
An Index for User-Friendly Proximal Detection of Water Requirements to Optimized Irrigation Management in Vineyards

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sites
2.2. Thermal Time for Phenological Succession
2.3. Vine Water Relations and Physiological Performances
2.4. Vineyard Water Balance, Atmospheric Evaporative Demand
2.5. Canopy Thermal Regime
2.6. Index for Optimized Irrigation Management
2.7. Statistical Analysis
3. Results and Discussion
3.1. Vineyard Thermal Regime, Water Balance and Productivity
3.2. Accuracy and Representativeness of Midday Stem Water Potential for Vine Water Status and Irrigation Opportunity Evaluation
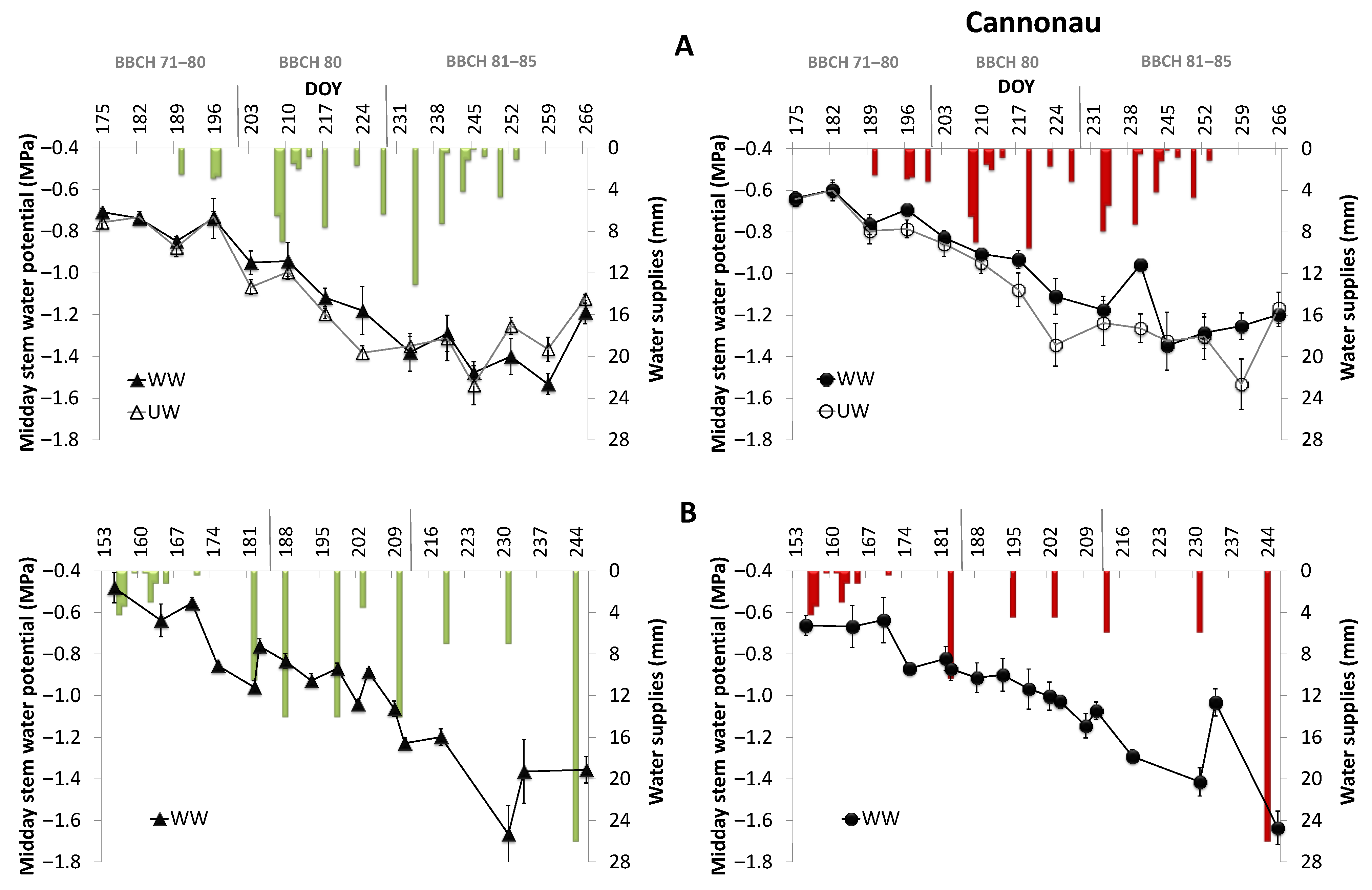
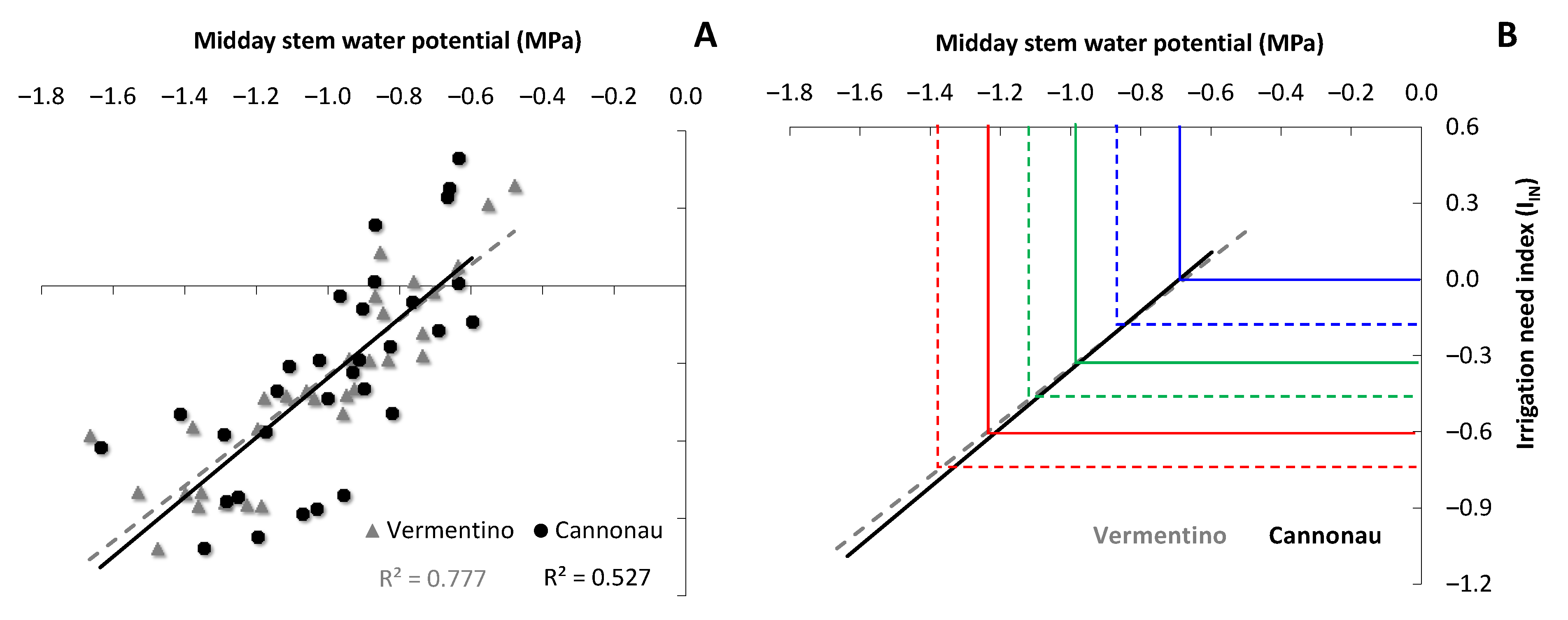
3.3. Representativeness of the Midday Stem Water Potential for Vermentino and Cannonau Varieties and Validation of Variety-Specific Thresholds
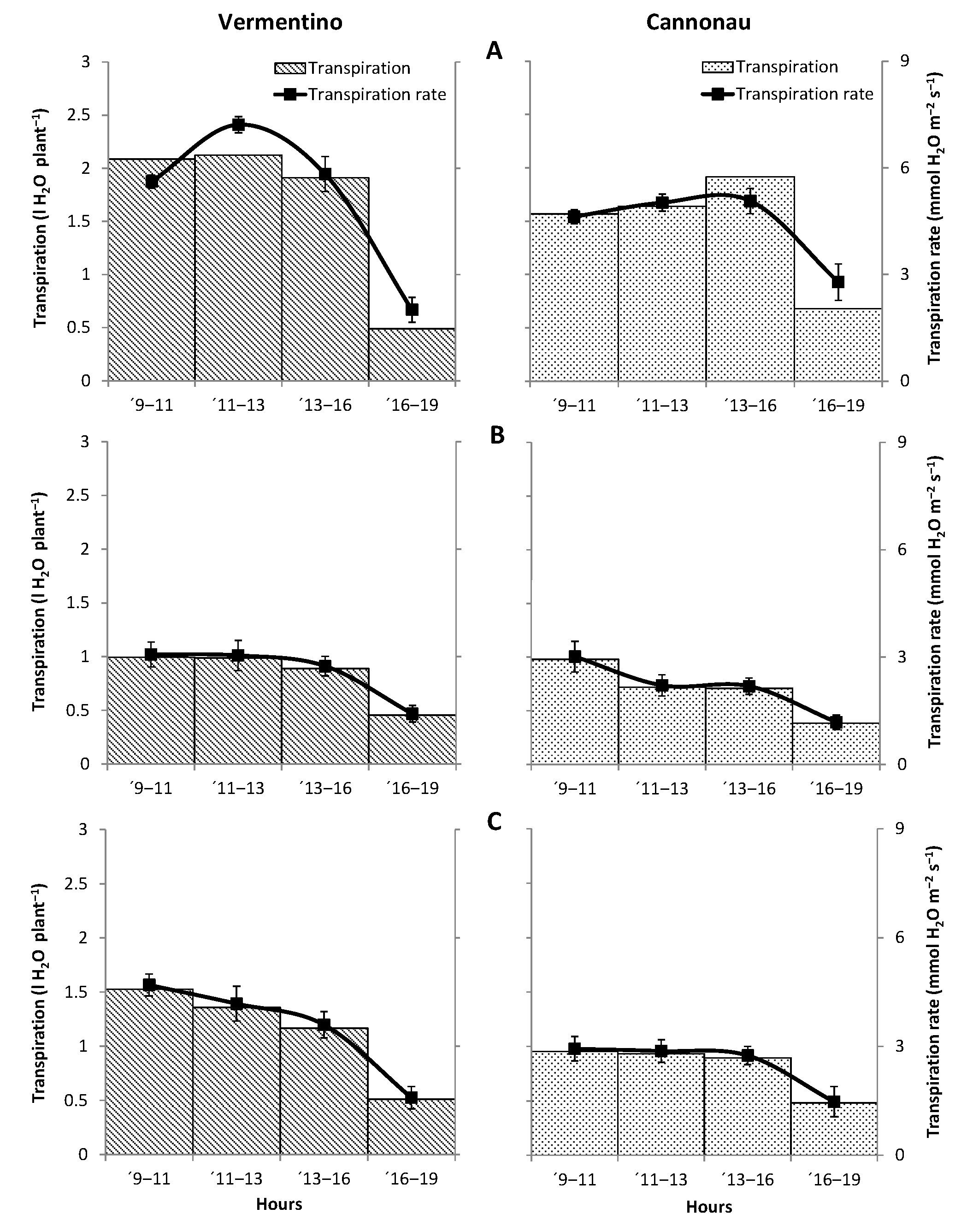
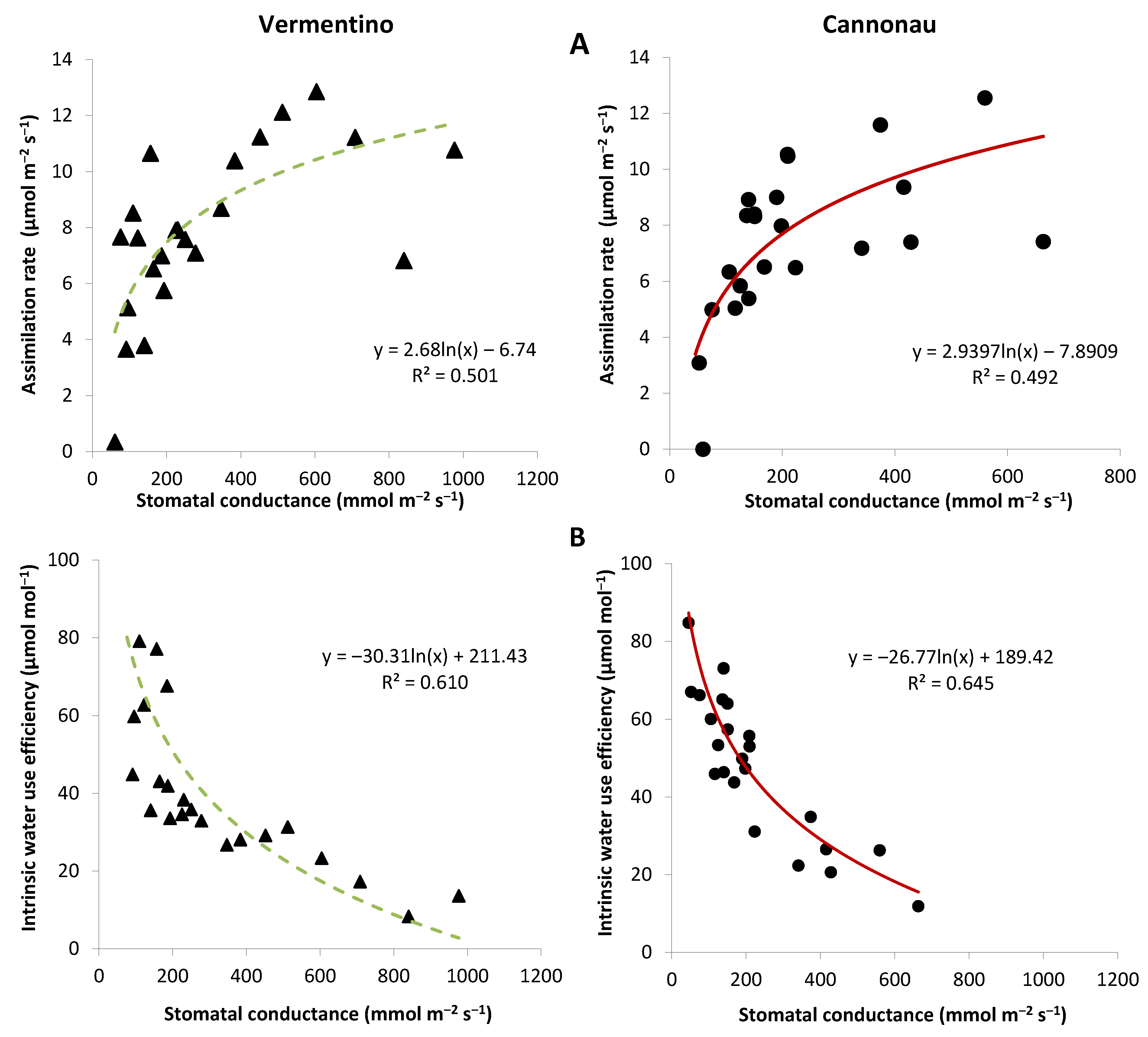
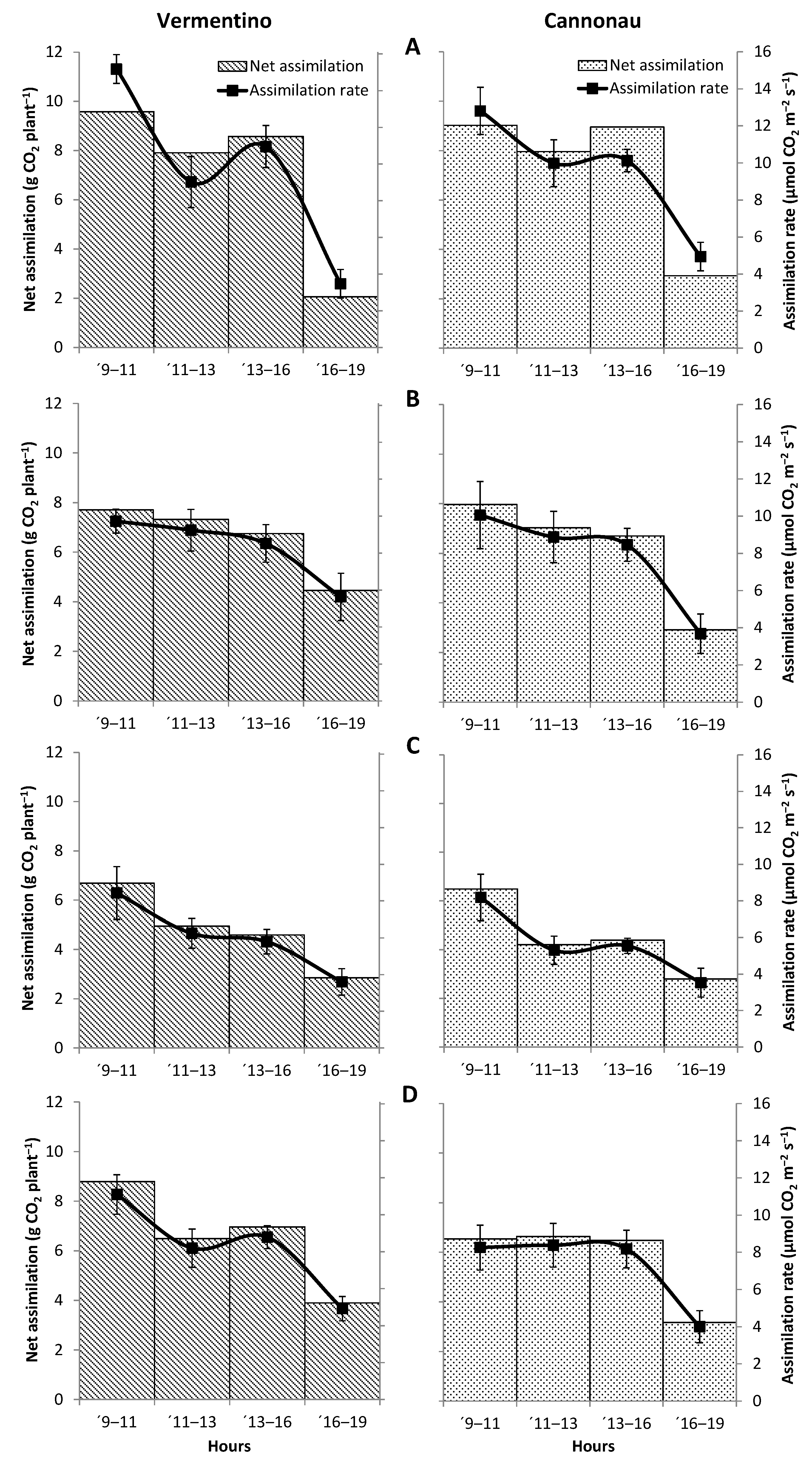
3.4. Evaluation of Photosynthetic and Transpiration Performances of Vermentino and Cannonau Grapevines in Three Different Viticultural Contexts
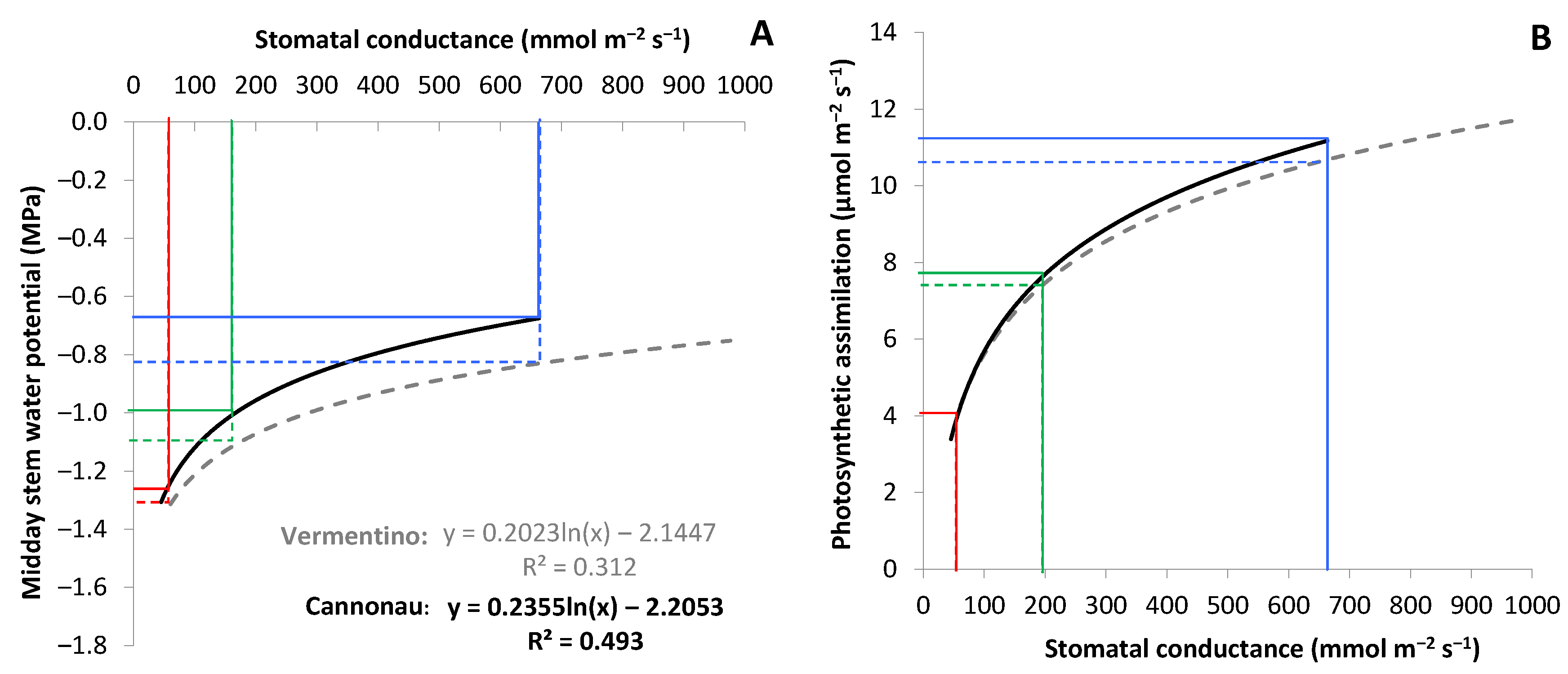
3.5. Validation and Accuracy of the Index for Irrigation Automation in Vermentino and Cannonau Vineyards
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | CV | Texture | ρ (g cm−3) | pH | O.M. (%) | CaCO3 (%) | CEC (cmol kg−1) | Salinity (dS m−1) | FC (g 100 g−1) | WP (g 100 g−1) | AW (mm m−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Romangia | VMT | Clay loam | 1.2 | 8.6 | 1.1 | 42 | 78.5 | 0.16 | 34.28 | 16.54 | 213.29 |
| CNN | Clay loam | 1.21 | 8.6 | 1.6 | 40 | 16.1 | 0.05 | 33.81 | 16.28 | 211.49 | |
| Gallura | VMT | Sandy loam | 1.47 | 6.8 | 3.9 | - | 27.4 | 0.12 | 12.55 | 5.55 | 102.94 |
| CNN | Sandy loam | 1.47 | 6.1 | 1.8 | - | 16.1 | 0.05 | 12.35 | 5.46 | 101.6 | |
| Parteolla | VMT | Clay loam | 1.3 | 8.4 | 1.3 | 17.2 | 23.7 | 0.09 | 26.34 | 12.57 | 179.54 |
| CNN | Clay loam | 1.3 | 8.5 | 1.4 | 26.3 | 26.1 | 0.11 | 26.78 | 12.76 | 181.73 |
References
- Schultz, H.R. Issues to be considered for strategic adaptation to climate evolution—Is atmospheric evaporative demand changing? OENO One 2017, 51, 107–114. [Google Scholar] [CrossRef]
- Mosedale, J.R.; Wilson, R.J.; Maclean, I.M.D. Climate change and crop exposure to adverse weather: Changes to frost risk and grapevine flowering conditions. PLoS ONE 2015, 10, e0141218. [Google Scholar] [CrossRef] [Green Version]
- Nicholas, K.A. Will we still enjoy Pinot noir? Sci. Am. 2015, 312, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.G.; Grant, O. Remote sensing and other imaging technologies to monitor grapevine performance. In Grapevine under Environmental Stress: From Ecophysiology to Molecular Mechanisms; Gerós, H., Chaves, M.M., Medrano, H., Delrot, S., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2015; pp. 179–201. [Google Scholar] [CrossRef]
