
Can Soil pH Correction Reduce the Animal Supplementation Needs in the Critical Autumn Period in Mediterranean Montado Ecosystem?

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
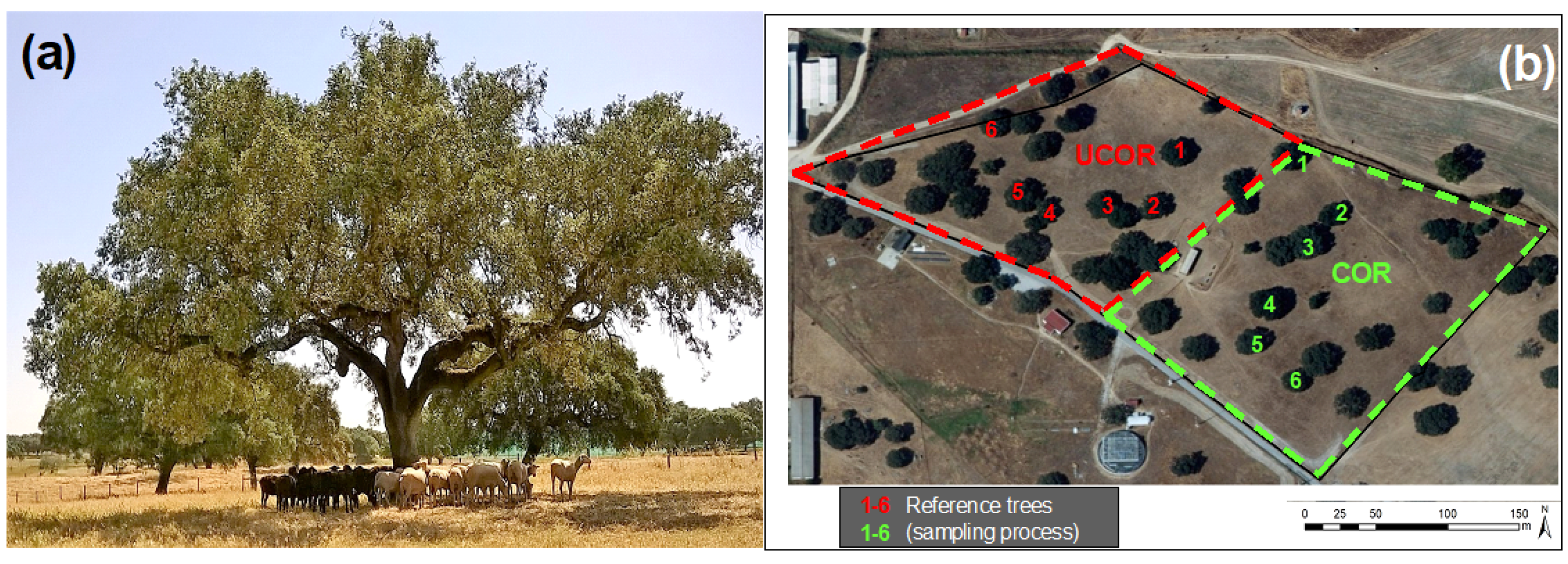
2.1. Experimental Field and Sampling Scheme
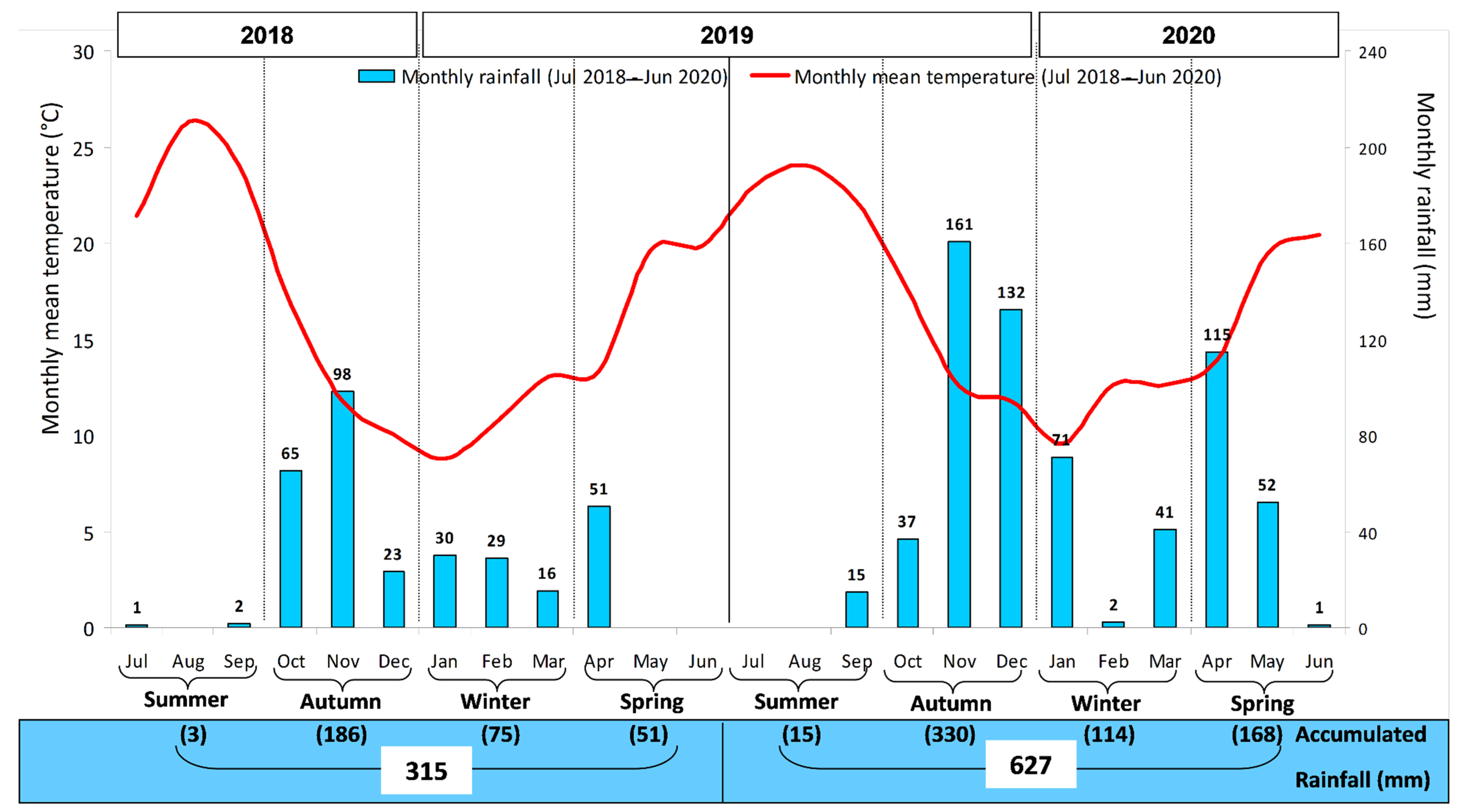
2.2. Characterization of the Climate
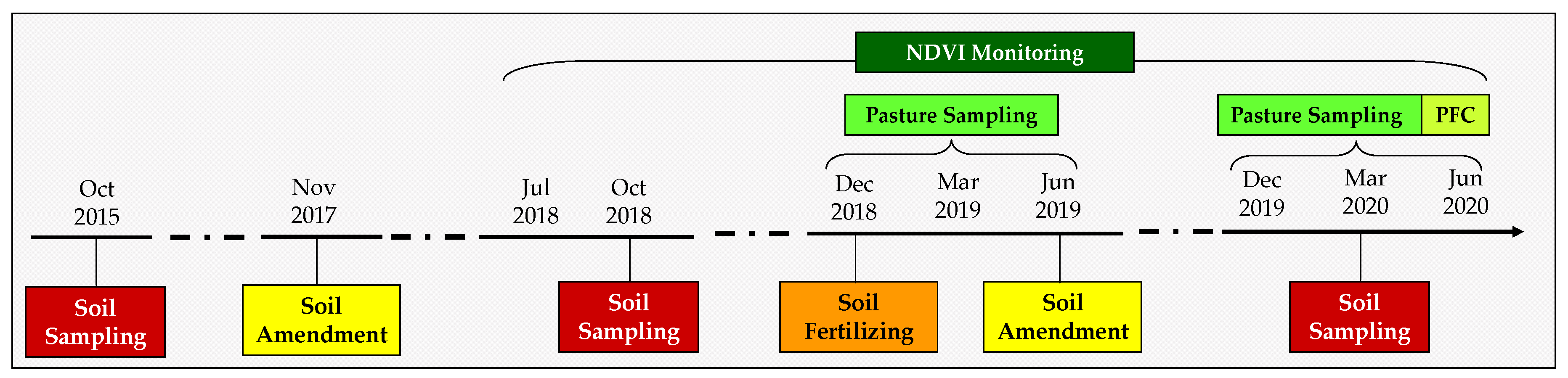
2.3. Chronological Sequence of the Interventions and Measurements in the Experimental Field
2.4. Soil Sample Collection and Analysis
2.5. Pasture Samples Collection and Analysis
2.6. Statistical Analysis of the Data
3. Results
3.1. Temporal and Spatial Variability Pattern of the Soil Parameters
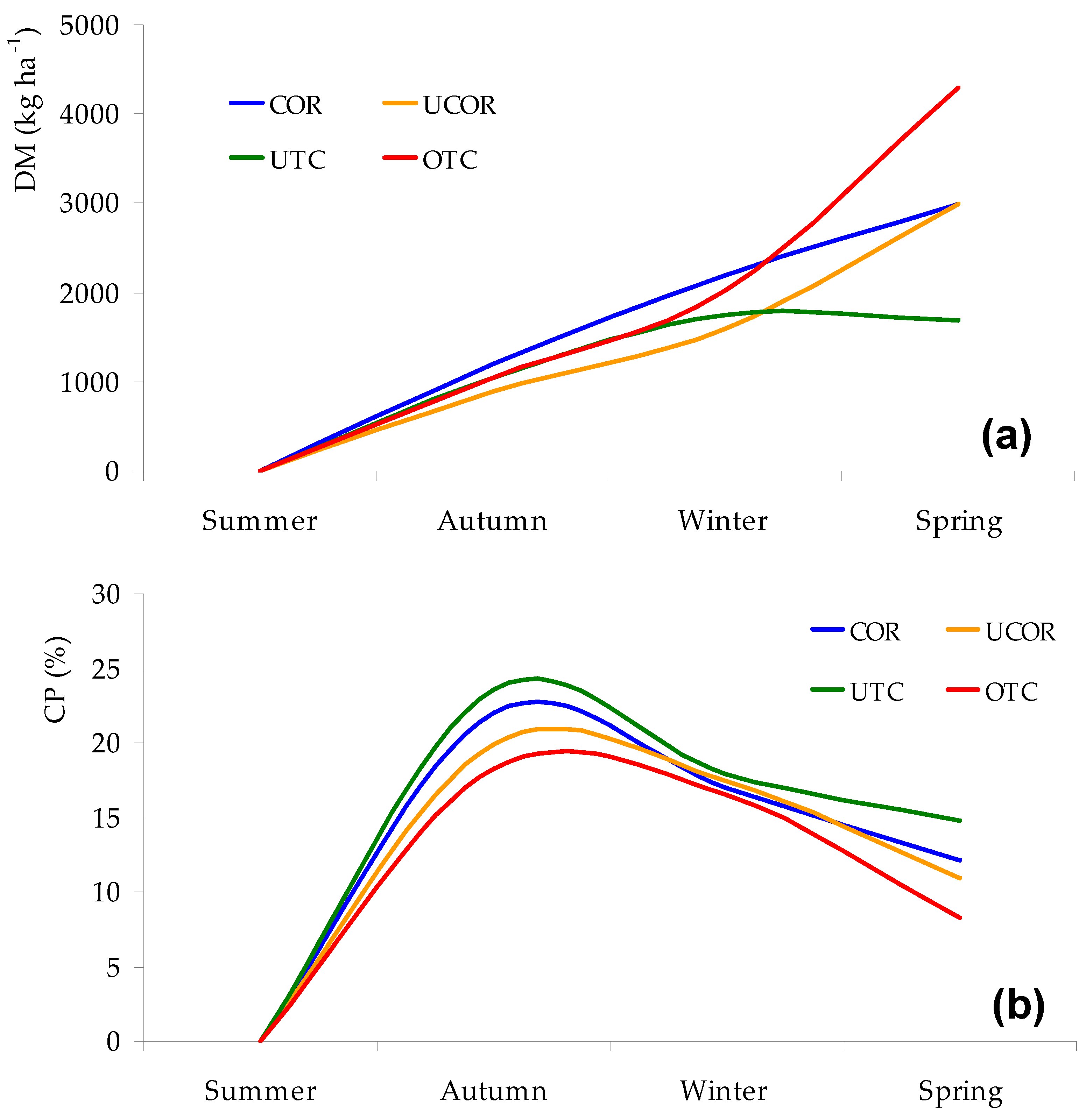
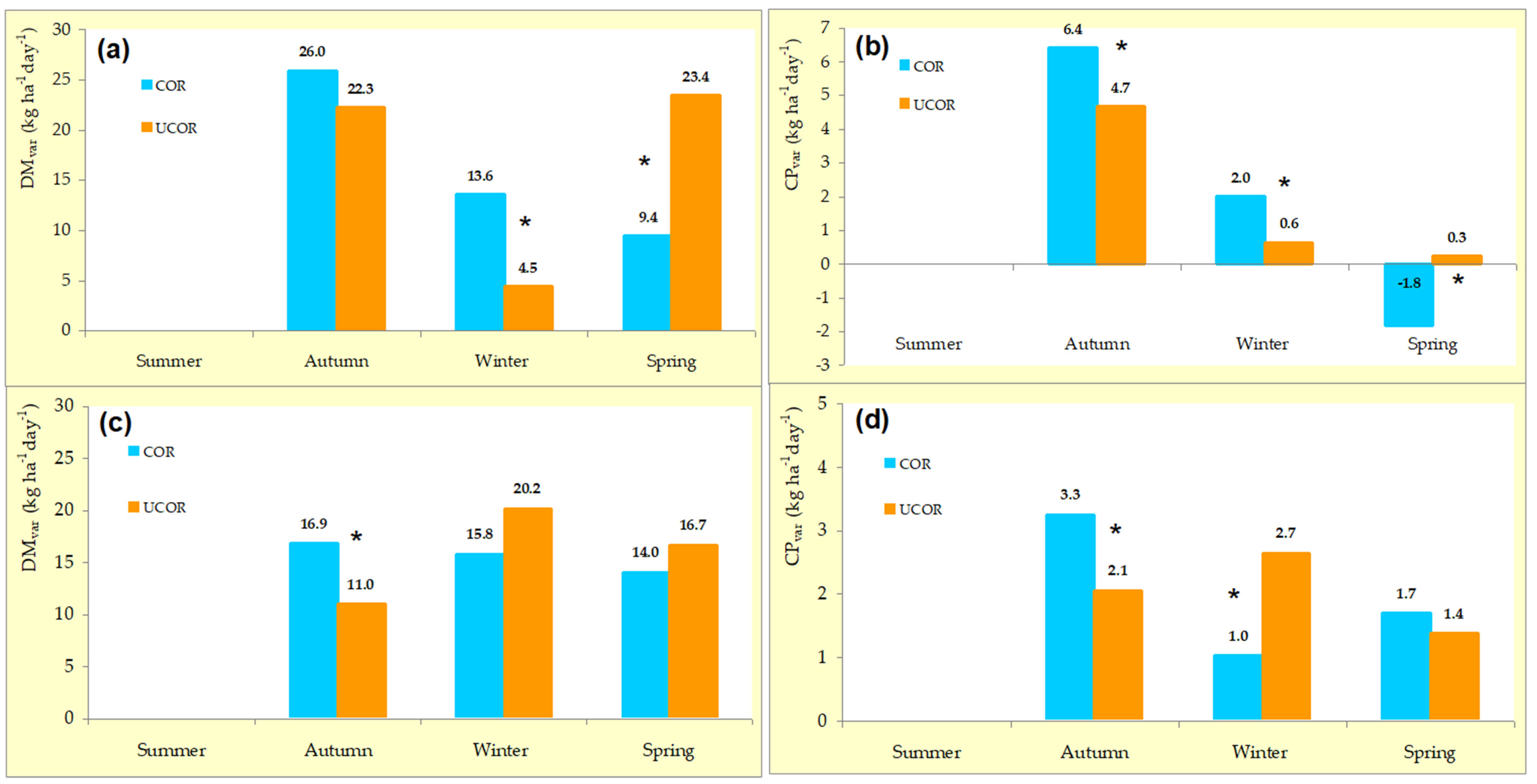
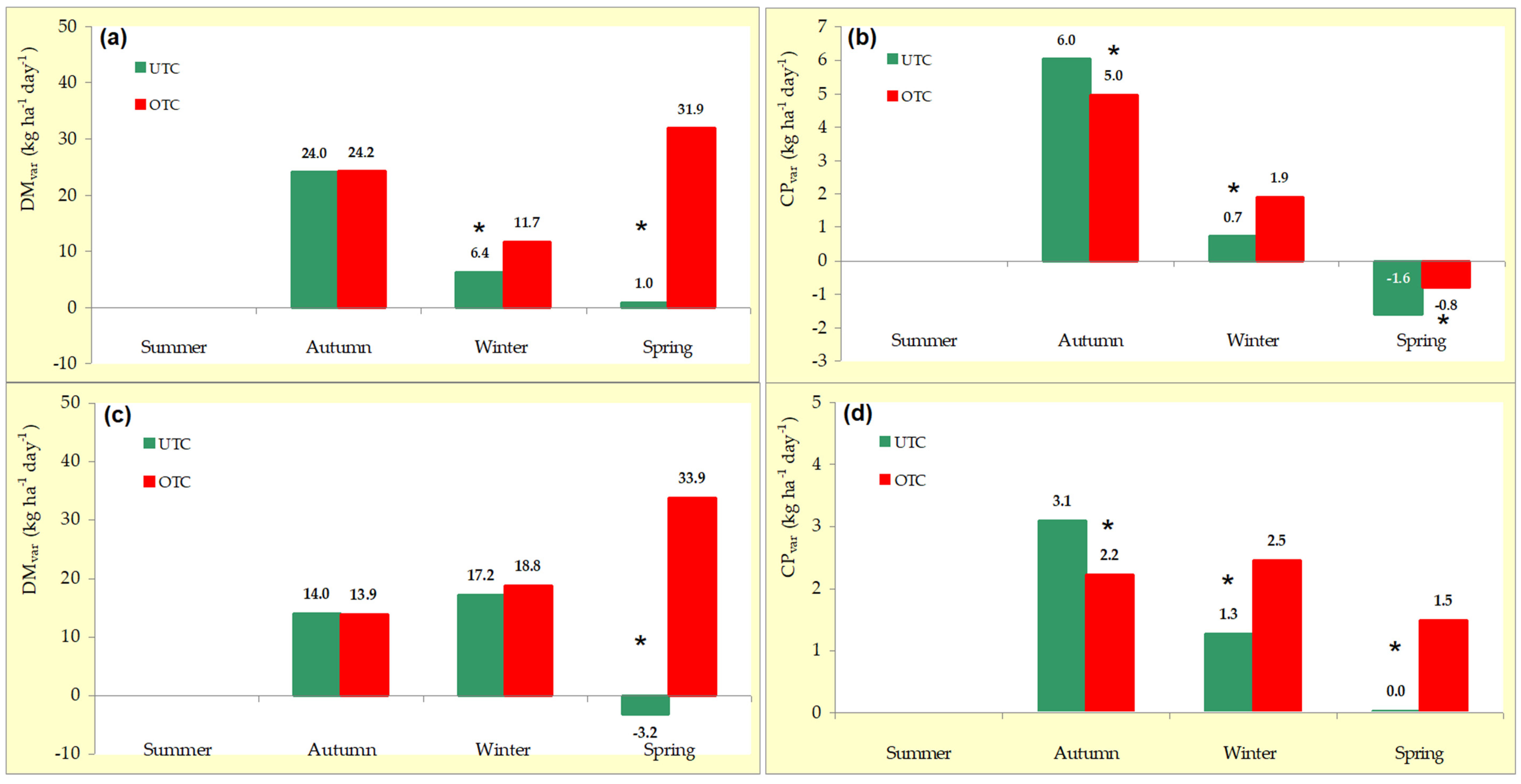
3.2. Variability Pattern of Pasture Productivity and Quality
3.3. Temporal Pattern of Evolution of Normalized Difference Vegetation Index (NDVI)
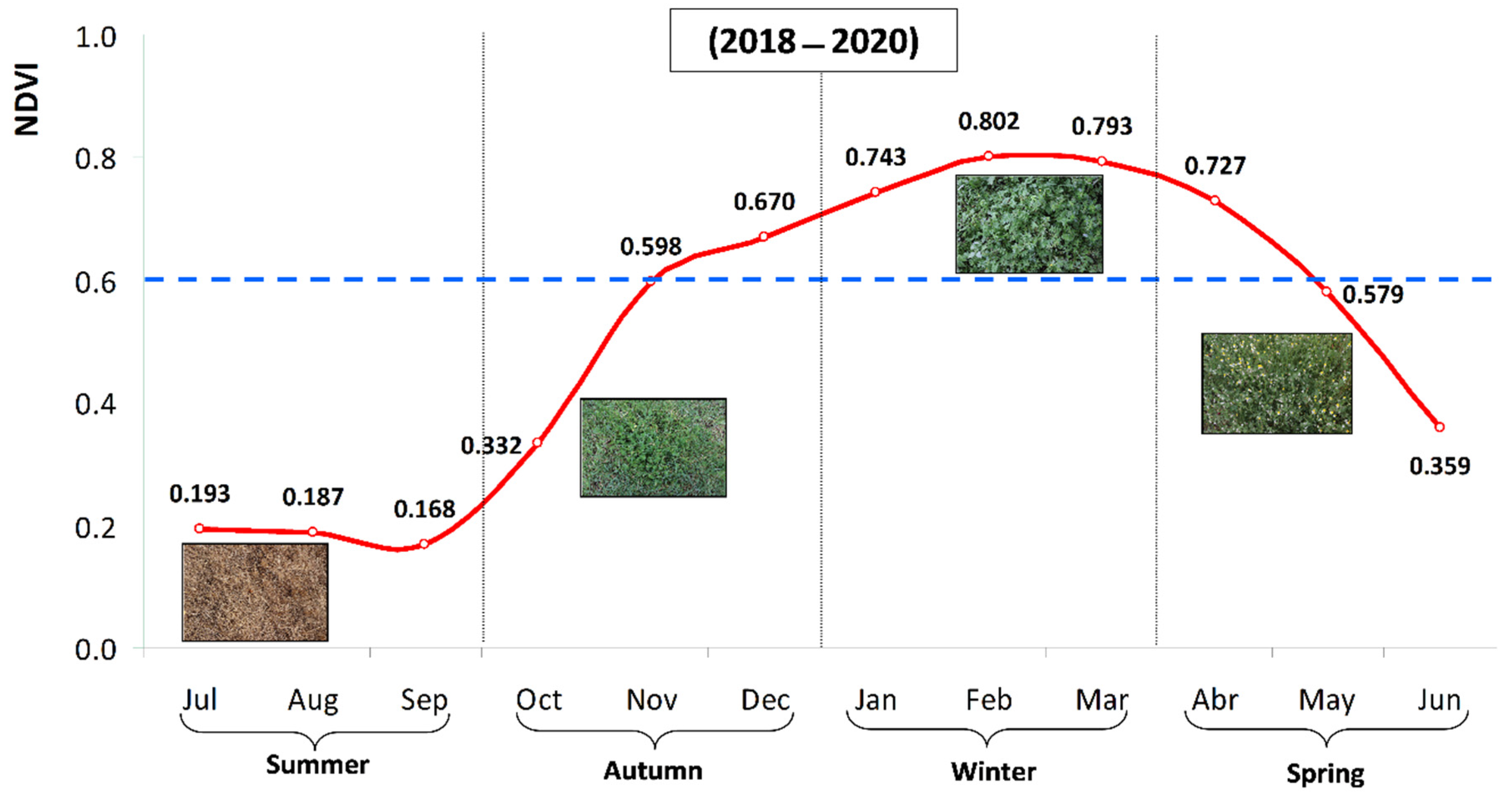
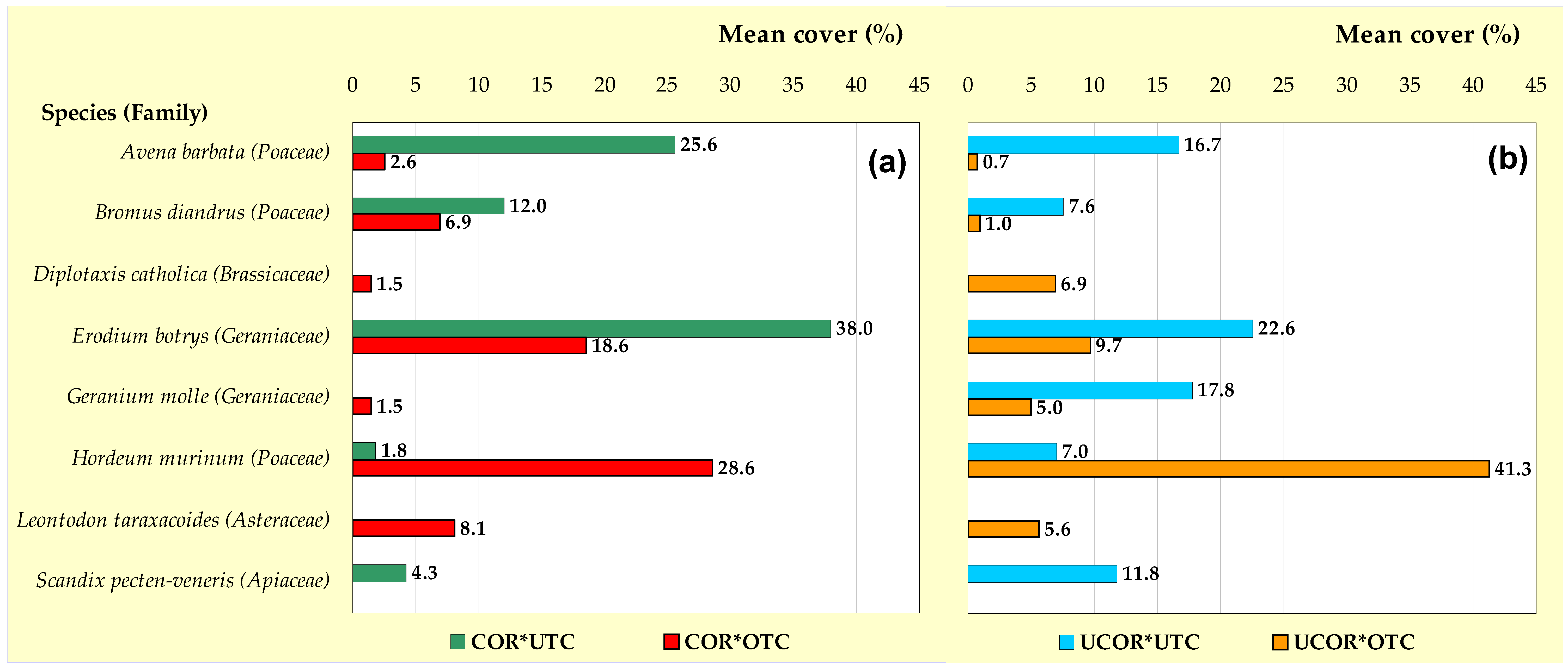
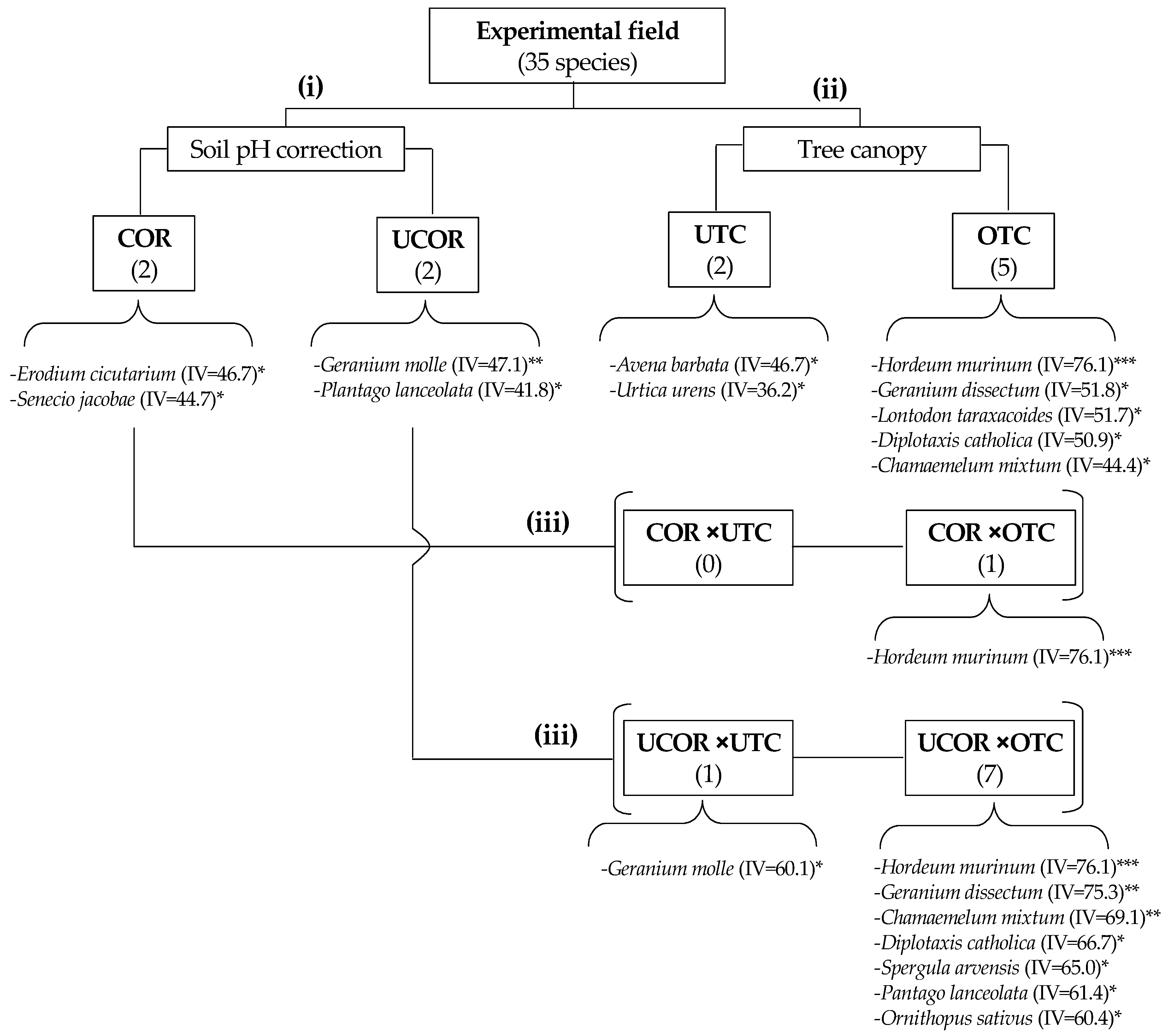
3.4. Spatial Variability of Pasture Floristic Composition (PFC)
4. Discussion
4.1. Variability Pattern of Pasture Productivity and Quality
4.2. Variability of Spatial Pattern of Pasture Floristic Composition (PFC)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Serrano, J.; Sales-Baptista, E.; Shahidian, S.; Marques da Silva, J.; Ferraz de Oliveira, I.; Lopes de Castro, J.; Pereira, A.; Cancela d’Abreu, M.; Carvalho, M. Proximal sensors for monitoring seasonal changes of feeding sites selected by grazing ewes. Agroforest. Syst. 2018, 95, 55–69. [Google Scholar] [CrossRef]
- Hanson, J.; Ellis, R.H. Progress and Challenges in Ex Situ Conservation of forage germplasm: Grasses, herbaceous legumes and fodder trees. Plants 2020, 9, 446. [Google Scholar] [CrossRef] [Green Version]
- Santos, S.K.; Falbo, M.K.; Sandini, I.E.; Pacentchuk, F.; Neumann, M.; Garbossa, G. Concentrate supplementation strategies in ryegrass pasture for productive performance in lambs. Span. J. Agric. Res. 2018, 16, 1–6. [Google Scholar] [CrossRef]
