Winter Wheat Adaptation to Climate Change in Turkey


Department of Crop Science, Faculty of Agriculture, Siirt University, Siirt 56100, Turkey
Agronomy 2021, 11(4), 689; https://doi.org/10.3390/agronomy11040689
Submission received: 19 February 2021
/
Revised: 31 March 2021
/
Accepted: 1 April 2021
/
Published: 4 April 2021
(This article belongs to the Section Crop Breeding and Genetics)
Abstract
:Climate change scenarios reveal that Turkey’s wheat production area is under the combined effects of heat and drought stresses. The adverse effects of climate change have just begun to be experienced in Turkey’s spring and the winter wheat zones. However, climate change is likely to affect the winter wheat zone more severely. Fortunately, there is a fast, repeatable, reliable and relatively affordable way to predict climate change effects on winter wheat (e.g., testing winter wheat in the spring wheat zone). For this purpose, 36 wheat genotypes in total, consisting of 14 spring and 22 winter types, were tested under the field conditions of the Southeastern Anatolia Region, a representative of the spring wheat zone of Turkey, during the two cropping seasons (2017–2018 and 2019–2020). Simultaneous heat (>30 °C) and drought (<40 mm) stresses occurring in May and June during both growing seasons caused drastic losses in winter wheat grain yield and its components. Declines in plant characteristics of winter wheat genotypes, compared to those of spring wheat genotypes using as a control treatment, were determined as follows: 46.3% in grain yield, 23.7% in harvest index, 30.5% in grains per spike and 19.4% in thousand kernel weight, whereas an increase of 282.2% in spike sterility occurred. On the other hand, no substantial changes were observed in plant height (10 cm longer than that of spring wheat) and on days to heading (25 days more than that of spring wheat) of winter wheat genotypes. In general, taller winter wheat genotypes tended to lodge. Meanwhile, it became impossible to avoid the combined effects of heat and drought stresses during anthesis and grain filling periods because the time to heading of winter wheat genotypes could not be shortened significantly. In conclusion, our research findings showed that many winter wheat genotypes would not successfully adapt to climate change. It was determined that specific plant characteristics such as vernalization requirement, photoperiod sensitivity, long phenological duration (lack of earliness per se) and vulnerability to diseases prevailing in the spring wheat zone, made winter wheat difficult to adapt to climate change. The most important strategic step that can be taken to overcome these challenges is that Turkey’s wheat breeding program objectives should be harmonized with the climate change scenarios.
1. Introduction
According to global warming models, it is estimated that the rise in the global temperature will reach 1.5 °C in 2030, 2.5 °C in 2050, and 4 °C in 2100 [1]. Temperature increases have reached record levels (0.68–1.00 °C) in eight of the last ten years (2011–2020) [2]. Already approaching the 2030 temperature limit (1.5 °C) is causing humanity to be alarmed. Undoubtedly, climate change (CC) models will need to be reconstructed with new data to make more realistic predictions.
Based on long-term data (1901–2015), global precipitation increases approximately 2 mm every decade [3]. Even so, regional and local precipitation patterns (amount, distribution and frequency) are likely to deviate from the global trend [4]. For example, precipitation recorded (607 mm) in Turkey during the last decade (2011–2020) was 35 mm lower than the long-term (1950–2020) average precipitation (642 mm) [5].
In Turkey, during the last decade (2011–2020) the average temperature (14.4 °C) measured was 0.5 °C higher than the long-term (1950–2020) average temperature (13.9 °C). In Turkey, since 1998 (except 2011), the incidence of positive temperature anomalies has increased [5]. By the end of this century, in Turkey, located in the Mediterranean basin, it is estimated that temperature will gradually increase, and precipitation will primarily decrease along with an unpredictable pattern [6]. Since the 2000s, the effects of CC have been observed in wheat zones. For instance, it is expected that the cold and snowy winter season would turn to mild and rainy, and the summer season to hot and dry periods in the winter wheat zone (WWZ) [7].
Different strategies have been followed in studies conducted to determine the effect of CC on wheat. Some have focused solely on heat stress in wheat [8,9,10]. Others have studied the combined effects of heat and drought (HD) stresses on wheat [11,12,13,14].
The effects of heat stress on wheat are not yet fully understood. It is not known precisely at what temperature level heat stress starts to damage wheat. The research in [15] divided heat stress into two categories: chronic stress (20–30 °C) and heat shock (>32 °C). The research in [9] classified heat stress depending on the maximum temperature limit: heat stress (>30 °C) and hot days (>35 °C). On the other hand, there are no standard protocols to be followed when studying the effects of heat stress on wheat. Furthermore, it is unknown exactly which plant characters can reduce the adverse effects of heat stress on wheat.
HD stresses negatively affect all growth and development stages of wheat [16]. More precisely, the anthesis and grain filling periods are most sensitive to them [17]. They result in declining grains per spike by reducing pollen viability (i.e., spike infertility). Likewise, during the grain filling periods, they give rise to a decrease in grain size and weight, thereby negatively affecting yield and quality [12,18,19].
According to the CC projections for Turkey, yield reduction driven by HD stresses is estimated to reach 25–29% in winter wheat (WW) and facultative wheat (FW), and 15–16% in spring wheat (SW) [20,21]. It has been reported that the only way to prevent the decrease in yield could be by adapting the wheat phenology to the changing climate (i.e., changing the time to heading) [22,23,24].
Research in [25] reported that under the CC scenarios, yield could be increased by early planting of slow-growing (winter type) wheat instead of late planting of fast-growing (spring type) wheat, but facilitating wheat adaptation to CC in Turkey may require the exact opposite of their findings. There are at least four pieces of evidence that CC has begun to affect WW in Turkey. First, SW has started to be grown, albeit a little, in some parts of the WWZ, provided it is sown mostly in late autumn or early winter [26]. It has been experienced that a late sowing practice could prevent SW from being damaged by the cold. Second, some diseases prevailing in spring wheat zone (SWZ) (e.g., Septoria tritici) have begun to cause epidemics in WWZ of Turkey [27,28]. Third, recently, heat stress has occurred during drought in Turkey [5]. It was observed that during recent drought spells (e.g., in 2000, 2008, 2013, 2017, 2020) [5], WW could not tolerate HD stresses as much as SW, since varieties with shorter phenological cycles could be less affected by HD stresses [29,30]. Fourth, WW planting time has been shifted further in its zone (i.e., WWZ). Therefore, early WW varieties have been preferred in recent years [24,26].