- Douthe, C.; Medrano, H.; Tortosa, I.; Escalona, J.M.; Hernández-Montes, E.; Pou, A. Whole-plant water use in field grown grapevine: Seasonal and environmental effects on water and carbon balance. Front. Plant Sci. 2018, 1540. [Google Scholar] [CrossRef]
- Medrano, H.; Tomás, M.; Martorell, S.; Escalona, J.M.; Pou, A.; Fuentes, S.; Flexas, J.; Bota, J. Improving water use efficiency of vineyards in semi-arid regions. A review. Agron. Sustain. Dev. 2015, 35, 499–517. [Google Scholar] [CrossRef] [Green Version]
- Mozell, M.R.; Thach, L. The impact of climate change on the global wine industry: Challenges & solutions. J. Wine Econ. 2014, 3, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.M.; Grant, O.M.; Chaves, M.M. Thermography to explore plant–environment interactions. J. Exp. Bot. 2013, 64, 3937–3949. [Google Scholar] [CrossRef]
- Pou, A.; Diago, M.P.; Medrano, H.; Baluja, J.; Tardaguila, J. Validation of thermal indices for water status identification in grapevine. Agric. Water Manag. 2014, 134, 60–72. [Google Scholar] [CrossRef]
- Zarrouk, O.; Costa, M.; Francisco, R.; Lopes, C.; Chaves, M.M. Drought and water management in Mediterranean vineyards. In Grapevine under Environmental Stress: From Ecophysiology to Molecular Mechanisms; Gerós, H., Chaves, M.M., Medrano, H., Delrot, S., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2015; pp. 38–67. [Google Scholar] [CrossRef]
- Matese, A.; Baraldi, R.; Berton, A.; Cesaraccio, C.; Di Gennaro, S.; & Duce, P.; Facini, O.; Mameli, M.; Piga, A.; Zaldei, A. Estimation of water stress in grapevines using proximal and remote sensing methods. Remote Sens. 2018, 10, 114. [Google Scholar] [CrossRef] [Green Version]
- van Leeuwen, C.; Destrac, A. Modified grape composition under climate change conditions requires adaptations in the vineyard. OENO One 2017, 51, 147–154. [Google Scholar] [CrossRef]
- Costa, J.M.; Egipto, R.; Sánchez-Virosta, A.; Lopes, C.M.; Chaves, M.M. Canopy and soil thermal patterns to support water and heat stress management in vineyards. Agric. Water Manag. 2019, 216, 484–496. [Google Scholar] [CrossRef]
- Jones, H.G. Use of infrared thermometry for estimation of stomatal conductance in irrigation scheduling. Agr. For. Meteorol. 1999, 95, 139–149. [Google Scholar] [CrossRef]
- Jones, H.; Stoll, M.; Santos, T.; Souza, C.; Chaves, M.M.; Grant, O. Use of infrared thermography for monitoring stomatal closure in the field: Application to grapevine. J. Exp. Bot. 2002, 378, 2249–2260. [Google Scholar] [CrossRef]
- Loveys, B.R.; Jones, H.G.; Theobald, J.C.; McCarthy, M.-G. An assessment of plant-based measures of grapevine performance as irrigation scheduling tools. Acta Hortic. 2008, 792, 421–427. [Google Scholar] [CrossRef]
- Grant, O.M.; Tronina, L.; Jones, H.G.; Chaves, M.M. Exploring thermal imaging variables for the detection of stress responses in grapevine under different irrigation regimes. J. Exp. Bot. 2007, 58, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, K.; Durham, W. Farm-scale adaptation and vulnerability to environmental stresses: Insights from winegrowing in Northern California. Glob. Environ. Chang. 2012, 22, 483–494. [Google Scholar] [CrossRef]
