
Variation among Tanzania Rice Germplasm Collections Based on Agronomic Traits and Resistance to Rice Yellow Mottle Virus

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Description of Experimental Sites
2.3. Experimental Design and Management
2.4. Data Collection
2.5. Data Analysis
3. Results
3.1. Analysis of Variance (ANOVA) for Grain Yield and Yield-Related Traits, and the RYMV Disease Parameter
3.2. Mean Performance of Genotypes for Agronomic Traits and the RYMVD Parameter
3.3. Correlations among Agronomic Traits and RYMVD Reaction
3.4. Principal Components Analysis (PCA)
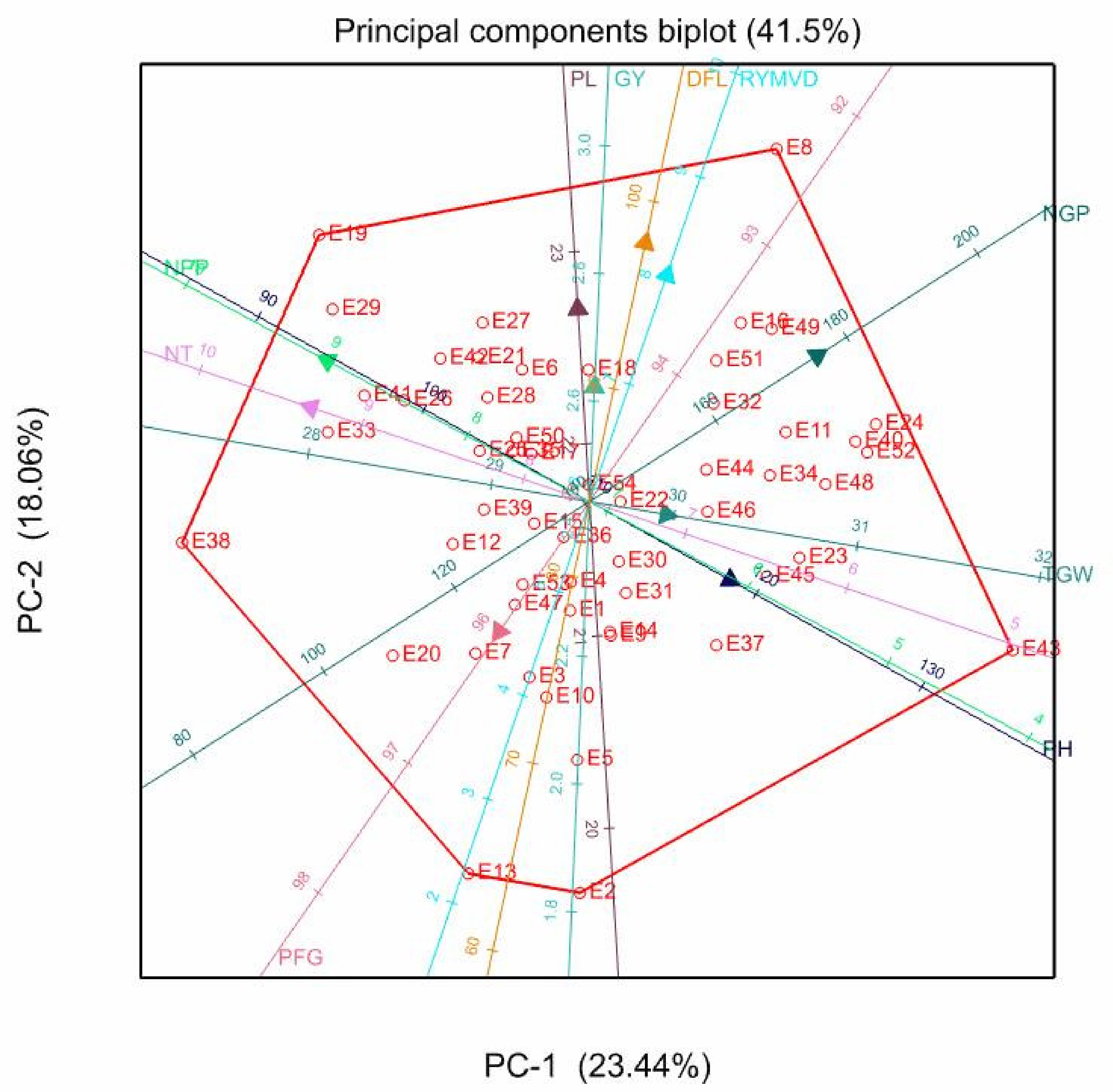
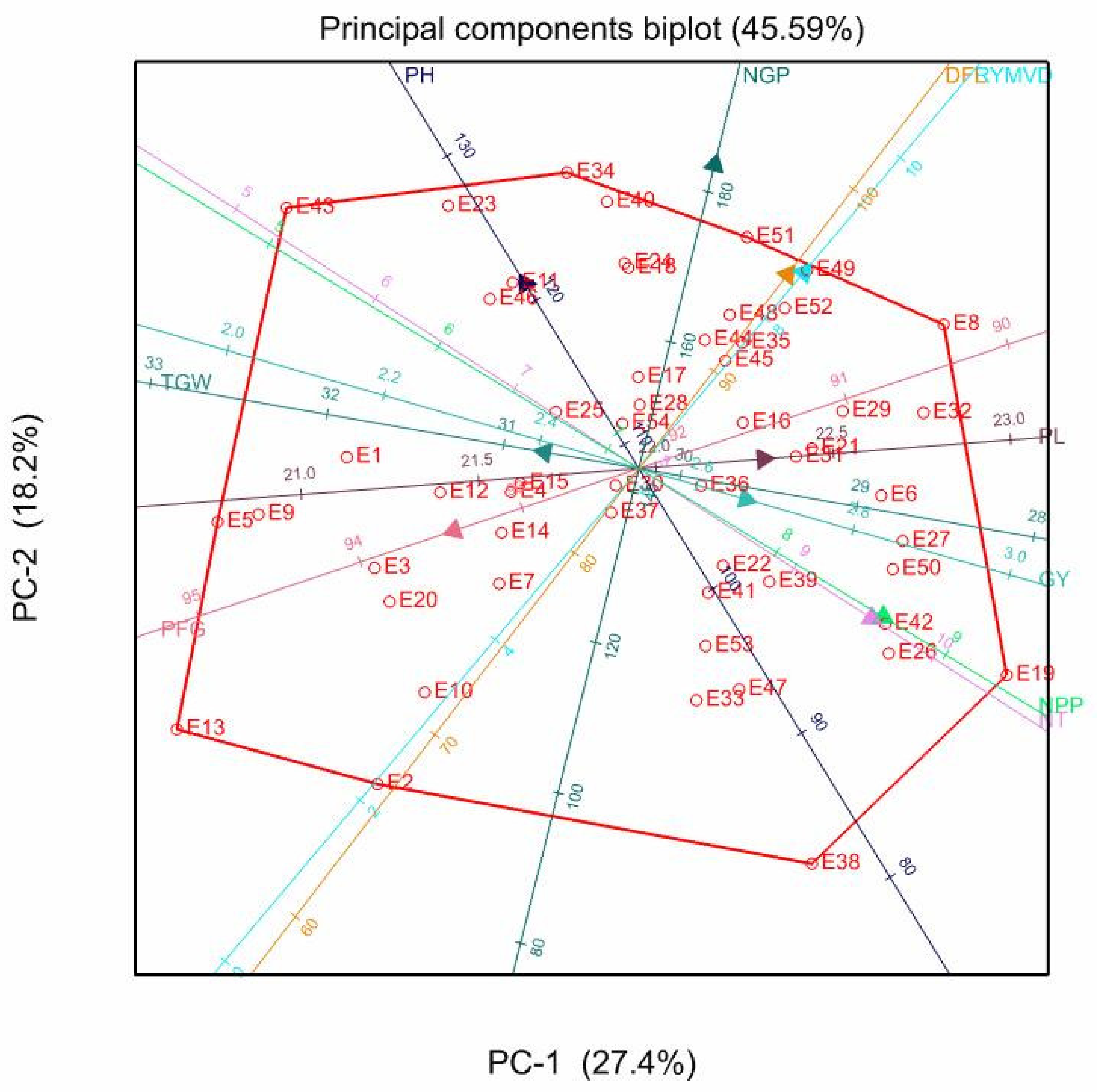
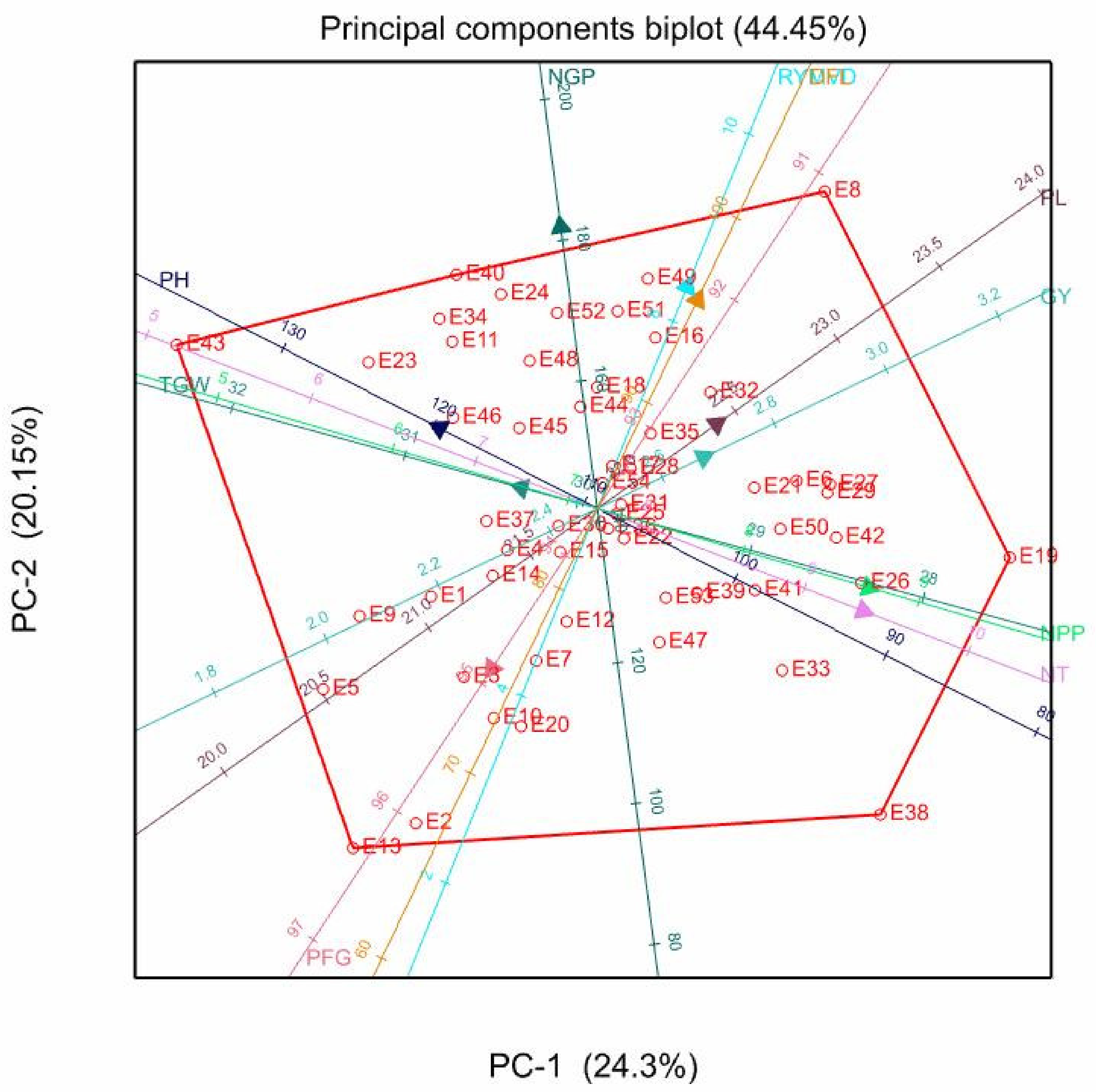
3.5. Principal Component Biplot Analysis
4. Discussion
4.1. Genotypic Variation and Mean Performance
4.2. Traits Associations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mwalyego, F.S.; Maganga, R.M.; Luzi-Kihupi, A.; Ndunguru, A.A.; Kabungo, D.A. Evaluation and deployment of rice genotypes resistant to rice yellow mottle virus disease in the southern highlands of Tanzania. Huria J. 2017, 2, 33–45. [Google Scholar]
- Zhang, Q.J.; Zhu, T.; Xia, E.H.; Shi, C.; Liu, Y.L.; Zhang, Y.; Liu, Y.; Jiang, W.K.; Zhao, Y.J.; Mao, S.Y. Rapid diversification of five Oryza AA genomes associated with rice adaptation. Proc. Natl. Acad. Sci. USA 2014, 111, E4954–E4962. [Google Scholar] [CrossRef] [Green Version]
- Tao, J.; Li, Y.O. Edible insects as a means to address global malnutrition and food insecurity issues. Food Qual. Saf. 2018, 2, 17–26. [Google Scholar] [CrossRef]
- Balasubramanian, V.; Sie, M.; Hijmans, R.J.; Otsuka, K. Increasing rice production in sub-Saharan Africa: Challenges and opportunities. Adv. Agron. 2007, 94, 55–133. [Google Scholar]
- Hubert, J.; Lyimo, H.J.F.; Luzi-Kihupi, A. Geographical variation, distribution and diversity of rice yellow mottle virus phylotypes in Tanzania. Am. J. Plant Sci. 2017, 8, 1264–1284. [Google Scholar]
- Kilimo-Trust. Expanding Rice Markets in the EAC: An Opportunity for Actors in the Value Chain; Kilimo Trust: Dar es Salaam, Tanzania, 2012; 45p. [Google Scholar]
- Food and Agriculture Organization. The Rice Value Chain in Tanzania. A Report from the Southern Highlands food Systems Programme; Wilson, R.T., Lewis, I., Eds.; FAO: Dar es Salaam, Tanzania, 2015; 9p. [Google Scholar]
- Mghase, J.J.; Shiwachi, H.; Nakasone, K.; Takahashi, H. Agronomic and socio-economic constraints to high yield of upland rice in Tanzania. Afr. J. Agric. Res. 2010, 5, 150–158. [Google Scholar]
- Hubert, J.; Luzi-Kihupi, A.; Hébrard, E.; Lyimo, H.J.F. Farmers’ knowledge and perceptions of Rice yellow mottle virus in selected rice growing areas in Tanzania. Int. J. Sci. Res. 2016, 5, 549–559. [Google Scholar]
- Suvi, W.T.; Shimelis, H.; Laing, M. Breeding rice for rice yellow mottle virus resistance in Sub-Saharan Africa: A review. Acta Agric. Scand. Sect. B Soil Plant Sci. 2018, 69, 181–188. [Google Scholar] [CrossRef]
- Lamo, L.; Cho, G.; Jane, I.; Dartey, P.K.A.; James, E.; Ekobu, M.; Alibu, S.; Okanya, S.; Oloka, B.; Otim, M.; et al. Developing lowland rice germplasm with resistance to multiple biotic stresses through another culture in Uganda. Korean Soc. J. Int. Agric. 2015, 27, 415–420. [Google Scholar] [CrossRef]
- Kouassi, N.K.; N’guessan, P.; Albar, L.; Fauquet, C.; Ghesquiere, A. Distribution and characterization of rice yellow mottle virus: A threat to African farmers. Plant Dis. 2005, 89, 124–133. [Google Scholar] [CrossRef] [Green Version]
- Luzi-Kihupi, A.; Zakayo, J.A.; Tusekelege, H.; Mkuya, M.; Kibanda, N.J.M.; Khatib, K.J.; Maerere, A. Mutation breeding for rice improvement in Tanzania. In Induced Plant Mutations in the Genomics Era; FAO: Rome, Italy, 2009; pp. 385–387. [Google Scholar]
- Longué, R.D.S.; Zinga, I.; Semballa, S.; Barro, N.; Traoré, O. Detection and serological characterization of rice yellow mottle virus in Central African Republic. Agric. Sci. 2016, 7, 911–919. [Google Scholar]
