepa L.), lettuce, soybean, and sweet corn (Zea mays L.) [100].
A review of current literature indicates that antioxidant machinery has not been extensively studied for PEG-primed plants under the condition of temperature stress. Bailly et al. [101] noted that CAT played an important role in promoting sunflower (Helianthus annuus L.) seed germination after priming with PEG. Recent evidences indicated that other osmotica can significantly enhance the enzymatic activity in plants growing under low temperature stresses [99,102,103,104,105]. For instance, Guan et al. (2009) noted that seed priming with chitosan enhanced CAT and POD activities, along with faster germination rate, increased shoot and root length, and overall plant biomass in two corn cultivars with distinct chilling stress tolerance. Kaur and Goyal [104] recently reported that antioxidants play a key role in improving tolerance of Egyptian clover (Trifolium alexandrinum L.) to low temperature stress after priming with salicylic acid.
Although PEG-primed plants exhibited superior tolerance to abiotic stresses in a wide range of crop species [4,35], the use of PEG as an osmotica is not recommended for the seeds of high-tannin sorghum cultivars [106]. Tannins in the seed coat are desirable and protect seeds from weathering and reduce the vulnerability of seeds to birds, insects, and mold attacks [107]. High tannin content may also help improve seedling emergence under suboptimal temperature conditions [107]. However, tannins exhibit higher binding affinity with PEG (Silanikove et al., 1996). Patanè et al. [106] reported that after priming with PEG, tannin content consistently reduced in the seeds of high-tannin cultivar of sorghum [106,108], thus reducing the beneficial effect of seed priming. In order to preserve the benefits of seed priming, Patanè et al. [106] recommended the use of other osmotica (instead of PEG) or the adoption of biological agents for priming high-tannin sorghum cultivars.
2.3. Salinity Stress
Soil salinity is a serious problem limiting crop production. Salinity stress is especially problematic in arid and semi-arid regions of the world [13]. Salinity stress reduces soil water availability for plant roots by lowering osmotic potential [109]. The germination of most crops fails under severe or even moderate salinity stress [34,110,111]. Salinity negatively influences seed germination through ionic imbalances, osmotic stress, or mixed effects of these factors, causing detrimental impacts on enzymes, proteins, cell organelles, plasma membranes, thereby reducing respiration and photosynthetic rates [13,112]. In addition, salinity stress changes the signal balance in higher plants. Increasing salinity stress is linked with increases in abscisic acid and jasmonates as well as reductions in auxin, cytokinin, gibberellins [13,113]. Salinity stress can significantly delay seed germination and reduce the percentage of seeds that are capable of germinating [13]. Plants generally exhibit a decrease in biomass accumulation upon exposure to salinity stress [36].
The effectiveness of seed priming with inorganic salt, such as CaCl2, KCl, KNO3, and MgSO4, for overcoming salinity stress has been noted in a variety of crop species (Table 1) [4,114,115,116,117]. Shahi-Gharahlar et al. [117] reported that priming with PEG was less effective for decreasing salinity damage than with NaCl in summer squash (Cucurbita pepo L.) [117]. Nevertheless, extensive studies revealed that PEG priming is a successful approach to improve germination performance in various plant species including amaranth (Amaranthus spp.) [118], pepper (Capsicum annuum L.) [119], tomato (Solanum lycopersicum L.) [120], sugarcane (Saccharum officinarum L.) [121], soybean [122], wheat [123], and sweet corn [83] under salinity stress.
Salt tolerance conferred by seed priming is linked to an enhanced ability for osmotic adjustment as primed plants exhibited elevated concentrations of Ca2+, K+, and Mg2+ ions and reduced concentrations of Cl− or Na+ ions in roots and a greater amount of soluble sugars and organic acids in leaves compared to the unprimed plants [4,34,35]. Priming of wheat seeds with choline enhanced salinity tolerance by reducing the toxic elements of Na+ and Cl−, and by increasing the beneficial elements of Ca2+ and K+ [124]. Priming with melatonin enhanced indole-3-acetic acid, total phenolic content, as well as Ca2+ and K+ in the leaves of faba bean (Vicia faba L.) under salinity stress [125]. However, these physiological parameters have not been studied in PEG-primed plants under salinity stress, and therefore further research is needed to understand the specific mechanisms of PEG priming in protecting plants against salinity stress.
2.4. Multiple Abiotic Stresses
The concurrent occurrence of multiple abiotic stresses may result in additive or synergistic effects on plant growth. Consequently, combined occurrence of multiple abiotic stresses can be more damaging than when they occur sequentially or individually [4,68]. Under natural conditions, multiple abiotic stresses may commonly occur [55]. However, most PEG priming studies reported to date were conducted under the conditions of a single abiotic stress [5,33,64,75,80,83,126,127,128].