- Costa, J.M.; Marques da Silva, J.; Pinheiro, C.; Barón, M.; Mylona, P.; Centritto, M.; Haworth, M.; Loreto, F.; Uzilday, B.; Turkan, I.; et al. Opportunities and limitations of crop phenotyping in Southern European countries. Front. Plant Sci. 2019, 10, 1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojeda, H. Irrigation qualitative de précision de la vigne. PAV 2007, 7, 133–141. [Google Scholar]
- Schultz, H.R. Differences in hydraulic architecture account for near-isohydric and anisohydric behaviour of two field-grown Vitis vinifera L. cultivars during drought. Plant Cell Environ. 2003, 26, 1393–1405. [Google Scholar] [CrossRef]
- Jones, G.V. Climate change: Observations, projections and general implications for viticulture and wine production. In Work Paper No 7, Economics Department Whitman College; Essick, E., Griffin, P., Keefer, B., Miller, S., Storchmann, K., Eds.; Whitman College: Walla Walla, DC, USA, 2007; pp. 1–15. [Google Scholar]
- Chaves, M.M.; Zarrouk, O.; Francisco, R.; Costa, M.; Santos, T.; Regalado, A.; Rodrigues, M.; Lopes, C. Grapevine under deficit irrigation: Hints from physiological and molecular data. Ann. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef] [Green Version]
- Lovisolo, C.; Lavoie-Lamoureux, A.; Tramontini, S.; Ferrandino, A. Grapevine adaptations to water stress: New perspectives about soil/plant interactions. Theor. Exp. Plant Physiol. 2016, 28, 53–66. [Google Scholar] [CrossRef]
- Duchêne, E. How can grapevine genetics contribute to the adaptation to climate change? OENO One 2016, 50, 113–124. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Global warming of 1.5 °C. In An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable development, and Efforts to Eradicate Poverty; Masson-Delmotte, V., Zhai, P., Pörtner, H.O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; World Meteorological Organization: Geneva, Switzerland, 2018; p. 616. [Google Scholar]
- Martínez de Toda, F.; Ramos, M.C. Variability in grape composition and phenology of Tempranillo variety in zones located at different elevations and with differences in the climatic conditions. Vitis 2019, 58, 131–139. [Google Scholar] [CrossRef]
- Gambetta, G.A.; Herrera, J.C.; Dayer, S.; Feng, Q.; Hochberg, U.; Castellarin, S.D. The physiology of drought stress in grapevine: Towards an integrative definition of drought tolerance. J. Exp. Bot. 2020, 71, 4658–4676. [Google Scholar] [CrossRef]
- Florez-Sarasa, I.; Clemente-Moreno, M.J.; Cifre, J.; Capó, M.; Llompart, M.; Fernie, A.R.; Bota, J. Differences in Metabolic and Physiological Responses between Local and Widespread Grapevine Cultivars under Water Deficit Stress. Agronomy 2020, 10, 1052. [Google Scholar] [CrossRef]
- Gerzon, E.; Biton, I.; Yaniv, Y.; Zemach, H.; Netzer, Y.; Schwartz, A.; Fait, A.; Ben-Ari, G. Grapevine anatomy as a possible determinant of isohydric or anisohydric behavior. Am. J. Enol. Vitic. 2015, 66, 340–347. [Google Scholar] [CrossRef]
