
Combining Process Modelling and LAI Observations to Diagnose Winter Wheat Nitrogen Status and Forecast Yield

 , , ,
, , , 
Abstract
:1. Introduction
- What is the performance of an intermediate complexity model for the simulation of LAI and yields when calibrated across a range of N fertilizer treatments and are the model parameter consistent across treatments?
- Can a single model calibration (developed from research Question 1) reliably reproduce destructively sampled leaf N content observations at each of the treatments when constrained by climate and LAI data?
- Can a single model calibration (developed from Question 1) reliably reproduce yield variation across each of the treatments when constrained by climate and LAI inputs and what is the relative information content of LAI and climate data?
- Can yield prediction from fusing the model and EO data potentially be accurate enough to support timely management interventions?
2. Materials and Methods
2.1. Field Site and Measurements
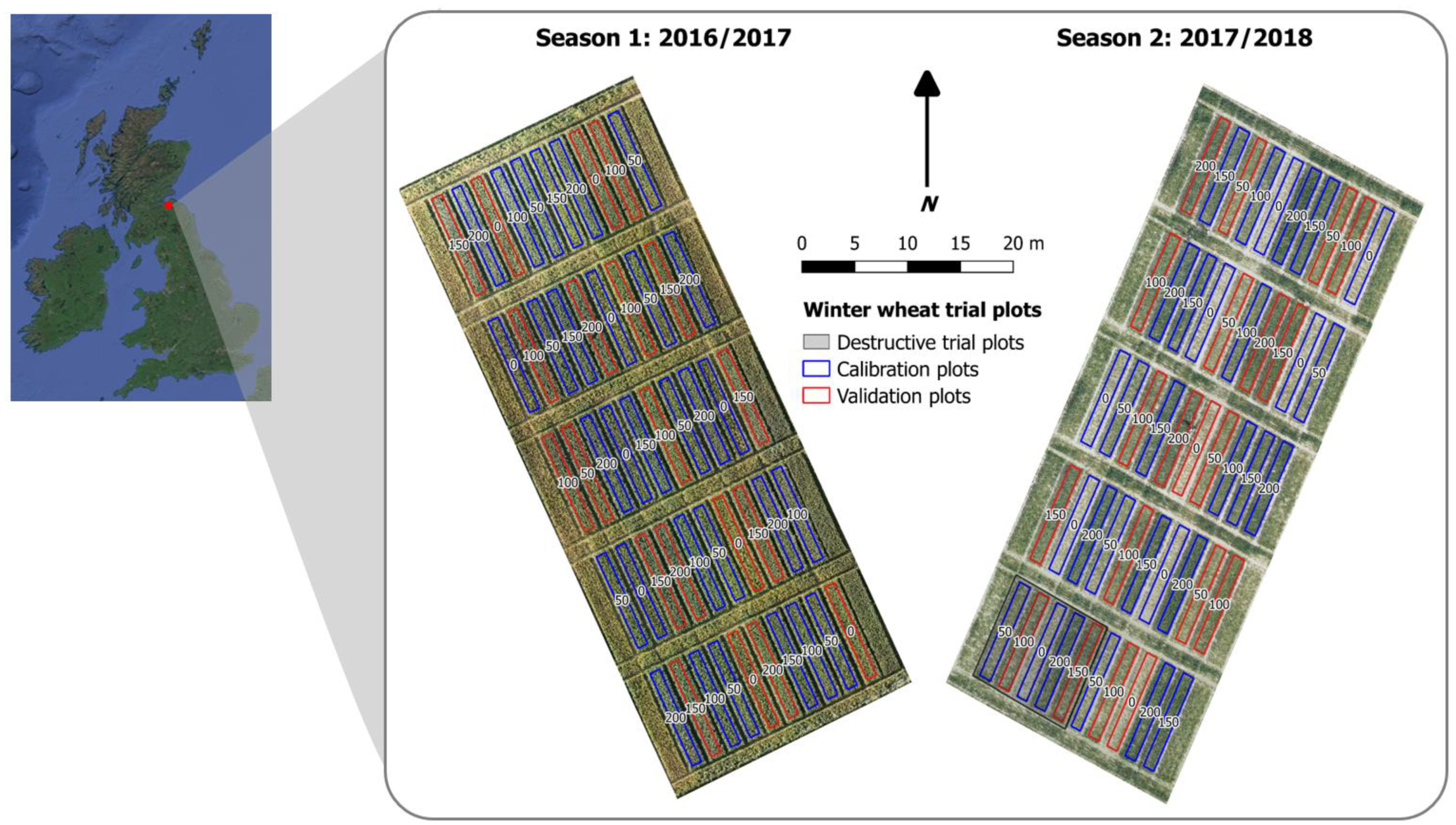
2.1.1. Experimental Trial Plot Description
2.1.2. Non-Destructive Measurements
2.1.3. Destructive Sampling and Post-Processing
2.2. Description of DALEC-Crop
2.2.1. Model Overview
2.2.2. Nitrogen Dilution Function
2.2.3. Model-Data Fusion Approach
2.3. Calibration and Experimental Protocol
2.3.1. Multi-Treatment Analysis and Calibration
2.3.2. Experiment 1: Leaf N Retrieval
2.3.3. Experiment 2: Linking Model Inputs to Yield Estimation Accuracy
2.3.4. Experiment 3: Synthetic EO Data Constraint
3. Results
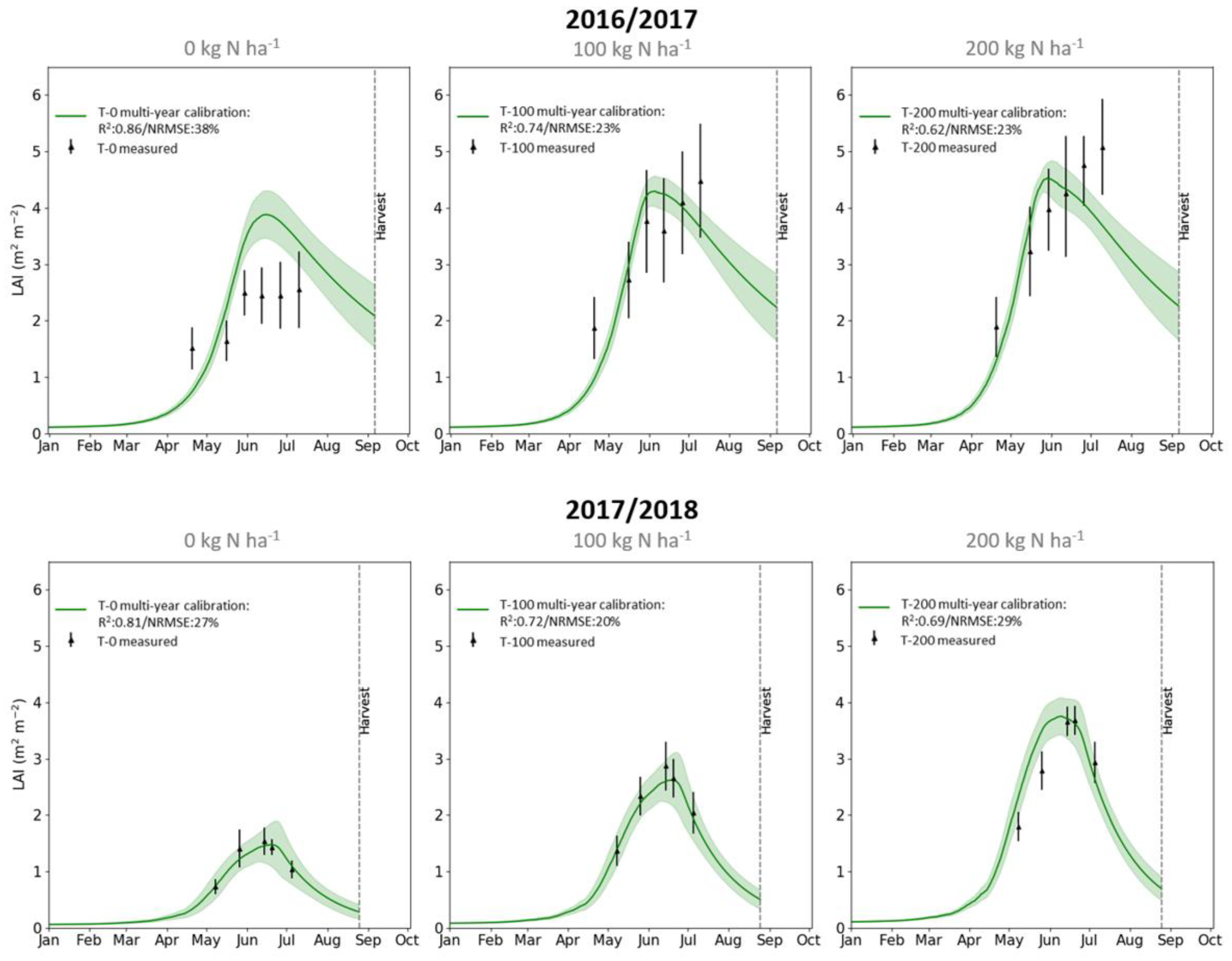
3.1. DALEC-Crop Analysis and Calibration
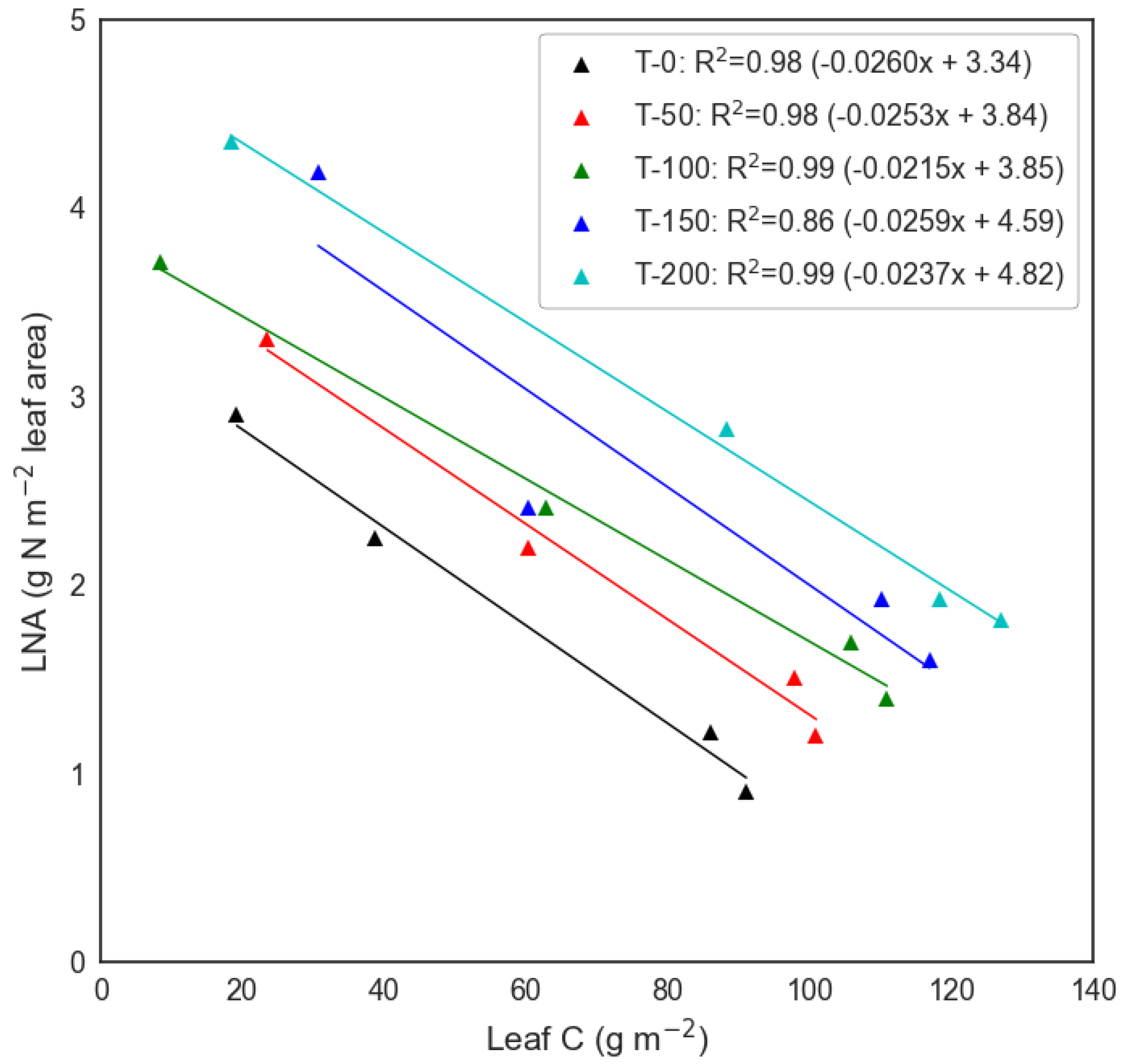
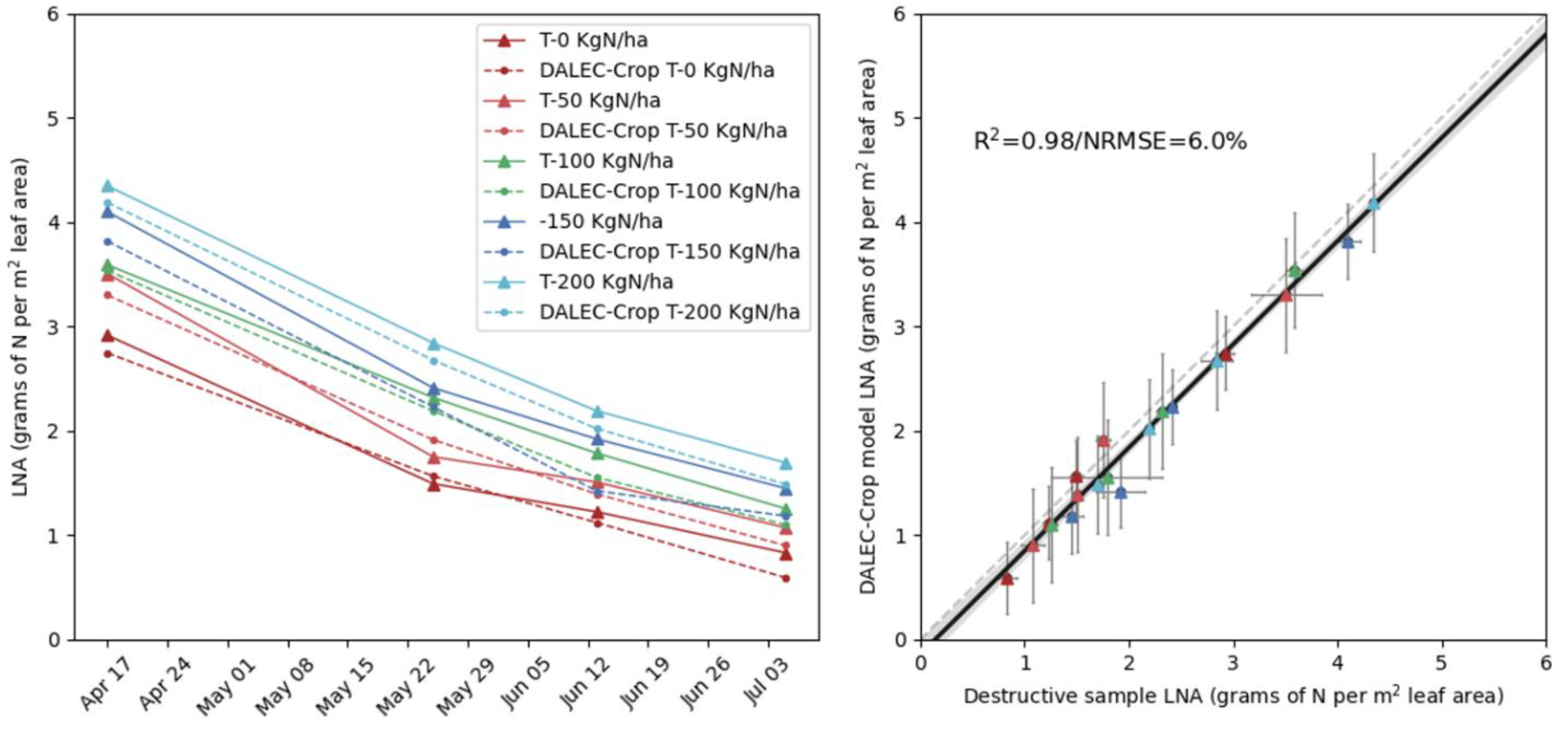
3.2. Multi-Temporal LNA Retrieval across Fertilisation Treatments
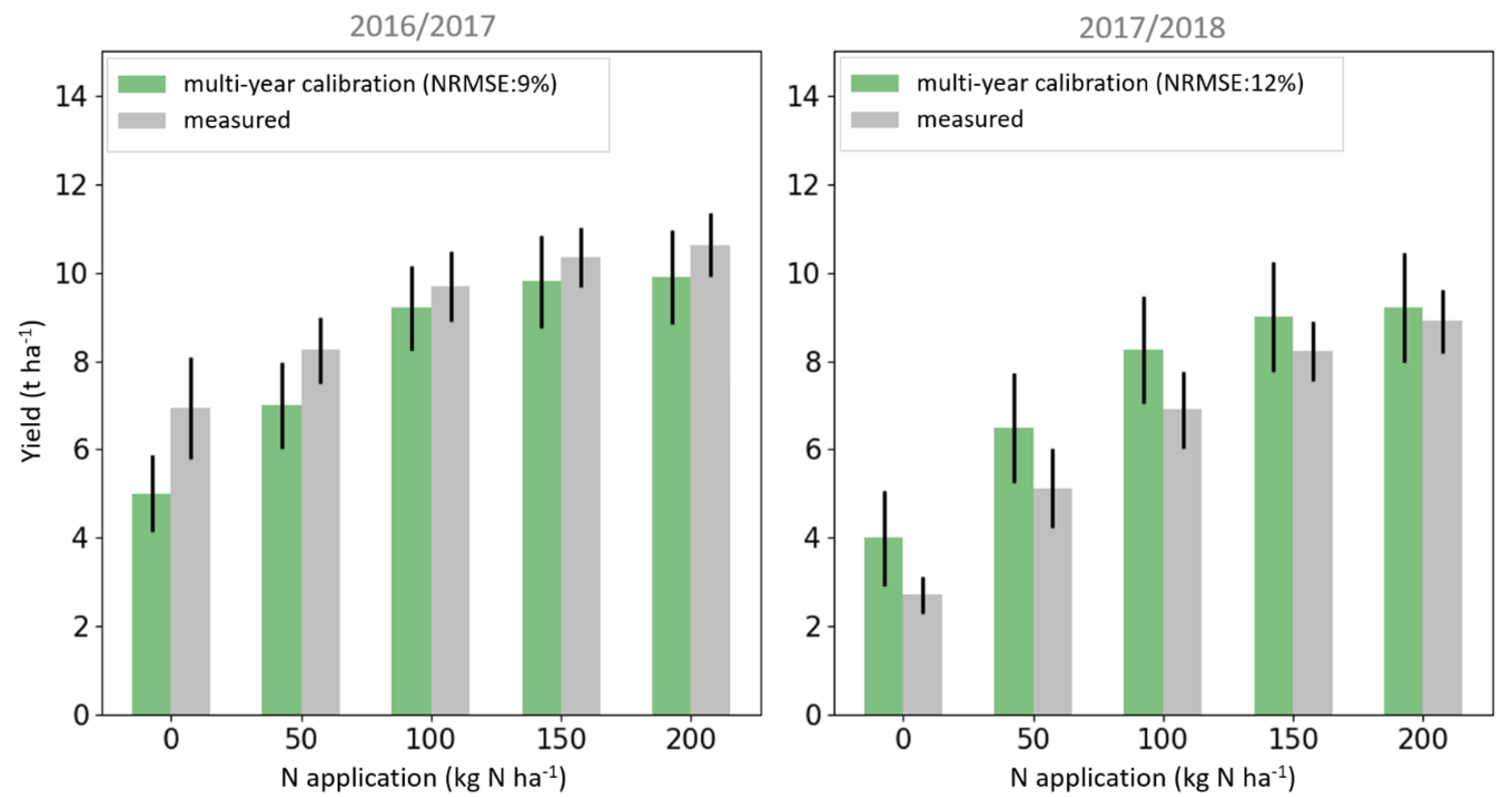
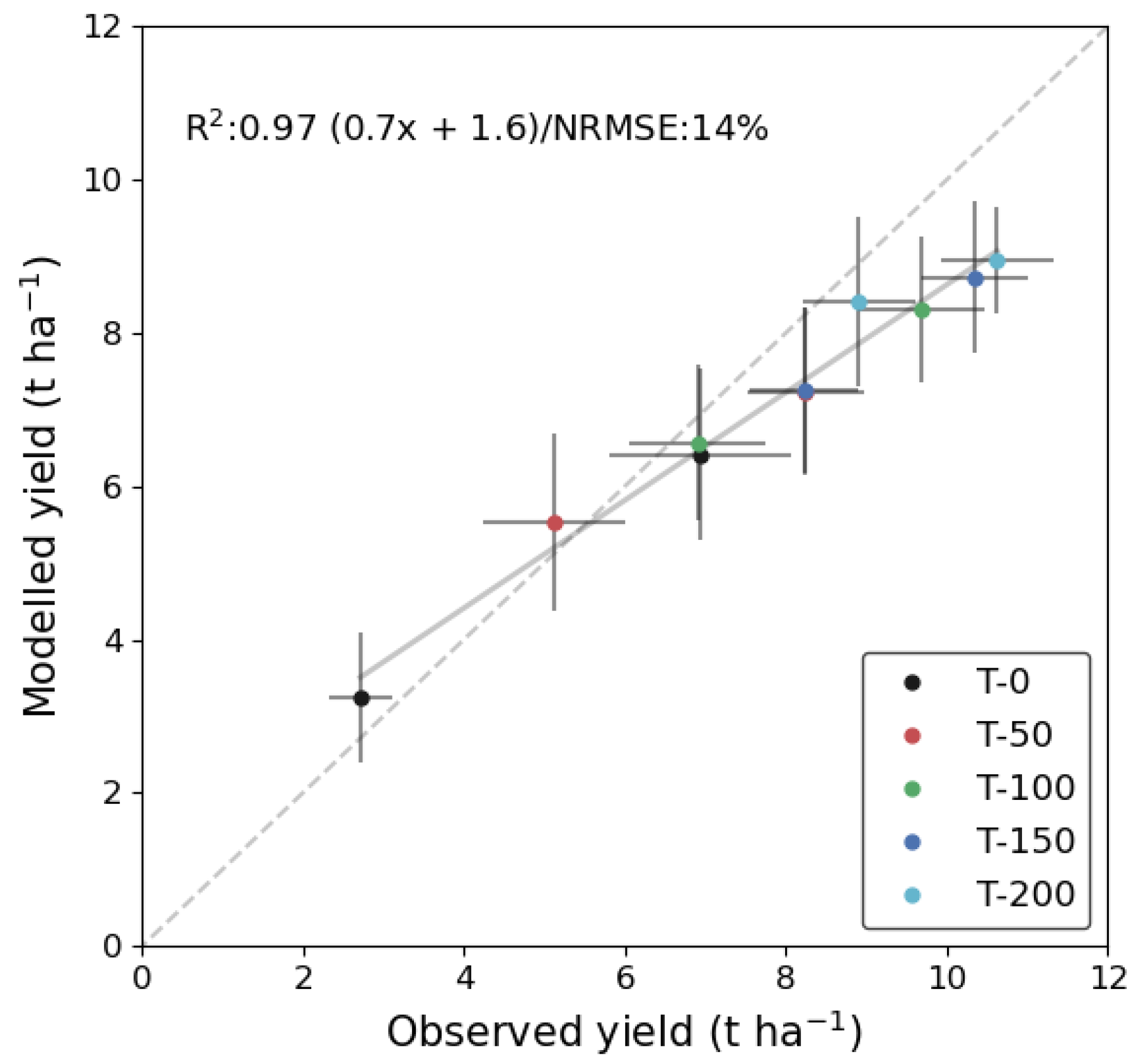
3.3. Validating Yield Retrievals Based on Leaf N Estimates Derived from LAI
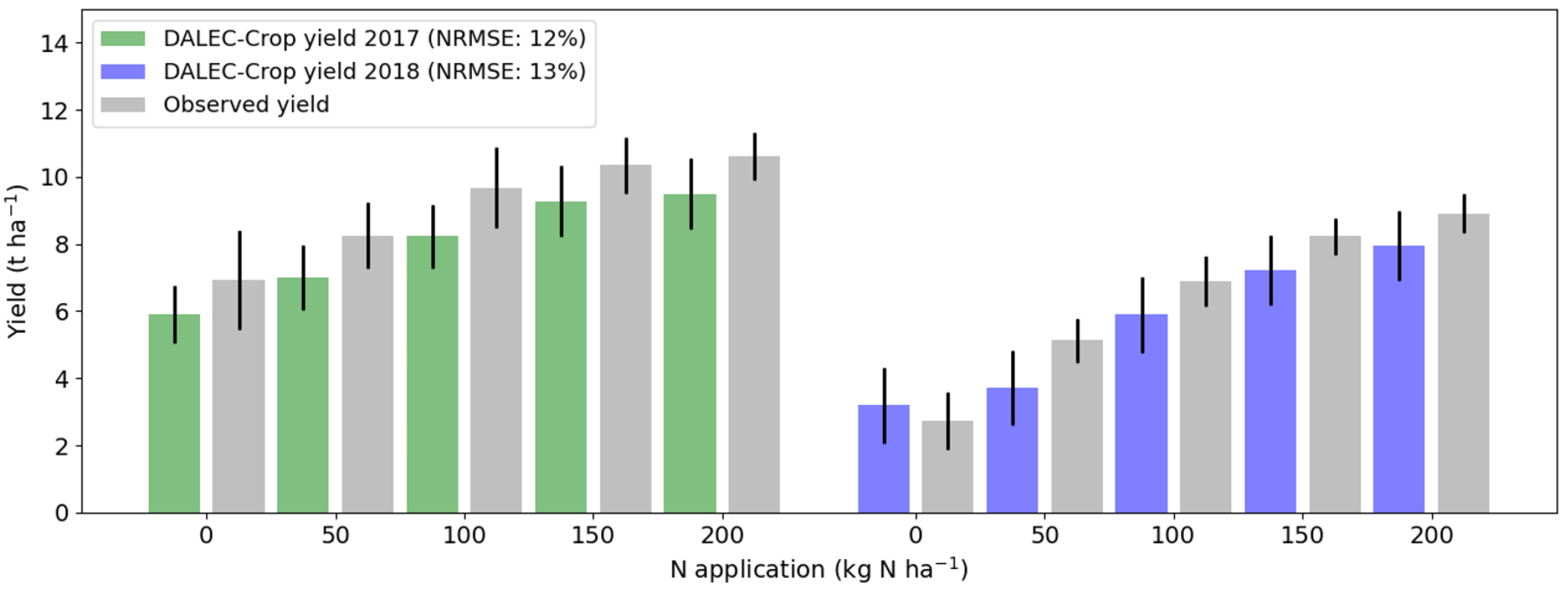
3.4. Estimating Yield Variability from LAI and Meteorology Data through Inference of Crop N Status
3.5. Yield Estimation Accuracy during the N Application Growth Stages
4. Discussion
4.1. Simulating LAI and Yield Responses to N
4.2. Leaf Nitrogen Dynamics—Constrained Estimates from LAI Observations
4.3. Impact of Model Inputs on Yield