- Bakker, W. Characterization and Ecological Aspects of Rice Yellow Mottle Virus in Kenya. Ph.D. Thesis, Wageningen Agricultural University, Wageningen, The Netherlands, 1974; 128p. [Google Scholar]
- Rossel, H.W.; Thottapilly, G.; Buddenhagen, I.W. Occurrence of rice yellow mottle virus in two important rice-growing areas of Nigeria. FAO Plant Prot. Bull. 1982, 30, 137–139. [Google Scholar]
- Hubert, J.G.; Pinel-Galzi, A.; Dibwe, D.; Cinyabuguma, E.; Kabore, A.; Fargette, D.; Silue, D.; Herbrard, E.; Sere, Y. First report of rice yellow mottle virus on rice in the Democratic Republic of Congo. Plant Dis. 2013, 97, 1664. [Google Scholar] [CrossRef] [Green Version]
- Longué, D.R.S.; Galzi-Pinel, A.; Semballa, S.; Zinga, I.; Fargette, D.; Traoré, O. First report of Rice yellow mottle virus in rice in the Central African Republic. Plant Dis. 2014, 98, 162. [Google Scholar] [CrossRef]
- Ochola, D.; Issaka, S.; Rakotomalala, M.; Pinel-Galzi, A.; Ndikumana, I.; Hubert, J.; Hébrard, E.; Séré, Y.; Tusiime, G.; Fargette, D. Emergence of rice yellow mottle virus in eastern Uganda: Recent and singular interplay between strains in East Africa and in Madagascar. J. Virus Res. 2015, 195, 64–72. [Google Scholar] [CrossRef]
- Kanyeka, Z.L.; Sangu, E.; Fargette, D.; Pinel-Galzi, A.; Hébrard, E. Distribution and diversity of local strains of Rice yellow mottle virus in Tanzania. Afr. J. Crop Sci. 2007, 15, 201–209. [Google Scholar]
- Sy, A.A.; Sere, Y. Manuel de Formation en Pathologie du Riz; Association pour le Developpment de la Riziculture en Afrique de L’OUEST (ADRAO): Bouake, Cote d’Ivoire, 1996. [Google Scholar]
- Traoré, O.; Pinel-Galzi, A.; Issaka, S.; Poulicard, N.; Aribi, J.; Aké, S.; Ghesquiéré, A.; Séré, Y.; Konaté, G.; Hébrard, E.; et al. The adaptation of Rice yellow mottle virus to the elF(iso)4G-mediated rice resistance. J. Virol. 2010, 408, 103–108. [Google Scholar] [CrossRef] [Green Version]
- N’Guessan, P.; Pinel, A.; Caruana, M.L.; Frutos, R.; Sy, A.; Ghesquière, A.; Fargette, D. Evidence of the presence of two serotypes of rice yellow mottle sobemovirus in Côte d’Ivoire. Eur. J. Plant Pathol. 2000, 106, 167–178. [Google Scholar] [CrossRef]
- Mpunami, A.; Ndikumana, I.; Hubert, J.; Pinel-Galzi, A.; Kibanda, N.; Mwalyego, F.; Tembo, P.; Kola, B.; Mkuya, M.; Kanyeka, Z.; et al. Tanzania, biodiversity hotspot of Rice yellow mottle virus. In Proceedings of the 12th International Plant Virus Epidemiology Symposium, Arusha, Tanzania, 29 January–1 February 2013. 70p. [Google Scholar]
- Longué, D.R.S.; Galzi-Pinel, A.; Zinga, I.; Semballa, S.; Fargette, D.; Baro, N.; Traoré, O. Genetic diversity and molecular epidemiology of rice yellow mottle virus in Central African Republic. Int. J. Virol. 2017, 13, 80–89. [Google Scholar] [CrossRef]
- Kam, H.; Laing, M.D.; Sere, Y.; Thiemele, D.; Ghesquiere, A.; Ahmadi, N.; Ndjiondjop, M.-N. Evaluation of a collection of rice landraces from Burkina Faso for resistance or tolerance to rice yellow mottle virus. J. Plant Pathol. 2013, 95, 485–492. [Google Scholar]
- Koudamilor, A.; Nwilene, F.E.; Togola, A.; Akogbeto, M. Insect vectors of rice yellow mottle virus. J. Insects 2015. [Google Scholar] [CrossRef] [Green Version]
- Abo, M.E.; Alegbejo, M.D.; Sy, A.A.; Misari, S.M. An overview of the mode of transmission, host plants and methods of detection of rice yellow mottle virus. J. Sustain. Agric. 2000, 17, 19–36. [Google Scholar] [CrossRef]
- Sereme, D.; Ouedraogo, I.; Neya, B.J.; Zida, P.E.; Yao, N.; Sie, M. Screening improved rice varieties (Oryza spp.) for their resistance /tolerance to rice yellow mottle virus in West Africa. Int. J. Agric. Innov. Res. 2016, 5, 481–486. [Google Scholar]
- Traoré, V.S.E.; Néya, B.J.; Camara, M.; Gracen, V.; Offei, S.K.; Traoré, O. Farmers’ perception and impact of rice yellow mottle disease on rice yields in Burkina Faso. Agric. Sci. 2015, 6, 943–952. [Google Scholar] [CrossRef] [Green Version]