- Tombesi, S.; Poni, S.; Palliotti, A. Stress idrico in Vitis vinifera: Variabilità delle risposte fisiologiche intraspecifiche e loro potenziale sfruttamento nella mitigazione degli effetti dei cambiamenti climatici. Italus Hortus 2016, 23, 45–53. [Google Scholar]
- Hochberg, U.; Degu, A.; Fait, A.; Rachmilevitch, S. Near isohydric grapevine cultivar displays higher photosynthetic efficiency and photorespiration rates under drought stress as compared with near anisohydric grapevine cultivar. Physiol. Plant. 2013, 147, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Poni, S.; Bernizzoni, F.; Civardi, S. Response of “Sangiovese” grapevines to partial root-zone drying: Gas-exchange, growth and grape composition. Sci. Hortic. 2007, 114, 96–103. [Google Scholar] [CrossRef]
- Soar, C.J.; Speirs, J.; Maffei, S.M.; Penrose, A.B.; Mccarthy, M.G.; Loveys, B.R. Grapevine varieties Shiraz and Grenache differ in their stomatal response to VPD: Apparent links with ABA physiology and gene expression in leaf tissue. Aust. J. Grape Wine Res. 2006, 12, 2–12. [Google Scholar] [CrossRef]
- Vandeleur, R.K.; Mayo, G.; Shelden, M.C.; Gilliham, M.; Kaiser, B.N.; Tyerman, S.D. The role of plasma membrane intrinsic protein aquaporins in water transport through roots: Diurnal and drought stress responses reveal different strategies between isohydric and anisohydric cultivars of grapevine. Plant Physiol. 2009, 149, 445–460. [Google Scholar] [CrossRef] [Green Version]
- Tombesi, S.; Nardini, A.; Frioni, T.; Soccolini, M.; Zadra, C.; Farinelli, D.; Poni, S.; Palliotti, A. Stomatal closure is induced by hydraulic signals and maintained by ABA in drought-stressed grapevine. Sci. Rep. 2015, 5, 12449. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, U.; Rockwell, F.E.; Holbrook, N.M.; Cochard, H. Iso/anisohydry: A plant-environment interaction rather than a simple hydraulic trait. Trends Plant Sci. 2018, 23, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Ollat, N.; Cookson, S.J.; Destrac-Irvine, A.; Lauvergeat, V.; Ouaked-Lecourieux, F.; Marguerit, E.; Barrieu, F.; Dai, Z.; Duchêne, E.; Gambetta, G.A.; et al. Grapevine adaptation to abiotic stress: An overview. Acta Hortic. 2019, 1248, 497–512. [Google Scholar] [CrossRef]
- Medrano, H.; Tortosa, I.; Montes, E.; Pou, A.; Balda, P.; Bota, J.; Escalona, J.M. Genetic improvement of grapevine (Vitis vinifera L.) water use efficiency. In Water Scarcity and Sustainable Agriculture in Semiarid Environment: Tools, Strategies, and Challenges for Woody Crops; Tejero, I.F.C., Zuazo, V.H.D., Eds.; Academic Press: London, UK; San Diego, CA, USA; Cambridge, UK; Kidlington, UK, 2018; pp. 377–401. [Google Scholar] [CrossRef]
- Tramontini, S.; Döring, J.; Vitali, M.; Ferrandino, A.; Stoll, M.; Lovisolo, C. Soil water-holding capacity mediates hydraulic and hormonal signals of near-isohydric and near-anisohydric Vitis cultivars in potted grapevines. Funct. Plant Biol. 2014, 41, 1119–1128. [Google Scholar] [CrossRef]