As shown in Table 1, a limited number of studies have investigated the effectiveness of PEG priming against multiple abiotic stresses and shown promising results [32,126,129,130,131,132]. Priming with PEG effectively enhanced seed germination and early seedling establishment in asparagus and spinach (Spinacia oleracea L.), especially when exposed to combined drought and low temperature stresses [32,129,130,132,133]. Seed priming with PEG also enhanced tolerance to simultaneously occurring drought and salinity stresses in sunflower [131]. However, the specific mechanisms of improved seed germination and emergence for PEG priming under multiple abiotic stresses, particularly at the cellular and molecular levels, have not yet been elucidated.
3. Conclusions
The effectiveness of PEG priming against different abiotic stresses has been shown in a wide range of crop species. PEG primed seeds generally resulted in earlier and synchronized seed germination largely due to enzyme activation, increased germination-promoting metabolites, and osmotic adjustment. The reported mechanisms of PEG priming against drought, temperature, or salinity stress are summarized in Figure 1. However, only a very few studies have examined the effects PEG priming against combined abiotic stresses. The specific mechanisms of PEG priming against abiotic stresses, especially under the conditions of multiple stresses, are not yet fully understood and our knowledge of the cellular and molecular levels changes following plant seeds after seed priming remains limited. Future research in this report is expected to facilitate broader adoption of this technology.
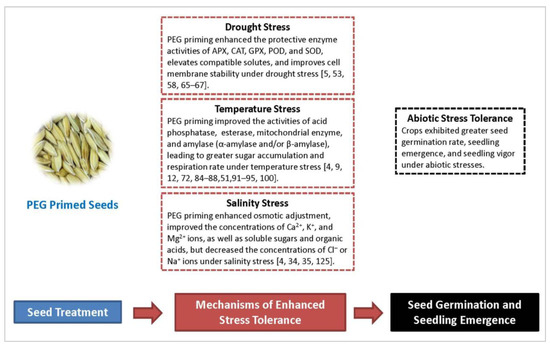
Figure 1.
The effects of PEG priming on germination and emergence of crop seeds under drought, temperature, and salinity stress conditions and associated mechanisms of stress tolerance. Abbreviations: APX, ascorbate peroxidase; CAT, catalase; GPX, glutathione peroxidase; PEG, polyethylene glycol; POD, peroxidase; SOD, superoxide dismutase.
Author Contributions
The idea on the structure and organization of this review paper came from C.L. and J.Y., C.L., M.B., S.M.S. and J.Y. wrote and/or revised the draft. H.W., W.M. and J.Y. collected the necessary references and prepared the table. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (No. 32072498).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
The authors thank Yiping Gao for his help in preparing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ahmed, M.; Qadeer, U.; Ahmed, Z.I.; Hassan, F. Improvement of wheat (Triticum aestivum) drought tolerance by seed priming with silicon. Arch. Agron. Soil Sci. 2016, 62, 299–315. [Google Scholar] [CrossRef]
- Ansari, O.; Azadi, M.; Sharif-Zadeh, F.; Younesi, E. Effect of hormone priming on germination characteristics and enzyme activity of mountain rye (Secale montanum) seeds under drought stress conditions. J. Stress Physiol. Biochem. 2013, 9, 61–71. [Google Scholar]
- Kazemi, K.; Eskandari, H. Does priming improve seed performance under salt and drought stress. J. Basic Appl. 2012, 2, 3503–3507. [Google Scholar]
- Savvides, A.; Ali, S.; Tester, M.; Fotopoulos, V. Chemical priming of plants against multiple abiotic stresses: Mission possible? Trends Plant. Sci. 2016, 21, 329–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Yu, J.; Johnston, C.R.; Wang, Y.; Zhu, K.; Lu, F.; Zhang, Z.; Zou, J. Seed priming with polyethylene glycol induces physiological changes in sorghum (Sorghum bicolor L. Moench) seedlings under suboptimal soil moisture environments. PLoS ONE 2015, 10, e0140620. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Tao, Y.; Hussain, S.; Jiang, Q.; Peng, S.; Huang, J.; Cui, K.; Nie, L. Seed priming in dry direct-seeded rice: Consequences for emergence, seedling growth and associated metabolic events under drought stress. Plant. Growth Regul. 2016, 78, 167–178. [Google Scholar] [CrossRef]
- Bai, Y.; Kissoudis, C.; Zhe, Y.; Visser, R.G.F.; Van Der Linden, G. Plant behaviour under combined stress. Plant. J. 2018, 93, 781–793. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Back, K. 2-Hydroxymelatonin promotes the resistance of rice plant to multiple simultaneous abiotic stresses (combined cold and drought). J. Pineal Res. 2016, 61, 303–316. [Google Scholar] [CrossRef]