Research in [31] highlighted that improvements in wheat management, such as early sowing, more irrigation and more nitrogen fertilizer, regardless of the wheat phenological cycle, could facilitate wheat adaption to CC. Obviously, this proposal does not appear to be a sustainable approach because of higher input requirement. On the other hand, there is an easy and practical way to enhance wheat adaptation to CC, i.e., harmonizing wheat phenology with changing climate. However, it has been speculated that genes regulating vernalization (VRN), photoperiod (PPD) and thermal time, or growing degree days (GDD), in WW could prevent its phenological profile flexible against CC [32]. More precisely, WW’s gene pool enrichment may be a proper approach to develop flexible phenology. By crossing WW with SW, new varieties with flexible phenology adaptable to CC can be created. What is suggested here has been implemented for many years. The improvement strategy of the IWWIP program coordinated by CIMMYT-ICARDA-Turkey has been built on the principle we mentioned [33]. The IWWIP program creates WW × SW crossing combinations to expand the WW adaptation boundaries and makes simultaneous selections over the multi-environment trials conducted in SWZ, WWZ, and facultative wheat zone (FWZ) of Turkey. In this article, the breeding method we propose to develop climate-resilient wheat varieties has been developed by inspiration from the IWWIP program.
This study’s objective was to determine the likely impact of CC on WW. For this purpose, a total of 36 wheat genotypes, consisting of 22 WW and 14 SW, were tested in one rain-fed field trial in the Southeastern Anatolia Region, representing the SWZ of Turkey, during two cropping seasons (2017–2018 and 2019–2020). By doing so, changes in grain yield and its relevant characteristics in WW were estimated, compared with those of SW, which was used as a control treatment. Throughout this research, we tried to answer specific questions: (a) how and to what extent could CC affect WW? (b) could the effect of CC on WW be measured under the field trials? (c) how could Turkey’s wheat breeding program be adapted to CC? Of course, the method we suggest has pros and cons. Despite those, our study brought up a primary topic. For the time being, Turkey’s wheat breeding program does not have a strategy to mitigate the effects of CC on wheat. For this reason, we have presented our suggestions on improving the adaptation of wheat to CC.
2. Materials and Methods
2.1. Experimental Design
A field trial was conducted in the experimental area (37°55′30″ N, 41°56′45″ E; 902 m asl) of the Faculty of Agriculture of Siirt University, Siirt, in the Southeastern Anatolia Region (SAR), a representative of the SWZ of Turkey, during two growing seasons (2017–2018 and 2019–2020) (Figure S1) [34]. All 36 bread wheat genotypes (Table S1), including 22 winter and 14 spring types, were tested in one trial. Experimental lay-out was a triple lattice square design. The trial was conducted over three replications. The plots consisted of four rows, 20 cm apart and 4 m long.
2.2. Wheat Genotypes
WW genotypes included 20 registered varieties, one landrace (Tir, from Van, Turkey), and one introduction from Canada (Broadview) (Table S1) [35]. Broadview is a strong winter type cultivar, selected 25th FAWWON-SA (entry#298) (25th Facultative and Winter Wheat Observation Nursery for Semi-arid Areas) [36]. On the other hand, SW genotypes included 12 registered varieties, one landrace (Siyare, from Siirt, Turkey), and one advanced breeding line (selected from 50th IBWSN (entry#19) (50th International Bread Wheat Screening Nursery) [37].
2.3. Plant Characteristcs
In this study, seven plant characteristics were observed, measured and evaluated as following [38]. Grain yield (GY): plants were harvested with a sickle (Zadoks-Z93) and spikes were threshed by hand. The grain yield (g m−2) obtained from the plot was converted into ton ha−1. Days to heading (DH) were calculated as Julian days starting from January 1 until half the heads emerged (Z55). Harvest index (HI): plants in one-meter-long row per plot were cut with a sickle above the soil surface, and their biomass was weighed (Z93). The spikes were threshed by hand and the grain weight was proportioned to the above-ground biomass (%). Thousand kernel weight (TKW): a random sample was taken of the whole grains harvested from each plot, 1000 seeds were counted by hand and weighed (g). Plant height (PH): the length of 10 main stems randomly selected from each plot were measured from the ground surface to the tip of the spike, excluding awn, recorded to the nearest cm (Z85). Grains per spike (GPS): 10 spikes were randomly sampled from main stems per plot, and grains per spike were counted (Z93). Spike sterility (SS): grains were developed in the first two (basal) florets of the spikelets per spike of all wheat genotypes tested in the experiment. However, the third floret grain formation exhibited a wide genotypic variation. Therefore, 10 spikes were randomly sampled from main stems per plot during the postfertilization stage (Z75). Grain abortion and infertility rates in the third floret were calculated and expressed as spike sterility (%).
2.4. Soil Properties
The soil at the experimental site was calcareous, slightly alkaline (pH = 7.97) and clayey. Organic matter was 14.2 g kg−1. Extractable P and K concentrations were 32 and 125 mg kg−1, respectively.
2.5. Crop Management
Both WW and SW genotypes were treated equally. The field trial was set up on December 8 of 2017 in the first season, and on November 28 of 2019 in the second season. The seeding rate was 500 seeds m−2 for all wheat genotypes. At sowing, 40 kg N ha−1 and 70 kg P ha−1 were applied. Besides, 80 kg N ha−1 was applied just before the stem elongation stage (Zadoks 30) in the early spring. Weeds were controlled manually. Pesticides were not applied, so that the reactions of wheat genotypes to diseases could be observed under the field conditions. SW genotypes were harvested by hand (with a sickle) in mid-June, while WW genotypes were harvested in early July. The trial was conducted under rain-fed conditions (without irrigation) in both cropping seasons.