- Fernandes de Oliveira, A.; Mameli, M.G.; De Pau, L.; Satta, D.; Nieddu, G. Deficit irrigation strategies in Vitis vinifera L. cv. Cannonau under Mediterranean climate. Part I—Physiological responses, growth-yield balance and berry composition. S. Afr. J. Enol. Vitic. 2013, 34, 170–183. [Google Scholar] [CrossRef] [Green Version]
- Fernandes de Oliveira, A.; Mercenaro, L.; Azzena, M.; Nieddu, G. Effects of pre and post-veraison water deficit on Vermentino cluster microclimate and berry composition. In BIO Web of Conferences; EDP Sciences: Ulis, France, 2019; Volume 13, p. 04015. [Google Scholar] [CrossRef]
- Alsina, M.M.; Smart, D.R.; Bauerle, T.; de Herralde, F.; Biel, C.; Stockert, C.; Negron, C.; Save, R. Seasonal changes of whole root system conductance by a drought-tolerant grape root system. J. Exp. Bot. 2011, 62, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Intrigliolo, D.S.; Castel, J.R. Evaluation of grapevine water status from trunk diameter variations. Irrig. Sci. 2007, 26, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Lovisolo, C.; Perrone, I.; Carra, A.; Ferrandino, A.; Flexas, J.; Medrano, H.; Schubert, A. Drought-induced changes in development and function of grapevine (Vitis spp.) organs and in their hydraulic and non-hydraulic interactions at the whole-plant level: A physiological and molecular update. Funct. Plant. Biol. 2010, 37, 98–116. [Google Scholar] [CrossRef]
- Santos, J.A.; Fraga, H.; Malheiro, A.C.; Moutinho-Pereira, J.; Dinis, L.-T.; Correia, C.; Moriondo, M.; Leolini, L.; Dibari, C.; Costafreda-Aumedes, S.; et al. A review of the potential climate change impacts and adaptation options for European viticulture. Appl. Sci. 2020, 10, 3092. [Google Scholar] [CrossRef]
- Choné, X.; Van Leeuwen, C.; Dubourdieu, D.; Gaudillères, J.-P. Stem water potential is a sensitive indicator of grapevine water status. Ann. Bot. 2001, 87, 477–483. [Google Scholar] [CrossRef] [Green Version]
- Myburgh, P.A. Responses of Vitis vinifera L. cv. Merlot to low frequency deficit irrigation and partial root zone drying in western Cape Coast region—Part, I. Soil and plant water status. S. Afr. J. Enol. Vitic. 2011, 32, 89–103. [Google Scholar] [CrossRef] [Green Version]
- Acevedo-Opazo, C.; Ortega-Farias, S.; Fuentes, S. Effects of grapevine (Vitis vinifera L.) water status on water consumption, vegetative growth and grape quality: An irrigation scheduling application to achieve regulated deficit irrigation. Agric. Water Manag. 2010, 97, 956–964. [Google Scholar] [CrossRef]
- Cola, G.; Failla, O.; Maghradze, D.; Meghrelidze, L.; Mariani, L. Grapevine phenology and climate change in Georgia. Int. J. Biometeorol. 2017, 61, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Mariani, L.; Alilla, R.; Cola, G.; Dal Monte, G.; Epifani, C.; Puppi, G.; Failla, O. IPHEN—A real-time network for phenological monitoring and modelling in Italy. Int. J. Biometeorol. 2013, 57, 881–893. [Google Scholar] [CrossRef] [PubMed]
- Simonneau, T.; Lebon, E.; Coupel-Ledru, A.; Marguerit, E.; Rossdeutsch, L.; Ollat, N. Adapting plant material to face water stress in vineyards: Which physiological targets for an optimal control of plant water status? OENO One 2017, 51, 167–179. [Google Scholar] [CrossRef]