- Zhang, H.; Dou, W.; Jiang, C.-X.; Wei, Z.-J.; Liu, J.; Jones, R.L. Hydrogen sulfide stimulates β-amylase activity during early stages of wheat grain germination. Plant. Signal. Behav. 2010, 5, 1031–1033. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Liu, W.; Zhang, S. Different responses of plant growth and antioxidant system to the combination of cadmium and heat stress in transgenic and non-transgenic rice. J. Integr. Plant. Biol. 2009, 51, 942–950. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R. Silicon (Si): Review and future prospects on the action mechanisms in alleviating biotic and abiotic stresses in plants. Ecotoxicol. Environ. Saf. 2017, 147, 881–896. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Song, Z.; Yan, Z.; Qian, H.; Song, A.; Liu, L.; Yang, X.; Xia, S.; Liang, Y. Silicon enhancement of estimated plant biomass carbon accumulation under abiotic and biotic stresses. a meta-analysis. Agron. Sustain. Dev. 2018, 38, 26. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Bioph. Res. Co. 2018, 495, 286–291. [Google Scholar] [CrossRef]
- Per, T.S.; Khan, M.I.R.; Anjum, N.A.; Masood, A.; Khan, N.A. Jasmonates in plants under abiotic stresses: Crosstalk with other phytohormones matters. Environ. Exp. Bot. 2018, 145, 104–120. [Google Scholar] [CrossRef]
- Abedi, T.; Pakniyat, H. Antioxidant enzymes changes in response to drought stress in ten cultivars of oilseed rape (Brassica napus L.). Czech., J. Gene. Plant. Breed. 2010, 46, 27–34. [Google Scholar] [CrossRef]
- Hossain, A.; Skalicky, M.; Brestic, M.; Maitra, S.; Ashraful Alam, M.; Syed, M.A.; Hossain, J.; Sarkar, S.; Saha, S.; Bhadra, P. Consequences and mitigation strategies of abiotic stresses in wheat (Triticum aestivum L.) under the changing climate. Agronomy 2021, 11, 241. [Google Scholar] [CrossRef]
- Marthandan, V.; Geetha, R.; Kumutha, K.; Renganathan, V.G.; Karthikeyan, A.; Ramalingam, J. Seed priming: A feasible strategy to enhance drought tolerance in crop plants. Inter. J. Mol. Sci. 2020, 21, 8258. [Google Scholar] [CrossRef] [PubMed]
- Abumhadi, N.M.; Atanassov, A.I. Future challenges of plant biotechnology and genomics. Rom. Biotechnol. Let. 2010, 15, 127–142. [Google Scholar]
- Bansal, K.C.; Lenka, S.K.; Mondal, T.K. Genomic resources for breeding crops with enhanced abiotic stress tolerance. Plant. Breed. 2014, 133, 1–11. [Google Scholar] [CrossRef]
- Boscaiu, M.; Donat, P.M.; Llinares, J.; Vicente, O. Stress-tolerant wild plants: A source of knowledge and biotechnological tools for the genetic improvement of stress tolerance in crop plants. Not. Bot. Horti. Agrobot. Cluj. Napoca. 2012, 40, 323–327. [Google Scholar] [CrossRef] [Green Version]
- Rosielle, A.; Hamblin, J. Theoretical aspects of selection for yield in stress and non-stress environment. Crop. Sci. 1981, 21, 943–946. [Google Scholar] [CrossRef]
- Vinocur, B.; Altman, A. Recent advances in engineering plant tolerance to abiotic stress: Achievements and limitations. Curr. Opin. Biotechnol. 2005, 16, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Jaganathan, D.; Ramasamy, K.; Sellamuthu, G.; Jayabalan, S.; Venkataraman, G. CRISPR for crop improvement: An update review. Front. Plant. Sci. 2018, 9, 985. [Google Scholar] [CrossRef]
- Lone, A.A.; Khan, M.H.; Dar, Z.A.; Wani, S.H. Breeding strategies for improving growth and yield under waterlogging conditions in maize: A review. Maydica 2018, 61, 11. [Google Scholar]
- Wang, Y.; Li, D.; Gao, J.; Li, X.; Chen, P. The 2’-O-methyladenosine nucleoside modification gene OsTRM13 positively regulates salt stress tolerance in rice. J. Exp. Bot. 2017, 68, 1479–1491. [Google Scholar] [CrossRef]
- Bhatnagar-Mathur, P.; Vadez, V.; Sharma, K.K. Transgenic approaches for abiotic stress tolerance in plants: Retrospect and prospects. Plant. Cell Rep. 2008, 27, 411–424. [Google Scholar] [CrossRef]
- Raina, M.; Pandotra, P.; Salgotra, R.; Ali, S.; Mir, Z.A.; Bhat, J.A.; Ali, A.; Tyagi, A.; Upadhahy, D. Genetic Engineering and Environmental Risk. In Modern Age Environmental Problems and Their Remediation; Oves, M., Zain Khan, M., Ismail, M.I.I., Eds.; Springer: Cham, Switzerland, 2018; pp. 69–82. [Google Scholar]
