
Utilisation of Miscanthus x giganteus L. Based C-Rich Fertilisers for N Immobilisation and Microbial Biomass Build-Up in a Crop Rotation


Abstract
:1. Introduction
2. Materials and Methods
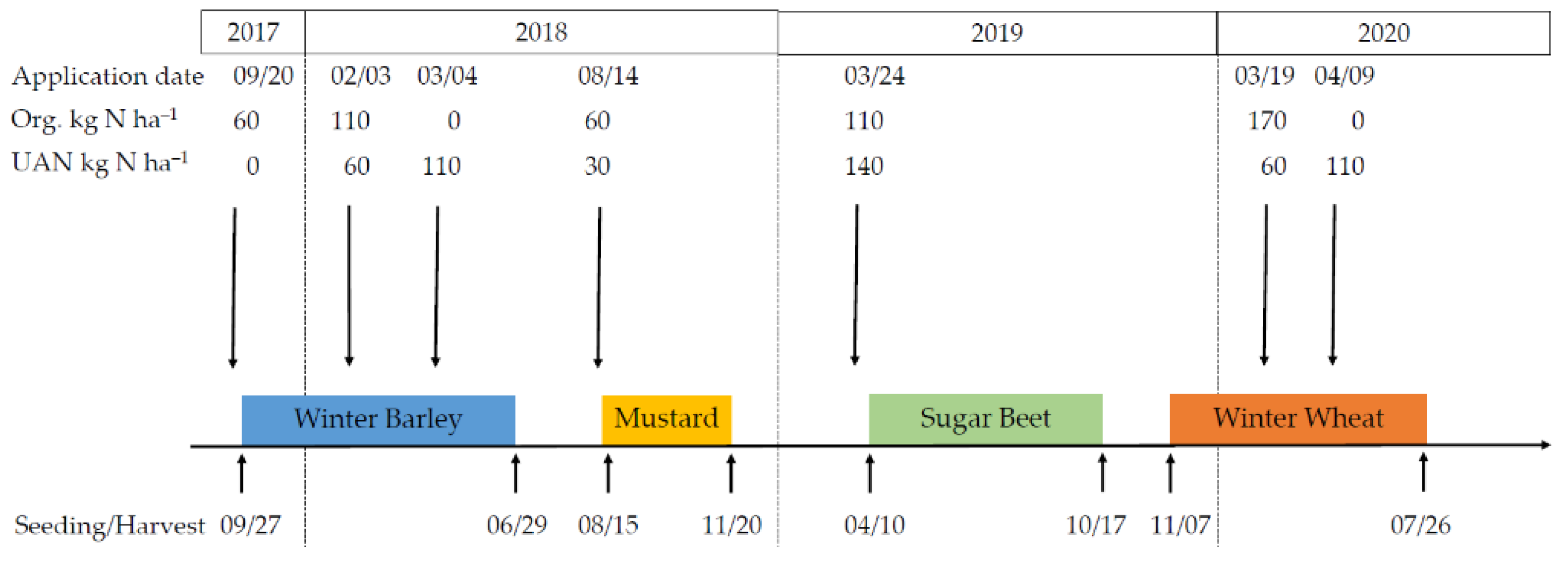
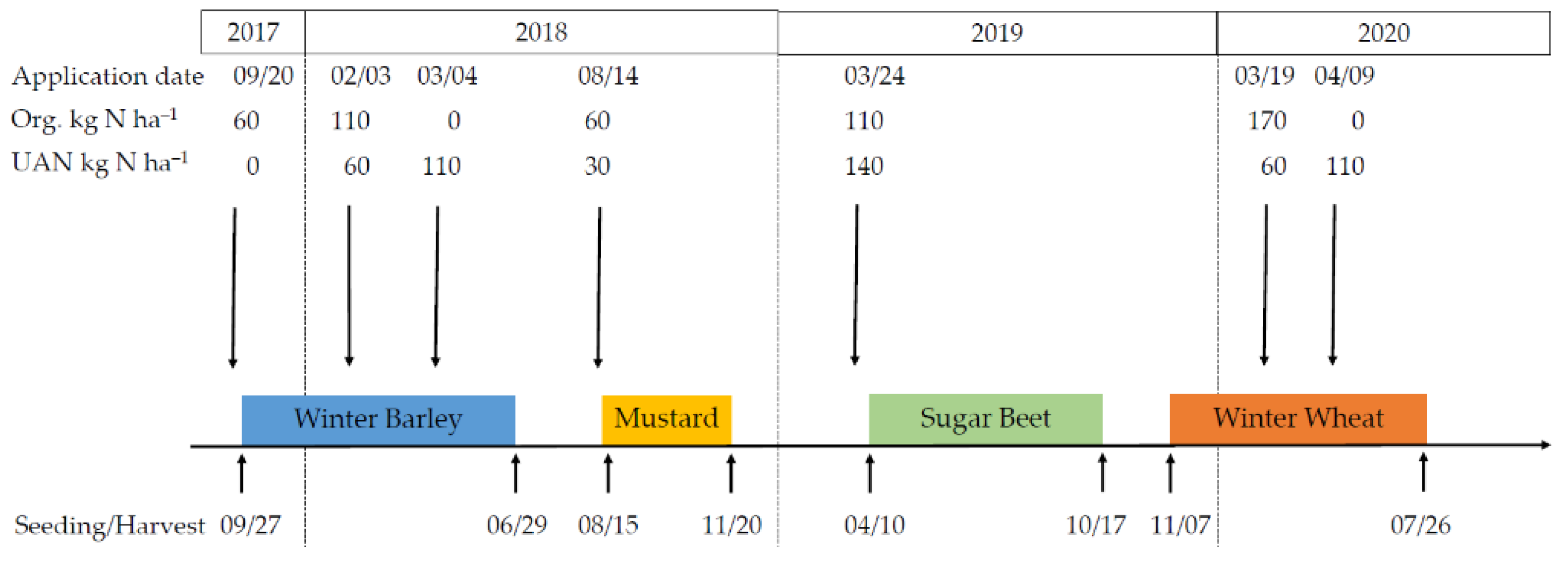
2.1. Site Description
2.2. Soil Analyses
2.3. Plant Analyses
2.4. Statistical Analyses
3. Results
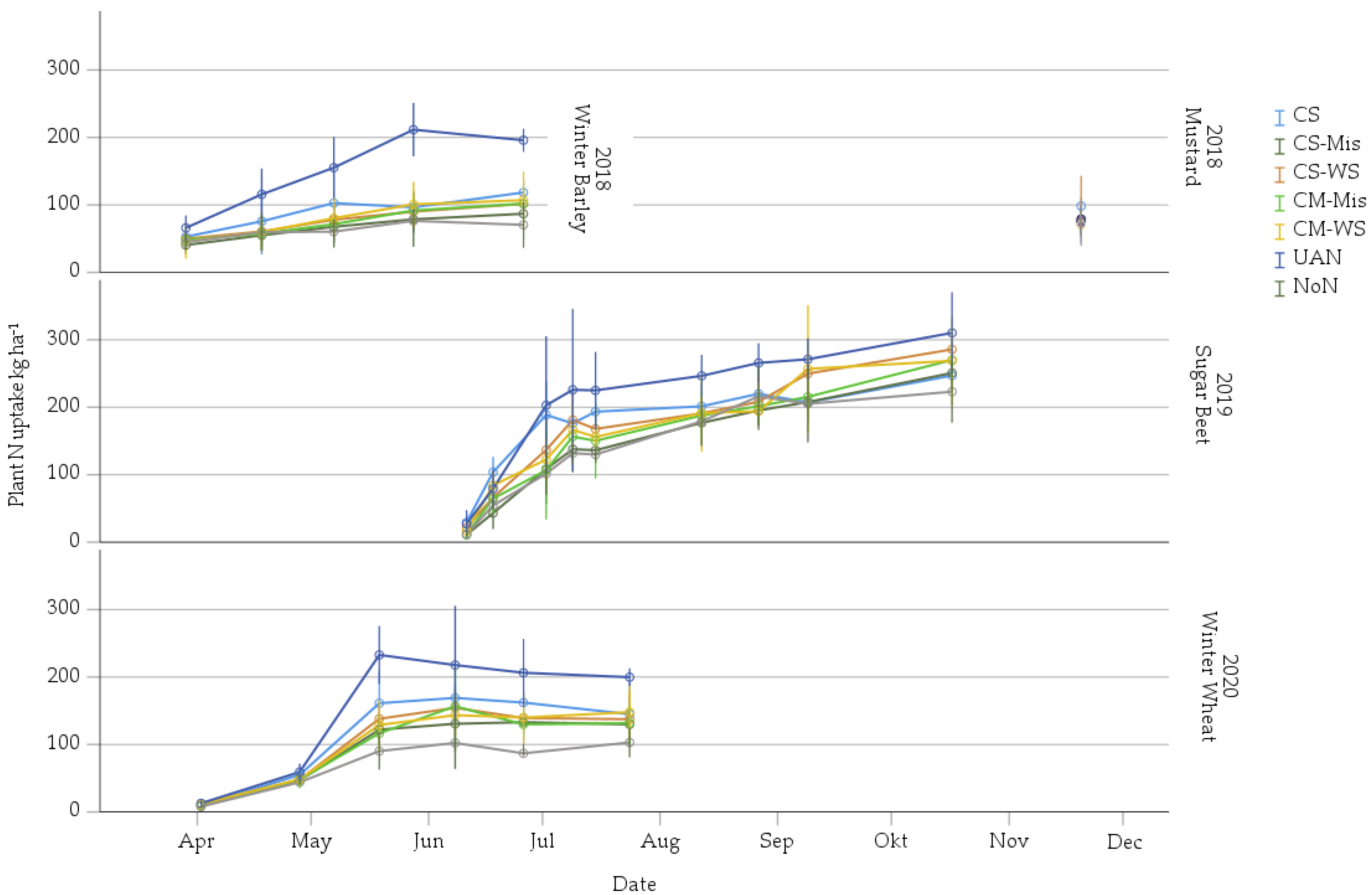
3.1. Plant N Uptake
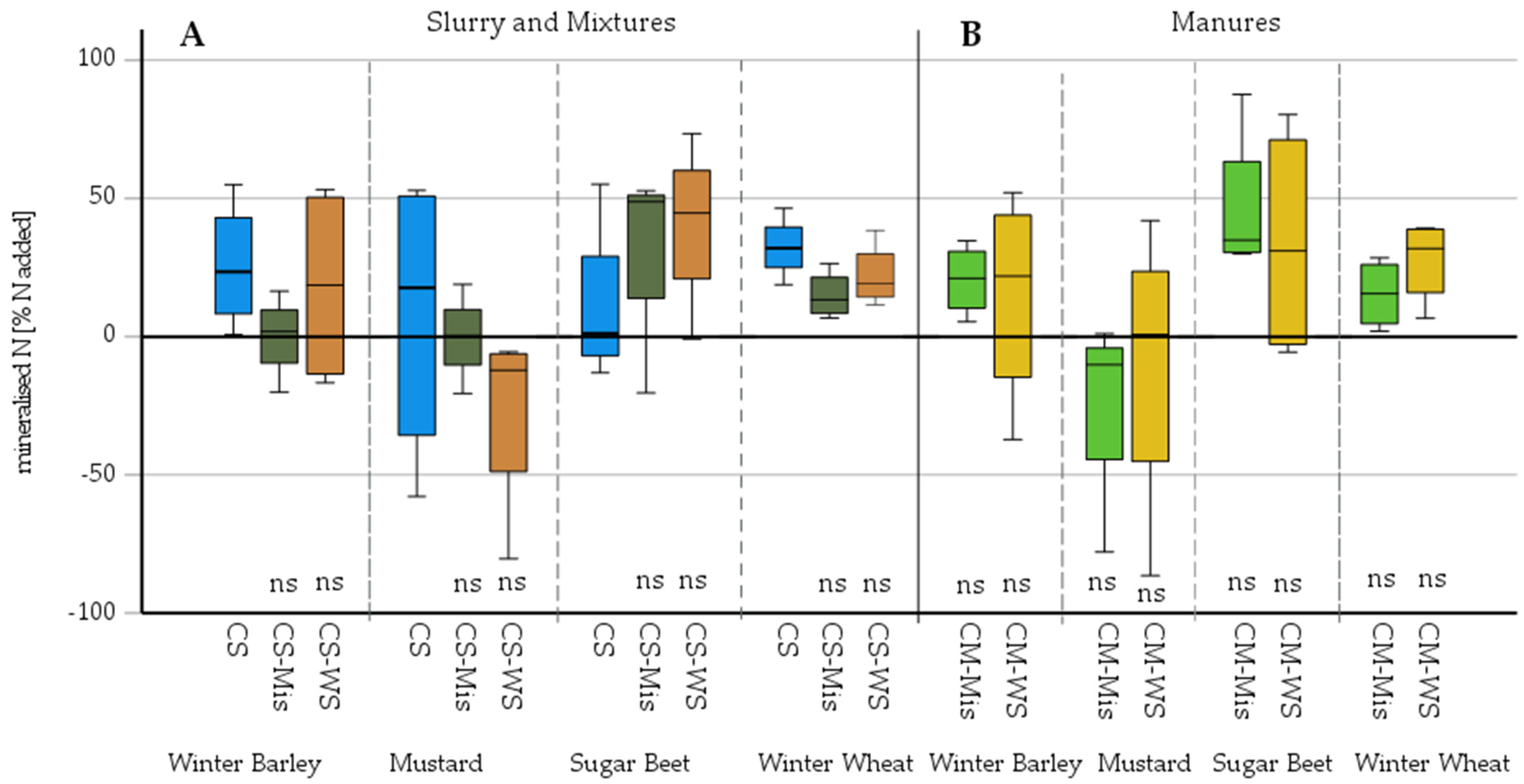
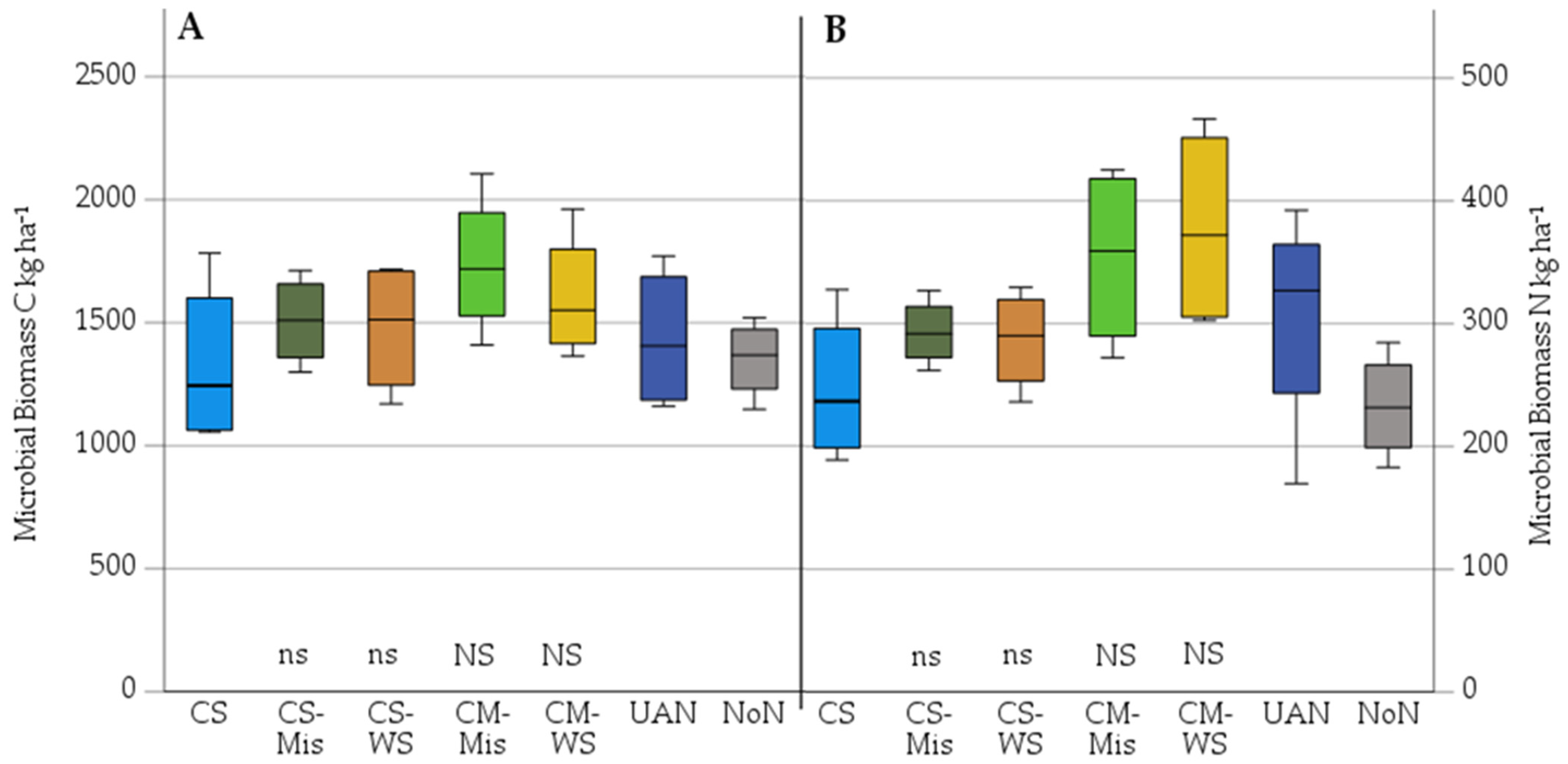
3.2. Microbial Mineralisation–Immobilisation as Affected by Added Miscanthus Straw
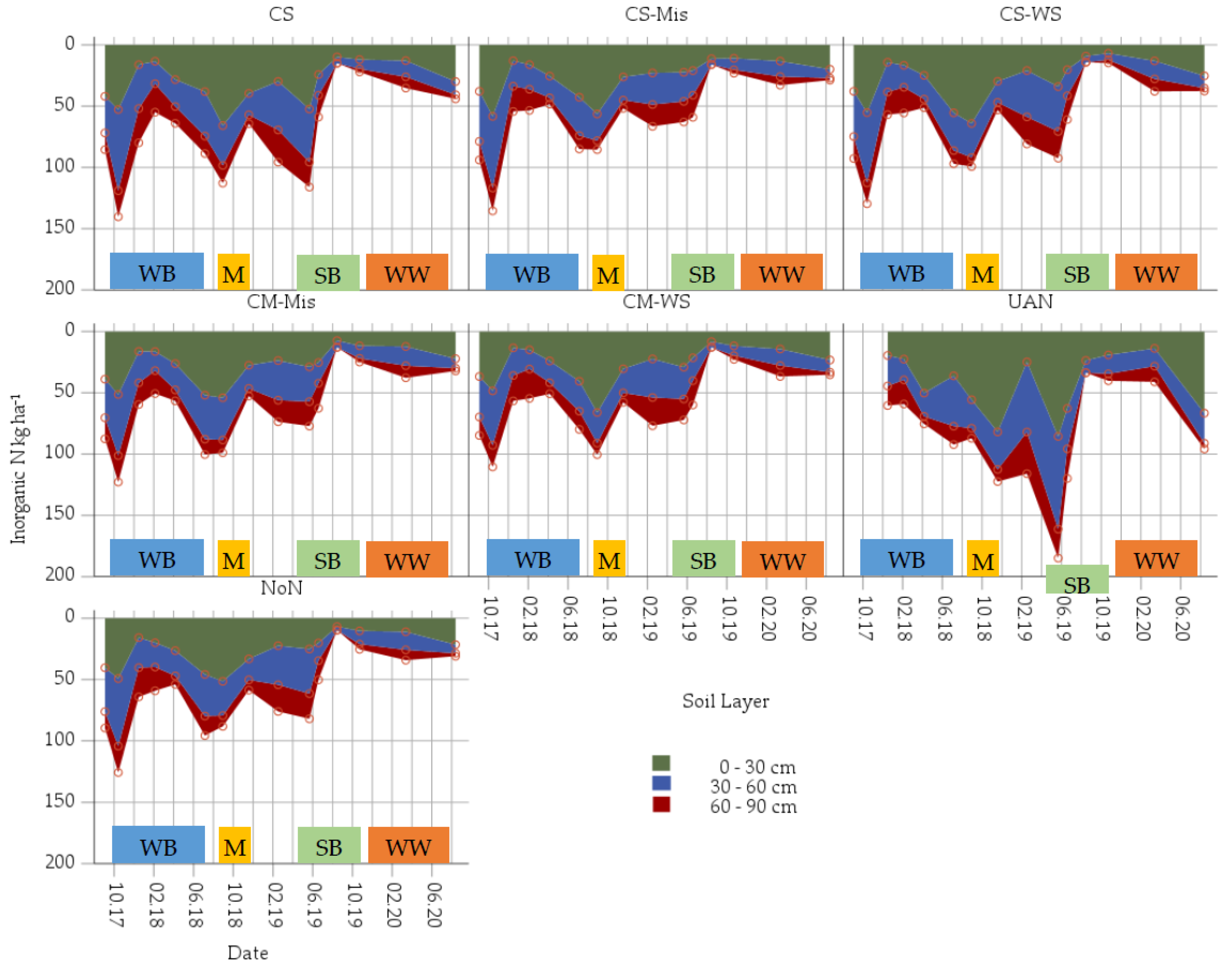
3.3. Inorganic N as Affected by Added Miscanthus Straw
3.4. Yield and Quality Parameters as Affected by Added Miscanthus Straw
4. Discussion
4.1. Miscanthus as C Source for Microbial N Immobilisation
4.2. Miscanthus as C Source for Microbial-Derived C Sequestration
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bouwman, L.; Goldewijk, K.K.; van der Hoek, K.W.; Beusen, A.; van Vuuren, D.; Willems, J.; Rufino, M.; Stehfest, E. Exploring global changes in nitrogen and phosphorus cycles in agriculture induced by livestock production over the 1900–2050 period. Proc. Natl. Acad. Sci. USA 2013, 110, 20882–20887. [Google Scholar] [CrossRef] [Green Version]
- Deutschland Statistisches Bundesamt. Produzierendes Gewerbe Düngemittelversorgung Wirtschaftsjahr 2019/2020; Fachserie 4, Reihe 8.2; Destatis: Wiesbaden, Germany, 2020; pp. 1–24.
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef]