- Jones, H.G. Irrigation scheduling: Advantages and pitfalls of plant-based methods. J. Exp. Bot. 2004, 55, 2427–2436. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop evapotranspiration—guidelines for computing crop water requirements. In FAO Irrigation and Drainage Paper 56; Food and Agriculture Organization: Rome, Italy, 1998. [Google Scholar]
- Idso, S.B.; Jackson, R.D.; Pinter, P.J.; Reginato, R.J.; Hatfield, J.L. Normalizing the stress degree day parameter for environmental variability. Agric. Meteorol. 1981, 24, 45–55. [Google Scholar] [CrossRef]
- Maes, W.H.; Steppe, K. Estimating evapotranspiration and drought stress with ground-based thermal remote sensing in agriculture: A review. J. Exp. Bot. 2012, 63, 4671–4712. [Google Scholar] [CrossRef] [Green Version]
- Stull, R. Wet-Bulb temperature from relative humidity and air temperature. J. Appl. Meteorol. Climatol. 2011, 50, 2267–2269. [Google Scholar] [CrossRef] [Green Version]
- ARPAS. Riepilogo Mensile Meteorologico e Agrometeorologico. Aprile-Ottobre 2019. Agenzia Regionale per la Protezione Dell’ambiente Della Sardegna Dipartimento Meteoclimatico. Available online: http://www.sar.sardegna.it/pubblicazioni/riepiloghimensili/mensili.asp (accessed on 1 October 2020).
- ARPAS. Riepilogo Mensile Meteorologico e Agrometeorologico. Aprile-Ottobre 2020. Agenzia Regionale per la Protezione Dell’ambiente Della Sardegna Dipartimento Meteoclimatico. Available online: http://www.sar.sardegna.it/pubblicazioni/riepiloghimensili/mensili.asp (accessed on 1 October 2020).
- Belfiore, N.; Vinti, R.; Lovat, L.; Chitarra, W.; Tomasi, D.; de Bei, R.; Meggio, F.; Gaiotti, F. Infrared thermography to estimate vine water status: Optimizing canopy measurements and thermal indices for the varieties Merlot and Moscato in northern Italy. Agronomy 2019, 9, 821. [Google Scholar] [CrossRef] [Green Version]
- Alsina, M.M.; Herralde, F.D.; Aranda, X.; Savé, R.; Biel, C. Water relations and vulnerability to embolism are not related: Experiments with eight grapevine cultivars. Vitis 2007, 46, 1–6. [Google Scholar] [CrossRef]
- Fernandes de Oliveira, A.; Rais, F.; Dettori, I.; Azzena, M.; Nieddu, G. UV light acclimation capacity of leaf photosynthetic and photochemical behaviour in near-isohydric and anisohydric grapevines in hot and dry environments. S. Afr. J. Enol. Vitic. 2019, 40, 188–205. [Google Scholar] [CrossRef] [Green Version]
- Medrano, H.; Pou, A.; Tomás, M.; Martorell, S.; Gulias, J.; Flexas, J.; Escalona, J.M. Average daily light interception determines leaf water use efficiency among different canopy locations in grapevine. Agric. Water Manag. 2012, 114, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Medrano, H.; Magdalena, T.; Martorell, S.; Flexas, J.; Hernández, E.; Rosselló, J.; Pou, A.; Escalona, J.M.; Bota, J. From leaf to whole-plant water use efficiency (WUE) in complex canopies: Limitations of leaf WUE as a selection target. Crop J. 2015, 3, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, H.; Fiorani, F.; Pieruschka, R.; Müller-Linow, M.; Jedmowski, C.; Schreiber, L.; Schurr, U.; Rascher, U. Quantitative estimation of leaf heat transfer coefficients by active thermography at varying boundary layer conditions. Front. Plant Sci. 2020, 10, 1684. [Google Scholar] [CrossRef] [Green Version]