4.4. Expected Yield Prediction Accuracy during the Nitrogen Application Period
4.5. Research Implications and Recommendations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, B.; Ata-Ul-Karim, S.T.; Duan, A.; Liu, Z.; Wang, X.; Xiao, J.; Liu, Z.; Qin, A.; Ning, D.; Zhang, W.; et al. Determination of critical nitrogen concentration and dilution curve based on leaf area index for summer maize. Field Crop. Res. 2018, 228, 195–203. [Google Scholar] [CrossRef]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [Green Version]
- Gerten, D.; Heck, V.; Jägermeyr, J.; Bodirsky, B.; Fetzer, I.; Jalava, M.; Kummu, M.; Lucht, W.; Rockström, J.; Schaphoff, S.; et al. Feeding ten billion people is possible within four terrestrial planetary boundaries. Nat. Sustain. 2020, 3, 200–208. [Google Scholar] [CrossRef]
- Lemaire, G.; Jeuffroy, M.-H.; Gastal, F. Diagnosis tool for plant and crop N status in vegetative stage. Eur. J. Agron. 2008, 28, 614–624. [Google Scholar] [CrossRef]
- Dumont, B.; Basso, B.; Bodson, B.; Destain, J.-P.; Destain, M.-F. Climatic risk assessment to improve nitrogen fertilisation recommendations: A strategic crop model-based approach. Eur. J. Agron. 2015, 65, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Rockström, J.; Steffen, W.; Noone, K.; Persson, Å.; Chapin, F.S.; Lambin, E.F.; Lenton, T.M.; Scheffer, M.; Folke, C.; Schellnhuber, H.J.; et al. A safe operating space for humanity. Nature 2009, 461, 472–475. [Google Scholar] [CrossRef] [PubMed]
- Grace, P.R.; Robertson, G.P.; Millar, N.; Colunga-Garcia, M.; Basso, B.; Gage, S.H.; Hoben, J. The contribution of maize cropping in the Midwest USA to global warming: A regional estimate. Agric. Syst. 2011, 104, 292–296. [Google Scholar] [CrossRef]
- Olin, S.; Schurgers, G.; Lindeskog, M.; Wårlind, D.; Smith, B.; Bodin, P.; Holmér, J.; Arneth, A. Modelling the response of yields and tissue C: N to changes in atmospheric CO2 and N management in the main wheat regions of western Europe. Biogeosciences 2015, 12, 2489–2515. [Google Scholar] [CrossRef] [Green Version]
- Greenwood, D.J.; Neeteson, J.J.; Draycott, A. Quantitative relationships for the dependence of growth rate of arable crops on their nitrogen content, dry weight and aerial environment. Plant Soil 1986, 91, 281–301. [Google Scholar] [CrossRef]
