
Single- versus Double-Species Cover Crop Effects on Soil Health and Yield in Mississippi Soybean Fields


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design, Site History, and Management
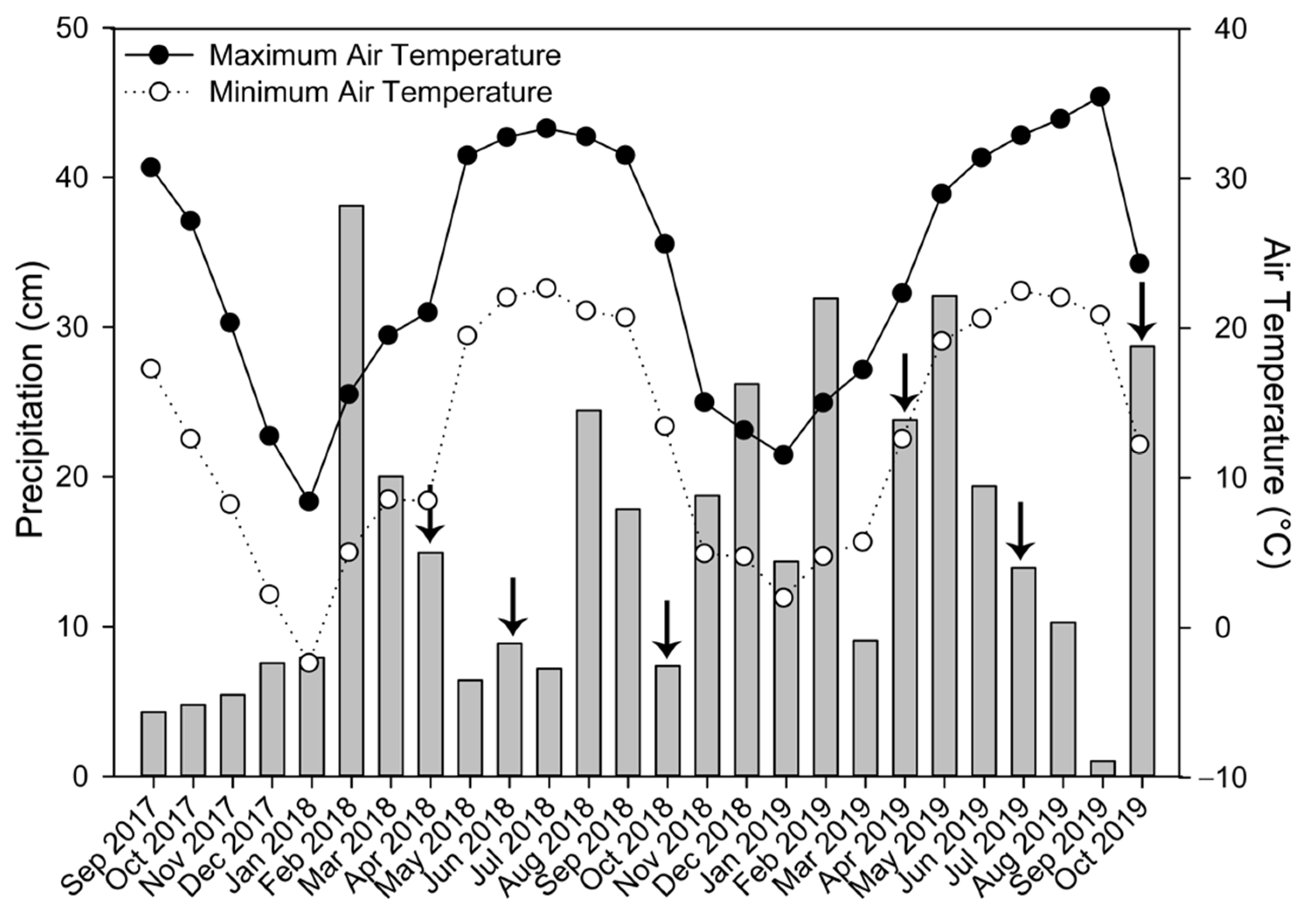
2.2. Sample Collection
2.3. Soil Sample Analyses
2.4. Statistics
3. Results and Discussion
3.1. Cover Crop Biomass
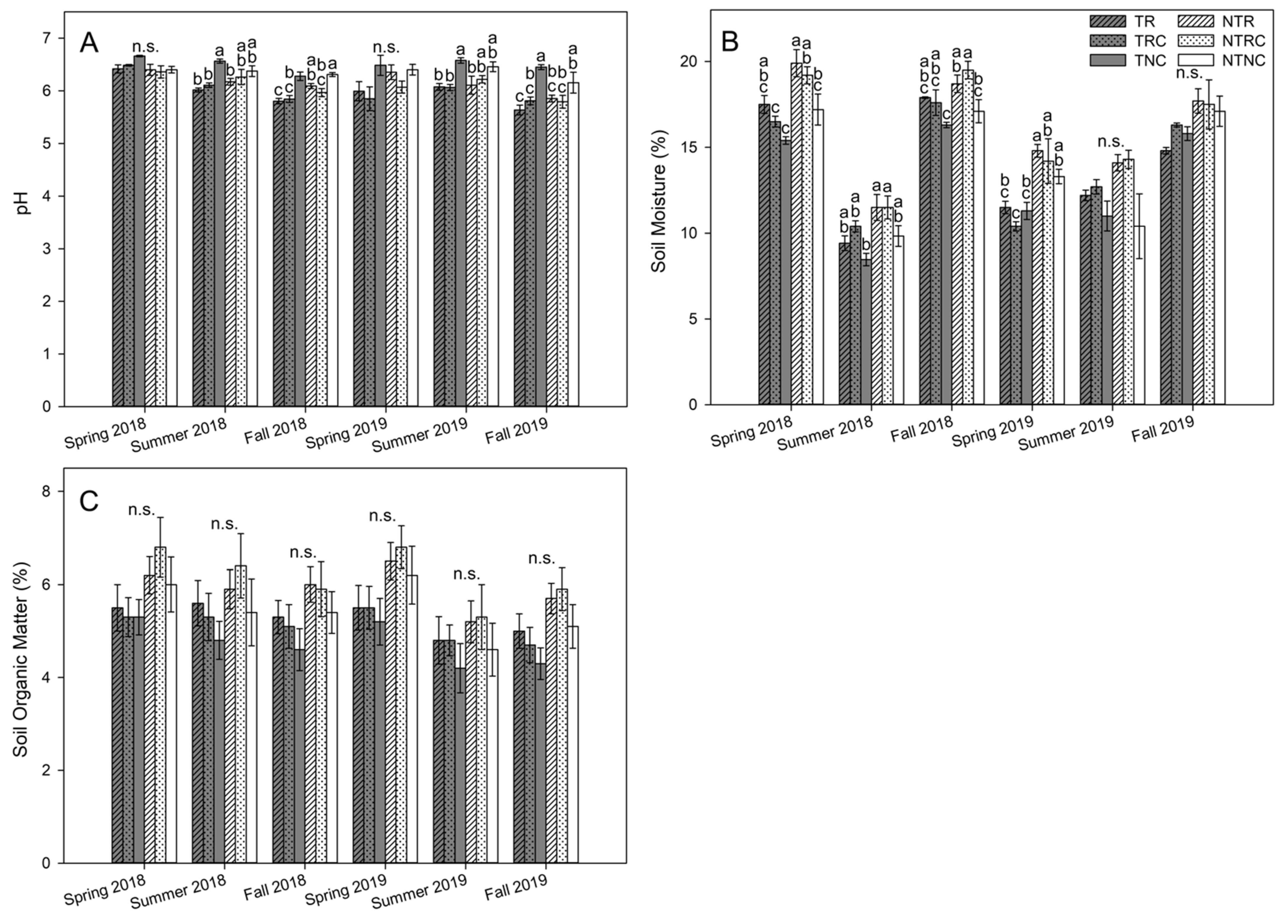
3.2. Soil Chemical Characteristics
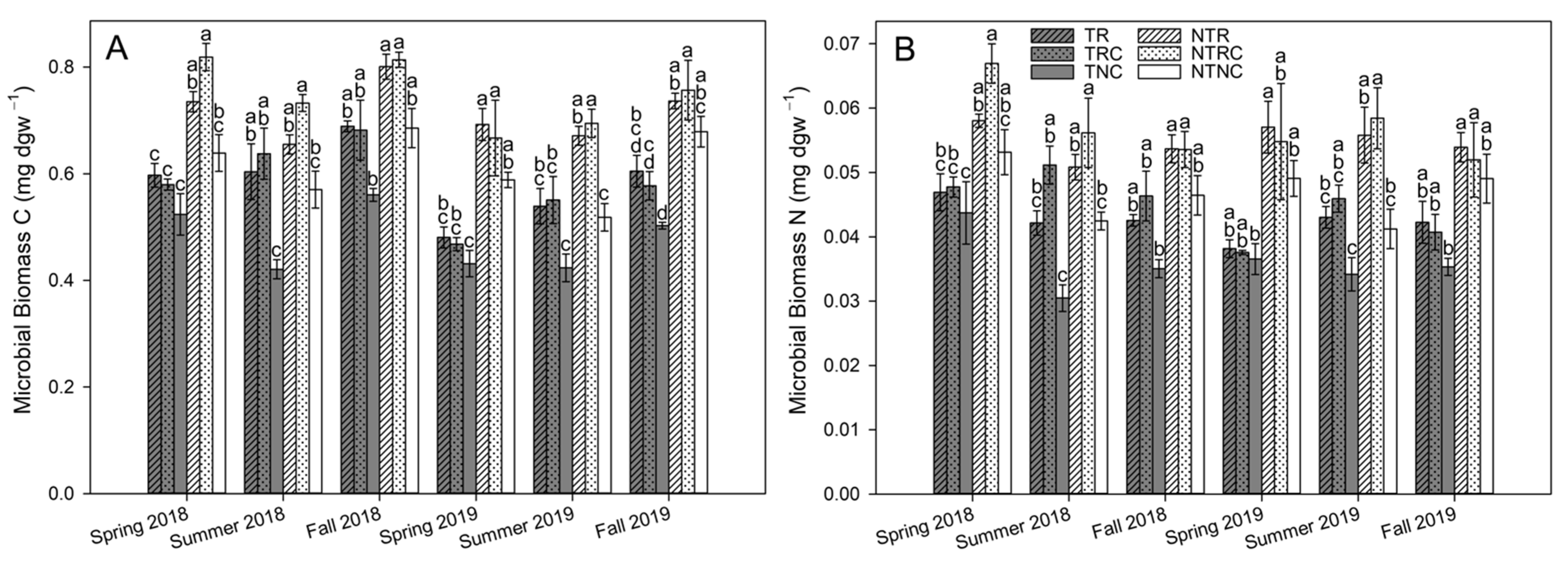
3.3. Microbial Biomass
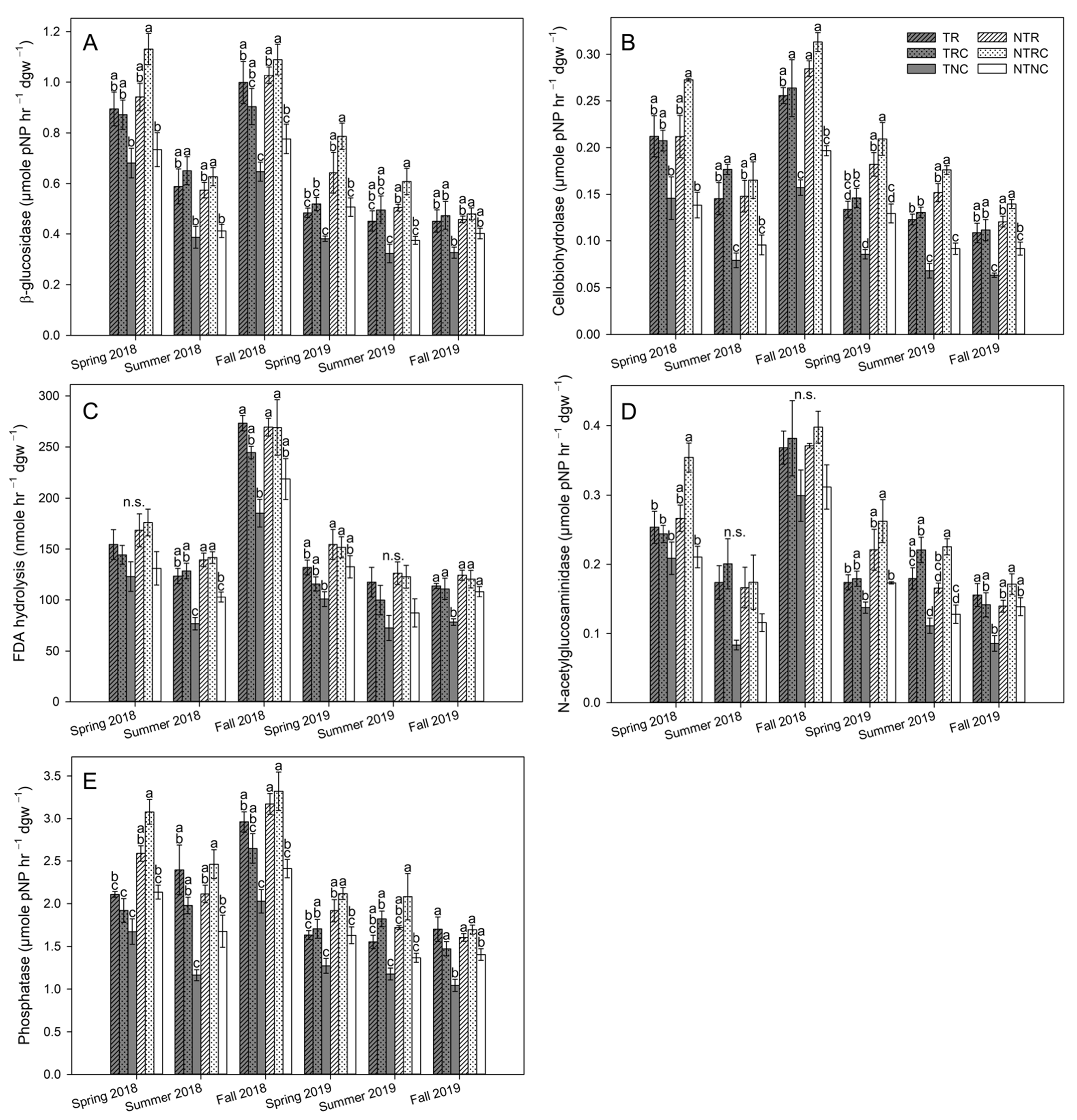
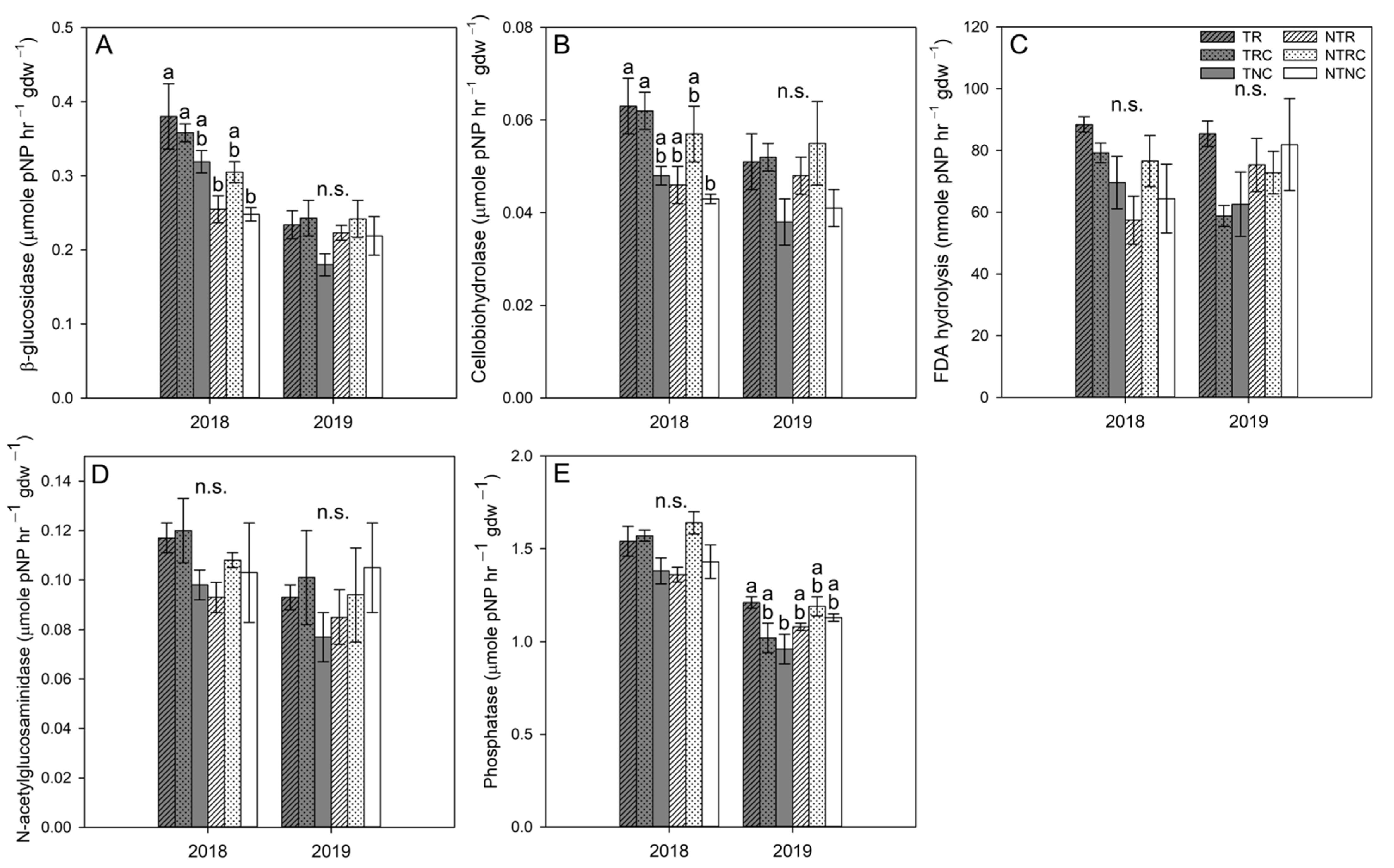
3.4. Soil Enzyme Activities
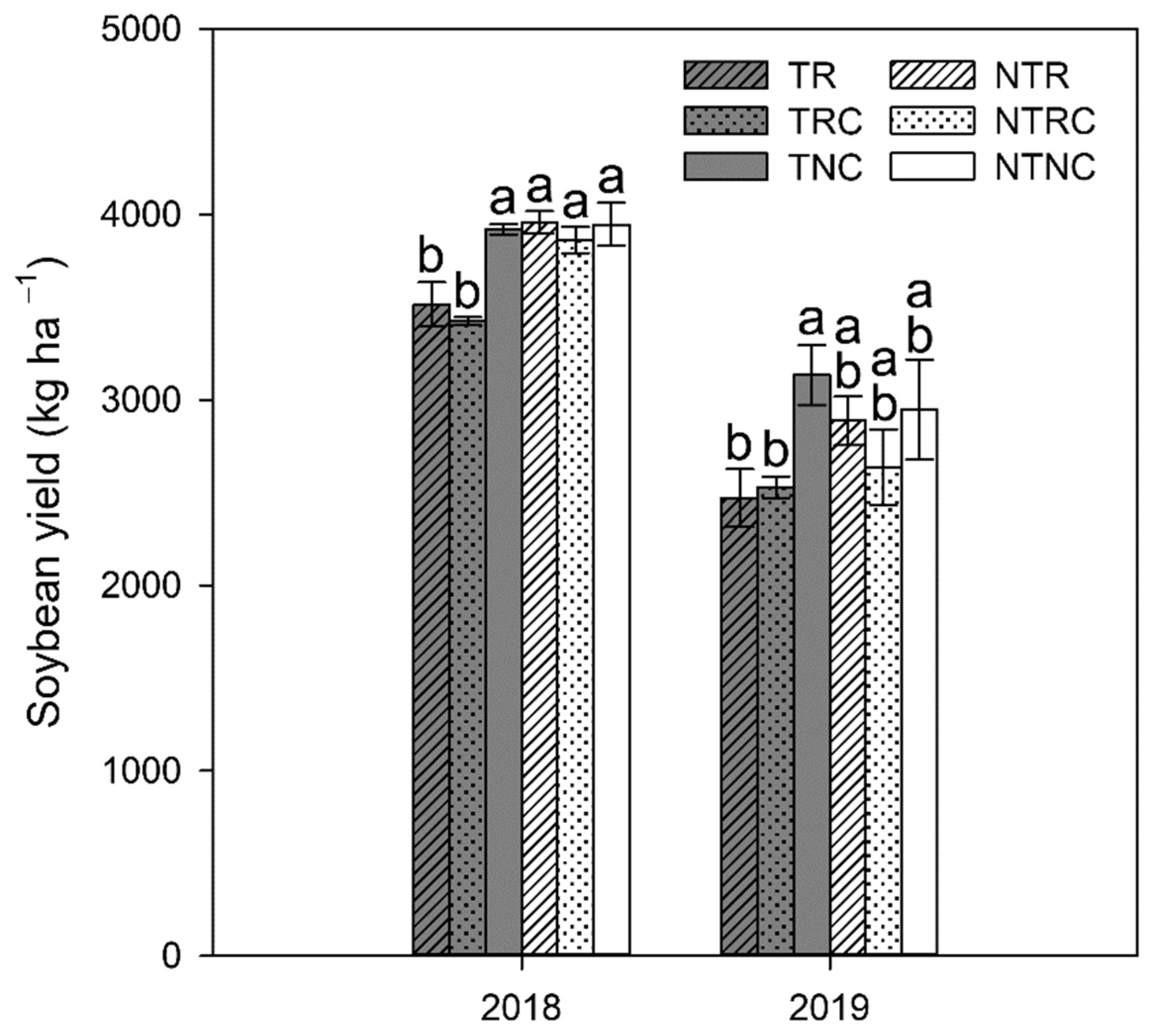
3.5. Effects on Soybean Yield
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Locke, M.A.; Bryson, C.T. Herbicide-soil interactions in reduced tillage and plant residue management systems. Weed Sci. 1997, 45, 307–320. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Shaver, T.M.; Lindquist, J.L.; Shapiro, C.A.; Elmore, R.W.; Francis, C.A.; Hergert, G.W. Cover Crops and Ecosystem Services: Insights from Studies in Temperate Soils. Agron. J. 2015, 107, 2449–2474. [Google Scholar] [CrossRef] [Green Version]
- Teasdale, J.R.; Brandsæter, L.O.; Calegari, A.; Neto, F.S. Cover crops and weed management. In Non-Chemical Weed Management: Principles, Concepts and Technology; CABI: Egham, UK, 2007; pp. 49–64. [Google Scholar]
- Bergtold, J.S.; Ramsey, S.; Maddy, L.; Williams, J.R. A review of economic considerations for cover crops as a conservation practice. Renew. Agric. Food Syst. 2019, 34, 62–76. [Google Scholar] [CrossRef]
- Mischler, R.A.; Curran, W.S.; Duiker, S.W.; Hyde, J.A. Use of a Rolled-rye Cover Crop for Weed Suppression in No-Till Soybeans. Weed Technol. 2010, 24, 253–261. [Google Scholar] [CrossRef]