- Scott, S.E.; Inbar, Y.; Wirz, C.D.; Brossard, D.; Rozin, P. An overview of attitudes toward genetically engineered food. Ann. Rev. Nutr. 2018, 38, 459–479. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, A.; Mariani, L. Agronomic management for enhancing plant tolerance to abiotic stresses: High and low values of temperature, light intensity, and relative humidity. Horticulturae 2018, 4, 21. [Google Scholar] [CrossRef] [Green Version]
- Mariani, L.; Ferrante, A. Agronomic management for enhancing plant tolerance to abiotic stresses—Drought, salinity, hypoxia, and lodging. Horticulturae 2017, 3, 52. [Google Scholar] [CrossRef] [Green Version]
- Amini, R. Drought stress tolerance of barley (Hordeum vulgare L.) affected by priming with PEG. Inter. J. Farm. All. Sci. 2013, 2, 803–808. [Google Scholar]
- Bittencourt, M.; Dias, D.; Dias, L.; Araújo, E. Effects of priming on asparagus seed germination and vigour under water and temperature stress. Seed Sci. Technol. 2004, 32, 607–616. [Google Scholar] [CrossRef]
- Bush, E.W.; Wilson, P.; Shepard, D.P.; McClure, G. Enhancement of seed germination in common carpetgrass and centipedegrass seed. HortScience 2000, 35, 769–770. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J. Plant. Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Jisha, K.; Vijayakumari, K.; Puthur, J.T. Seed priming for abiotic stress tolerance: An overview. Acta Physiol. Plant. 2013, 35, 1381–1396. [Google Scholar] [CrossRef]
- Abdelhamid, M.T.; El-Masry, R.R.; Darwish, D.S.; Abdalla, M.M.; Oba, S.; Ragab, R.; Sabagh, A.E.; El Kholy, M.H.; Omer, E. Mechanisms of Seed Priming Involved in Salt Stress Amelioration. In Priming and Pretreatment of Seeds and Seedlings; Hasanuzzaman, M., Fotopoulos, V., Eds.; Springer: Singapore, 2019; pp. 219–251. [Google Scholar]
- Farooq, M.; Basra, S.; Wahid, A.; Cheema, Z.; Cheema, M.; Khaliq, A. Physiological role of exogenously applied glycinebetaine to improve drought tolerance in fine grain aromatic rice (Oryza sativa L.). J. Agron. Crop. Sci. 2008, 194, 325–333. [Google Scholar] [CrossRef]
- Farooq, M.; Irfan, M.; Aziz, T.; Ahmad, I.; Cheema, S. Seed priming with ascorbic acid improves drought resistance of wheat. J. Agron. Crop. Sci. 2013, 199, 12–22. [Google Scholar] [CrossRef]
- Kaya, M.D.; Okçu, G.; Atak, M.; Cıkılı, Y.; Kolsarıcı, Ö. Seed treatments to overcome salt and drought stress during germination in sunflower (Helianthus annuus L.). Europ. J. Agron. 2006, 24, 291–295. [Google Scholar] [CrossRef]
- Li, J.; Yin, L.; Jongsma, M.; Wang, C. Effects of light, hydropriming and abiotic stress on seed germination, and shoot and root growth of pyrethrum (Tanacetum cinerariifolium). Indus. Crop. Prod. 2011, 34, 1543–1549. [Google Scholar] [CrossRef]
- Varier, A.; Vari, A.K.; Dadlani, M. The subcellular basis of seed priming. Cur. Sci. 2010, 99, 450–456. [Google Scholar]
- Whitesides, G.M. Poly(ethylene glycol) chemistry biotechnical and biomedical applications. App. Biochem. Biotech. 1993, 41, 233–234. [Google Scholar] [CrossRef]
- Branch, D.W.; Wheeler, B.C.; Brewer, G.J.; Leckband, D.E. Long-term stability of grafted polyethylene glycol surfaces for use with microstamped substrates in neuronal cell culture. Biomaterials 2001, 22, 1035–1047. [Google Scholar] [CrossRef]
- Janes, B.E. The effect of molecular size, concentration in nutrient solution, and exposure time on the amount and distribution of polyethylene glycol in pepper plants. Plant. Physiol. 1974, 54, 226–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, B.E.; Kaufmann, M.R. The osmotic potential of polyethylene glycol 6000. Plant. Physiol. 1973, 51, 914–916. [Google Scholar] [CrossRef]
- Parera, C.A.; Cantliffe, D.J. Presowing seed priming. Hort. Rev. 1994, 16, 109–141. [Google Scholar]
- Farooq, S.; Hussain, M.; Jabran, K.; Hassan, W.; Rizwan, M.S.; Yasir, T.A. Osmopriming with CaCl2 improves wheat (Triticum aestivum L.) production under water-limited environments. Environ. Sci. Pol. Res. 2017, 24, 13638–13649. [Google Scholar] [CrossRef]