2.6. Vernalization, Photoperiod and Growing Degree Days
Out of 36 wheat genotypes tested in the trial, vernalization (VRN) and photoperiod (PPD) genes for 18 were determined by [39] (Table S1). VRN and PPD data for one genotype (25th FAWWON-SA-298 = Broadview) were obtained from the IWWIP program [36]. The VRN and PPD responses of the remaining (17) genotypes were determined by visually comparing them with the previously defined 19 genotypes tested in the trial conducted during the two growing seasons.
Wheat is inherently sensitive to at least 14 h of light period (i.e., photoperiod) [40]. At the Siirt location, long days start on April 30 [5]. The PPD sensitivities of genotypes tested were determined by considering both the findings of [39] and DH values (before or after 30 April).
Growth and development periods in wheat are often explained in growing degree days (GDD) (i.e., thermal time or heat accumulation). Traditionally, it is assumed that wheat is functional at temperatures between 0 °C (base temperature-Tbase) and 30 °C (ceiling temperature-Tceiling). Temperatures below Tbase and above Tceiling are excluded in the GDD calculation. GDD values for the genotypes tested were calculated using the following Equation (1) [41]:
where Tmax is the maximum temperature (<Tceiling), and Tmin is the minimum temperature (>Tbase).
2.7. Comparing Winter Wheat with Spring Wheat: Relative Differences between Plant Characteristics
Two wheat groups (WW and SW) were planted simultaneously in a single trial and subjected to the same agronomic practices. Thus, the level of change in the GY and related traits of WW genotypes was determined by comparing SW genotypes (as a control treatment) [33,42]. For example, the relative difference in the GY of WW genotypes was calculated with the following Equation (2):
where GY is grain yield, WWGYmean is winter wheat grain yield mean, and SWGYmean is spring wheat grain yield mean.
The relative differences in the rest of plant characteristics observed were calculated as in GY.
2.8. Statistical Analyses
Most of the wheat genotypes, especially WW, used in the trial were not tested previously at the location of Siirt, Turkey. Therefore, the genotypic effects were considered random in the statistical models of variance analyses. Correspondingly, year, replication and block effects standing for the environment (variations in climate and soil over the years) in which the trial was conducted were also considered random in the statistical models (Table S2).
A combined analysis of variance (ANOVA) for the seven plant characters studied was performed over the two years (Table S2). Means of wheat genotypes were compared with the LSD multiple comparison test (Figures 3–9). ANOVA and LSD tests were conducted in SAS software.
Combined analysis of variance (ANOVA) showed statistically significant differences (p < 0.05 or p < 0.01) between years (Y), among wheat genotypes (G), and their interactions with each other (G × Y) for the seven plant characters (Table S2).
Correlation analysis was performed in Minitab software to determine pair-wise relationships between the seven plant characters (Figure S2). The relationships between plant characters and wheat genotypes were displayed by two multivariate analyses: principal components (PCA) and cluster (CA) analyses. PCA (Figure 10) was conducted in Microsoft Office Excel [43]. CA (Figures S3 and S4) was executed in Minitab software.
3. Results
3.1. Precipitation
According to climate data [5,6], the precipitation (522.4 mm) recorded in the first growing season (2017–2018) was 138 mm lower than the long-term (1939–2020) average precipitation (660.4 mm) (LTAP) (Table S3). On the other hand, the precipitation (757 mm) recorded in the second growing season (2019–2020) was 96.6 mm higher than LTAP. We noticed that precipitation decreased by 34.1 mm when comparing LTAP (660.4) with the precipitation (626.3 mm on average) recorded in the last three decades (from 1980 to 2020).
In the first season, the low precipitation (47.2 mm < LTAP = 111.2 mm) received in March accelerated the transition from stem elongation to reproductive phases in SW genotypes (Table S3). Fortunately, the high precipitation (146.6 mm > 63.8 mm) received in May contributed positively to the grain filling periods of SW genotypes, and the formation of reproductive organs (i.e., spike and florets) in WW genotypes (Figure 1).
In the second season, the precipitation received between November and January (51.4 mm, 75.8 mm, and 63.8 mm, respectively), when the germination, emergence, and tillering stages occurred, remained behind the LTAP (82.2 mm, 95.8 mm, and 97.5 mm, respectively) (Table S3). On the other hand, the precipitation received in February (137.2 mm), March (229.6 mm), and April (158.6 mm) was above LTAP (97.6 mm, 111.2 mm, and 105 mm, respectively). Thus, the high precipitation, together with mild weather conditions, provoked diseases such as Septoria tritici and Stagonospora nodorum, to infect plants. Moreover, low precipitation (40.4 mm < 63.8 mm) recorded in May in the second season, compared to that of the first season (146.6 mm > 63.8 mm), negatively affected the grain filling periods of the SW and the formation of the reproductive organs of the WW genotypes (Figure 1).
In June of both seasons, when the grain filling stages of WW genotypes coincided, almost no precipitation (0.2 mm and 2.8 mm) was received (Table S3). Compared to SW, WW genotypes exhibited longer phenological cycles, causing them to face DH stresses during grain filling stages.
3.2. Temperature
The minimum temperatures (Tmin) measured in both seasons (4.7 °C and 4.2 °C, respectively) were approximately 2.2 °C lower than the long-term average Tmin (6.7 °C) (LTATmin) (Table S3). The lowest Tmin (−9 °C) recorded in February of the second season gave rise to low to moderate cold damage only in SW genotypes. In other words, it caused leaf chlorosis and burning leaf tips on SW genotypes, indicating that they were susceptible to cold stress. [44]. However, WW genotypes were winter hardy.
The maximum temperatures (Tmax) measured in both growing seasons (23.8 °C and 21.7 °C, respectively) were 5.5 and 7.6 °C higher than the long-term average Tmax (16.2 °C) (LTATmax) (Table S3). Tmaxes (from 10.9 °C to 17.8 °C) observed during the winter season reached approximately twice the LTATmaxes (from 6.6 °C to 8.8 °C). The temperatures measured in May and June (32.2–36.2 °C) were well above the values (15–18 °C) considered optimal temperatures for wheat growth and development (Figure 2) [45].