- Cordell, D.; Drangert, J.-O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Galloway, J.N.; Dentener, F.J.; Capone, D.G.; Boyer, E.W.; Howarth, R.W.; Seitzinger, S.P.; Asner, G.P.; Cleveland, C.C.; Green, P.A.; Holland, E.A.; et al. Nitrogen Cycles: Past, Present, and Future. Biogeochemistry 2004, 70, 153–226. [Google Scholar] [CrossRef]
- Good, A.G.; Shrawat, A.K.; Muench, D.G. Can less yield more? Is reducing nutrient input into the environment compatible with maintaining crop production? Trends Plant Sci. 2004, 9, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Hietz, P.; Turner, B.J.; Wanek, W.; Richter, A.; Nock, C.A.; Wright, S.J. The influence of Late Quaternary climate-change velocity on species endemism. Science 2011, 334, 664–666. [Google Scholar] [CrossRef] [Green Version]
- Nacry, P.; Bouguyon, E.; Gojon, A. Nitrogen acquisition by roots: Physiological and developmental mechanisms ensuring plant adaptation to a fluctuating resource. Plant Soil 2013, 370, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Goidts, E.; van Wesemael, B. Regional assessment of soil organic carbon changes under agriculture in Southern Belgium (1955–2005). Geoderma 2007, 141, 341–354. [Google Scholar] [CrossRef]
- Sleutel, S.; de Neve, S.; Hofman, G. Assessing causes of recent organic carbon losses from cropland soils by means of regionalscaled input balances for the case of Flanders (Belgium). Nutr. Cycl. Agroecosyst. 2007, 78, 265–278. [Google Scholar] [CrossRef]