- Kubala, S.; Wojtyla, Ł.; Quinet, M.; Lechowska, K.; Lutts, S.; Garnczarska, M. Enhanced expression of the proline synthesis gene P5CSA in relation to seed osmopriming improvement of Brassica napus germination under salinity stress. J. Plant. Physiol. 2015, 183, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouradi, M.; Bouizgaren, A.; Farissi, M.; Latrach, L.; Qaddoury, A.; Ghoulam, C. Seed osmopriming improves plant growth, nodulation, chlorophyll fluorescence and nutrient uptake in alfalfa (Medicago sativa L.)—Rhizobia symbiosis under drought stress. Sci. Hortic. 2016, 213, 232–242. [Google Scholar] [CrossRef]
- Tabassum, T.; Ahmad, R.; Farooq, M.; Basra, S.M.A. Improving salt tolerance in barley by osmopriming and biopriming. Int. J. Agric. Biol. 2018, 20, 2455–2464. [Google Scholar]
- Paparella, S.; Araújo, S.; Rossi, G.; Wijayasinghe, M.; Carbonera, D.; Balestrazzi, A. Seed priming: State of the art and new perspectives. Plant. Cell Rep. 2015, 34, 1281–1293. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant. Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trend. Plant. Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trend. Plant. Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Oladosu, Y.; Rafii, M.Y.; Samuel, C.; Fatai, A.; Magaji, U.; Kareem, I.; Kamarudin, Z.S.; Muhammad, I.i.; Kolapo, K. Drought resistance in rice from conventional to molecular breeding: A review. Inter. J. Mol. Sci. 2019, 20, 3519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, D.; Joshi, A.; Khan, P.; Gothkar, P.; Sodhi, P. On-farm seed priming in semi-arid agriculture: Development and evaluation in maize, rice and chickpea in India using participatory methods. Experi. Agric. 1999, 35, 15–29. [Google Scholar] [CrossRef]
- Goswami, A.; Banerjee, R.; Raha, S. Drought resistance in rice seedlings conferred by seed priming. Protoplasma 2013, 250, 1115–1129. [Google Scholar] [CrossRef] [PubMed]
- Jisha, K.C.; Puthur, J.T. Seed halopriming outdo hydropriming in enhancing seedling vigor and osmotic stress tolerance potential of rice varieties. J. Crop. Sci. Biotechnol. 2014, 17, 209–219. [Google Scholar] [CrossRef]
- Azadi, M.; Younesi, E.; Tabatabaei, S. Seed germination, seedling growth and enzyme activity of wheat seed primed under drought and different temperature conditions. J. Stress Physiol. Biochem. 2013, 9, 310–318. [Google Scholar]
- Bradford, K.J.; Somasco, O.A. Water relations of lettuce seed thermoinhibition. I. Priming and endosperm effects on base water potential. Seed Sci. Res. 1994, 4, 1–10. [Google Scholar] [CrossRef]
- Corbineau, F.; Picard, M.; Côme, D. Germinability of leek seeds and its improvement by osmopriming. Acta Hortic. 1993, 371, 45–52. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Suresh Kumar, J.; Suprasanna, P. Seed ‘primeomics’: Plants memorize their germination under stress. Biol. Rev. 2021, 96, 1723–1743. [Google Scholar] [CrossRef] [PubMed]
- Tiryaki, I.; Kizilsimsek, M.; Kaplan, M. Rapid and enhanced germination at low temperature of alfalfa and white clover seeds following osmotic priming. Trop. Grassland. 2009, 43, 171–177. [Google Scholar]
- Ahmad, P.; Jaleel, C.A.; Azooz, M.; Nabi, G. Generation of ROS and non-enzymatic antioxidants during abiotic stress in plants. Bot. Res. Inter. 2009, 2, 11–20. [Google Scholar]
- Ahmad, P.; Sarwat, M.; Sharma, S. Reactive oxygen species, antioxidants and signaling in plants. J. Plant. Biol. 2008, 51, 167–173. [Google Scholar] [CrossRef]
- Zhang, C.; He, P.; Yu, Z.; Hu, S. Effect of zinc sulphate and PEG priming on ageing seed germination and antioxidase activities of Perilla frutescens seedlings. China J. Chin. Mater. Med. 2010, 35, 2372–2377. [Google Scholar]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S. Crop production under drought and heat stress: Plant responses and management options. Front. Plant. Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guy, C.; Kaplan, F.; Kopka, J.; Selbig, J.; Hincha, D.K. Metabolomics of temperature stress. Physiol. Plant. 2008, 132, 220–235. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urano, K.; Kurihara, Y.; Seki, M.; Shinozaki, K. ‘Omics’ analyses of regulatory networks in plant abiotic stress responses. Curr. Opin. Plant. Biol. 2010, 13, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Zinn, K.E.; Tunc-Ozdemir, M.; Harper, J.F. Temperature stress and plant sexual reproduction: Uncovering the weakest links. J. Exp. Bot. 2010, 61, 1959–1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahan, J.; McMichael, B.; Wanjura, D. Methods for reducing the adverse effects of temperature stress on plants: A review. Environ. Exp. Bot. 1995, 35, 251–258. [Google Scholar] [CrossRef]