3.3. Grain Yield
Grain yield (GY) means for SW and WW genotypes were 4.77 t ha−1 and 2.56 t ha−1, respectively (Figure 3). In other words, the GY mean of the WW genotypes grown in the SWZ was significantly lower [(2.56–4.77)/(4.77)×100 = −46.3%], see Equation (2) than that of the SW genotypes, which were used as a control treatment for comparison.
Within the SW group, the highest yielding genotype was Dinc (5.88 t ha−1), while the lowest was Siyare (2.59 t ha−1), the landrace grown locally in Siirt, Turkey (Table S1 and Figure 3). That is, Dinc had approximately twice the yielding capacity of Siyare.
Among the WW genotypes, the highest yielder was Pehlivan (3.60 t ha−1), and the lowest was Broadview (25thFAWWON-SA-298) (1.50 t ha−1) (Figure 3 and Table S1) [37]. This showed that the GY of Pehlivan was more than twice that of Broadview.
When all wheat genotypes tested in the trial were considered, it was recognized that Dinc had four times more GY than Broadview. The substantial difference in GY between SW and WW genotypes could indicate that the latter could not adapt to SWZ [46].
3.4. Days to Heading
In our experiment, 105 days (Julian days) were taken to initiate the heading stage in SW genotypes, while 130 days in WW genotypes (Figure 4). This underlined that the WW group headed 25 days (=130−105) later than the SW group. It is a typical characteristic that WW heads later than SW under the same growing conditions [47].
Among the SW genotypes, Tahirova had the longest heading time (115 days), while the 50th IBWSN-19, an advanced breeding line [38], had the shortest (98 days) (Figure 4 and Table S1). A period of 17 days occurred for days to heading (DH) within the SW genotypes.
In the WW group, the highest DH (142 days) was recorded for Broadview and the lowest DH (123 days) for Pehlivan (Figure 4 and Table S1). In other words, there occurred a 19-day interval for DH within WW genotypes. As a result, the heading time of the WW group was two days (19–17) longer than that of the SW group, indicating a wider genotypic variation.
3.5. Harvest Index
In our experiment, the average harvest index (HI) of SW genotypes was 38% (medium), and for WW genotypes was 29% (low), indicating that the WW group had a lower HI value (23.7%) than the SW group had (Figure 5).
Among the WW genotypes, Pehlivan had the highest HI (34%), and Broadview (25th FAWWON-SA-298) had the lowest HI (21%) (Figure 5 and Table S1). Specifically, Pehlivan had a higher HI value (17.2%) than the average HI (29%) of WW genotypes.
Variation in the HI values (from 35% for Siyare to 42% for Dinc) was found within SW genotypes (Figure 5 and Table S1). In other words, the HI of Dinc was 27% higher than that of Siyare. We realized a wide variation among the HI values of the genotypes tested in the experiment. It was evident that the highest HI value in the trial (42% for Dinc) reached exactly twice the lowest HI value (21% for Broadview).
3.6. Plant Height
The average plant height (PH) of the WW genotypes was 106 cm, and for the SW genotypes was 96 cm (Figure 6), indicating that the former was 10 cm taller than the latter.
Among the WW group, a wide genotypic variation (141−90 = 51 cm) was observed for PH (Figure 6 and Table S1), mainly due to the three tallest genotypes: Dagdas-94, Tir and Dogu-88 (141 cm, 130 cm, and 120 cm, respectively). We assume that if those were excluded from WW group, the genotypic variation for PH would be narrowed by more than 50% (114−90 = 24 cm). However, it would still be large enough because modern cultivars currently range 90 cm to 110 cm in height [48].
In the SW group, the landrace Siyare was the highest (120 cm) in PH, and three genotypes (Pandas, Altindane, and one advanced breeding line, selected from 50th IBWSN-19) were the lowest (90 cm) (Figure 6 and Table S1). As in the WW group, the genotypic variation for PH was large enough in the SW group (120−90 = 30 cm). However, if Siyare was excluded from the SW group, PH would be narrowed by 50% (105−90 = 15 cm).
In this experiment, we found that landraces and WW cultivars developed for rain-fed areas were quite taller. That was the main reason why genotypic variation for PH in our trials was so wide [49].
3.7. Thousand Kernel Weight
The WW group had a 19.4% lower (29 g) thousand kernel weight (TKW) than the SW group (36 g) (Figure 7). In the SW group, Altindane had the highest TKW (39 g), while Tahirova had the lowest (33 g) (Figure 7 and Table S1). Therefore, genotypic variation (39−33 = 6 g) for TKW within the SW group was not large enough. As for the WW group, TKW values ranged from 22 g (Broadview = 25th FAWWON-SA-298) to 34 g (Pehlivan) (Figure 7 and Table S1). It showed that genotypic variation (34−22 = 12 g) for TKW within the WW group reached twice that of the SW group (6 g).
3.8. Grains per Spike
In our experiment, the average grains per spike (GPS) of the SW group was 36, and for the WW group was 25, indicating that the former had a 30.5% higher GPS than the latter (Figure 8). There was a considerable difference (11 grains = 36−25) between wheat groups for GPS. Within the SW group, GPS varied between 39 for Adana-99 and 34 for Tahirova (Figure 8 and Table S1). However, genotypic variation (5 grains = 39−34) for GPS in the SW group was limited. As for WW group, GPS ranged from 30 for Pehlivan to 20 for Broadview (25th FAWWON-SA-298) (Figure 8 and Table S1). Compared to the SW group (5 grains), the genotypic variation for GPS within the WW group was slightly broader (10 grains = 30−20).