- Yue, S.; Meng, Q.; Zhao, R.; Li, F.; Chen, X.; Zhang, F.; Cui, Z. Critical Nitrogen Dilution Curve for Optimizing Nitrogen Management of Winter Wheat Production in the North China Plain. Agron. J. 2012, 104, 523–529. [Google Scholar] [CrossRef]
- Revill, A.; Florence, A.; Mac Arthur, A.; Hoad, S.P.; Rees, R.; Williams, M. The Value of Sentinel-2 Spectral Bands for the Assessment of Winter Wheat Growth and Development. Remote Sens. 2019, 11, 2050. [Google Scholar] [CrossRef] [Green Version]
- Clevers, J.G.P.W.; Kooistra, L.; Brande, M.M.M.V.D. Using Sentinel-2 Data for Retrieving LAI and Leaf and Canopy Chlorophyll Content of a Potato Crop. Remote Sens. 2017, 9, 405. [Google Scholar] [CrossRef] [Green Version]
- Cammarano, D.; Fitzgerald, G.J.; Casa, R.; Basso, B. Assessing the Robustness of Vegetation Indices to Estimate Wheat N in Mediterranean Environments. Remote Sens. 2014, 6, 2827–2844. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Chen, S.; Shen, S. Assimilating remote sensing information with crop model using Ensemble Kalman Filter for improving LAI monitoring and yield estimation. Ecol. Model. 2013, 270, 30–42. [Google Scholar] [CrossRef]
- Revill, A.; Florence, A.; Mac Arthur, A.; Hoad, S.P.; Rees, R.; Williams, M. Quantifying Uncertainty and Bridging the Scaling Gap in the Retrieval of Leaf Area Index by Coupling Sentinel-2 and UAV Observations. Remote Sens. 2020, 12, 1843. [Google Scholar] [CrossRef]
- Becker-Reshef, I.; Vermote, E.; Lindeman, M.; Justice, C. A generalized regression-based model for forecasting winter wheat yields in Kansas and Ukraine using MODIS data. Remote Sens. Environ. 2010, 114, 1312–1323. [Google Scholar] [CrossRef]
- Sakamoto, T.; Gitelson, A.A.; Arkebauer, T.J. MODIS-based corn grain yield estimation model incorporating crop phenology information. Remote Sens. Environ. 2013, 131, 215–231. [Google Scholar] [CrossRef]
- Lashkari, A.; Salehnia, N.; Asadi, S.; Paymard, P.; Zare, H.; Bannayan, M. Evaluation of different gridded rainfall datasets for rainfed wheat yield prediction in an arid environment. Int. J. Biometeorol. 2018, 62, 1543–1556. [Google Scholar] [CrossRef] [PubMed]
- Boote, K.J.; Jones, J.W.; White, J.W.; Asseng, S.; Lizaso, J.I. Putting mechanisms into crop production models. Plant Cell Environ. 2013, 36, 1658–1672. [Google Scholar] [CrossRef]