- Reddy, K.N. Effects of Cereal and Legume Cover Crop Residues on Weeds, Yield, and Net Return in Soybean (Glycine max). Weed Technol. 2001, 15, 660–668. [Google Scholar] [CrossRef]
- Westgate, L.R.; Singer, J.W.; Kohler, K.A. Method and Timing of Rye Control Affects Soybean Development and Resource Utilization. Agron. J. 2005, 97, 806–816. [Google Scholar] [CrossRef]
- Sainju, U.M.; Singh, B.P. Tillage, Cover Crop, and Kill-Planting Date Effects on Corn Yield and Soil Nitrogen. Agron. J. 2001, 93, 878–886. [Google Scholar] [CrossRef]
- Acharya, J.; Bakker, M.G.; Moorman, T.B.; Kaspar, T.C.; Lenssen, A.W.; Robertson, A.E. Time Interval Between Cover Crop Termination and Planting Influences Corn Seedling Disease, Plant Growth, and Yield. Plant Dis. 2017, 101, 591–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirsky, S.B.; Curran, W.S.; Mortenseny, D.M.; Ryany, M.R.; Shumway, D.L. Timing of Cover-Crop Management Effects on Weed Suppression in No-Till Planted Soybean using a Roller-Crimper. Weed Sci. 2011, 59, 380–389. [Google Scholar] [CrossRef]
- Cookson, W.; Beare, M.; Wilson, P. Effects of prior crop residue management on microbial properties and crop residue decomposition. Appl. Soil Ecol. 1998, 7, 179–188. [Google Scholar] [CrossRef]
- Jahanzad, E.; Barker, A.V.; Hashemi, M.; Eaton, T.; Sadeghpour, A.; Weis, S.A. Nitrogen Release Dynamics and Decomposition of Buried and Surface Cover Crop Residues. Agron. J. 2016, 108, 1735–1741. [Google Scholar] [CrossRef]
- Kessavalou, A.; Walters, D.T. Winter Rye as a Cover Crop Following Soybean under Conservation Tillage. Agron. J. 1997, 89, 68–74. [Google Scholar] [CrossRef]