- Ruelland, E.; Zachowski, A. How plants sense temperature. Environ. Exp. Bot. 2010, 69, 225–232. [Google Scholar] [CrossRef]
- Lee, S.; Kim, J. Total sugars, α-amylase activity, and germination after priming of normal and aged rice seeds. Korean J. Crop. Sci. 2000, 45, 108–111. [Google Scholar]
- Murray, G.; Swensen, J.B.; Gallian, J.J. Emergence of sugar beet seedlings at low soil temperature following seed soaking and priming. HortScience 1993, 28, 31–32. [Google Scholar] [CrossRef] [Green Version]
- Murray, G.A.; Swensen, J.B.; Beaver, G. Emergence of spring-and summer-planted onions following osmotic priming. HortScience 1992, 27, 409–410. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, W.; Huber, D.; Cantliffe, D. Carrot seed germination and respiration at high temperature in response to seed maturity and priming. Seed Sci. Technol. 2013, 41, 164–169. [Google Scholar] [CrossRef]
- Nascimento, W.M.; Huber, D.J.; Cantliffe, D.J. Carrot seed germination and ethylene production at high temperature in response to seed osmopriming. Hortic. Bras. 2013, 31, 554–558. [Google Scholar] [CrossRef]
- Parera, C.A.; Cantliffe, D.J. Priming leek seed for improved germination and emergence at high temperature. HortScience 1992, 27, 1077–1079. [Google Scholar] [CrossRef] [Green Version]
- Bodsworth, S.; Bewley, J. Osmotic priming of seeds of crop species with polyethylene glycol as a means of enhancing early and synchronous germination at cool temperatures. Can. J. Bot. 1981, 59, 672–676. [Google Scholar] [CrossRef]
- Yu, F.; Liu, Y. Effects of PEG pretreatment on seed vigour of Masson pine. J. Nanjing For. Uni. 2000, 24, 38–40. [Google Scholar]
- Patanè, C.; Cavallaro, V.; Cosentino, S.L. Germination and radicle growth in unprimed and primed seeds of sweet sorghum as affected by reduced water potential in NaCl at different temperatures. Indust. Crop. Prod. 2009, 30, 1–8. [Google Scholar] [CrossRef]
- Fu, K.; Lu, D. Reaction kinetics study of α-amylase in the hydrolysis of starch size on cotton fabrics. J. Text. Inst. 2014, 105, 203–208. [Google Scholar] [CrossRef]
- Goswami, A.; Jain, M.; Paul, B. α-and β-Amylases in seed germination. Biolog. Plant. 1977, 19, 469–471. [Google Scholar] [CrossRef]
- Kirschenbaum, D.M. The action of α-amylase on starch. Biochem. Educ. 1983, 11, 152–153. [Google Scholar] [CrossRef]
- Martínez, J.L.; Meza, E.; Petranovic, D.; Nielsen, J. The impact of respiration and oxidative stress response on recombinant α-amylase production by Saccharomyces cerevisiae. Met. Eng. Com. 2016, 3, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Henson, C.A. A quantitative assessment of the importance of barley seed α-amylase, β-amylase, debranching enzyme, and α-glucosidase in starch degradation. Arch. Biochem. Biophys. 1991, 284, 298–305. [Google Scholar] [CrossRef]
- Lee, S.; Kim, J.; Hong, S.; Yun, S.; Park, E. Priming effect of rice seeds on seedling establishment under adverse soil conditions. Korean J. Crop. Sci. 1998, 43, 194–198. [Google Scholar]
- Lee, W.S.; Slaughter, D.; Giles, D. Robotic weed control system for tomatoes. Precis. Agric. 1999, 1, 95–113. [Google Scholar] [CrossRef]
- Chiu, K.; Chen, C.; Sung, J. Effect of priming temperature on storability of primed sh-2 sweet corn seed. Crop. Sci. 2002, 42, 1996–2003. [Google Scholar] [CrossRef]
- Debbarma, A.; Devi, J.; Barua, M. Seed priming durations and concentrations influence on germination and seedling growth of bitter gourd. Veg. Sci. 2018, 45, 137–139. [Google Scholar]
- Girolamo, D.G.; Barbanti, L. Treatment conditions and biochemical processes influencing seed priming effectiveness. Ita. J. Agron. 2012, 7, e25. [Google Scholar] [CrossRef] [Green Version]
- Stephen, K.; Khan, F.; Bhat, S.; Narayan, S.; Mir, S.; Mir, M.; Hussain, K.; Gul, M.; Khurshid, A.; Siddiqi, I. Optimizing priming concentration and duration of various priming agents for improved seed germination in chilli (Capsicum annum L.). J. Pharm. Phytochem. 2018, 7, 2689–2693. [Google Scholar]