- Polo, J.L.M.; Bellido, I.G.; Rodriguez, M.E.S. Plant production and nutritive quality of savannah-like grasslands (dehesas) in semi-arid zones of the province of Salamanca. Span. J. Agric. Res. 2003, 1, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Scocco, P.; Piermarteri, K.; Malfatti, A.; Tardella, F.M.; Catorci, A. Effects of summer rainfall variations on sheep body state and farming sustainability in sub-Mediterranean pastoral systems. Span. J. Agric. Res. 2016, 14, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Perera, R.S.; Cullen, B.R.; Eckard, R.J. Growth and physiological responses of temperate pasture species to consecutive heat and drought stresses. Plants 2019, 8, 227. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, F.A.; Vázquez, M.; Camuñez, J.A.; Castel, J.M.; Mena, Y. Characterization and challenges of livestock farming in Mediterranean protected mountain areas. Span. J. Agric. Res. 2020, 18, 1–14. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014; International Soil Classification System for Naming Soils and Creating Legends for Soil Maps. Update 2015; World Soil Resources Report 106; Food and Agriculture Organization (FAO): Rome, Italy, 2015; 188p. [Google Scholar]
- Efe Serrano, J. Pastures in Alentejo: Technical Basis for Characterization, Grazing and Improvement; Universidade de Évora—ICAM, Ed.; Gráfica Eborense: Évora, Portugal, 2006; pp. 165–178. (In Portuguese) [Google Scholar]
- Carvalho, M.; Goss, M.J.; Teixeira, D. Manganese toxicity in Portuguese Cambisols derived from granitic rocks: Causes, limitations of soil analyses and possible solutions. Rev. Cienc. Agrar. 2015, 38, 518–527. [Google Scholar] [CrossRef]
- Paço, A.; da-Silva, J.R.; Torres, D.P.; Glick, B.R.; Brígido, C. Exogenous ACC Deaminase is key to improving the performance of pasture legume-rhizobial symbioses in the presence of a high manganese concentration. Plants 2020, 9, 1630. [Google Scholar] [CrossRef]
- Serrano, J.; Shahidian, S.; Marques da Silva, J.; Moral, F.; Carvajal-Ramirez, F.; Carreira, E.; Pereira, A.; Carvalho, M. Evaluation of the effect of dolomitic lime application on pastures—Case study in the Montado Mediterranean ecosystem. Sustainability 2020, 12, 3758. [Google Scholar] [CrossRef]
- Demarchi, L.O.; Scudeller, V.V.; Moura, L.C.; Dias-Terceiro, R.G.; Lopes, A.; Wittmann, F.K.; Piedade, M.T.F. Floristic composition, structure and soil-vegetation relations in three white-sand soil patches in central Amazonia. Acta Amaz. 2018, 48, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Serrano, J.; Shahidian, S.; da Silva, J.M.; Carvalho, M. A holistic approach to the evaluation of the Montado ecosystem using proximal sensors. Sensors 2018, 18, 570. [Google Scholar] [CrossRef] [Green Version]