- Basso, B.; Ritchie, J.T.; Cammarano, D.; Sartori, L. A strategic and tactical management approach to select optimal N fertilizer rates for wheat in a spatially variable field. Eur. J. Agron. 2011, 35, 215–222. [Google Scholar] [CrossRef]
- Ewert, F.; Van Ittersum, M.K.; Heckelei, T.; Therond, O.; Bezlepkina, I.; Andersen, E. Scale changes and model linking methods for integrated assessment of agri-environmental systems. Agric. Ecosyst. Environ. 2011, 142, 6–17. [Google Scholar] [CrossRef]
- Phelan, D.C.; Harrison, M.T.; McLean, G.; Cox, H.; Pembleton, K.G.; Dean, G.J.; Parsons, D.; Richter, M.E.D.A.; Pengilley, G.; Hinton, S.J.; et al. Advancing a farmer decision support tool for agronomic decisions on rainfed and irrigated wheat cropping in Tasmania. Agric. Syst. 2018, 167, 113–124. [Google Scholar] [CrossRef]
- Asseng, S.; Ewert, F.; Rosenzweig, C.; Jones, J.W.; Hatfield, J.L.; Ruane, A.C.; Boote, K.J.; Thorburn, P.J.; Rotter, R.; Cammarano, D.; et al. Uncertainty in simulating wheat yields under climate change. Nat. Clim. Chang. 2013, 3, 827–832. [Google Scholar] [CrossRef] [Green Version]
- Sus, O.; Williams, M.; Bernhofer, C.; Béziat, P.; Buchmann, N.; Ceschia, E.; Doherty, R.; Eugster, W.; Grünwald, T.; Kutsch, W.; et al. A linked carbon cycle and crop developmental model: Description and evaluation against measurements of carbon fluxes and carbon stocks at several European agricultural sites. Agric. Ecosyst. Environ. 2010, 139, 402–418. [Google Scholar] [CrossRef]
- Di Paola, A.; Valentini, R.; Santini, M. An overview of available crop growth and yield models for studies and assessments in agriculture. J. Sci. Food Agric. 2016, 96, 709–714. [Google Scholar] [CrossRef]
- De Wit, A.; Boogaard, H.; Fumagalli, D.; Janssen, S.; Knapen, R.; Van Kraalingen, D.; Supit, I.; Van Der Wijngaart, R.; Van Diepen, K. 25 years of the WOFOST cropping systems model. Agric. Syst. 2019, 168, 154–167. [Google Scholar] [CrossRef]
- Keenan, T.F.; Davidson, E.; Moffat, A.M.; Munger, W.; Richardson, A.D. Using model-data fusion to interpret past trends, and quantify uncertainties in future projections, of terrestrial ecosystem carbon cycling. Glob. Chang. Biol. 2012, 18, 2555–2569. [Google Scholar] [CrossRef] [Green Version]
- Ziehn, T.; Scholze, M.; Knorr, W. On the capability of Monte Carlo and adjoint inversion techniques to derive posterior parameter uncertainties in terrestrial ecosystem models. Glob. Biogeochem. Cycles 2012, 26, 3025. [Google Scholar] [CrossRef] [Green Version]