- Kessavalou, A.; Walters, D.T. Winter Rye Cover Crop Following Soybean Under Conservation Tillage: Residual Soil Nitrate. Agron. J. 1999, 91, 643–649. [Google Scholar] [CrossRef]
- Shekoofa, A.; Safikhan, S.; Raper, T.B.; Butler, S.A. Allelopathic Impacts of Cover Crop Species and Termination Timing on Cotton Germination and Seedling Growth. Agronomy 2020, 10, 638. [Google Scholar] [CrossRef]
- Barnes, J.P.; Putnam, A.R. Rye residues contribute weed suppression in no-tillage cropping systems. J. Chem. Ecol. 1983, 9, 1045–1057. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, A.L.; Sheaffer, C.C.; Wyse, D.L.; Staley, C.; Gould, T.J.; Sadowsky, M.J. Associations between soil bacterial community structure and nutrient cycling functions in long-term organic farm soils following cover crop and organic fertilizer amendment. Sci. Total Environ. 2016, 566–567, 949–959. [Google Scholar] [CrossRef] [Green Version]
- Locke, M.A.; Zablotowicz, R.M.; Steinriede, R.W.; Testa, S.; Reddy, K.N. Conservation Management in Cotton Production: Long-Term Soil Biological, Chemical, and Physical Changes. Soil Sci. Soc. Am. J. 2013, 77, 974–984. [Google Scholar] [CrossRef]
- Tyler, H.L. Winter cover crops and no till management enhance enzyme activities in soybean field soils. Pedobiologia 2020, 81-82, 150666. [Google Scholar] [CrossRef]
- Salvagiotti, F.; Cassman, K.; Specht, J.; Walters, D.; Weiss, A.; Dobermann, A. Nitrogen uptake, fixation and response to fertilizer N in soybeans: A review. Field Crop. Res. 2008, 108, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, P.G.; Parrish, D.J.; Luna, J.M. Cover crop contributions to N supply and water conservation in corn production. Am. J. Altern. Agric. 1991, 6, 106–113. [Google Scholar] [CrossRef]