- Benavides, R.; Douglas, G.B.; Osoro, K. Silvopastoralism in New Zealand: Review of effects of evergreen and deciduous trees on pasture dynamics. Agrofor. Syst. 2009, 76, 327–350. [Google Scholar] [CrossRef]
- Graß, R.; Malec, S.; Wachendorf, M. Biomass performance and competition effects in an established temperate agroforestry system of willow and grassland—Results of the 2nd rotation. Agronomy 2020, 10, 1819. [Google Scholar] [CrossRef]
- Ramírez, N.; Dezzeo, N.; Chacón, N. Floristic composition, plant species abundance, and soil properties of montane savannas in the Gran Sabana, Venezuela. Flora 2007, 202, 316–327. [Google Scholar] [CrossRef]
- Irume, M.V.; Morais, M.L.C.S.; Zartman, C.E.; Amaral, I.L. Floristic composition and community structure of epiphytic angiosperms in a terra firme forest in central Amazonia. Acta Bot. Bras. 2013, 27, 378–393. [Google Scholar] [CrossRef] [Green Version]
- Serrano, J.; Shahidian, S.; da Silva, J.M.; Sales-Baptista, E.; de Oliveira, I.F.; de Castro, J.L.; Pereira, A.; d’Abreu, M.C.; Machado, E.; Carvalho, M. Tree influence on soil and pasture: Contribution of proximal sensing to pasture productivity and quality estimation in montado ecosystems. Int. J. Remote Sens. 2018, 39, 4801–4829. [Google Scholar] [CrossRef]
- Egner, H.; Riehm, H.; Domingo, W.R. Untersuchungen über die chemische Bodenanalyse als Grudlage für die Beurteilung des Nahrstoffzunstandes der Boden. II: Chemische extraktion methoden zur phosphor- und kalium-bestimmung. K. Lantbrhogsk. Annlr 1960, 26, 199–216. (In German) [Google Scholar]
- AOAC. Official Method of Analysis of AOAC International, 18th ed.; AOAC International: Arlington, AT, USA, 2005. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie, 3rd ed.; Grundzüge der Vegetationskunde; Springer: Vienna, Austria; New York, NY, USA, 1964. [Google Scholar]
- Shore, A. DESeq and Indicator Species Analysis R script. Figshare. Software. 2020. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- McGeoch, M.A.; Chown, S.L. Scaling up the value of bioindicators. Trends Ecol. Evol. 1998, 13, 46–47. [Google Scholar] [CrossRef]