3.9. Spike Sterility
HD stresses occurring at preanthesis and during anthesis cause severe spike sterility (SS) [50]. Grains were developed in the first two florets (basal florets f1 and f2) in each spikelet of all wheat genotypes tested. However, almost all WW genotypes (88%) could not form grains in the third florets of spikelets per spike (Figure 9). A few genotypes from the SW group (23%) could not either. Therefore, WW genotypes had about a three-fold (282.6%) higher SS rate than SW genotypes.
3.10. Vernalization, Photoperiod, and Growing Degree Days
The wheat genotypes tested in this experiment were categorized into two groups in terms of the vernalization (VRN) requirement based on molecular data and visual observations: genotypes’ high vs low responsiveness to VRN (i.e., genotypes winter vs. spring growth habit). On the other hand, all SW, and some WW genotypes, were insensitive to photoperiod (PPD) (Table S1) [39]. All SW genotypes headed before April 30 (light period <14 h). In our case, DH could be irrelevant with PPD sensitivity, because all WW genotypes, including the PPD insensitive group, headed after April 30 (light period >14 h) (Figure 4). In fact, DH’s main driving force was growing degree days (GDD). The GDD values calculated for the SW genotypes (from 1567 to 1801 °Cd) were lower than those of the WW genotypes (from 1927 to 2293 °Cd) (Table S1). It has been pointed out that WW genotypes accumulate more GDD to initiate heading [40].
3.11. Multivariate Analyses for Plant Characteristics and Genotypes
The positive and negative relationships between all plant characteristics studied were found to be statistically significant (**, p < 0.01) (Figure S2). Among them, the lowest correlation coefficient (r = 0.448 **) was calculated between DH and PH, while the highest (r = −0.966 ** and r = 0.966 **, respectively) was calculated between SS and GPS, and SS and DH. Significant positive relationships were determined between GY and TKW, GPS and HI, and negatively significant relationships between GY and PH, DH and SS.
A biplot graph created by the principal components analysis (PCA) allows us to visually interpret the relationships between genotypes and their plant characters (Figure 10). The first two PCA components (Principal Components 1 vs. 2 (PC1 vs. PC2)) explained 94% of the relationships between genotypes and plant characters involved.
GY, HI, GPS and TKW were located on the positive axis of PC1 in the biplot, while PH, DH, and SS were on the negative axis. This showed that plant characters formed two main groups (Figure 10). However, the second axis (PC2) of the biplot separated PH from DH and SS, creating two subgroups. PH was located on the positive axis of PC2, and DH and SS on the negative axis. This grouping type was verified with the dendrogram obtained from the cluster analysis (CA) (Figure S3). CA separated all plant characters into three clusters. CA revealed that HI, GPS, and TKW, which were in the first cluster, had a positive effect on GY, and that SS and DH in the third cluster had an adverse impact. The fact that PH created a single cluster (cluster 2) and a separate group showed that it did not have a direct contribution (negative or positive) to the GY.
SW genotypes were located on the positive axis of PC1 in the biplot graph (Figure 10), indicating that they had GY, HI, TKW and GPS values higher than the average. On the other hand, WW genotypes were located on the negative axis of PC1, meaning that the BH, DH and SS values for WW genotypes were above the average. Wheat genotypes formed two main clusters (WW vs. SW) on the biplot, just like plant traits. Interestingly, the genotypes that deviated from the origin of the biplot (for example, G4, G21, G35, and G22) formed different clusters: four clusters (clusters 1–4) for WW genotypes and three clusters (clusters 5–7) for SW genotypes (Figure S4). Genotypic variations in PH (for clusters 3 and 4) and GY and its components (i.e., low adaptability for clusters 1 and 2), showed a differential effect in different clustering of WW genotypes. As for the SW genotypes, locally adaptiveness (for cluster 6) and facultative growth type (for cluster 5) resulted in different sets in the CA.
4. Discussion
4.1. Heat and Drought Tolerance
We believe that focusing only on heat stress is not a holistic approach to studying the possible effects of CC on wheat [10,11]. The frequency of concurrent heat and drought (HD) stress is anticipated to increase in the future [13]. All growth and development periods of wheat can be adversely affected by HD stresses. However, HD stresses occurring during the flowering and grain filling periods of wheat may result in dramatic economic losses [51]. This was confirmed by lack of precipitation, and the high Tmax values recorded at anthesis (32.2 and 33.7 °C in May) and grain filling phases (34.3 and 36.2 °C in June) during the two seasons this study was conducted.
The limit (upper optimal temperature -Topt) at which wheat starts to be adversely affected by high temperature differs according to studies: 15 °C [52], 18 °C [45,53,54,55,56] and 22 °C [57] and 30 °C [9]. However, in most of these studies, 15–18 °C was determined as Topt for wheat. The Topt (15–18 °C) proposed by the new findings coincides with the upper limit where the VRN requirement can be met [53,54,56]. In other words, it is an interesting coincidence that the upper-temperature limit (15–18 °C) at which the VRN requirement is met, and the boundary where temperature is optimal (15–18 °C), are equivalent. There have found no literature on this subject. We believe that the Topt range for WW is determined explicitly by the VRN genes. Research in [58] found that VRN genes affected the formation of freezing tolerance, stem elongation (i.e., plant height), spike architecture and flowering characteristics, as well as meeting the low-temperature requirement of wheat. The findings of [58] support our suggestion.
In interpreting the findings obtained from our study, 18 °C was accepted as the limit where high-temperature stress started [45,52,53,54,55,56]. Declines ranging from 2.95% to 3.31% in the GY of WW, compared with SW, tested in our experiment, occurred for each one °C temperature increase above 18 °C. Research in [52,55] found that each one °C temperature increase caused decreases between 3.3% and 6.4% in the GY of wheat. The reasons for the differences between the research findings were due to the different Topt temperatures in each study, the application of heat stress in different temperature ranges, the different heat stress application times and the location where the heat stress was applied in different geographic positions (in different altitudes and latitudes). Unfortunately, there is no standard protocol for measuring the effects of heat stress on wheat.