- Pagano, M.C.; Miransari, M. The importance of soybean production worldwide. In Abiotic and Biotic Stresses in Soybean Production; Miransari, M., Ed.; Academic Press: San Diego, CA, USA, 2016; pp. 1–26. [Google Scholar]
- Horwath, W.R.; Paul, E.A. Microbial Biomass. In Methods of Soil Analysis: Part 2—Microbiological and Biochemical Properties; Bottomley, P.S., Angle, J.S., Weaver, R.W., Eds.; Soil Science Society of America Book Series: Madison, WI, USA, 1994; pp. 753–773. [Google Scholar]
- Jackson, C.R.; Tyler, H.L.; Millar, J. Determination of Microbial Extracellular Enzyme Activity in Waters, Soils, and Sediments using High Throughput Microplate Assays. J. Vis. Exp. 2013, 80, e50399. [Google Scholar] [CrossRef] [Green Version]
- Schnürer, J.; Rosswall, T. Fluorescein Diacetate Hydrolysis as a Measure of Total Microbial Activity in Soil and Litter. Appl. Environ. Microbiol. 1982, 43, 1256–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, E.S. Barley uptake of N deposited in the rhizosphere of associated field pea. Soil Biol. Biochem. 1996, 28, 159–168. [Google Scholar] [CrossRef]
- Keating, B.A.; Carberry, P.S. Resource capture and use in intercropping: Solar radiation. Field Crop. Res. 1993, 34, 273–301. [Google Scholar] [CrossRef]
- Reddy, K.N.; Zablotowicz, R.M.; Locke, M.A.; Koger, C.H. Cover crop, tillage, and herbicide effects on weeds, soil properties, microbial populations, and soybean yield. Weed Sci. 2003, 51, 987–994. [Google Scholar] [CrossRef]
- Moore, E.B.; Wiedenhoeft, M.H.; Kaspar, T.C.; Cambardella, C.A. Rye Cover Crop Effects on Soil Quality in No-Till Corn Silage-Soybean Cropping Systems. Soil Sci. Soc. Am. J. 2014, 78, 968–976. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Lal, R. No-Tillage and Soil-Profile Carbon Sequestration: An On-Farm Assessment. Soil Sci. Soc. Am. J. 2008, 72, 693–701. [Google Scholar] [CrossRef] [Green Version]