- Meersmans, J.; van Wesemael, B.; Goidts, E.; van Molle, M.; de Baets, S.; de Ridder, F. Spatial analysis of soil organic carbon evolution in Belgian croplands and grasslands, 1960–2006. Glob. Chang. Biol. 2010, 17, 466–479. [Google Scholar] [CrossRef]
- Steinmann, T.; Welp, G.; Holbeck, B.; Amelung, W. Long-term development of organic carbon contents in arable soil of North Rhine-Westphalia, Germany, 1979–2015. Eur. J. Soil Sci. 2016, 67, 616–623. [Google Scholar] [CrossRef]
- Steinmann, T.; Welp, G.; Wolf, A.; Holbeck, B.; Amelung, W.; Große-Rüschkamp, T. Repeated monitoring of organic carbon stocks after eight years reveals carbon losses from intensively managed agricultural soils in Western Germany. J. Plant Nutr. Soil Sci. 2016, 179, 355–366. [Google Scholar] [CrossRef]
- Greef, J.; Deuter, M.; Jung, C.; Schondelmaier, J. Genetic diversity of European Miscanthus species revealed by AFLP fingerprinting. Genet. Resour. Crop Evol. 1997, 44, 185–195. [Google Scholar] [CrossRef]
- Hodkinson, T.R.; Renvoize, S. Nomenclature of Miscanthus x giganteus (Poaceae). Kew Bull. 2001, 56, 759–760. [Google Scholar] [CrossRef]
- Felten, D.; Emmerling, C. Accumulation of Miscanthus-derived carbon in soils in relation to soil depth and duration of land use under commercial farming conditions. Z. Pflanzenernähr. Bodenk. 2012, 175, 661–670. [Google Scholar] [CrossRef]
- Geisseler, D.; Horwath, W.R. Investigating amino acid utilization by soil microorganisms using compound specific stable isotope analysis. Soil Biol. Biochem. 2014, 74, 100–105. [Google Scholar] [CrossRef]