Due to the combined effect of HD stresses, WW’s GY was 46% lower than that of SW (control treatment) in our trial. Research in [55,59,60] determined that heat stress reduced the GY of wheat by 52–54%. These findings agreed with ours. As in GY, HD stresses had adverse effects on other plant characters such as TKW, SS, GPS, and HI. They also reported that, like our findings (44%), heat stress significantly reduced HI and GPS values (84% and 61%, respectively) in wheat. Contrary to our results, they stated that heat stress did not adversely affect the PH, DH and TKW in wheat. The difference between the two studies was mainly due to the genetic makeup of wheat. They studied spring durum wheat genotypes with low genotypic variation (88–95 days for HD and 75–85 cm for PH), while we studied SW and WW genotypes with a broad genetic variation (98–142 days for DH and 90–141 cm for PH). On the other hand, in the same study, [55], no adverse effect of heat stress on TKW (34–39 g) was determined, while a large effect (22–39 g) was determined in this study. Like our findings, [14] determined that the combined impact of HD stresses led TKW to a decrease (23%) in wheat.
Under normal conditions, the grain formation rate per average per spikelet of the wheat spike is around 2.7, and it is well known that the sterility rate in the third florets of the wheat spike is about 30% [61]. One of the most important reasons why the WW genotypes tested had low GY was sterility in the spike (SS). Time to initiate anthesis in WW, covering pollination and fertilization, coincided with which heat (32.2–33.7 °C in May) and drought (low rainfall in the second season, 40 mm in May) stresses occurred. Therefore, infertility in the third floret of the spikelet was developed at a rate of 88% on average. [19] and highlighted that the combined effect of drought (5 days) and heat (32 °C) applied to wheat during flowering caused functional irregularities in female and male organs at the level of 34% and 66%, respectively. Our research findings were supported by theirs.
Research in [14] concluded that wheat genotypes with thicker, greener leaves, higher photosynthetic capacity and increased water use index, could overcome the simultaneous effects of HD stresses with less damage during both anthesis and grain filling periods. Our findings showed that WW genotypes (e.g., Pehlivan), which could mature faster in a shortened period, were better able to avoid the destructive effects of both stresses. Among the WW genotypes tested in our experiment, there were registered varieties tolerant to drought (e.g., Bayraktar-2000 and Karahan-99). However, we confirmed that drought tolerance alone was insufficient for wheat to adapt to CC, because of low GY. Early heading is often considered a type of escape mechanism [62]. We elucidated that early heading genotypes within the WW group generally had lower SS and higher TKW and HI, and consequently higher GY. All of this underlines that earliness per se can be a crucial characteristic for wheat adapting to CC.
To summarize, our research findings revealed that due to the long phenological cycle of WW, especially anthesis and grain filling periods were severely affected by HD stresses. Therefore, we made suggestions on how WW could adapt to CC.
4.2. Suggestions to Mitigate Possible Effects of Climate Change on Winter Wheat
4.2.1. Wheat Breeding Program
Wheat breeding activities carried out in Turkey are executed in two independent programs: the WW breeding program (WWBP) and the SW breeding program (SWBP) (Figure S1). [63]. Today, there is hardly ever a germplasm exchange between the two programs. [64]. However, it is thought that WW would not adapt to the WWZ in the future owing to the effect of CC [16,65]. Each year, two-thirds of the wheat cultivation area of Turkey is reserved for WW [66]. If CC would threaten WWZ more frequently than before, a new wheat breeding strategy should be put forward urgently.
4.2.2. Crossing Block
Currently, parents in the WWBP’s crossing block (CB) are characterized only by the winter growth type. Therefore, WW × WW crossing combinations are created regularly. However, the winter wheat crossing block (WWCB) gene pool should be enriched to adapt to changing climatic conditions. Parents of the spring growth type should be included in the WWCB. Gene frequencies of parents of the winter growth type, mainly standing for VRN, PPD and cold tolerance (CT) characters in the WWCB should be altered gradually. It is estimated that these characters would make it challenging to adapt WW to CC (Table 1). The easy way to cope with this challenge is to create WW × SW crossing combinations [42]. We think that WW carrying SW genes would adapt to CC more effectively. Thus, we hope that the facultative wheat (FW) type obtained from WW × SW crossings will begin the first phase of adaptation to CC [6]. Since the climate is changing dynamically, we must run a dynamic wheat breeding program to adapt to it.
4.2.3. Early Generations
Today, populations (F2, F3, and F4) in the segregating generations of WWBP are tested commonly just in one location, where good agronomic practices are provided. However, the selection environment must also be changed for WWBP to adapt to changing climatic conditions. F2, F3, and F4 generations created from WW × SW crossing combinations should be tested simultaneously in contrasting locations, representing both WWZ and SWZ, to make an efficient selection (Figure S1). While choosing the locations from SWZ, different temperatures (testing for heat stress), and precipitation patterns (testing for drought stress), should be considered. Besides, diseases that cause economic losses in SWZ have managed to reach WWZ due to atmospheric barriers being broken by CC and creating epidemics in the WWZ [27,28]. Therefore, segregating generations (WW × SW) should be tested against diseases causing epidemics in SWZ.
4.2.4. Advanced Generations
According to the breeding strategy we propose for CC adaptation, advanced lines selected from WW × SW crossing combinations should be simultaneously tested against yield performance, quality, diseases, pests and HD stresses. Preliminary yield and advanced yield trials must be conducted over the locations to be selected from both the WWZ and SWZ (Figure S1). For instance, to carry out multienvironment trials for WW advanced lines, the following locations from the SWZ (and FWZ) would be preferred: testing for HD stresses in location Sanlıurfa, screening for high yield potential, disease resistance, high humidity, and heat stress in location Adana, and selecting for disease resistance in location Sakarya, Turkey.
4.2.5. Plant Characteristics Essential for Adapting to Climate Change
Plant characteristics that would improve WW’s adaptation to CC are given in Table 1, for more details, see [17,67,68]. We envisage that WW’s typical characters (e.g., VRN, PPD, and CT) would gradually lose their priorities due to global warming [32]. On the other hand, there is no doubt that tolerance to HD stresses, which are among the classical breeding aims of wheat, would be much more critical than ever before [13,14]. It is predicted that if the atmospheric CO2 concentration, which is the primary driver of CC, continues to rise, the photosynthetic capacity of wheat, a C3 plant, will increase, thereby increasing its yield potential. A new subject should be included in wheat’s breeding objectives: CO2 use efficiency [11].