- Bakker, J.D. Increasing the utility of Indicator Species Analysis. J. App. Ecol. 2008, 45, 1829–1835. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Sheep, 6th ed.; National Academy Press: Washington, DC, USA, 1985; Volume 5. [Google Scholar]
- Pinto-Correia, T.; Ribeiro, N.; Sá-Sousa, P. Introducing the montado, the cork and holm oak agroforestry system of Southern Portugal. Agrofor. Syst. 2011, 82, 99. [Google Scholar] [CrossRef] [Green Version]
- Sade, H.; Meriga, B.; Surapu, V.; Gadi, J.; Sunita, M.S.; Suravajhala, P.; Kavi Kishor, P.B. Toxicity and tolerance of aluminum in plants: Tailoring plants to suit to acid soils. Biometals 2016, 29, 187–210. [Google Scholar] [CrossRef]
- Kochian, L.V.; Piñeros, M.A.; Hoekenga, O.A. The physiology, genetics and molecular biology of plant aluminum resistance and toxicity. Plant Soil 2005, 274, 175–195. [Google Scholar] [CrossRef]
- Alho, L.; Carvalho, M.; Brito, I.; Goss, M.J. The effect of arbuscular mycorrhiza fungal propagules on the growth of subterranean clover (Trifolium subterraneum L.) under Mn toxicity in ex situ experiments. Soil Use Manag. 2015, 31, 337–344. [Google Scholar] [CrossRef]
- Luna, I.M.; Fernández-Quintanilla, C.; Dorado, J. Is Pasture Cropping a Valid Weed Management Tool? Plants 2020, 9, 135. [Google Scholar] [CrossRef] [Green Version]
- Serrano, J.; Shahidian, S.; da Silva, J.M. Monitoring seasonal pasture quality degradation in the Mediterranean montado ecosystem: Proximal versus remote sensing. Water 2018, 10, 1422. [Google Scholar] [CrossRef] [Green Version]
- Halim, N.; Abdullah, R.; Karsani, S.; Osman, N.; Panhwar, Q.; Ishak, C. Influence of soil amendments on the growth and yield of rice in acidic soil. Agronomy 2018, 8, 165. [Google Scholar] [CrossRef] [Green Version]
- Li, G.D.; Conyers, M.K.; Helyar, K.R.; Lisle, C.J.; Poile, G.J.; Cullis, B.R. Long-term surface application of lime ameliorates subsurface soil acidity in the mixed farming zone of south-eastern Australia. Geoderma 2019, 338, 236–246. [Google Scholar] [CrossRef]
- Rayburn, E.B.; Griggs, T.C. Light interception and the growth of pastures under ideal and stressful growing conditions on the Allegheny Plateau. Plants 2020, 9, 734. [Google Scholar] [CrossRef]