- Pidon, H.; Ghesquière, A.; Chéron, S.; Issaka, S.; Hébrard, E.; Sabot, F.; Kolade, O.; Silué, D.; Albar, L. Fine mapping of RYMV3: A new resistance gene to rice yellow mottle virus from Oryza glaberrima. Theor. Appl. Genet. 2017, 130, 807–818. [Google Scholar] [CrossRef]
- Nkuba, J.; Ndunguru, A.; Madulu, R.; Lwezaura, D.; Kajiru, G.; Babu, A.; Chalamila, B.; Ley, G. Rice value chain analysis in Tanzania: Identification of constraints, opportunities, and upgrading strategies. Afr. Crop Sci. J. 2016, 24, 73–87. [Google Scholar] [CrossRef] [Green Version]
- Ndjiondjop, M.N.; Albar, L.; Fargette, D.; Fauquet, C.; Ghesquiere, A. The genetic basis of high resistance to rice yellow mottle virus (RYMV) in cultivars of two cultivated rice species. Plant Dis. 1999, 83, 931–935. [Google Scholar] [CrossRef]
- Thiémélé, D.; Boisnard, A.; Ndjiondjop, M.N.; Chéron, S.; Séré, Y.; Aké, S.; Ghesquière, A.; Albar, L. Identification of a second major resistance gene to Rice yellow mottle virus, RYMV2, in the African cultivated rice species, O. glaberrima. J. Theor. Appl. Genet. 2010, 121, 169–179. [Google Scholar]
- Pinel-Galzi, A.; Dubreuil-Tranchant, C.; Hébrard, E.; Mariac, C.; Ghesquière, A.; Albar, L. Mutations in rice yellow mottle virus polyprotein P2a involved in RYMV2 gene resistance breakdown. Front. Plant Sci. 2016, 7, 1779. [Google Scholar] [CrossRef] [Green Version]
- Munganyinka, E.; Edema, R.; Lamo, J.; Gibson, P. The reaction of intraspecific and interspecific rice cultivars for resistance to rice yellow mottle virus disease. Eur. J. Exp. Biol. 2016, 6, 13–18. [Google Scholar]
- Xiao, X.Y.; Wang, Y.P.; Zhang, J.Y.; Li, S.G.; Rong, T.Z. SSR marker based genetic diversity fingerprinting of hybrid rice in Sichuan, China. China J. Rice Sci. 2016, 20, 1–7. [Google Scholar]
- Mausa, E.A. Genetic Diversity of Rice (Oryza sativa L.) Landraces Conserved at the National Genebank as Revealed by Simple Sequence Repeat (SSR) Dna Markers. Master’s Thesis, Degree at Sokoine University of Agriculture, Morogoro, Tanzania, 2014; p. 83. [Google Scholar]
- Bakari, M.M. Genetic Diversity of Some Rice (Oryza sativa L.) landraces Grown in Tanzania Using Simple Sequence Repeats (SSR). Master’s Thesis, Sokoine University of Agriculture, Morogoro, Tanzania, 2010; 72p. [Google Scholar]
- Suvi, W.T.; Shimelis, H.; Laing, M.; Mathew, I.; Shayanowako, A.I.T. Assessment of the genetic diversity and population structure of rice genotypes using SSR markers. Acta Agric. Scand. Sect. B Soil Plant Sci. 2020, 70, 76–86. [Google Scholar] [CrossRef]
- Aida, A.; Huseain, M.; Mohamad, A.S.; Hani, S.; Sameer, M. Assessment of genetic variation among Jordanian barely landraces (Hordeum vulgare) as revealed by molecular markers. Am. Eurasian J. Agric. Environ. Sci. 2007, 2, 68–74. [Google Scholar]
- Mangosongo, H.M.; Lyaruu, H.V.; Mneney, E.E. Agro-morphological characterization of the wild rice (Oryza longistaminata) populations from selected areas of Tanzania. Tanzan. J. Sci. 2019, 45, 9–17. [Google Scholar]
- Wilson, R.T. The rice value chain in Tanzania. Int. J. Agric. Innov. Res. 2018, 7, 237–244. [Google Scholar]
- International Rice Research Institute (Ed.) Standard Evaluation System for Rice, 5th ed.; Genetic Resources Center, IRRI: Manila, Philippines, 2002; 27p. [Google Scholar]
- Payne, R.W.; Murray, D.A.; Harding, S.A. An Introduction to the GenStat Command Language, 18th ed.; VSN International: Hemel Hempstead, UK, 2017. [Google Scholar]
- SPSS. Statistical Package for Social Scientists; SPSS for Windows Release 24.0; SPSS: Chicago, IL, USA, 2017. [Google Scholar]
- Adhikari, B.N.; Joshi, B.P.; Shrestha, J.; Bhatta, N.R. Genetic variability, heritability, genetic advance and correlation among yield and yield components of rice (Oryza sativa L.). J. Agric. Nat. Resour. 2018, 1, 149–160. [Google Scholar] [CrossRef]
- Gyawali, S.; Poudel, A.; Poudel, S. Genetic variability and association analysis in different rice genotypes in mid hill of western Nepal. Acta Sci. Agric. 2018, 2, 69–76. [Google Scholar]