- Zablotowicz, R.M.; Reddy, K.N.; Weaver, M.A.; Mengistu, A.; Krutz, L.J.; Gordon, R.E.; Bellaloui, N. Cover crops, tillage, and glyphosate effects on chemical and biological properties of a lower Mississippi Delta soil and soybean yield. Environ. Res. J. 2010, 4, 227–251. [Google Scholar]
- Lupwayi, N.Z.; Larney, F.J.; Blackshaw, R.E.; Kanashiro, D.A.; Pearson, D.C. Phospholipid fatty acid biomarkers show positive soil microbial community responses to conservation soil management of irrigated crop rotations. Soil Tillage Res. 2017, 168, 1–10. [Google Scholar] [CrossRef]
- Cordeiro, C.F.D.S.; Rodrigues, D.R.; Rocha, C.H.; Araujo, F.F.; Echer, F.R. Glomalin and microbial activity affected by cover crops and nitrogen management in sandy soil with cotton cultivation. Appl. Soil Ecol. 2021, 167, 104026. [Google Scholar] [CrossRef]
- Calderón, F.J.; Nielsen, D.; Acosta-Martínez, V.; Vigil, M.F.; Lyon, D. Cover Crop and Irrigation Effects on Soil Microbial Communities and Enzymes in Semiarid Agroecosystems of the Central Great Plains of North America. Pedosphere 2016, 26, 192–205. [Google Scholar] [CrossRef]
- Bending, G.D.; Turner, M.K.; Jones, J.E. Interactions between crop residue and soil organic matter quality and the functional diversity of soil microbial communities. Soil Biol. Biochem. 2002, 34, 1073–1082. [Google Scholar] [CrossRef]
- Woodward, J. Synergism in cellulase systems. Bioresour. Technol. 1991, 36, 67–75. [Google Scholar] [CrossRef]
- Ekenler, M.; Tabatabai, M.A. β-Glucosaminidase Activity as an Index of Nitrogen Mineralization in Soils. Commun. Soil Sci. Plant Anal. 2004, 35, 1081–1094. [Google Scholar] [CrossRef]