- Hussain, Z.; Kemp, P.D.; Horne, D.J. Pasture production under densely planted young willow and poplar in a silvopastoral system. Agrofor. Syst. 2009, 76, 351–362. [Google Scholar] [CrossRef]
- Marcos, G.M.; Obrador, J.J.; Garcia, E.; Cubera, E.; Montero, M.J.; Pulido, F.; Dupraz, C. Driving competitive and facilitative interactions in oak dehesas through management practices. Agrofor. Syst. 2007, 70, 25–40. [Google Scholar] [CrossRef]
- Sousa, L.F.; Maurício, R.M.; Moreira, G.R.; Gonçalves, L.C.; Borges, I.; Pereira, L.G.R. Nutritional evaluation of “Braquiaro” grass in association with “Aroeira” trees in a silvopastoral system. Agrofor. Syst. 2010, 79, 189–199. [Google Scholar] [CrossRef]
- Avdiu, B.; Aliu, S.; Fetahu, S.; Zeka, D.; Rusinovci, I. The floristic composition of the natural pastures in Massive of Novoberba. Agric. For. 2018, 64, 235–241. [Google Scholar]
- Sales-Baptista, E.; Oliveira, I.F.; Santos, M.B.; Castro, J.A.; Pereira, A.; Rafael, J.; Serrano, J. Tecnologia GNSS de baixo custo na monitorizacão de ovinos em pastoreio. Rev. Cienc. Agrar. 2016, 39, 251–260. (In Portuguese) [Google Scholar] [CrossRef]
- Solefack, M.C.M.; Fedoung, E.F.; Temgoua, L.F. Factors determining floristic composition and functional diversity of plant communities of Mount Oku forests, Cameroon. J. Asia Pac. Biodiver. 2018, 11, 284–293. [Google Scholar] [CrossRef]
- Gómez-Rey, M.X.; Garcês, A.; Madeira, M. Soil organic-C accumulation and N availability under improved pastures established in Mediterranean Oak Woodlands. Soil Use Manag. 2012, 28, 497–507. [Google Scholar] [CrossRef]
- Guevara-Escobar, A.; Kemp, P.D.; Mackay, A.D.; Hodgson, J. Pasture production and composition under poplar in a hill environment in New Zealand. Agrofor. Syst. 2007, 3, 199–213. [Google Scholar] [CrossRef]
- Ferraz de Oliveira, M.I.; Lamy, E.; Bugalho, M.N.; Vaz, M.; Pinheiro, C.; Cancela d’Abreu, M.; Capela e Silva, F.; Sales-Baptista, E. Assessing foraging strategies of herbivores in Mediterranean oak woodlands: A review of key issues and selected methodologies. Agrofor. Syst. 2013, 87, 1421–1437. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Parameters | GLOBAL | COR | UCOR | Prob. | UTC | OTC | Prob. | Ref. |
|---|---|---|---|---|---|---|---|---|
| October 2015 | - | - | - | [19] | ||||
| pH | 5.4 ± 0.3 | 5.4 ± 0.4 | 5.3 ± 0.2 | ns | ||||