- Sumanth, V.; Suresh, B.G.; Ram, B.J.; Srujana, G. Estimation of genetic variability, heritability and genetic advance for grain yield components in rice (Oryza sativa L.). J. Pharmacogn. Phytochem. 2017, 6, 1437–1439. [Google Scholar]
- Bandi, H.R.K.; Satyanarayana, P.V.; Babu, D.R.; Chamundeswari, N.; Rao, V.S.; Raju, S.K. Genetic variability estimates for yield and yield components traits and quality traits in rice (Oryza sativa L.). Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 551–559. [Google Scholar] [CrossRef]
- Mercer, K.L.; Perales, H.G. Evolutionary response of landraces to climate change in centres of crop diversity. Evol. Appl. 2010, 3, 480–493. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, N.; Yadaw, R.B.; Chaudhary, B.; Prasai, H.; Iftekharuddaula, K.; Venkateshwarlu, C.; Annamalai, A.; Xangsayasane, P.; Battan, K.R.; Ram, M.; et al. Evaluating the performance of rice genotypes for improving yield and adaptability under direct seeded aerobic cultivation conditions. Front. Plant Sci. 2019, 10, 159. [Google Scholar] [CrossRef]
- Joseph, A.; Olufolaji, D.B.; Mwilene, F.E.; Onasanya, A.; Omale, M.M.; Onasanya, R.O.; Sere, Y. Effect of leaf age on rice yellow mottle virus severity chlorophyll content with mechanical inoculation and vector transmission method. Trends Appl. Sci. Res. 2011, 6, 1345–1351. [Google Scholar] [CrossRef] [Green Version]
- Hubert, J.; Lyimo, H.J.F.; Luzi-Kihupi, A. Pathogenic variation and occurrence of multiple resistance-breaking rice yellow mottle virus strains in Tanzania. Afr. J. Biotechnol. 2017, 24, 1331–1337. [Google Scholar]
- Bustos-Korts, D.; Romagosa, I.; Borràs-Gelonch, G.; Casas, A.M.; Slafer, G.A.; van Eeuwijk, F.A. Genotype by Environment Interaction and Adaptation. Encyclopedia of Sustainability Science and Technology; Springer Science and Business Media, LLC: Berlin/Heidelberg, Germany, 2018; pp. 29–71. [Google Scholar]
- Romagosa, I.; Fox, P.N. Genotype × Environment Interaction and Adaptation; Hayward, M.D., Bosemark, N.O., Romagosa, I., Cerezo, M., Eds.; Plant Breeding; Plant Breeding Series; Springer: Dordrecht, The Netherlands, 1993. [Google Scholar]
- Bhuvaneswari, S.; Kumar, S.; Singh, I.M.; Takhellambam, S.; Shasidhar, K.S.; Singh, R.Y.; Prakash, N. Genetic variability and association studies on grain yield components in F2 populations of black rice (Oryza sativa L.) of Manipur. Indian J. Hill Farming 2015, 28, 85–89. [Google Scholar]
- Getachew, M.; Burhan, T. Growth and yield of rice (Oryza sativa L.) as affected by time and ratio of nitrogen application at Jimma, South-West Ethiopia. Int. J. Agric. Innov. Res. 2017, 4, 175–182. [Google Scholar]
- Li, F.; Xie, J.; Zhu, X.; Wang, X.; Zhao, Y.; Ma, X.; Zhang, Z.; Rashid, M.A.R.; Zhang, Z.; Zhi, L.; et al. Genetic basis underlying correlations among growth duration and yield traits revealed by GWAS in rice (Oryza sativa L.). Front. Plant Sci. 2018, 9, 650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, Q.; Chen, L.; Mei, H.; Wei, H.; Feng, F.; Wang, P.; Xia, H.; Li, T.; Luo, L. Quantitative trait locus mapping of deep rooting by linkage and association analysis in rice. J. Exp. Bot. 2015, 66, 4749–4757. [Google Scholar] [CrossRef] [Green Version]
- Onwughalu, J.T.; Abo, M.E.; Okoro, J.K.; Onasanya, A.; Sere, Y. Rice yellow mottle virus infection and reproductive losses in rice (Oryza sativa Linn.). Trends Appl. Sci. Res. 2011, 6, 182–189. [Google Scholar] [CrossRef] [Green Version]
- Sahu, H.; Saxena, R.R.; Verulkar, S.B.; Kumar, V. Principal component analysis of agro-morphological traits in RIL population of rice (Oryza sativa L.) grown in a irrigated lowland water regime. Green Farming 2017, 8, 11–15. [Google Scholar]
- Yugandhar, P.R.; Kota, S.; Kiran, B.U.; Sridhar, M. Principal component analysis for agro-morphological and quality characters in germplasm of rice (Oryza sativa L.). Int. J. Adv. Biol. Res. 2018, 8, 268–273. [Google Scholar]
- Ranjith, P.; Sahu, S.; Dash, S.K.; Bastia, D.N.; Pradhan, B.D. Principal component analysis of rice varieties under bacterial leaf blight incidence. Int. J. Chem. Stud. 2019, 7, 4184–4187. [Google Scholar]