- Geisseler, D.; Horwath, W.R.; Joergensen, R.G.; Ludwig, B. Pathways of nitrogen utilization by soil microorganisms—A review. Soil Biol. Biochem. 2010, 42, 2058–2067. [Google Scholar] [CrossRef]
- Joergensen, R.; Wichern, F. Quantitative assessment of the fungal contribution to microbial tissue in soil. Soil Biol. Biochem. 2008, 40, 2977–2991. [Google Scholar] [CrossRef]
- Dilly, O.; Blume, H.-P.; Munch, J.C. Soil Microbial Activities in Luvisols and Anthrosols during 9 Years of Region-Typical Tillage and Fertilisation Practices in Northern Germany. Biogeochemistry 2003, 65, 319–339. [Google Scholar] [CrossRef]
- Daudén, A.; Quílez, D.; Martínez, C. Residual effects of pig slurry applied to a Mediterranean soil on yield and N uptake of a subsequent wheat crop. Soil Use Manag. 2004, 20, 156–162. [Google Scholar] [CrossRef]
- Sørensen, P. Immobilisation, remineralisation and residual effects in subsequent crops of dairy cattle slurry nitrogen compared to mineral fertiliser nitrogen. Plant Soil 2004, 267, 285–296. [Google Scholar] [CrossRef]
- Sørensen, P.; Thomsen, I.K. Separation of Pig Slurry and Plant Utilization and Loss of Nitrogen-15-labeled Slurry Nitrogen. Soil Sci. Soc. Am. J. 2005, 69, 1644–1651. [Google Scholar] [CrossRef]
- Eurostat. Agri-Environmental Indicator—Nitrate Pollution of Water. Archive—Agriculture. 2012. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=Archive:Agri-environmental_indicator_-_nitrate_pollution_of_water (accessed on 3 September 2021).