| Mg (mg kg−1) | 95.6 ± 43.7 | 115.0 ± 38.8 | 76.3 ± 40.9 | 0.0503 | ||||
| Mn (mg kg−1) | 76.4 ± 44.9 | 100.0 ± 45.7 | 52.8 ± 30.1 | 0.0131 | ||||
| October 2018 | [12] | |||||||
| pH | 5.4 ± 0.2 | 5.6 ± 0.2 | 5.3 ± 0.2 | 0.0193 | 5.5 ± 0.2 | 5.3 ± 0.2 | 0.0232 | |
| Mg (mg kg−1) | 78.1 ± 33.0 | 82.9±32.1 | 73.3±34.6 | ns | 84.2 ± 21.2 | 72.1 ± 41.8 | ns | |
| Mn (mg kg−1) | 50.2 ± 29.7 | 33.6±16.1 | 66.8±31.4 | 0 | 38.4 ± 23.4 | 62.1 ± 31.5 | 0 | |
| March 2020 | - | |||||||
| pH | 5.7 ± 0.3 | 5.8 ± 0.4 | 5.6 ± 0.2 | 0.0225 | 5.8 ± 0.2 | 5.6 ± 0.4 | 0.0331 | |
| Mg (mg kg−1) | 94.8 ± 29.2 | 108.1 ± 22.3 | 71.3 ± 16.8 | 0.0442 | 102.4 ± 25.8 | 79.8 ± 19.3 | 0.0215 | |
| Mn (mg kg−1) | 40.1 ± 17.3 | 32.5 ± 13.6 | 51.6 ± 24.6 | 0.0182 | 47.6 ± 22.3 | 34.3 ± 16.1 | 0.0441 |
| Pasture Parameters | COR | UCOR | Prob. | UTC | OTC | Prob. | COR × UTC | COR × OTC | UCOR × UTC | UCOR × OTC |
|---|---|---|---|---|---|---|---|---|---|---|
| GM (t ha−1) | ||||||||||
| 2018/2019, Autumn | 7.9 ± 3.4 | 6.8 ± 2.4 | ns | 6.9 ± 3.5 | 7.9 ± 2.9 | ns | 7.9a | 7.8a | 5.6b | 8.0a |
| Winter | 16.3 ± 9.8 | 8.7 ± 4.9 | 0.0486 | 9.2 ± 8.1 | 15.8 ± 7.8 | 0.0205 | 13.7a | 18.8a | 4.6b | 12.8a |
| Spring | 4.9 ± 2.6 | 4.7 ± 2.8 | ns | 2.7 ± 1.4 | 6.9 ± 1.9 | 0.0001 | 3.3b | 6.6a | 2.2b | 7.2a |
| 2019/2020, Autumn | 8.3 ± 3.4 | 5.1 ± 2.2 | 0.0277 | 7.6 ± 3.8 | 5.8 ± 2.4 | 0.0959 | 9.5a | 7.2b | 5.7bc | 4.4c |
| Winter | 14.4 ± 6.7 | 13.0 ± 5.5 | ns | 12.5 ± 5.3 | 14.8 ± 6.7 | ns | 14.1a | 14.7a | 10.9b | 15.0a |
| Spring | 12.7 ± 4.6 | 14.7 ± 8.7 | ns | 9.8 ± 2.5 | 17.8 ± 7.4 | 0.0010 | 9.3b | 16.2a | 9.9b | 19.4a |
| DM (t ha−1) | ||||||||||
| 2018/2019, Autumn | 1.1 ± 0.6 | 0.9 ± 0.2 | ns | 1.0 ± 0.6 | 1.0 ± 0.3 | ns | 1.2a | 1.0a | 0.9a | 1.1a |
| Winter | 2.3 ± 0.9 | 1.4 ± 0.4 | 0.0198 | 1.6 ± 0.8 | 2.1 ± 0.8 | 0.0307 | 2.1ab | 2.6a | 1.1c | 1.6bc |
| Spring | 3.0 ± 1.8 | 3.1 ± 1.9 | ns | 1.7 ± 0.9 | 4.5 ± 1.3 | 0.0002 | 1.8b | 4.2a | 1.5b | 4.7a |
| 2019/2020, Autumn | 1.3 ± 0.4 | 0.8 ± 0.4 | 0.0226 | 1.1 ± 0.5 | 1.0 ± 0.4 | ns | 1.2a | 1.3a | 0.9b | 0.8b |
| Winter | 2.1 ± 1.0 | 1.8 ± 0.4 | ns | 1.9 ± 0.7 | 2.0 ± 0.9 | ns | 2.0a | 2.1a | 1.8b | 1.9b |
| Spring | 2.9 ± 1.7 | 2.9 ± 1.3 | ns | 1.7 ± 0.5 | 4.1 ± 1.0 | 0.0000 | 1.5b | 4.4a | 2.0b | 3.8a |
| CP (%) | ||||||||||