- Turner, B.L.; McKelvie, I.D.; Haygarth, P.M. Characterisation of water-extractable soil organic phosphorus by phosphatase hydrolysis. Soil Biol. Biochem. 2002, 34, 27–35. [Google Scholar] [CrossRef]
- Guilbault, G.G.; Kramer, D.N. Fluorometric Determination of Lipase, Acylase, Alpha-, and Gamma-Chymotrypsin and Inhibitors of These Enzymes. Anal. Chem. 1964, 36, 409–412. [Google Scholar] [CrossRef]
- Rotman, B.; Papermaster, B.W. Membrane properties of living mammalian cells as studied by enzymatic hydrolysis of fluorogenic esters. Proc. Natl. Acad. Sci. USA 1966, 55, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Webber, C.L.; Gebhardt, M.R.; Kerr, H.D. Effect of Tillage on Soybean Growth and Seed Production. Agron. J. 1987, 79, 952–956. [Google Scholar] [CrossRef]
- Tyler, H.L. Shifts in bacterial community in response to conservation management practices within a soybean production system. Biol. Fertil. Soils 2021, 57, 1–12. [Google Scholar] [CrossRef]
- Bakker, M.G.; Acharya, J.; Moorman, T.B.; Robertson, A.E.; Kaspar, T.C. The Potential for Cereal Rye Cover Crops to Host Corn Seedling Pathogens. Phytopathology 2016, 106, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Sturz, A.V.; Carter, M.R.; Johnston, H.W. A review of plant disease, pathogen interactions and microbial antagonism under conservation tillage in temperate humid agriculture. Soil Tillage Res. 1997, 41, 169–189. [Google Scholar] [CrossRef]
- Singh, G.; Thilakarathne, A.D.G.M.; Williard, K.W.J.; Schoonover, J.E.; Cook, R.L.; Gage, K.L.; McElroy, R. Tillage and legume non-legume cover cropping effects on corn–soybean production. Agron. J. 2020, 112, 2636–2648. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | 2018 | 2019 | ||||