- Nachimuthu, V.V.; Robin, S.; Sudhakar, D.; Raveendran, M.; Rajeswari, S.; Manonmani, S. Evaluation of rice genetic diversity and variability in a population panel by principal component analysis. Indian J. Sci. Technol. 2014, 7, 1555–1562. [Google Scholar] [CrossRef]
- Gana, A.S.; Shaba, S.Z.; Tsado, E.K. Principal component analysis of morphological traits in thirty-nine accessions of rice (Oryza sativa L.) grown in a rainfed lowland ecology of Nigeria. J. Plant Breed. Crop Sci. 2013, 5, 120–126. [Google Scholar]
- Flores, F.; Moreno, M.T.; Cubbero, J.I. A comparison of univariate and multivariate methods to analyze G × E interaction. Field Crop Res. 1998, 56, 271–286. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sr. No | Genotypes | Origin/Source | Sr. No. | Genotypes | Origin/Source |
|---|---|---|---|---|---|
| 1 | Salama M-57 | SUA/Tanzania | 28 | Kalubangala | Landrace/Tanzania |
| 2 | SSD 1 | SUA/Tanzania | 29 | Mpaka wa bibi | Landrace/Tanzania |
| 3 | Nerica 7 | AfricaRice/Benin | 30 | Mbawambili nyekundu | Landrace/Tanzania |
| 4 | Kalalu | SUA/Tanzania | 31 | Limota | Landrace/Tanzania |
| 5 | IRAT 256 | AfricaRice/Benin | 32 | Moshi | Landrace/Tanzania |
| 6 | SARO | TARI/Tanzania | 33 | Shingo ya mwali | Landrace/Tanzania |
| 7 | Nerica 1 | AfricaRice/Benin | 34 | Kalundi | Landrace/Tanzania |
| 8 | Serena | Landrace/Tanzania | 35 | IR54 | IRRI/Philippines |
| 9 | Nerica 4 | AfricaRice/Benin | 36 | TXD 88 | TARI/Tanzania |
| 10 | WAB450 | AfricaRice/Benin | 37 | IR 56 | IRRI/Philippines |
| 11 | Mbega | Landrace/Tanzania | 38 | IR64 | IRRI/Philippines |
| 12 | Salama M-55 | SUA/Tanzania | 39 | Mzinga | Landrace/Tanzania |
| 13 | Mwangaza | SUA/Tanzania | 40 | Afaa mwanza | Landrace/Tanzania |
| 14 | Nerica 2 | AfricaRice/Benin | 41 | TXD 85 | TARI/Tanzania |
| 15 | Lunyuki | TARI/Tanzania | 42 | TXD 307 | TARI/Tanzania |
| 16 | Turiani | Landrace/Tanzania | 43 | Sumbawanga | Landrace/Tanzania |
| 17 | Mbawa ya njiwa | Landrace/Tanzania | 44 | Supa | Landrace/Tanzania |
| 18 | Chamota | Landrace/Tanzania | 45 | Rangi mbili nyekundu | Landrace/Tanzania |
| 19 | IR72 | IRRI/Philippines | 46 | Faya mzinga | Landrace/Tanzania |
| 20 | Salama M-19 | SUA/Tanzania | 47 | TAI | TARI/Tanzania |
| 21 | Masantula | Landrace/Tanzania | 48 | Gombe | Landrace/Tanzania |
| 22 | IR 68 | IRRI/Philippines | 49 | Kisegese | Landrace/Tanzania |
| 23 | Kalamata | Landrace/Tanzania | 50 | Gigante | AfricaRice |
| 24 | Zambia | Landrace/Tanzania | 51 | Sindano nyeupe | Landrace/Tanzania |
| 25 | Ringa | Landrace/Tanzania | 52 | Kihogo red | Landrace/Tanzania |
| 26 | Wahiwahi | Landrace/Tanzania | 53 | Cherehani | Landrace/Tanzania |
| 27 | Mwanza | Landrace/Tanzania | 54 | ITA 303 | TARI/Tanzania |
| Source of Variation | DF | DFL | NT | NPP | PH | PL | NGP | PFG | TGW | RYMVD | GY |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Site | 1 | 0.00 ns | 6.97 * | 21.41 *** | 797.72 *** | 228.93 *** | 1533.33 *** | 1561.80 *** | 97.34 *** | 1.85 *** | 52.25 *** |
| Rep (Site) | 1 | 0.93 ns | 0.47 ns | 1.16 ns | 22.89 ns | 5.55 * | 22.26 ns | 772.20 *** | 2.08 | 0.00 | 0.17 * |
| Block (Rep) | 32 | 167.83 *** | 2.82 *** | 2.20 ** | 371.84 *** | 3.25 *** | 623.91 *** | 26.53 *** | 10.19 *** | 7.73 *** | 0.42 *** |
| Genotype | 53 | 450.42 *** | 3.95 *** | 4.13 *** | 945.34 *** | 3.25 *** | 2539.17 *** | 10.57 *** | 52.20 *** | 7.62 *** | 1.65 *** |
| Genotype × Site | 53 | 0.00 ns | 1.27 ns | 1.34 ns | 120.77 *** | 4.11 *** | 146.09 *** | 11.67 *** | 4.98 ** | 1.73 *** | 0.59 *** |
| Residual | 106 | 1.40 | 1.22 | 1.15 | 31.37 | 1.28 | 36.83 | 4.78 | 2.39 | 0.08 | 0.04 |
| Entry | Genotype | DFL | NT | NPP | PH (cm) | PL (cm) | NGP | PFG | TGW (g) | RYMVD | GY (t/ha) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ifa | Mk | Ifa | Mk | Ifa | Mk | Ifa | Mk | Ifa | Mk | Ifa | Mk | Ifa | Mk | Ifa | Mk | Ifa | Mk | Ifa | Mk | ||