| 2018/2019, Autumn | 24.7 ± 8.6 | 21.0 ± 3.2 | ns | 25.1 ± 8.0 | 20.5 ± 3.9 | 0.0708 | 28.5a | 20.9b | 21.8ab | 20.2b |
| Winter | 19.6 ± 6.2 | 19.1 ± 4.8 | ns | 20.3 ± 6.1 | 18.4 ± 4.7 | ns | 23.5a | 15.7b | 17.1b | 21.2ab |
| Spring | 10.5 ± 5.1 | 8.9 ± 1.9 | ns | 12.2 ± 4.0 | 7.2 ± 1.3 | 0.0004 | 14.1a | 6.8c | 10.2b | 7.6bc |
| 2019/2020, Autumn | 19.3 ± 4.8 | 18.8 ± 4.1 | ns | 22.1 ± 3.4 | 16.0 ± 2.8 | 0.0015 | 22.5a | 16.0b | 21.8a | 15.9b |
| Winter | 14.4 ± 4.0 | 15.7 ± 3.3 | ns | 15.53 ± 4.2 | 14.6 ± 3.2 | ns | 15.6a | 13.2b | 15.5a | 15.9a |
| Spring | 13.7 ± 6.2 | 13.0 ± 3.9 | ns | 17.4 ± 3.5 | 9.3 ± 2.5 | 0.0000 | 19.1a | 8.2b | 15.7a | 10.4b |
| NDF (%) | ||||||||||
| 2018/2019, Autumn | 48.5 ± 9.7 | 50.4 ± 6.9 | ns | 52.2 ± 6.6 | 46.7 ± 9.2 | ns | 50.7a | 46.3a | 53.8a | 47.1a |
| Winter | 43.1 ± 6.9 | 48.7 ± 9.0 | 0.0424 | 50.6 ± 9.1 | 41.2 ± 3.8 | 0.0014 | 44.7b | 41.5b | 56.7a | 40.8b |
| Spring | 63.6 ± 5.0 | 65.6 ± 2.8 | ns | 62.7 ± 4.0 | 66.5 ± 3.4 | 0.0457 | 60.6b | 66.7a | 64.9ab | 66.3a |
| 2019/2020, Autumn | 36.3 ± 8.8 | 39.0 ± 6.8 | ns | 35.5 ± 7.4 | 39.9 ± 8.0 | 0.0976 | 34.8b | 37.9b | 36.2b | 41.9a |
| Winter | 41.6 ± 6.3 | 48.7 ± 4.3 | 0.0126 | 43.1 ± 5.0 | 47.3 ± 7.2 | 0.0452 | 40.2c | 43.1bc | 46.0b | 51.5a |
| Spring | 57.3 ± 6.9 | 58.7 ± 6.2 | ns | 53.9 ± 5.7 | 62.1 ± 4.3 | 0.0002 | 51.5b | 63.2a | 56.3ab | 61.1a |
| FAMILY | Botanical Species | COR | UCOR | ||
|---|---|---|---|---|---|
| UTC | OTC | UTC | OTC | ||
| Vegetation cover | 65.8 ± 14.3 | 91.7 ± 7.5 | 64.2 ± 15.3 | 98.3 ± 4.1 | |
| Bare soil | 34.2 ± 14.3 | 8.3 ± 7.5 | 35.8 ± 15.3 | 1.7 ± 4.1 | |
| APIACEA | Daucus carota | 5.3 ± 13.0 | 0 | 6.9 ± 13.1 | 0 |
| Scandix pecten-veneris | 4.3 ± 6.7 | 0 | 11.8 ± 26.3 | 0 | |
| ARACEAE | Arum italicum | 1.0 ± 2.5 | 0 | 1.5 ± 3.7 | 0 |
| ASTERACEAE | Chamaemelum mixtum | 0 | 0.3 ± 0.4 | 0 | 3.2 ± 3.1 |
| Leontodon taraxacoides | 0 | 8.1 ± 9.2 | 0 | 5.6 ± 8.1 | |
| Senecio jacobae | 2.6 ± 5.5 | 5.3 ± 1.9 | 0 | 2.0 ± 1.9 | |
| Tolpis barbata | 0 | 1.3 ± 3.2 | 0 | 0.7 ± 1.7 | |
| BRASSICACEAE | Diplotaxis catholica | 0 | 1.5 ± 3.8 | 0 | 6.9 ± 5.5 |
| Raphanus raphanistrum | 0 | 2.1 ± 3.8 | 0 | 0.7 ± 1.8 | |
| BORAGINACEAE | Echium plantagineum | 0 | 3.0 ± 7.2 | 0 | 0 |
| CARYOPHYLLACEAE | Cerastium glomeratum | 0 | 1.3 ± 3.2 | 1.1 ± 2.2 | 1.9 ± 2.1 |
| Spergula arvensis | 0 | 0 | 0 | 3.7 ± 4.1 | |