- Thorup-Kristensen, K.; Magid, J.; Jensen, L.S. Catch crops and green manures as biological tools in nitrogen management in temperate zones. Adv. Agron. 2003, 79, 227–302. [Google Scholar]
- Fritz, C.; Wichern, F. In the land of plenty: Catch crops trigger nitrogen uptake by soil microorganisms. Plant Soil 2018, 423, 549–562. [Google Scholar] [CrossRef]
- Meier, I.C.; Finzi, A.C.; Phillips, R.P. Root exudates increase N availability by stimulating microbial turnover of fast-cycling N pools. Soil Biol. Biochem. 2017, 106, 119–128. [Google Scholar] [CrossRef] [Green Version]
- García-Ruiz, R.; Carranza-Gallego, G.; Aguilera, E.; de Molina, M.G.; Guzmán, G.I. C and N mineralisation of straw of traditional and modern wheat varieties in soils of contrasting fertility. Nutr. Cycl. Agroecosyst. 2019, 113, 167–179. [Google Scholar] [CrossRef]
- Nishio, T.; Oka, N. Effect of Organic matter application on the fate of 15 N-labeled ammonium fertilizer in an upland soil. Soil Sci. Plant Nutr. 2003, 49, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Shindo, H.; Nishio, T. Immobilization and remineralization of N following addition of wheat straw into soil: Determination of gross N transformation rates by 15N-ammonium isotope dilution technique. Soil Biol. Biochem. 2005, 37, 425–432. [Google Scholar] [CrossRef]
- Šimon, T.; Kunzová, E.; Friedlová, M. The effect of digestate, cattle slurry and mineral fertilization on the winter wheat yield and soil quality parameters. Plant Soil Environ. 2016, 61, 522–527. [Google Scholar] [CrossRef] [Green Version]
- Reichel, R.; Wei, J.; Islam, M.S.; Schmid, C.; Wissel, H.; Schröder, P.; Schloter, M.; Brüggemann, N. Potential of Wheat Straw, Spruce Sawdust, and Lignin as High Organic Carbon Soil Amendments to Improve Agricultural Nitrogen Retention Capacity: An Incubation Study. Front. Plant Sci. 2018, 9, 900. [Google Scholar] [CrossRef]
- Wei, J.; Reichel, R.; Islam, M.S.; Wissel, H.; Amelung, W.; Brüggemann, N. Chemical Composition of High Organic Carbon Soil Amendments Affects Fertilizer-Derived N2O Emission and Nitrogen Immobilization in an Oxic Sandy Loam. Front. Environ. Sci. 2020, 8, 115. [Google Scholar] [CrossRef]
- Stotter, M.; Wichern, F.; Pude, R.; Hamer, M. Nitrogen Immobilisation and Microbial Biomass Build-Up Induced by Miscanthus × giganteus L. Based Fertilisers. Agronomy 2021, 11, 1386. [Google Scholar] [CrossRef]
- Pude, R. Nachwachsende Rohstoffe aus der region und für die region. Berichte Über Zeitschrift Agrarpolitik Landwirtschaft Aktuelle Beiträge 2021, 99, 1–12. [Google Scholar] [CrossRef]
- Publications Office of the European Union. Regulation (EU) No. 2017/2393 of the European Parliament and of the Council-of 13 December 2017-Amending Regulations (EU) No 1305/2013 on Support for Rural Development by the European Agricultural Fund for Rural Development (EAFRD), (EU) No 1306/2013 on the Financing, Management and Monitoring of the Common Agricultural Policy, (EU) No 1307/2013 Establishing Rules for Direct Payments to Farmers under Support Schemes within the Framework of the Common Agricultural Policy, (EU) No 1308/2013 Establishing a Common Organisation of the Markets in Agricultural Products and (EU) No 652/2014 Laying Down Provisions for the Management of Expenditure Relating to the Food Chain, Animal Health and Animal Welfare, and Relating to Plant Health and Plant Reproductive Material; Publications Office of the European Union: Luxembourg, 2017; pp. 15–49. [Google Scholar]
- Emmerling, C.; Pude, R. Introducing Miscanthus to the greening measures of the EU Common Agricultural Policy. GCB Bioenergy 2017, 9, 274–279. [Google Scholar] [CrossRef]
- Bellamy, P.E.; Croxton, P.J.; Heard, M.S.; Hinsley, S.A.; Hulmes, L.; Hulmes, S.; Nuttall, P.; Pywell, R.F.; Rothery, P. The impact of growing Miscanthus for biomass on farmland bird populations. Biomass Bioenergy 2009, 33, 191–199. [Google Scholar] [CrossRef]
- Semere, T.; Slater, F.M. Ground flora, small mammal and bird species diversity in Miscanthus (Miscanthus × giganteus) and reed canary-grass (Phalaris arundinacea) fields. Biomass Bioenergy 2007, 31, 20–29. [Google Scholar] [CrossRef]
- Butler, S.J.; Boccaccio, L.; Gregory, R.D.; Vorisek, P.; Norris, K. Quantifying the impact of land-use change to European farmland bird populations. Agric. Ecosyst. Environ. 2010, 137, 348–357. [Google Scholar] [CrossRef]
- Flade, M.; Schwarz, J. Bestandsentwicklung von Vogelarten der Agrarlandschaft in Deutschland 1991–2010 und Schlüsselfaktoren. Fachgespräch „Agrarvögel—Ökologische Bewertungsgrundlage für Biodiversitätsziele in Ackerbaugebieten”, Kleinmachnow, Germany, 1–2 März 2013. Julius Kühn Archiv 2013, 442, 8–17. [Google Scholar] [CrossRef]
- Green, R.E.; Cornell, S.J.; Scharlemann, J.P.W.; Balmford, A. Farming and the fate of wild nature. Science 2005, 307, 550–555. [Google Scholar] [CrossRef] [Green Version]