4.2.6. New Approaches in Wheat Agronomy for Adapting to Climate Change
Due to the influence of CC, WW’s planting time has started to shift. New cultivars that mature earlier, suitable for late planting and tolerant to HD stresses, should be preferred [69]. Soil sustainability is of paramount importance for adapting to CC. Therefore, a conservation tillage system should be chosen in wheat cultivation [70]. Besides, smart irrigation systems should be preferred for irrigated wheat areas [71].
4.2.7. A Particular Topic: Genetically Modified Wheat
Genetically modified (GM) wheat can be conceptualized, in that traditional wheat breeding methods may be unsuccessful in developing new wheat varieties to adapt to CC [72]. In such a case, due to food security concerns driven by CC, the GM wheat issue would be brought back to the table. Thus, some would favor the GM wheat approach, and even try to put it into practice. To illustrate this, in 2020, one drought-resistant GM wheat variety (HB4) was announced to have been developed in Argentina, where wheat production has decreased significantly due to CC in recent years [73].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Priority rankings of plant characteristics for adapting to climate change in Turkey.
| Plant Characteristics | Current | Future | Reference |
|---|---|---|---|
| Vernalization | +++ | + | [53,74] |
| Photoperiod | ++ | + | [40,56] |
| Cold tolerance | +++ | + | [16,75] |
| Grain yield (Food security) | ++++ | +++++ | [55] |
| Grain quality | +++ | ++++ | [65] |
| Phenology | + | ++++ | [76,77] |
| Thermal time | +++ | + | [41,78] |
| Earliness | + | ++++ | [45] |
| Heat tolerance | ++ | +++++ | [9,10,13,17] |
| Drought tolerance | ++++ | +++++ | [14,17,19] |
| Waterlogging | + | ++ | [79] |
| Disease resistance | +++ | ++++ | [27,28] |
| Insect pest resistance | + | +++ | [80,81] |
| Water use efficiency | + | ++++ | [82] |
| CO2 use efficiency | + | +++ | [11] |
| Genetically modified wheat | ? | ? | [73] |
+ not important, ++ low priority, +++ medium priority, ++++ high priority, +++++ the most important, ? unknown.
5. Conclusions
WW tested in SWZ exhibited a lower performance for GY, and related plant characters, compared to SW because, we think, WW’s VRN and PPD genes and high GDD accumulation make it challenging to adapt to CC. The way to overcome this is by shortening the phenological cycle of WW (i.e., reduce days to heading) and thus facilitating adaptation to CC.
How CC will affect wheat is not fully understood yet. Therefore, there is no concrete suggestion on what breeding strategy should be followed to facilitate the adaptation of wheat to CC. Most of the studies on CC are only predictions derived from CC models. However, an easy-to-implement and effective strategy would be to test breeding nurseries developed from WW × SW combinations simultaneously in both the WWZ and the SWZ. By doing so, we can effectively determine whether WW will adapt to the warmer and drier conditions envisaged by the CC scenarios. The method we propose can help breeders carry out wheat breeding activities in developed and developing countries to improve new varieties adapted to CC.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/agronomy11040689/s1, Figure S1: Wheat zones of Turkey, Figure S2: Correlations between plant characteristics, Figure S3: Dendrogram for plant characteristics, Figure S4: Dendrogram for genotypes, Table S1: Wheat genotypes tested, Table S2: Analyses of variance for plant characteristics, Table S3: Climate data and wheat growth and development stages.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The author declares no conflict of interest.
References
- Intergovernmental Panel on Climate Change. Available online: https://www.ipcc.ch/site/assets/uploads/sites/2/2019/05/SR15_Chapter2_Low_Res.pdf (accessed on 18 February 2021).
- National Centers for Environmental Information. Available online: https://www.ncdc.noaa.gov/sotc/global/202013 (accessed on 18 February 2021).
- Environmental Protection Agency. Available online: https://www.epa.gov/climate-indicators/climate-change-indicators-us-and-global-precipitation (accessed on 18 February 2021).
- Dai, A.; Zhao, T.; Chen, J. Climate Change and Drought: A Precipitation and Evaporation Perspective. Curr. Clim. Chang. Rep. 2018, 4, 301–312. [Google Scholar] [CrossRef]
- Turkish State Meteorological Service. Available online: https://www.mgm.gov.tr/eng/yearly-climate.aspx (accessed on 18 February 2021).
- Climate Change and Its Projections with New Scenarios in Turkey (in Turkish). Available online: https://mgm.gov.tr/FILES/iklim/iklim-degisikligi-projeksiyon2015.pdf (accessed on 18 February 2021).