- Yang, Y.; Chen, W.; Guo, J. Effects of PVA and PEG pretreatment on development and ultrastructure of plumular root mitochondria in soybean seed during low temperature imbibition process. J. Integr. Plant. Biol. 1992, 34, 432–436. [Google Scholar]
- Morohashi, Y. Patterns of mitochondrial development in reserve tissues of germinated seeds: A survey. Physiol. Plant. 1986, 66, 653–658. [Google Scholar] [CrossRef]
- Noctor, G.; De Paepe, R.; Foyer, C.H. Mitochondrial redox biology and homeostasis in plants. Trends Plant. Sci. 2007, 12, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Smiri, M.; Chaoui, A.; El Ferjani, E. Respiratory metabolism in the embryonic axis of germinating pea seed exposed to cadmium. J. Plant. Physiol. 2009, 166, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhang, Z.; Liu, J.; Chen, M.; Pan, R.; Hu, W.; Guan, Y.; Hu, J. Seed priming with spermidine and trehalose enhances chilling tolerance of rice via different mechanisms. J. Plant. Growth Regul. 2020, 39, 669–679. [Google Scholar] [CrossRef]
- Khan, A.A.; Tao, K.-L.; Knypl, J.; Borkowska, B.; Powell, L.E. Osmotic conditioning of seeds: Physiological and biochemical changes. Acta Hortic. 1977, 83, 267–278. [Google Scholar] [CrossRef]
- Bailly, C.; Benamar, A.; Corbineau, F.; Côme, D. Antioxidant systems in sunflower (Helianthus annuus L.) seeds as affected by priming. Seed Sci. Res. 2000, 10, 35–42. [Google Scholar] [CrossRef]
- Guan, Y.; Hu, J.; Wang, X.; Shao, C. Seed priming with chitosan improves maize germination and seedling growth in relation to physiological changes under low temperature stress. J. Zhejiang Univ. Sci. 2009, 10, 427–433. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Hussain, S.; Khaliq, A.; Ali, S.; Khan, I. Physiological, Biochemical, and Molecular Aspects of Seed Priming. In Priming and Pretreatment of Seeds and Seedlings; Hasanuzzaman, M., Fotopoulos, V., Eds.; Springer: Singapore, 2019; pp. 43–62. [Google Scholar]
- Kaur, H.; Goyal, M. Salicylic acid priming enhances low temperature stress tolerance in Egyptian clover (Trifolium alexandrinum L.) by influencing antioxidant system. Indian J. Exp. Biol. 2019, 24, 291–295. [Google Scholar]
- Shah, T.; Latif, S.; Khan, H.; Munsif, F.; Nie, L. Ascorbic acid priming enhances seed germination and seedling growth of winter wheat under low temperature due to late sowing in Pakistan. Agronomy 2019, 9, 757. [Google Scholar] [CrossRef] [Green Version]
- Patanè, C.; Cavallaro, V.; D’Agosta, G.; Cosentino, S. Plant emergence of PEG-osmoprimed seeds under suboptimal temperatures in two cultivars of sweet sorghum differing in seed tannin content. J. Agron. Crop. Sci. 2008, 194, 304–309. [Google Scholar] [CrossRef]
- Harris, H.B.; Burns, R.E. Influence of tannin content on preharvest seed germination in sorghum. Agron. J. 1970, 62, 835–836. [Google Scholar] [CrossRef]
- Kantar, F.; Pilbeam, C.; Hebblethwaite, P. Effect of tannin content of faba bean (Vicia faba) seed on seed vigour, germination and field emergence. Ann. App. Biol. 1996, 128, 85–93. [Google Scholar] [CrossRef]
- Sheldon, A.R.; Dalal, R.C.; Kirchhof, G.; Kopittke, P.M.; Menzies, N.W. The effect of salinity on plant-available water. Plant. Soil 2017, 418, 477–491. [Google Scholar] [CrossRef]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pol. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef] [PubMed]
- Volkmar, K.; Hu, Y.; Steppuhn, H. Physiological responses of plants to salinity: A review. Can. J. Plant. Sci. 1998, 78, 19–27. [Google Scholar] [CrossRef]
- Porcel, R.; Aroca, R.; Ruiz-Lozano, J.M. Salinity stress alleviation using arbuscular mycorrhizal fungi. A review. Agron. Sust. Develop. 2012, 32, 181–200. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Goyal, M.R.; Singh, A. Physiological and biochemical changes in plants under soil salinity stress: A review. In Engineering Practices for Management of Soil Salinity; Apple Academic Press: New York, NY, USA, 2018; pp. 159–200. [Google Scholar]
- Biswas, S.; Rasal-Monir, M.; Islam, M.; Modak, S.; Kabir, M.H. Induction of salt tolerance in tomato through seed priming. Plant. 2019, 7, 47. [Google Scholar] [CrossRef]