| FABACEAE | Medicago polymorpha | 0.2 ± 0.4 | 0 | 0 | 0 |
| Ornithopus pinnatus | 0 | 0 | 0 | 1.1 ± 1.8 | |
| Ornithopus sativus | 0 | 0 | 0 | 5.3 ± 6.6 | |
| Trifolium repens | 0 | 4.3 ± 9.4 | 0 | 0.7 ± 1.8 | |
| GERANIACEAE | Erodium botrys | 38.0 ± 24.2 | 18.6 ± 26.5 | 22.6 ± 26.4 | 9.7 ± 11.2 |
| Erodium cicutarium | 2.8 ± 3.2 | 6.5 ± 8.7 | 0 | 0 | |
| Geranium dissectum | 0 | 0.6 ± 1.6 | 0 | 4.8 ± 3.4 | |
| Geranium molle | 0 | 1.5 ± 2.3 | 17.8 ± 18.1 | 5.0 ± 6.6 | |
| IRIDACEAE | Gynandrisis sisyrinchium | 0 | 0.6 ± 1.6 | 0 | 0.1 ± 0.3 |
| PLANTAGINACEAE | Plantago lanceolata | 0 | 0 | 0.2 ± 0.4 | 1.4 ± 1.6 |
| POACEAE | Avena barbata | 25.6 ± 31.5 | 2.6 ± 4.1 | 16.7 ± 23.9 | 0.7 ± 1.8 |
| Bromus diandrus | 12.0 ± 12.1 | 6.9 ± 14.9 | 7.6 ± 13.4 | 1.0 ± 1.8 | |
| Bromus hordeaceus | 0 | 0.4 ± 0.9 | 0 | 0 | |
| Hordeum murinum | 1.8 ± 4.4 | 28.6 ± 20.3 | 7.0 ± 10.9 | 41.3 ± 12.3 | |
| Lolium multiflorum | 2.3 ± 3.0 | 0.6 ± 1.6 | 0 | 0 | |
| Lolium rigidum | 0 | 0.4 ± 0.9 | 0 | 0 | |
| Poa annua | 0.2 ± 0.5 | 0 | 0 | 0.1 ± 0.4 | |
| Vulpia geniculata | 1.3 ± 1.8 | 3.3 ± 2.6 | 3.0 ± 4.1 | 0.7 ± 0.3 | |
| POLYGONACEAE | Rumex angiocarpus | 0 | 0 | 0 | 3.0 ± 4.6 |
| Rumex bucephalophorus | 0 | 1.0 ± 1.7 | 0 | 0.3 ± 0.4 | |
| Rumex conglomeratus | 0 | 0.5 ± 1.1 | 2.4 ± 4.4 | 0 | |
| RUBIACEAE | Sherardia arvensis | 0 | 0.6 ± 1.6 | 0 | 0 |
| URTICACEAE | Urtica urens | 2.5 ± 4.8 | 0 | 1.4 ± 2.6 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serrano, J.; Shahidian, S.; Costa, F.; Carreira, E.; Pereira, A.; Carvalho, M. Can Soil pH Correction Reduce the Animal Supplementation Needs in the Critical Autumn Period in Mediterranean Montado Ecosystem? Agronomy 2021, 11, 514. https://doi.org/10.3390/agronomy11030514
Serrano J, Shahidian S, Costa F, Carreira E, Pereira A, Carvalho M. Can Soil pH Correction Reduce the Animal Supplementation Needs in the Critical Autumn Period in Mediterranean Montado Ecosystem? Agronomy. 2021; 11(3):514. https://doi.org/10.3390/agronomy11030514
Chicago/Turabian StyleSerrano, João, Shakib Shahidian, Francisco Costa, Emanuel Carreira, Alfredo Pereira, and Mário Carvalho. 2021. "Can Soil pH Correction Reduce the Animal Supplementation Needs in the Critical Autumn Period in Mediterranean Montado Ecosystem?" Agronomy 11, no. 3: 514. https://doi.org/10.3390/agronomy11030514