- Gregory, R.D.; van Strien, A.; Vorisek, P.; Gmelig Meyling, A.W.; Noble, D.G.; Foppen, R.P.B.; Gibbons, D.W. Developing indicators for European birds. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 269–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruf, T.; Emmerling, C. Impact of premature harvest of Miscanthus x giganteus for biogas production on organic residues, microbial parameters and earthworm community in soil. Appl. Soil Ecol. 2017, 114, 74–81. [Google Scholar] [CrossRef]
- Schmidt, A.; Lemaigre, S.; Ruf, T.; Delfosse, P.; Emmerling, C. Miscanthus as biogas feedstock: Influence of harvest time and stand age on the biochemical methane potential (BMP) of two different growing seasons. Biomass Conv. Bioref. 2018, 8, 245–254. [Google Scholar] [CrossRef]
- Nguyen, V.T.H.; Elfers, J.; Kühn, H.; Kraska, T.; Pude, R. Different Miscanthus genotypes as growing media in soilless tomato cultivation and its subsequent use for combustion. Acta Hortic. 2021, 1305, 301–308. [Google Scholar] [CrossRef]
- Nowak, A.; Slizewska, K.; Gajecka, M.; Piotrowska, M.; Zakowska, Z.; Zielonka, L.; Gajecki, M. The genotoxicity of caecal water from gilts following experimentally induced Fusarium mycotoxicosis. Veterinarni Med. 2016, 60, 133–140. [Google Scholar] [CrossRef] [Green Version]
- ISO; DIN. Bodenbeschaffenheit–Bestimmung der Partikelgrößenverteilung in Mineralböden-Verfahren Mittels Siebung und Sedimentation; Deutsches Institut für Normung e. V.: Berlin, Germany, 2002; p. 11277. [Google Scholar]
- VDLUFA. Methode A 5.1.1. bestimmung des pH-wertes. In Methodenbuch I Die Untersuchung von Böden; VDLUFA-Verlag: Darmstadt, Germany, 2016. [Google Scholar]
- VDLUFA. Methode A 6.2.1.1. Bestimmung von phosphor und kalium im calcium-acetat-lactat-auszug. In Methodenbuch I Die Untersuchung von Böden; VDLUFA-Verlag: Darmstadt, Germany, 2012. [Google Scholar]
- VDLUFA. Methode A 6.2.4.1. Bestimmung des pflanzenverfügbaren Magnesiums im calciumchlorid-Auszug. In Methodenbuch I Die Untersuchung von Böden; VDLUFA-Verlag: Darmstadt, Germany, 1997. [Google Scholar]
- VDLUFA. Methode A 6.4.1. Bestimmung von magnesium, natrium, und den spurennährstoffen kupfer, mangan, zink und bor im calciumchlorid/DTPA-auszug. In Methodenbuch I Die Untersuchung von Böden; VDLUFA-Verlag: Darmstadt, Germany, 2002. [Google Scholar]
- ISO; DIN. Bodenbeschaffenheit-Bestimmung von Organischem Kohlenstoff und Gesamtkohlenstoff nach Trockener Verbrennung (Elementaranalyse); Deutsches Institut für Normung e. V.: Berlin, Germany, 1996; p. 10694. [Google Scholar]
- ISO; DIN. Bodenbeschaffenheit-Bestimmung des Gesamt-Stickstoffs Durch Trockene Verbrennung (Elementaranalyse); Deutsches Institut für Normung e. V.: Berlin, Germany, 1998; p. 13878. [Google Scholar]
- Vance, E.; Brookes, P.; Jenkinson, D. Microbial biomass measurements in forest soils: Determination of kC values and tests of hypotheses to explain the failure of the chloroform fumigation-incubation method in acid soils. Soil Biol. Biochem. 1987, 19, 689–696. [Google Scholar] [CrossRef]
- Brookes, P.; Landman, A.; Pruden, G.; Jenkinson, D. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Wu, J.; Joergensen, R.; Pommerening, B.; Chaussod, R.; Brookes, P. Measurement of soil microbial biomass C by fumigationextraction—An automated procedure. Soil Biol. Biochem. 1990, 19, 1167–1169. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Mueller, T. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kEN value. Soil Biol. Biochem. 1996, 28, 33–37. [Google Scholar] [CrossRef]
- Aiken, G.R. Dissolved organic matter in aquatic systems. In Comprehensive Water Quality and Purification; Elsevier: Amsterdam, The Netherlands, 2014; Volume 1, pp. 205–220. ISBN 9780123821836. [Google Scholar]
- Bhogal, A.; Williams, J.R.; Nicholson, F.A.; Chadwick, D.R.; Chambers, K.H.; Chambers, B.J. Mineralization of organic nitrogen from farm manure applications. Soil Use Manag. 2016, 32, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Cabrera, M.L.; Kissel, D.E.; Vigil, M.F. Nitrogen Mineralization from Organic Residues. J. Environ. Qual. 2005, 34, 75–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eiland, F.; Leth, M.; Klamer, M.; Lind, A.-M.; Jensen, H.E.K.; Iversen, J.J.L. C and N Turnover and Lignocellulose Degradation During Composting of Miscanthus Straw and Liquid Pig Manure. Compost. Sci. Util. 2001, 9, 186–196. [Google Scholar] [CrossRef]
- Corbeels, M.; Hofman, G.; van Cleemput, O. Nitrogen cycling associated with the decomposition of sunflower stalks and wheat straw in a Vertisol. Plant Soil 2000, 218, 71–82. [Google Scholar] [CrossRef]