| 1 | Salama M-57 | 86 | 84 | 8 | 6 | 7 | 6 | 133.2 | 130.9 | 22.8 | 22.4 | 141 | 143 | 96.3 | 92.9 | 30.5 | 34.5 | 1 | 1 | 2.0 | 2.3 |
| 2 | SSD1 | 60 | 63 | 7 | 7 | 6 | 5 | 123.9 | 124.0 | 20.0 | 20.9 | 100 | 99 | 95.1 | 92.7 | 33.0 | 35.5 | 1 | 1 | 2.7 | 2.8 |
| 3 | Nerica 7 | 74 | 76 | 8 | 7 | 7 | 7 | 108.7 | 102.3 | 19.2 | 20.8 | 142 | 135 | 96.1 | 97.4 | 26.5 | 30.5 | 3 | 3 | 2.2 | 2.0 |
| 4 | Kalalu | 77 | 74 | 7 | 6 | 7 | 6 | 100.1 | 95.0 | 19.3 | 22.3 | 142 | 137 | 94.5 | 87.1 | 23.0 | 25.5 | 3 | 3 | 2.3 | 2.4 |
| 5 | IRAT 256 | 73 | 76 | 7 | 5 | 7 | 4 | 156.7 | 146.2 | 24.0 | 25.3 | 106 | 105 | 96.3 | 95.5 | 29.5 | 33.0 | 1 | 1 | 1.4 | 1.7 |
| 6 | Gigante | 95 | 93 | 10 | 11 | 10 | 10 | 97.1 | 98.5 | 19.7 | 22.0 | 176 | 181 | 96.4 | 91.4 | 30.0 | 30.0 | 5 | 5 | 3.7 | 3.5 |
| 7 | Nerica 1 | 74 | 72 | 8 | 7 | 7 | 6 | 85.1 | 89.9 | 19.4 | 20.2 | 127 | 164 | 95.7 | 89.5 | 29.5 | 32.5 | 3 | 3 | 2.5 | 2.6 |
| 8 | Serena | 91 | 94 | 8 | 9 | 7 | 8 | 110.2 | 112.9 | 23.6 | 23.9 | 183 | 184 | 84.1 | 93.3 | 31.0 | 31.5 | 5 | 3 | 3.6 | 3.3 |
| 9 | Nerica 4 | 78 | 76 | 7 | 5 | 6 | 5 | 101.5 | 96.9 | 20.1 | 22.3 | 116 | 119 | 95.1 | 91.6 | 29.0 | 29.5 | 5 | 3 | 1.0 | 1.1 |
| 10 | WAB450 | 65 | 63 | 8 | 8 | 6 | 7 | 103.1 | 99.4 | 20.2 | 21.1 | 118 | 98 | 94.8 | 95.9 | 28.0 | 30.5 | 5 | 5 | 1.8 | 1.7 |
| 11 | Mbega | 85 | 82 | 7 | 6 | 6 | 6 | 127.5 | 126.8 | 23.2 | 22.2 | 160 | 156 | 95.4 | 87.2 | 36.4 | 36.5 | 5 | 5 | 3.9 | 3.6 |
| 12 | Salama M-55 | 86 | 89 | 9 | 6 | 9 | 7 | 133.2 | 117.6 | 21.1 | 21.3 | 149 | 163 | 96.0 | 95.6 | 35.0 | 35.0 | 1 | 1 | 1.5 | 3.2 |
| 13 | Mwangaza | 67 | 70 | 8 | 7 | 7 | 6 | 116.5 | 99.2 | 19.6 | 21.0 | 91 | 85 | 94.7 | 94.8 | 37.0 | 37.3 | 1 | 1 | 1.2 | 1.3 |
| 14 | Nerica 2 | 77 | 79 | 7 | 7 | 6 | 6 | 85.6 | 86.1 | 19.2 | 21.1 | 137 | 140 | 93.4 | 90.0 | 25.5 | 32.0 | 3 | 3 | 1.9 | 2.1 |
| 15 | Lunyuki | 78 | 76 | 8 | 7 | 8 | 7 | 124.0 | 122.1 | 18.9 | 21.5 | 146 | 139 | 95.5 | 89.5 | 29.5 | 32.5 | 1 | 1 | 3.2 | 3.3 |
| 16 | SARO | 90 | 93 | 7 | 7 | 6 | 6 | 89.0 | 87.4 | 23.0 | 22.9 | 154 | 153 | 91.0 | 91.8 | 34.0 | 34.0 | 5 | 5 | 3.9 | 4.0 |
| 17 | Mbawa ya njiwa | 76 | 80 | 8 | 7 | 8 | 7 | 112.8 | 109.7 | 20.2 | 22.9 | 142 | 156 | 95.1 | 88.7 | 28.0 | 27.4 | 7 | 5 | 1.9 | 2.1 |
| 18 | Chamota | 91 | 89 | 8 | 7 | 8 | 6 | 118.0 | 127.6 | 19.1 | 22.6 | 164 | 168 | 95.5 | 88.4 | 25.5 | 23.5 | 7 | 5 | 2.7 | 2.2 |
| 19 | IR72 | 92 | 90 | 9 | 9 | 8 | 8 | 89.5 | 84.5 | 20.8 | 23.8 | 155 | 152 | 93.6 | 89.4 | 29.5 | 30.5 | 5 | 3 | 2.0 | 2.5 |
| 20 | Salama M-19 | 79 | 81 | 9 | 6 | 8 | 5 | 115.1 | 114.4 | 20.5 | 22.4 | 129 | 115 | 96.9 | 91.4 | 30.5 | 32.5 | 1 | 1 | 1.7 | 1.8 |
| 21 | Masantula | 102 | 101 | 8 | 9 | 8 | 8 | 124.0 | 126.7 | 20.1 | 22.8 | 109 | 123 | 96.9 | 88.7 | 23.0 | 26.5 | 7 | 5 | 2.1 | 2.5 |
| 22 | IR68 | 94 | 90 | 7 | 9 | 6 | 8 | 87.6 | 87.9 | 19.2 | 22.9 | 147 | 143 | 96.2 | 89.9 | 25.0 | 26 | 3 | 3 | 1.9 | 2.1 |
| 23 | Kalamata | 91 | 96 | 7 | 6 | 6 | 6 | 126.4 | 118.4 | 18.7 | 19.1 | 168 | 174 | 95.6 | 89.6 | 34.0 | 35.0 | 5 | 5 | 2.7 | 2.7 |
| 24 | Zambia | 90 | 91 | 6 | 7 | 6 | 6 | 115.8 | 125.2 | 21.5 | 20.5 | 177 | 179 | 90.9 | 92.3 | 30.0 | 27.5 | 5 | 5 | 2.5 | 2.8 |
| 25 | Ringa | 73 | 69 | 9 | 7 | 9 | 7 | 116.2 | 113.3 | 20.8 | 21.5 | 163 | 161 | 95.4 | 90.9 | 31.0 | 33.0 | 6 | 7 | 1.5 | 2.0 |
| 26 | Rangimbili nyekundu | 73 | 75 | 10 | 10 | 8 | 9 | 105.6 | 112.9 | 21.3 | 23.4 | 97 | 139 | 93.6 | 89.9 | 32.5 | 34.0 | 7 | 5 | 3.7 | 3.8 |
| 27 | Mwanza | 88 | 87 | 8 | 7 | 8 | 6 | 78.5 | 77.0 | 19.6 | 23.1 | 143 | 154 | 94.5 | 88.3 | 26.5 | 32.0 | 7 | 5 | 1.5 | 1.7 |
| 28 | Kalubangala | 88 | 89 | 8 | 6 | 7 | 6 | 84.5 | 108.3 | 19.0 | 24.7 | 115 | 160 | 96.0 | 85.4 | 29.5 | 35.5 | 7 | 3 | 2.8 | 2.2 |
| 29 | Mpaka wa bibi | 103 | 104 | 9 | 8 | 9 | 7 | 104.3 | 113.3 | 22.2 | 23.6 | 114 | 145 | 96.1 | 89.1 | 23.0 | 23.5 | 5 | 7 | 1.7 | 2.3 |
| 30 | Mbawambili | 71 | 72 | 7 | 8 | 7 | 7 | 116.6 | 116.4 | 21.2 | 22.3 | 123 | 134 | 93.9 | 89.7 | 28.5 | 27.5 | 7 | 5 | 2.3 | 2.0 |