- Poblete-Echeverría, C.; Espinace, D.; Sepúlveda-Reyes, D.; Zúñiga, M.; Sanchez, M. Analysis of crop water stress index (CWSI) for estimating stem water potential in grapevines: Comparison between natural reference and baseline approaches. Acta Hortic. 2017, 1150, 189–193. [Google Scholar] [CrossRef]
- Hochberg, U.; Bonel, A.G.; David-Schwartz, R.; Degu, A.; Fait, A.; Cochard, H.; Peterlunger, E.; Herrera, J.C. Grapevine acclimation to water deficit: The adjustment of stomatal and hydraulic conductance differs from petiole embolism vulnerability. Planta 2017, 245, 1091–1104. [Google Scholar] [CrossRef] [PubMed]
- Schultz, H.R.; Matthews, M.A. Xylem development and hydraulic conductance in sun and shade shoots of grapevine (Vitis vinifera L.): Evidence that low light uncouples water transport capacity from leaf area. Planta 1993, 190, 393–406. [Google Scholar] [CrossRef]
- Martorell, S.; Medrano, H.; Tomàs, M.; Escalona, J.M.; Flexas, J.; Diaz-Espejo, A. Plasticity of vulnerability to leaf hydraulic dysfunction during acclimation to drought in grapevines: An osmotic-mediated process. Physiol. Plant. 2015, 153, 381–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardo, S.; Dinis, L.T.; Machado, N.; Moutinho Pereira, J. Grapevine abiotic stress assessment and search for sustainable adaptation strategies in Mediterranean-like climates. A review. Agron. Sustain. Dev. 2018, 38, 66. [Google Scholar] [CrossRef] [Green Version]
- Fernandes de Oliveira, A.; Nieddu, G. Deficit irrigation strategies in Vitis vinifera L. cv. Cannonau under Mediterranean climate. Part II—Cluster microclimate and anthocyanin accumulation patterns. S. Afr. J. Enol. Vitic. 2013, 34, 184–195. [Google Scholar] [CrossRef]
- Fernandes de Oliveira, A.; Mercenaro, L.; Nieddu, G. Assessing thermal efficiency for berry anthocyanin accumulation in four different sites and field-growing conditions. Acta Hortic. 2017, 1188, 181–188. [Google Scholar] [CrossRef]


| Fruit Growth | Fruit Ripening | ||||
|---|---|---|---|---|---|
| Flowering/Fruit Set | Fruit Set/Cluster Closure | Beginning of Veraison | Veraison | Mid-Ripening/Harvest | |
| Vermentino (MPa) | −0.7 | −0.9 | −0.9 | −1.2 | −1.4 |
| Cannonau (MPa) | −0.7 | −0.9 | −1.2 | −1.2 | −1.4 |
| BBCH stage | 65/70 | 71/79 | 80 | 83 | 85/89 |
| BBCH Stages | NHHc (Hours) | P (mm) | ETo (mm) | ETc (mm Day−1) | SWAV (mm Day−1 | CV | Irrigation (mm) | ETa (mm) (Ks) | Yield (t ha−1) | IWP (g L−1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | ||
| Romangia | |||||||||||||||||||
| 10–65 | 629 | 878 | 121 | 109 | 263 | 224 | 3.2 | 2.8 | 7.5 | 8.0 | VMT | 24 | 9 | 354 (6.4) | 233 (3.7) | 9.3 | 14.0 | 38.9 | 155.6 |