|---|---|---|---|---|---|---|
| pH | SM | SOM | pH | SM | SOM | |
| Till Rye | 6.5 ± 0.07 | 17.0 ± 0.32 | 4.9 ± 0.49 | 6.4 ± 0.17 | 14.8 ± 0.34 | 4.9 ± 0.42 |
| Till Rye+Clover | 6.6 ± 0.07 | 16.5 ± 0.36 | 4.8 ± 0.42 | 6.4 ± 0.07 | 13.9 ± 0.95 | 4.8 ± 0.41 |
| Till No Cover | 6.7 ± 0.08 | 15.0 ± 0.88 | 4.8 ± 0.43 | 6.8 ± 0.04 | 14.8 ± 1.06 | 4.7 ± 0.42 |
| No-Till Rye | 6.7 ± 0.08 | 17.0 ± 0.32 | 4.8 ± 0.51 | 6.4 ± 0.16 | 14.2 ± 0.98 | 5.0 ± 0.47 |
| No-Till Rye+Clover | 6.7 ± 0.14 | 13.9 ± 1.94 | 5.2 ± 0.70 | 6.3 ± 0.23 | 13.4 ± 0.54 | 5.2 ± 0.64 |
| No-Till No Cover | 6.6 ± 0.04 | 16.0 ± 0.81 | 4.9 ± 0.57 | 6.4 ± 0.18 | 14.5 ± 1.81 | 5.0 ± 0.47 |
| Treatment | 2018 | 2019 | ||
|---|---|---|---|---|
| MBC | MBN | MBC | MBN | |
| Till Rye | 0.066 ± 0.008 | 0.0031 ± 0.0008 | 0.369 ± 0.013 | 0.02 ± 0.001 |
| Till Rye+Clover | 0.066 ± 0.004 | 0.0041 ± 0.0001 | 0.316 ± 0.014 | 0.017 ± 0.002 |
| Till No Cover | 0.068 ± 0.006 | 0.0046 ± 0.0006 | 0.305 ± 0.018 | 0.015 ± 0.002 |
| No-Till Rye | 0.067 ± 0.005 | 0.0031 ± 0.0012 | 0.336 ± 0.021 | 0.018 ± 0.002 |
| No-Till Rye+Clover | 0.066 ± 0.005 | 0.0038 ± 0.0006 | 0.33 ± 0.026 | 0.018 ± 0.004 |
| No-Till No Cover | 0.069 ± 0.001 | 0.0041 ± 0.0006 | 0.356 ± 0.052 | 0.019 ± 0.004 |
| 2018 | 2019 | |||||
|---|---|---|---|---|---|---|
| Spring | Summer | Fall | Spring | Summer | Fall | |
| β-glucosidase | 0.431 * | 0.618 ** | 0.407 * | 0.567 ** | 0.636 *** | 0.526 ** |
| Cellobiohydrolase | 0.598 ** | 0.716 **** | 0.635 *** | 0.637 *** | 0.805 **** | 0.712 **** |
| NAGase | 0.407 * | 0.509 * | 0.378 | 0.525 ** | 0.769 **** | 0.481 * |
| Phosphatase | 0.310 | 0.429 * | 0.361 | 0.534 ** | 0.609 ** | 0.515 * |