- Fox, A.; Williams, M.; Richardson, A.D.; Cameron, D.; Gove, J.H.; Quaife, T.; Ricciuto, D.; Reichstein, M.; Tomelleri, E.; Trudinger, C.M.; et al. The REFLEX project: Comparing different algorithms and implementations for the inversion of a terrestrial ecosystem model against eddy covariance data. Agric. For. Meteorol. 2009, 149, 1597–1615. [Google Scholar] [CrossRef] [Green Version]
- Kuppel, S.; Peylin, P.; Chevallier, F.; Bacour, C.; Maignan, F.; Richardson, A.D. Constraining a global ecosystem model with multi-site eddy-covariance data. Biogeosci. Discuss. 2012, 9, 3317–3380. [Google Scholar] [CrossRef] [Green Version]
- Salo, T.J.; Palosuo, T.; Kersebaum, K.C.; Nendel, C.; Angulo, C.; Ewert, F.; Bindi, M.; Calanca, P.; Klein, T.W.; Moriondo, M.; et al. Comparing the performance of 11 crop simulation models in predicting yield response to nitrogen fertilization. J. Agric. Sci. 2016, 154, 1218–1240. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Tian, L.; Liang, S.; Ma, H.; Becker-Reshef, I.; Huang, Y.; Su, W.; Zhang, X.; Zhu, D.; Wu, W. Improving winter wheat yield estimation by assimilation of the leaf area index from Landsat TM and MODIS data into the WOFOST model. Agric. For. Meteorol. 2015, 204, 106–121. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.; Huang, J.; Wu, W.; Fan, J.; Zou, J.; Wu, S. Assimilation of MODIS-LAI into the WOFOST model for forecasting regional winter wheat yield. Math. Comput. Model. 2013, 58, 634–643. [Google Scholar] [CrossRef]
- Novelli, F.; Spiegel, A.; Sandén, T.; Vuolo, F. Assimilation of Sentinel-2 Leaf Area Index Data into a Physically-Based Crop Growth Model for Yield Estimation. Agronomy 2019, 9, 255. [Google Scholar] [CrossRef] [Green Version]
- Berrisford, P.; Dee, D.P.; Fielding, K.; Fuentes, M.; Kållberg, P.; Kobayashi, S.; Uppala, S.M. The ERA-Interim Archive; ECMWF: Reading, UK, 2009. [Google Scholar]
- Robinson, E.L.; Blyth, E.; Clark, D.B.; Comyn-Platt, E.; Finch, J.; Rudd, A.C. Climate Hydrology and Ecology Research Support System Meteorology Dataset for Great Britain (1961–2015), v1.2.; NERC Environmental Information Data Centre: Oxford, UK, 2016. [Google Scholar] [CrossRef]