| 70–80 | 1433 | 1624 | 4 | 12 | 276 | 177 | 4.8 | 3.1 | 0.0 | 0.1 | |||||||||
| 81–83 | 2244 | 2367 | 20 | 0 | 263 | 170 | 4.7 | 3.0 | 0.0 | 0.0 | CNN | 21.6 | 9 | 356 (5.7) | 233 (3.7) | 8.5 | 9.0 | 39.3 | 100 |
| 85–89 | 3054 | 3100 | 13 | 101 | 186 | 117 | 2.9 | 1.8 | 0.0 | 8.0 | |||||||||
| 90–95 | 3542 | 3447 | 80 | 64 | 95 | 53 | 1.5 | 0.9 | 17.0 | 51.8 | |||||||||
| Gallura | |||||||||||||||||||
| 10–65 | 635 | 843 | 122 | 185 | 263 | 294 | 3.2 | 3.6 | 5.7 | 40.0 | VMT | 60 | 70 | 318 (15.9) | 328 (17.6) | 4.0 | 8.1 | 6.7 | 11.6 |
| 70–80 | 1310 | 1574 | 0 | 14 | 276 | 285 | 4.8 | 5.0 | 0.0 | 0.0 | |||||||||
| 81–83 | 2010 | 2232 | 24 | 0 | 263 | 283 | 4.7 | 5.1 | 0.1 | 0.0 | CNN | 30 | 31 | 348 (7.9) | 367 (7.8) | 5.2 | 7.1 | 17.3 | 22.9 |
| 85–89 | 2775 | 2941 | 122 | 122 | 186 | 199 | 2.9 | 3.1 | 5.2 | 8.3 | |||||||||
| 90–95 | 3263 | 3304 | 53 | 5 | 95 | 91 | 1.5 | 1.5 | 6.9 | 13.4 | |||||||||
| Parteolla | |||||||||||||||||||
| 10–65 | 652 | 859 | 87 | 191 | 261 | 274 | 3.2 | 3.4 | 2.8 | 31.2 | VMT | 60 | 60 | 292 (17.2) | 279 (17.7) | 6.4 | 14.8 | 10.6 | 24.7 |
| 70–80 | 1407 | 1568 | 0 | 18 | 255 | 239 | 4.5 | 4.2 | 0.0 | 0.1 | |||||||||
| 81–83 | 2080 | 2259 | 0 | 4 | 242 | 246 | 4.3 | 4.4 | 0.0 | 0.0 | CNN | 60 | 60 | 292 (17.2) | 279 (17.7) | 5.1 | 10.6 | 8.5 | 17.7 |
| 85–89 | 2819 | 2957 | 150 | 101 | 162 | 168 | 2.5 | 2.6 | 14.2 | 8.0 | |||||||||
| 90–95 | 3317 | 3317 | 53 | 20 | 79 | 73 | 1.3 | 1.2 | 38.4 | 5.9 | |||||||||
| Descriptive Statistics | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| df1 | df2 | Regression Significance | R | R2 | Adj. R2 | ||||||
| Vermentino | 1 | 29 | 0 | 0.881 | 0.777 | 0.769 | |||||
| Cannonau | 1 | 29 | 0 | 0.726 | 0.527 | 0.511 | |||||
| Model Performances | |||||||||||
| Predictors | Unstandardized Coefficients | Variables Sig. | Durbin Watson | MAE | MA%E | RMSE | ENS | ||||
| α | ß | Std. Error | |||||||||
| Vermentino | Intercept | −0.757 | 0.038 | 0.000 | 2.003 | 0.1 | −9.65 | 0.14 | 0.78 | ||
| IIN | 0.725 | 0.000 | 0.000 | ||||||||
| Cannonau | Intercept | −0.831 | 0.042 | 0.000 | 1.893 | 0.13 | −13.68 | 0.17 | 0.53 | ||
| IIN | 0.457 | 0.08 | 0.000 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes de Oliveira, A.; Mameli, M.G.; Lo Cascio, M.; Sirca, C.; Satta, D. An Index for User-Friendly Proximal Detection of Water Requirements to Optimized Irrigation Management in Vineyards. Agronomy 2021, 11, 323. https://doi.org/10.3390/agronomy11020323
Fernandes de Oliveira A, Mameli MG, Lo Cascio M, Sirca C, Satta D. An Index for User-Friendly Proximal Detection of Water Requirements to Optimized Irrigation Management in Vineyards. Agronomy. 2021; 11(2):323. https://doi.org/10.3390/agronomy11020323
Chicago/Turabian StyleFernandes de Oliveira, Ana, Massimiliano Giuseppe Mameli, Mauro Lo Cascio, Costantino Sirca, and Daniela Satta. 2021. "An Index for User-Friendly Proximal Detection of Water Requirements to Optimized Irrigation Management in Vineyards" Agronomy 11, no. 2: 323. https://doi.org/10.3390/agronomy11020323