| FDA Hydrolysis | 0.405 * | 0.678 *** | 0.515 * | 0.302 | 0.448 * | 0.658 *** |
| MBC | 0.299 | 0.509 * | 0.352 | 0.308 | 0.630 ** | 0.390 |
| MBN | 0.279 | 0.718 **** | 0.391 | 0.255 | 0.696 *** | 0.316 |
| pH | SM | SOM | MBC | MBN | BG | Cello | NAGase | Phosphatase | FDA | |
|---|---|---|---|---|---|---|---|---|---|---|
| pH | 1 | −0.099 | −0.187 | −0.224 | 0.027 | −0.033 | −0.145 | −0.151 | −0.231 | −0.179 |
| SM | 1 | 0.107 | 0.649 | 0.465 | 0.582 | 0.561 | 0.581 | 0.463 | 0.593 | |
| SOM | 1 | 0.43 | 0.256 | 0.458 | 0.368 | 0.296 | 0.486 | 0.183 | ||
| MBC | 1 | 0.822 | 0.644 | 0.697 | 0.608 | 0.695 | 0.615 | |||
| MBN | 1 | 0.5 | 0.554 | 0.419 | 0.459 | 0.36 | ||||
| BG | 1 | 0.922 | 0.873 | 0.868 | 0.777 | |||||
| Cello | 1 | 0.887 | 0.854 | 0.838 | ||||||
| NAGase | 1 | 0.813 | 0.85 | |||||||
| Phosphatase | 1 | 0.78 | ||||||||
| FDA | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tyler, H.L. Single- versus Double-Species Cover Crop Effects on Soil Health and Yield in Mississippi Soybean Fields. Agronomy 2021, 11, 2334. https://doi.org/10.3390/agronomy11112334
Tyler HL. Single- versus Double-Species Cover Crop Effects on Soil Health and Yield in Mississippi Soybean Fields. Agronomy. 2021; 11(11):2334. https://doi.org/10.3390/agronomy11112334
Chicago/Turabian StyleTyler, Heather L. 2021. "Single- versus Double-Species Cover Crop Effects on Soil Health and Yield in Mississippi Soybean Fields" Agronomy 11, no. 11: 2334. https://doi.org/10.3390/agronomy11112334
APA StyleTyler, H. L. (2021). Single- versus Double-Species Cover Crop Effects on Soil Health and Yield in Mississippi Soybean Fields. Agronomy, 11(11), 2334. https://doi.org/10.3390/agronomy11112334