- AHDB Recommended Lists for Cereals and Oilseeds 2017/18. 2017. Available online: https://www.yumpu.com/en/document/view/57001707/ahdb-recommended-lists-for-cereals-and-oilseeds-2017-18 (accessed on 25 November 2019).
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Penning de Vries, F.W.T.; Jansen, D.M.; ten Berge, H.F.M.; Bakema, A.H. Simulation of ecophysiological processes in several annual crops. In Simulation Monographs; Pudoc: Wageningen, The Netherlands, 1989; Volume 29. [Google Scholar]
- Wang, E.; Engel, T. Simulation of phenological development of wheat crops. Agric. Syst. 1998, 58, 1–24. [Google Scholar] [CrossRef]
- Williams, M.; Rastetter, E.B.; Fernandes, D.N.; Goulden, M.L.; Shaver, G.R.; Johnson, L.C. Predicting Gross Primary Productivity in Terrestrial Ecosystems. Ecol. Appl. 1997, 7, 882–894. [Google Scholar] [CrossRef]
- Revill, A.; Bloom, A.A.; Williams, M. Impacts of reduced model complexity and driver resolution on cropland ecosystem photosynthesis estimates. Field Crop. Res. 2016, 187, 74–86. [Google Scholar] [CrossRef] [Green Version]
- Bloom, A.A.; Williams, M. Constraining ecosystem carbon dynamics in a data-limited world: Integrating ecological “common sense” in a model–data fusion framework. Biogeosciences 2015, 12, 1299–1315. [Google Scholar] [CrossRef] [Green Version]
- Patenaude, G.; Milne, R.; Van Oijen, M.; Rowland, C.; Hill, R.A. Integrating remote sensing datasets into ecological modelling: A Bayesian approach. Int. J. Remote Sens. 2008, 29, 1295–1315. [Google Scholar] [CrossRef] [Green Version]
- Myrgiotis, V.; Blei, E.; Clement, R.; Jones, S.K.; Keane, B.; Lee, M.A.; Levy, P.E.; Rees, R.M.; Skiba, U.M.; Smallman, T.L.; et al. A model-data fusion approach to analyse carbon dynamics in managed grasslands. Agric. Syst. 2020, 184, 102907. [Google Scholar] [CrossRef]
- AHDB. Winter Wheat Growth Guide; AHDB: Kenilworth, UK, 2018. [Google Scholar]
- Caldararu, S.; Purves, D.W.; Smith, M.J. The impacts of data constraints on the predictive performance of a general process-based crop model (PeakN-crop v1.0). Geosci. Model Dev. 2017, 10, 1679–1701. [Google Scholar] [CrossRef]
- Hansen, J.; Challinor, A.; Ines, A.; Wheeler, T.; Moron, V. Translating climate forecasts into agricultural terms: Advances and challenges. Clim. Res. 2006, 33, 27–41. [Google Scholar] [CrossRef]