- Rahn, C.R.; Bending, G.; Lillywhite, R.D.; Turner, M.K. Chemical characterisation of vegetable and arable crop residue materials: A comparison of methods. J. Sci. Food Agric. 1999, 79, 1715–1721. [Google Scholar] [CrossRef]
- Van Kuijk, S.J.A.; Sonnenberg, A.S.M.; Baars, J.J.P.; Hendriks, W.H.; del Río, J.C.; Rencoret, J.; Gutiérrez, A.; de Ruijter, N.C.A.; Cone, J.W. Chemical changes and increased degradability of wheat straw and oak wood chips treated with the white rot fungi Ceriporiopsis subvermispora and Lentinula edodes. Biomass Bioenergy 2017, 105, 381–391. [Google Scholar] [CrossRef]
- Dilly, O.; Bloem, J.; Vos, A.; Munch, J.C. Bacterial diversity in agricultural soils during litter decomposition. Appl. Environ. Microbiol. 2004, 70, 468–474. [Google Scholar] [CrossRef] [Green Version]
- Kallenbach, C.M.; Frey, S.D.; Grandy, A.S. Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat. Commun. 2016, 7, 13630. [Google Scholar] [CrossRef]
- Miltner, A.; Bombach, P.; Schmidt-Brücken, B.; Kästner, M. SOM genesis: Microbial biomass as a significant source. Biogeochemistry 2011, 111, 41–55. [Google Scholar] [CrossRef]
- Hobara, S.; Osono, T.; Hirose, D.; Noro, K.; Hirota, M.; Benner, R. The roles of microorganisms in litter decomposition and soil formation. Biogeochemistry 2014, 118, 471–486. [Google Scholar] [CrossRef]
- Khan, K.S.; Mack, R.; Castillo, X.; Kaiser, M.; Joergensen, R.G. Microbial biomass, fungal and bacterial residues, and their relationships to the soil organic matter C/N/P/S ratios. Geoderma 2016, 271, 115–123. [Google Scholar] [CrossRef]
- Liang, C.; Cheng, G.; Wixon, D.L.; Balser, T.C. An Absorbing Markov Chain approach to understanding the microbial role in soil carbon stabilization. Biogeochemistry 2010, 106, 303–309. [Google Scholar] [CrossRef]
- Bengtsson, J. Disturbance and resilience in soil animal communities. Eur. J. Soil Biol. 2002, 2002, 119–125. [Google Scholar] [CrossRef]
- Yesufu, J.; McCalmont, J.P.; Clifton-Brown, J.C.; Williams, P.; Hyland, J.; Gibbons, J.; Styles, D. Consequential life cycle assessment of Miscanthus livestock bedding, diverting straw to bioelectricity generation. GCB Bioenergy 2020, 12, 39–53. [Google Scholar] [CrossRef]
- Van Weyenberg, S.; Ulens, T.; de Reu, K.; Zwertvaegher, I.; Demeyer, P.; Pluym, L. Feasibility of Miscanthus as alternative bedding for dairy cows. Vet. Med. 2016, 60, 121–132. [Google Scholar] [CrossRef]
- Eiland, F.; Klamer, M.; Lind, A.-M.; Leth, M.; Bååth, E. Influence of Initial C/N Ratio on Chemical and Microbial Composition during Long Term Composting of Straw. Microb. Ecol. 2001, 41, 272–280. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH (H2O) | P (mg kg−1) | K (mg kg−1) | Mg (mg kg−1) |
| 6.3 ± 0.06 | 11.4 ± 2.7 | 10.4 ± 1.6 | 14 ± 1.9 |
| B (mg kg−1) | Cu (mg kg−1) | Mn (mg kg−1) | Fe (mg kg−1) |
| 0.5 ± 0.04 | 6.3 ± 0.5 | 169.4 ± 47.4 | 196.3 ± 18.6 |
| SOM (%) | SOC (%) | Nt (%) | C/N (ratio) |
| 3.9 ± 0.7 | 2.3 ± 0.4 | 0.27 ± 0.02 | 8.5 ± 1.2 |
| Clay (g kg−1) | Silt (g kg−1) | Sand (g kg−1) | |
| 229 | 597 | 173 |
| Test Parameter | Unit | CS 1 | CS–Mis 2 | CS–WS 2 | CM–Mis 2 | CM–WS 2 | UAN 1 | Mis 2 | WS 2 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Autumn 2017 | Dry matter | % | 9.7 | 20.9 | 16.4 | 32.7 | 25.4 | - | 87.8 | 86.2 |
| Organic matter | % | 7.2 | 18.3 | 13.4 | 29.2 | 17.3 | - | 85.2 | 79.2 | |
| Total N | kg m−3/kg t−1 | 4.6 | 4.2 | 5.1 | 5.6 | 8.3 | - | 1.7 | 6.3 | |
| NH4+-N | kg m−3/kg t−1 | 2.3 | 1.4 | 1.1 | 0.5 | 0.2 | - | <0.1 | 0.2 | |
| NH4+-N in total N | % | 50 | 33 | 22 | 9 | 2 | - | 5 | 3 | |
| pH | value | 7.3 | 8.4 | 8.2 | 8.3 | 8.2 | - | 6 | 6.8 | |
| C/N | ratio | 9 | 26 | 15 | 30 | 12 | - | 288 | 73 | |
| Spring 2018 | Dry matter | % | 9.2 | 20.1 | 16 | 32.5 | 26.1 | - | 87.8 | 86.2 |
| Organic matter | % | 6.7 | 17.6 | 13 | 29.8 | 22.6 | - | 85.2 | 79.2 | |
| Total N | kg m−3/kg t−1 | 4 | 3.9 | 4.7 | 6.5 | 6.4 | 358 | 1.7 | 6.3 | |
| NH4+-N | kg m−3/kg t−1 | 1.8 | 1 | 1 | 0.7 | 1.3 | 90 | <0.1 | 0.2 | |
| NH4+-N in total N | % | 45 | 26 | 21 | 11 | 20 | 25 | 5 | 3 | |
| pH | value | 7.7 | 8.1 | 7.8 | 8.2 | 8.3 | - | 6 | 6.3 | |
| C/N | ratio | 9.8 | 27 | 16 | 27 | 20 | - | 288 | 73 | |
| Autumn 2018 | Dry matter | % | 9.9 | 23.7 | 18.6 | 24.1 | 39.6 | - | 90.1 | 90.9 |
| Organic matter | % | 7.3 | 21 | 15.6 | 21.8 | 22.1 | - | 86.9 | 86.3 | |
| Total N | kg m−3/kg t−1 | 4.7 | 4.3 | 5.2 | 5.2 | 11.4 | 358 | 3 | 4.4 | |
| NH4+-N | kg m−3/kg t−1 | 2.2 | 1.1 | 1.4 | 1.4 | 2.1 | 90 | 0.2 | 0.2 | |
| NH4+-N in total N | % | 47 | 26 | 27 | 27 | 18 | 25 | 7 | 5 | |
| pH | value | - | - | - | - | - | - | - | - | |
| C/N | ratio | 8.9 | 28.2 | 17.5 | 24.5 | 11.2 | - | 166 | 114.8 | |
| Spring 2019 | Dry matter | % | 7 | 20.9 | 15.5 | 22.8 | 17.5 | - | 90.1 | 90.9 |
| Organic matter | % | 5.2 | 17.4 | 12.5 | 20.1 | 14.1 | - | 86.9 | 86.3 | |
| Total N | kg m−3/kg t−1 | 3.8 | 2.9 | 3.8 | 4.6 | 5.2 | 358 | 3 | 4.4 | |