| 31 | Limota | 79 | 80 | 7 | 7 | 7 | 6 | 116.5 | 109.9 | 19.8 | 21.4 | 143 | 152 | 95.0 | 85.1 | 24.5 | 24.0 | 5 | 7 | 1.5 | 2.0 |
| 32 | Moshi | 92 | 93 | 7 | 7 | 7 | 6 | 129.0 | 126.6 | 21.6 | 23.7 | 169 | 174 | 96.5 | 87.3 | 28.0 | 29.2 | 5 | 5 | 2.8 | 3.1 |
| 33 | Shingo ya mwali | 73 | 74 | 9 | 9 | 9 | 8 | 110.7 | 104.1 | 21.9 | 25.0 | 102 | 103 | 96.5 | 91.7 | 33.5 | 36.0 | 5 | 5 | 3.1 | 3.3 |
| 34 | Kalundi | 99 | 101 | 7 | 6 | 6 | 6 | 127.2 | 105.5 | 21.5 | 22.6 | 163 | 166 | 96.5 | 88.9 | 30.5 | 29.0 | 5 | 7 | 1.9 | 2.0 |
| 35 | IR54 | 90 | 94 | 8 | 6 | 8 | 5 | 95.2 | 91.1 | 19.6 | 21.5 | 148 | 176 | 94.8 | 84.9 | 27.0 | 27.5 | 5 | 3 | 2.0 | 2.0 |
| 36 | TXD88 | 92 | 95 | 7 | 9 | 7 | 7 | 90.0 | 86.0 | 19.8 | 21.4 | 126 | 149 | 95.2 | 86.0 | 29.5 | 32.0 | 5 | 3 | 3.1 | 2.9 |
| 37 | IR 56 | 77 | 74 | 6 | 7 | 6 | 7 | 96.4 | 97.8 | 19.8 | 21.7 | 163 | 141 | 94.4 | 86.1 | 22.5 | 26.0 | 3 | 3 | 2.3 | 2.4 |
| 38 | IR 64 | 75 | 79 | 9 | 10 | 8 | 9 | 86.9 | 85.8 | 20.7 | 21.7 | 98 | 102 | 96.6 | 88.8 | 27.0 | 28.5 | 3 | 3 | 2.8 | 3.2 |
| 39 | Mzinga | 92 | 95 | 8 | 9 | 7 | 8 | 98.3 | 87.3 | 20.4 | 21.8 | 117 | 129 | 95.8 | 88.8 | 26.5 | 27.5 | 5 | 5 | 2.0 | 2.3 |
| 40 | Afaa Mwanza | 89 | 92 | 6 | 7 | 6 | 6 | 117.3 | 117.6 | 22.3 | 22.1 | 168 | 166 | 93.5 | 87.3 | 31.5 | 35.5 | 7 | 5 | 1.8 | 1.8 |
| 41 | TXD 85 | 97 | 95 | 9 | 7 | 8 | 7 | 83.1 | 82.1 | 19.8 | 22.3 | 119 | 117 | 96.1 | 85.4 | 29.0 | 30.5 | 3 | 3 | 2.2 | 3.1 |
| 42 | TXD 307 | 98 | 100 | 8 | 10 | 8 | 8 | 89.4 | 78.9 | 19.1 | 23.8 | 110 | 113 | 93.2 | 85.1 | 29.0 | 30.5 | 3 | 3 | 1.8 | 2.5 |
| 43 | Sumbawanga | 80 | 81 | 5 | 5 | 4 | 5 | 123.3 | 123.6 | 22.7 | 20.4 | 179 | 173 | 96.2 | 93.1 | 34.0 | 35.0 | 5 | 5 | 2.6 | 2.8 |
| 44 | Supa | 84 | 87 | 7 | 7 | 7 | 7 | 130.0 | 115.9 | 20.9 | 23.2 | 153 | 169 | 95.9 | 89.8 | 34.0 | 33.0 | 5 | 7 | 1.9 | 2.5 |
| 45 | Wahiwahi | 80 | 83 | 6 | 6 | 6 | 5 | 121.3 | 116.3 | 22.4 | 22.5 | 159 | 168 | 83.7 | 84.5 | 25.0 | 26.0 | 5 | 7 | 1.5 | 1.4 |
| 46 | Faya mzinga | 87 | 88 | 8 | 7 | 6 | 6 | 128.0 | 119.3 | 20.9 | 21.0 | 156 | 172 | 96.4 | 91.3 | 34.5 | 35.0 | 5 | 5 | 3.2 | 3.4 |
| 47 | TAI | 79 | 80 | 7 | 9 | 7 | 8 | 95.0 | 87.6 | 20.5 | 22.0 | 116 | 112 | 96.1 | 85.5 | 26.0 | 28.0 | 3 | 3 | 3.5 | 3.7 |
| 48 | Gombe | 88 | 89 | 6 | 6 | 6 | 6 | 132.4 | 126.0 | 22.1 | 23.7 | 166 | 165 | 96.3 | 90.6 | 29.5 | 29.0 | 5 | 7 | 1.9 | 2.4 |
| 49 | Kisegese | 95 | 96 | 7 | 6 | 7 | 6 | 106.9 | 102.7 | 19.5 | 23.2 | 181 | 183 | 93.0 | 89.1 | 36.5 | 34.5 | 5 | 7 | 1.3 | 2.4 |
| 50 | Turiani | 88 | 89 | 8 | 7 | 8 | 6 | 94.4 | 93.2 | 20.9 | 21.0 | 145 | 157 | 96.2 | 85.5 | 32.5 | 34.5 | 5 | 5 | 2.6 | 3.1 |
| 51 | Sindano nyeupe | 97 | 98 | 7 | 8 | 7 | 7 | 127.7 | 136.2 | 22.2 | 23.0 | 160 | 169 | 93.8 | 90.7 | 26.5 | 27.0 | 5 | 7 | 2.1 | 2.7 |
| 52 | Kihogo red | 95 | 96 | 6 | 6 | 6 | 6 | 124.0 | 114.5 | 20.5 | 22.4 | 164 | 174 | 93.8 | 89.6 | 32.0 | 35.0 | 7 | 5 | 2.3 | 2.0 |
| 53 | Cherehani | 57 | 56 | 7 | 8 | 7 | 7 | 93.8 | 109.3 | 21.1 | 24.3 | 106 | 105 | 91.0 | 87.3 | 29.0 | 33.5 | 3 | 5 | 2.2 | 2.8 |
| 54 | ITA 303 | 85 | 81 | 8 | 9 | 8 | 7 | 131.3 | 126.1 | 21.0 | 22.8 | 150 | 147 | 96.2 | 86.4 | 33.0 | 27.0 | 5 | 5 | 2.3 | 2.6 |
| Mean | 84.0 | 85.0 | 7.6 | 7.3 | 7.1 | 6.6 | 110.0 | 107.4 | 20.3 | 22.3 | 140.6 | 146.0 | 95.0 | 89.6 | 29.5 | 30.9 | 4.9 | 4.1 | 2.3 | 2.6 | |
| CV (%) | 1.42 | 1.43 | 13.31 | 17.05 | 14.24 | 17.4 | 6.99 | 1.8 | 4.26 | 6.03 | 1.68 | 5.66 | 1.73 | 2.92 | 4.13 | 5.87 | 16.71 | 16.39 | 1.35 | 11.34 | |
| LSD (5%) | 2.38 | 2.86 | 1.99 | 2.43 | 2.04 | 2.28 | 15.48 | 3.85 | 1.74 | 2.71 | 4.74 | 16.62 | 3.32 | 5.27 | 2.46 | 3.65 | 0.60 | 0.55 | 0.04 | 0.60 | |
| Traits | DFL | NT | RYMVD | NPP | PH | PL | NGP | PFG | TGW | GY |
|---|---|---|---|---|---|---|---|---|---|---|
| DFL | 1 | −0.01 | −0.27 * | 0.05 | −0.02 | 0.22 | 0.41 ** | 0.13 | −0.07 | 0.13 |
| NT | 0.19 | 1 | −0.05 | 0.83 *** | −0.26 | 0.03 | 0.39 ** | 0.25 | 0.04 | 0.12 |
| RYMVD | −0.29 | 0.24 | 1 | 0.04 | 0.05 | 0.23 | −0.24 | −0.14 | 0.01 | −0.40 ** |