- Huang, J.; Gómez-Dans, J.L.; Huang, H.; Ma, H.; Wu, Q.; Lewis, P.E.; Liang, S.; Chen, Z.; Xue, J.-H.; Wu, Y.; et al. Assimilation of remote sensing into crop growth models: Current status and perspectives. Agric. For. Meteorol. 2019, 107609. [Google Scholar] [CrossRef]
- Parkes, B.; Higginbottom, T.P.; Hufkens, K.; Ceballos, F.; Kramer, B.; Foster, T. Weather dataset choice introduces uncertainty to estimates of crop yield responses to climate variability and change. Environ. Res. Lett. 2019, 14, 124089. [Google Scholar] [CrossRef]
- Revill, A.; Sus, O.; Barrett, B.; Williams, M. Carbon cycling of European croplands: A framework for the assimilation of optical and microwave Earth observation data. Remote Sens. Environ. 2013, 137, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Delloye, C.; Weiss, M.; Defourny, P. Retrieval of the canopy chlorophyll content from Sentinel-2 spectral bands to estimate nitrogen uptake in intensive winter wheat cropping systems. Remote Sens. Environ. 2018, 216, 245–261. [Google Scholar] [CrossRef]
- Söderström, M.; Piikki, K.; Stenberg, M.; Stadig, H.; Martinsson, J. Producing nitrogen (N) uptake maps in winter wheat by combining proximal crop measurements with Sentinel-2 and DMC satellite images in a decision support system for farmers. Acta Agric. Scand. Sect. B Plant Soil Sci. 2017, 67, 637–650. [Google Scholar] [CrossRef]
- Whitcraft, A.K.; Becker-Reshef, I.; Justice, C.O. A Framework for Defining Spatially Explicit Earth Observation Requirements for a Global Agricultural Monitoring Initiative (GEOGLAM). Remote Sens. 2015, 7, 1461–1481. [Google Scholar] [CrossRef] [Green Version]
- Kindred, D.R.; Milne, A.; Webster, R.; Marchant, B.; Sylvester-Bradley, R. Exploring the spatial variation in the fertilizer-nitrogen requirement of wheat within fields. J. Agric. Sci. 2015, 153, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Pallardy, S.G. Chapter 5—Photosynthesis. In Physiology of Woody Plants; Third, E., Ed.; Academic Press: San Diego, CA, USA, 2008; pp. 107–167. ISBN 978-0-12-088765-1. [Google Scholar]
- Spinoni, J.; Vogt, J.V.; Naumann, G.; Barbosa, P.; Dosio, A. Will drought events become more frequent and severe in Europe? Int. J. Clim. 2018, 38, 1718–1736. [Google Scholar] [CrossRef] [Green Version]
- Smallman, T.L.; Williams, M. Description and validation of an intermediate complexity model for ecosystem photosynthesis and evapotranspiration: ACM-GPP-ETv1. Geosci. Model Dev. 2019, 12, 2227–2253. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ground Measurement Date | Growth Stage Description |