- Chatterjee, P. Sodium chloride primed seeds modulate glutathione metabolism in legume cultivars under NaCl stress. Amer. J. Plant. Physiol. 2018, 13, 8–22. [Google Scholar] [CrossRef] [Green Version]
- Senturk, B.; Sivritepe, H.O. NaCl priming alleviates the inhibiting effect of salinity during seeding growth of peas (Pisum sativum L.). Fresen. Environ. Bull. 2016, 11, 4202. [Google Scholar]
- Shahi-Gharahlar, A.; Farhoudi, R.; Mosavi, M. Effect of seed pretreatment on summer squash (Cucurbita pepo) seed germination and seedling characteristics under salinity condition. Seed Sci. Biotechnol. 2009, 3, 15–23. [Google Scholar]
- Moosavi, A.; Tavakkol Afshari, R.; Sharif-Zadeh, F.; Aynehband, A. Seed priming to increase salt and drought stress tolerance during germination in cultivated species of Amaranth. Seed Sci.Technol. 2009, 37, 781–785. [Google Scholar] [CrossRef]
- Amjad, M.; Ziaf, K.; Iqbal, Q.; Ahmad, I.; Riaz, M.; Saqib, Z.A. Effect of seed priming on seed vigour and salt tolerance in hot pepper. Pak. J. Agri. Sci. 2007, 44, 408–416. [Google Scholar]
- Pradhan, N.; Prakash, P.; Manimurugan, C.; Tiwari, S.K.; Sharma, R.; Singh, P. Screening of tomato genotypes using osmopriming with PEG 6000 under salinity stress. Res. Environ. Life Sci. 2015, 8, 245–250. [Google Scholar]
- Patade, V.Y.; Bhargava, S.; Suprasanna, P. Better osmotic adjustment mediates salt and PEG stress tolerance in primed plants of contrasting cultivars of sugarcane. Sugar Tech. 2015, 17, 348–355. [Google Scholar] [CrossRef]
- Bejandi, T.K.; Sedghi, M.; Sharifi, R.S.; Namvar, A.; Molaei, P. Seed priming and sulfur effects on soybean cell membrane stability and yield in saline soil. Pes. Agro. Bra. 2009, 44, 1114–1117. [Google Scholar] [CrossRef] [Green Version]
- Ghiyasi, M.; Myandoab, M.P.; Tajbakhsh, M.; Salehzadeh, H.; Meshkat, M. Influence of different osmopriming treatments on emergency and yield of maize (Zea mays L.). Res. J. Biological. Sci. 2008, 3, 1452–1455. [Google Scholar]
- Salama, K.H.; Mansour, M.M.; Hassan, N.S. Choline priming improves salt tolerance in wheat (Triticum aestivum L.). Aust. J. Basic. Appl. Sci. 2011, 5, 126–132. [Google Scholar]
- Dawood, M.G.; El-Awadi, M.E. Alleviation of salinity stress on Vicia faba L. plants via seed priming with melatonin. Acta Biológica Colom. 2015, 20, 223–235. [Google Scholar] [CrossRef]
- Farooq, M.; Tabassum, R.; Afzal, I. Enhancing the performance of direct seeded fine rice by seed priming. Plant. Prod. Sci. 2006, 9, 446–456. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Jia, G.; Yu, X. Water uptake and WUE of apple tree-corn agroforestry in the loess hilly region of China. Agric. Wat. Manag. 2020, 234, 106138. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, H.; Yu, X.; Jia, G.; Jiang, J. Evidence of foliar water uptake in a conifer species. Agric. Wat. Manag. 2021, 255, 106993. [Google Scholar] [CrossRef]
- Chen, K.; Arora, R.; Arora, U. Osmopriming of spinach (Spinacia oleracea L. cv. Bloomsdale) seeds and germination performance under temperature and water stress. Seed Sci. Technol. 2010, 38, 36–48. [Google Scholar] [CrossRef]
- Frett, J.; Pill, W. Germination characteristics of osmotically primed and stored impatiens seeds. Sci. Hortic. 1989, 40, 171–179. [Google Scholar] [CrossRef]
- Moghanibashi, M.; Karimmojeni, H.; Nikneshan, P. Seed treatment to overcome drought and salt stress during germination of sunflower (Helianthus annuus L.). J. Agrobiol. 2013, 30, 89–96. [Google Scholar]
- Rahimi, A. Seed priming improves the germination performance of cumin (Cuminum syminum L.) under temperature and water stress. Ind. Crop. Prod. 2013, 42, 454–460. [Google Scholar] [CrossRef]
- Liu, Z.; Jia, G.; Yu, X. Variation of water uptake in degradation agroforestry shelterbelts on the North China Plain. Agric. Ecosys. Environ. 2020, 287, 106697. [Google Scholar] [CrossRef]
- Carpenter, W.J. Priming dusty miller seeds: Role of aeration, temperature, and relative humidity. HortScience 1990, 25, 299–302. [Google Scholar] [CrossRef] [Green Version]
- El-Saidy, A.E.; Farouk, S.; El-Ghany, H.A. Evaluating of different seed priming on seedling growth, yield and quality componenets in two sunflower (Helianthus annuus L.) cultivars. Trends App. Sci. Res. 2011, 6, 977–991. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).