| NPP | 0.20 | 0.85 *** | 0.21 | 1 | −0.19 | 0.16 | −0.32 * | 0.18 | −0.10 | 0.44 ** |
| PH | 0.03 | −0.21 | 0.21 | −0.27 | 1 | 0.07 | 0.29 * | 0.22 | 0.36 * | 0.05 |
| PL | 0.23 | 0.29 * | −0.26 | 0.31 * | 0.09 | 1 | −0.06 | −0.15 | 0.07 | 0.34 * |
| NGP | 0.47 ** | 0.31 * | −0.42 | −0.29 * | 0.33 * | −0.02 | 1 | −0.16 | 0.29 * | 0.28 * |
| PFG | −0.23 | −0.14 | −0.27 | −0.15 | 0.40 ** | −0.22 | −0.17 | 1 | 0.31 * | 0.36 * |
| TGW | −0.09 | −0.09 | −0.14 | −0.3 | 0.28 * | 0.06 | 0.46 ** | 0.41 ** | 1 | 0.48 ** |
| GY | 0.12 | 0.25 | −0.33 * | 0.29 * | 0.01 | 0.28 * | 0.54 *** | 0.34 * | 0.43 ** | 1 |
| Traits | DFL | NT | RYMVD | NPP | PH | PL | NGP | PFG | TGW | GY |
|---|---|---|---|---|---|---|---|---|---|---|
| DFL | 1 | −0.01 | −0.27 | 0.05 | −0.02 | 0.22 | 0.43 ** | 0.12 | −0.07 | 0.13 |
| NT | 1 | −0.05 | 0.83 *** | −0.31 * | 0.03 | 0.36 * | 0.25 | 0.04 | 0.12 | |
| RYMVD | 1 | 0.04 | 0.05 | 0.23 | −0.34 * | −0.14 | −0.01 | −0.37 * | ||
| NPP | 1 | −0.29 | 0.16 | −0.32 * | 0.28 * | −0.10 | 0.32 * | |||
| PH | 1 | 0.07 | 0.33 * | 0.22 | 0.34 * | 0.05 | ||||
| PL | 1 | −0.06 | −0.15 | 0.07 | 0.33 * | |||||
| NGP | 1 | −0.16 | 0.32 * | 0.45 ** | ||||||
| PFG | 1 | 0.37 * | 0.38 * | |||||||
| TGW | 1 | 0.47 ** | ||||||||
| GY | 1 |
| Trait | Ifakara | Mkindo | Across Locations | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC4 | PC5 | PC1 | PC2 | PC3 | PC4 | PC1 | PC2 | PC3 | PC4 | PC5 | |
| Eigen-values | 2.34 | 1.81 | 1.49 | 1.18 | 1.01 | 2.74 | 1.82 | 1.43 | 1.09 | 2.43 | 2.02 | 1.42 | 1.12 | 1.01 |
| Proportion variance (%) | 23.44 | 18.06 | 14.89 | 11.84 | 10.09 | 27.4 | 18.2 | 14.33 | 10.86 | 24.3 | 20.15 | 14.16 | 11.19 | 10.08 |
| Cumulative variance (%) | 23.44 | 41.5 | 56.39 | 68.22 | 78.32 | 27.4 | 45.5 | 59.92 | 70.78 | 24.3 | 44.45 | 58.61 | 69.8 | 79.88 |
| DFL | −0.17 | 0.71 | −0.2 | 0.41 | −0.17 | 0.49 | 0.52 | 0.01 | −0.04 | 0.32 | 0.62 | −0.08 | 0.36 | 0.26 |
| NT | 0.87 | 0.26 | 0.21 | 0.14 | −0.13 | 0.77 | −0.4 | 0.21 | 0.17 | 0.86 | −0.3 | 0.17 | 0.17 | 0.13 |
| RYMVD | −0.24 | 0.62 | −0.38 | 0.02 | 0.1 | 0.54 | 0.53 | −0.14 | 0.16 | 0.29 | 0.66 | −0.26 | 0.07 | −0.14 |
| NPP | 0.82 | 0.38 | 0.12 | 0.2 | 0.02 | 0.81 | −0.4 | 0.12 | 0.19 | 0.88 | −0.23 | 0.08 | 0.27 | 0.08 |
| PH | −0.44 | −0.21 | 0.41 | 0.56 | 0.32 | −0.37 | 0.5 | 0.43 | 0.49 | −0.51 | 0.24 | 0.43 | 0.52 | −0.18 |
| PL | 0.04 | 0.53 | 0.24 | −0.09 | 0.74 | 0.41 | 0.03 | 0.41 | 0.47 | 0.39 | 0.25 | 0.3 | 0.14 | −0.76 |
| NGP | −0.71 | 0.4 | −0.02 | 0.19 | −0.3 | 0.25 | 0.82 | 0.06 | −0.18 | −0.12 | 0.83 | 0.01 | 0.08 | 0.34 |
| PFG | 0.28 | −0.36 | −0.22 | 0.75 | −0.07 | −0.61 | −0.17 | 0.29 | 0.38 | −0.34 | −0.47 | 0.08 | 0.68 | 0.24 |
| TGW | −0.24 | −0.04 | 0.77 | 0.12 | −0.01 | −0.31 | 0.04 | 0.75 | −0.3 | −0.25 | 0.06 | 0.79 | −0.15 | 0.05 |
| GY | −0.02 | 0.31 | 0.61 | −0.16 | −0.47 | 0.35 | −0.08 | 0.6 | −0.52 | 0.34 | 0.15 | 0.63 | −0.32 | 0.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suvi, W.T.; Shimelis, H.; Laing, M.; Mathew, I.; Shayanowako, A.I.T. Variation among Tanzania Rice Germplasm Collections Based on Agronomic Traits and Resistance to Rice Yellow Mottle Virus. Agronomy 2021, 11, 391. https://doi.org/10.3390/agronomy11020391
Suvi WT, Shimelis H, Laing M, Mathew I, Shayanowako AIT. Variation among Tanzania Rice Germplasm Collections Based on Agronomic Traits and Resistance to Rice Yellow Mottle Virus. Agronomy. 2021; 11(2):391. https://doi.org/10.3390/agronomy11020391
Chicago/Turabian StyleSuvi, William Titus, Hussein Shimelis, Mark Laing, Isack Mathew, and Admire I. T. Shayanowako. 2021. "Variation among Tanzania Rice Germplasm Collections Based on Agronomic Traits and Resistance to Rice Yellow Mottle Virus" Agronomy 11, no. 2: 391. https://doi.org/10.3390/agronomy11020391
APA StyleSuvi, W. T., Shimelis, H., Laing, M., Mathew, I., & Shayanowako, A. I. T. (2021). Variation among Tanzania Rice Germplasm Collections Based on Agronomic Traits and Resistance to Rice Yellow Mottle Virus. Agronomy, 11(2), 391. https://doi.org/10.3390/agronomy11020391