|---|---|
| 20 April 2017 | Stem elongation—early (GS31) |
| 16 May 2017 | Stem elongation—early (GS32) |
| 30 May 2017 | Ear emergence (GS50) |
| 12 June 2017 | Ear emergence (GS59) |
| 26 June 2017 | Flowering (GS71) |
| 10 July 2017 | Milk development (GS78) |
| 17 April 2018 * | Tillering—late (GS30) |
| 08 May 2018 | Stem elongation—early (GS31) |
| 25 May 2018 * | Stem elongation—late (GS38) |
| 05 June 2018 | Ear emergence (GS54) |
| 20 June 2018 * | Flowering (GS68) |
| 04 July 2018 * | Milk development (GS79) |
| Index | EDC Description |
|---|---|
| 1 | C Pools cannot be negative |
| 2 | C Fluxes cannot be negative |
| 3 | LAI must be less than 6.5 m2 m−2 |
| 4 | GPP must be less than 25 g C m−2 |
| Multi-Year Calibration | ||||
|---|---|---|---|---|
| 2016–2017 | 2017–2018 | |||
| N Application (kg N ha−1) | R2 | NRMSE (%) | R2 | NRMSE (%) |
| 0 | 0.86 | 38 | 0.81 | 27 |
| 50 | 0.8 | 23 | 0.99 | 24 |
| 100 | 0.74 | 23 | 0.72 | 20 |
| 150 | 0.65 | 29 | 0.96 | 23 |
| 200 | 0.62 | 23 | 0.69 | 29 |
| Mean | 0.73 | 27 | 0.83 | 25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Revill, A.; Myrgiotis, V.; Florence, A.; Hoad, S.; Rees, R.; MacArthur, A.; Williams, M. Combining Process Modelling and LAI Observations to Diagnose Winter Wheat Nitrogen Status and Forecast Yield. Agronomy 2021, 11, 314. https://doi.org/10.3390/agronomy11020314
Revill A, Myrgiotis V, Florence A, Hoad S, Rees R, MacArthur A, Williams M. Combining Process Modelling and LAI Observations to Diagnose Winter Wheat Nitrogen Status and Forecast Yield. Agronomy. 2021; 11(2):314. https://doi.org/10.3390/agronomy11020314
Chicago/Turabian StyleRevill, Andrew, Vasileios Myrgiotis, Anna Florence, Stephen Hoad, Robert Rees, Alasdair MacArthur, and Mathew Williams. 2021. "Combining Process Modelling and LAI Observations to Diagnose Winter Wheat Nitrogen Status and Forecast Yield" Agronomy 11, no. 2: 314. https://doi.org/10.3390/agronomy11020314
APA StyleRevill, A., Myrgiotis, V., Florence, A., Hoad, S., Rees, R., MacArthur, A., & Williams, M. (2021). Combining Process Modelling and LAI Observations to Diagnose Winter Wheat Nitrogen Status and Forecast Yield. Agronomy, 11(2), 314. https://doi.org/10.3390/agronomy11020314