| NH4+-N | kg m−3/kg t−1 | 2.1 | 0.4 | 0.4 | 0.8 | 1.6 | 90 | 0.2 | 0.2 | |
| NH4+-N in total N | % | 55 | 14 | 11 | 17 | 31 | 25 | 7 | 5 | |
| pH | value | 7.8 | 8.3 | 8.1 | 7.1 | 7.4 | - | 6.1 | 6.3 | |
| C/N | ratio | 7.9 | 35 | 19.2 | 25.3 | 15.7 | - | 166 | 114.8 | |
| Spring 2020 | Dry matter | % | 8.7 | 21.7 | 16.5 | 32.6 | 17.9 | - | 89.8 | 92.1 |
| Organic matter | % | 6.7 | 18.3 | 13.3 | 28.4 | 12.6 | - | 86.8 | 87.8 | |
| Total N | kg m−3/kg t−1 | 4.6 | 3.8 | 4.3 | 6.5 | 4.9 | 358 | 1.9 | 3.7 | |
| NH4+-N | kg m−3/kg t−1 | 2.1 | 0.6 | 0.8 | 0.7 | 1.3 | 90 | 0.1 | 0.2 | |
| NH4+-N in total N | % | 46 | 16 | 19 | 11 | 27 | 25 | 5 | 5 | |
| pH | value | 7.3 | 8.4 | 8.3 | 8.3 | 8.1 | - | 6 | 6.4 | |
| C/N | ratio | 8.4 | 27.6 | 17.9 | 25.5 | 14.8 | - | 262.3 | 136.8 |
| Sampling | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | Harvest * | ||
| Cultivar | Treatment | N Uptake (% of CS) | ||||||||
| Winter Barley | CS | 100 ns | 100 ns | 100 ns | 100 ns | 100 ns | ||||
| CS–Mis | 77 ns | 73 ns | 66 ns | 82 ns | 73 ns | |||||
| CS–WS | 94 ns | 80 ns | 76 ns | 93 ns | 86 ns | |||||
| Mustard | CS | 100 a | ||||||||
| CS–Mis | 88 b | |||||||||
| CS–WS | 93 b | |||||||||
| Sugar Beet | CS | 100 a | 100 a | 100 a | 100 ns | 100 a | 100 ns | 100 ns | 100 b | 100 b |
| CS–Mis | 038 b | 42 b | 58 b | 78 ns | 70 b | 88 ns | 89 ns | 100 b | 102 b | |
| CS–WS | 074 ab | 64 b | 73 ab | 103 ns | 87 ab | 95 ns | 95 ns | 121 a | 116 a | |
| Winter Wheat | CS | 100 ns | 100 ns | 100 ns | 100 a | 100 ns | 100 ns | |||
| CS–Mis | 101 ns | 85 ns | 76 ns | 77 b | 82 ns | 90 ns | ||||
| CS–WS | 118 ns | 87 ns | 86 ns | 91 ab | 86 ns | 95 ns | ||||
| Cumulated | CS | 100 ns | ||||||||
| CS–Mis | 90 ns | |||||||||
| CS–WS | 99 ns | |||||||||
| Sampling | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | Harvest * | ||
| Cultivar | Treatment | N Uptake (% of CM–WS) | ||||||||
| Winter Barley | CM–Mis | 106 ns | 96 ns | 89 ns | 91 ns | 95 ns | ||||
| CM–WS | 100 ns | 100 ns | 100 ns | 100 ns | 100 ns | |||||
| Mustard | CM–Mis | 109 ns | ||||||||
| CM–WS | 100 ns | |||||||||
| Sugar Beet | CM–Mis | 72 ns | 77 ns | 87 ns | 94 ns | 96 ns | 98 ns | 104 ns | 84 ns | 100 ns |
| CM–WS | 100 ns | 100 ns | 100 ns | 100 ns | 100 ns | 100 ns | 100 ns | 100 ns | 100 ns | |
| Winter Wheat | CM–Mis | 81 ns | 99 ns | 90 ns | 109 ns | 93 ns | 89 ns | |||
| CM–WS | 100 ns | 100 ns | 100 ns | 100 ns | 100 ns | 100 ns | ||||
| Cumulated | CM–Mis | 98 ns | ||||||||
| CM–WS | 100 ns | |||||||||
| Treatment | Winter Barley | Mustard | Sugar Beet | Winter Wheat | ||||
|---|---|---|---|---|---|---|---|---|
| Corn Yield Mg ha−1 | Protein % | Biomass Yield Mg ha−1 | Beet Yield Mg ha−1 | Sugar Yield Mg ha−1 | AmN Mmol kg−1 | Corn Yield Mg ha−1 | Protein % | |
| CS | 5.0 ± 1.2 ns | 11.0 ± 0.7 ns | 2.3 ± 0.9 ns | 84.3 ± 2.9 b | 11.2 ± 4.0 b | 21.2 ± 4.4 ns | 8.9 ± 0.6 ns | 10.0 ± 0.4 ns |
| CS–Mis | 4.7 ± 1.1 ns | 10.6 ± 0.1 ns | 1.8 ± 0.3 ns | 98.0 ± 4.4 a | 12.1 ± 4.5 ab | 14.3 ± 2.7 ns | 8.4 ± 0.2 ns | 9.4 ± 0.4 ns |
| CS–WS | 4.9 ± 0.5 ns | 10.4 ± 0.1 ns | 1.6 ± 0.3 ns | 99.3 ± 10.7 a | 12.2 ± 4.7 b | 16.9 ± 3.6 ns | 8.9 ± 0.6 ns | 9.5 ± 0.4 ns |
| CM–Mis | 4.8 ± 0.7 NS | 10.3 ± 0.3 NS | 1.8 ± 0.6 NS | 94.5 ± 12.2 NS | 12.0 ± 4.5 NS | 15.3 ± 3.0 NS | 8.8 ± 0.5 NS | 9.4 ± 0.4 NS |
| CM–WS | 4.9 ± 1.0 NS | 10.6 ± 0.1 NS | 1.7 ± 0.3 NS | 92.9 ± 9.4 NS | 12.0 ± 4.4 NS | 17.7 ± 3.8 NS | 8.8 ± 0.7 NS | 9.9 ± 0.3 NS |
| UAN | 6.9 ± 1.7 | 14.1 ± 0.5 | 1.2 ± 0.3 | 79.7 ± 4.7 | 9.9 ± 3.8 | 37.3 ± 4.9 | 9.1 ± 0.2 | 12.0 ± 0.3 |
| NoN | 3.9 ± 0.5 | 10.2 ± 0.3 | 1.5 ± 0.2 | 85.7 ± 13.0 | 11.3 ± 4.4 | 16.6 ± 3.4 | 7.2 ± 0.4 | 9.1 ± 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stotter, M.; Wichern, F.; Pude, R.; Hamer, M. Utilisation of Miscanthus x giganteus L. Based C-Rich Fertilisers for N Immobilisation and Microbial Biomass Build-Up in a Crop Rotation. Agronomy 2021, 11, 2390. https://doi.org/10.3390/agronomy11122390
Stotter M, Wichern F, Pude R, Hamer M. Utilisation of Miscanthus x giganteus L. Based C-Rich Fertilisers for N Immobilisation and Microbial Biomass Build-Up in a Crop Rotation. Agronomy. 2021; 11(12):2390. https://doi.org/10.3390/agronomy11122390
Chicago/Turabian StyleStotter, Michael, Florian Wichern, Ralf Pude, and Martin Hamer. 2021. "Utilisation of Miscanthus x giganteus L. Based C-Rich Fertilisers for N Immobilisation and Microbial Biomass Build-Up in a Crop Rotation" Agronomy 11, no. 12: 2390. https://doi.org/10.3390/agronomy11122390
APA StyleStotter, M., Wichern, F., Pude, R., & Hamer, M. (2021). Utilisation of Miscanthus x giganteus L. Based C-Rich Fertilisers for N Immobilisation and Microbial Biomass Build-Up in a Crop Rotation. Agronomy, 11(12), 2390. https://doi.org/10.3390/agronomy11122390