- Demircan, M.; Gürkan, H.; Eskioğlu, O.; Arabacı, H.; Coşkun, M. Climate Change Projections for Turkey: Three Models and Two Scenarios. Turk. J. Water Sci. Manag. 2017, 1, 22–43. [Google Scholar] [CrossRef]
- Hütsch, B.W.; Jahn, D.; Schubert, S. Grain yield of wheat (Triticum aestivum L.) under long-term heat stress is sink-limited with stronger inhibition of kernel setting than grain filling. J. Agron. Crop. Sci. 2018, 205, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Balla, K.; Karsai, I.; Bónis, P.; Kiss, T.; Berki, Z.; Horváth, Á.; Mayer, M.; Bencze, S.; Veisz, O. Heat stress responses in a large set of winter wheat cultivars (Triticum aestivum L.) depend on the timing and duration of stress. PLoS ONE 2019, 14, e0222639. [Google Scholar] [CrossRef]
- Thistlethwaite, R.J.; Tan, D.K.; Bokshi, A.I.; Ullah, S.; Trethowan, R.M. A phenotyping strategy for evaluating the high-temperature tolerance of wheat. Field Crop. Res. 2020, 255, 107905. [Google Scholar] [CrossRef]
- Sabella, E.; Aprile, A.; Negro, C.; Nicolì, F.; Nutricati, E.; Vergine, M.; Luvisi, A.; De Bellis, L. Impact of Climate Change on Durum Wheat Yield. Agronomy 2020, 10, 793. [Google Scholar] [CrossRef]
- Sattar, A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Rizwan, M.S.; Hussain, M.; Jabran, K.; Cheema, M.A. Terminal drought and heat stress alter physiological and biochemical attributes in flag leaf of bread wheat. PLoS ONE 2020, 15, e0232974. [Google Scholar] [CrossRef]
- Schmidt, J.; Claussen, J.; Wörlein, N.; Eggert, A.; Fleury, D.; Garnett, T.; Gerth, S. Drought and heat stress tolerance screening in wheat using computed tomography. Plant Methods 2020, 16, 1–12. [Google Scholar] [CrossRef]
- Schmidt, J.; Garcia, M.; Brien, C.; Kalambettu, P.; Garnett, T.; Fleury, D.; Tricker, P.J. Transcripts of wheat at a target locus on chromosome 6B associated with increased yield, leaf mass and chlorophyll index under combined drought and heat stress. PLoS ONE 2020, 15, e0241966. [Google Scholar] [CrossRef]
- Wang, H.; Lemke, R.; Goddard, T.; Sprout, C. Tillage and root heat stress in wheat in central Alberta. Can. J. Soil Sci. 2007, 87, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Braun, H.J.; Payne, T. Mega-Environment breeding. In Physiological Breeding I: Interdisciplinary Approaches to Improve Crop Adaptation; Reynolds, M.P., Pask, A.J.D., Mullan, D.M., Eds.; CIMMYT: Mexico City, Mexico, 2012; pp. 6–17. Available online: https://repository.cimmyt.org/handle/10883/1287 (accessed on 18 February 2021).
- Snowdon, R.J.; Wittkop, B.; Chen, T.-W.; Stahl, A. Crop adaptation to climate change as a consequence of long-term breeding. Theor. Appl. Genet. 2020. [Google Scholar] [CrossRef]
- Hochman, Z.; Gobbett, D.L.; Horan, H. Climate trends account for stalled wheat yields in Australia since 1990. Glob. Chang. Biol. 2017, 23, 2071–2081. [Google Scholar] [CrossRef]
- Fábián, A.; Sáfrán, E.; Szabó-Eitel, G.; Barnabás, B.; Jäger, K. Stigma Functionality and Fertility Are Reduced by Heat and Drought Co-stress in Wheat. Front. Plant Sci. 2019, 10, 244. [Google Scholar] [CrossRef]
- Vanli, Ö.; Ustundag, B.B.; Ahmad, I.; Hernandez-Ochoa, I.M.; Hoogenboom, G. Using crop modeling to evaluate the impacts of climate change on wheat in southeastern turkey. Environ. Sci. Pollut. Res. 2019, 26, 29397–29408. [Google Scholar] [CrossRef]
- Yeşilköy, S.; Şaylan, L. Yields and Water Footprints of Sunflower and Winter Wheat under Different Climate Projections. J. Clean. Prod. 2021, 298, 126780. [Google Scholar] [CrossRef]
- Yang, C.; Fraga, H.; Van Ieperen, W.; Trindade, H.; Santos, J.A. Effects of climate change and adaptation options on winter wheat yield under rainfed Mediterranean conditions in southern Portugal. Clim. Chang. 2019, 154, 159–178. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Villegas, J.; Milan, A.M.; Alexandrov, N.; Asseng, S.; Challinor, A.J.; Crossa, J.; Van Eeuwijk, F.; Ghanem, M.E.; Grenier, C.; Heinemann, A.B.; et al. CGIAR modeling approaches for resource-constrained scenarios: I. Accelerating crop breeding for a changing climate. Crop. Sci. 2020, 60, 547–567. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, H.; Bentley, A. Changing times: Opportunities for altering winter wheat phenology. Plants People Planet 2021, 3, 113–123. [Google Scholar] [CrossRef]
- Hunt, J.R.; Lilley, J.M.; Trevaskis, B.; Flohr, B.M.; Peake, A.; Fletcher, A.; Zwart, A.B.; Gobbett, D.; Kirkegaard, J.A. Early sowing systems can boost Australian wheat yields despite recent climate change. Nat. Clim. Chang. 2019, 9, 244–247. [Google Scholar] [CrossRef]
- Karapinar, B.; Özertan, G. Yield implications of date and cultivar adaptation to wheat phenological shifts: A survey of farmers in Turkey. Clim. Chang. 2020, 158, 453–472. [Google Scholar] [CrossRef]
- Turgay, E.B.; Buyuk, O.; Olmez, F.; Yildirim, A.F. Determination of prevalence of the Septoria leaf blotch disease of wheat [Zymoseptoria tritici (Desm. Quaedvlieg & Crous)] in Central Anatolia and its molecular identification. Plant Prot. Bull. 2016, 56, 283–293. [Google Scholar]
- Impact of Climate Change on Wheat Production in Kentucky. Available online: http://dx.doi.org/10.13023/PSSRR.2014.3 (accessed on 18 February 2021).
- Zhao, Z.; Rebetzke, G.J.; Zheng, B.; Chapman, S.C.; Wang, E. Modelling impact of early vigour on wheat yield in dryland regions. J. Exp. Bot. 2019, 70, 2535–2548. [Google Scholar] [CrossRef] [PubMed]
- Zeleke, K. Simulating Agronomic Adaptation Strategies to Mitigate the Impacts of Climate Change on Wheat Yield in South-Eastern Australia. Agronomy 2021, 11, 337. [Google Scholar] [CrossRef]
- Dubey, R.; Pathak, H.; Chakrabarti, B.; Singh, S.; Gupta, D.K.; Harit, R. Impact of terminal heat stress on wheat yield in India and options for adaptation. Agric. Syst. 2020, 181, 102826. [Google Scholar] [CrossRef]
- Dowla, M.N.U.; Edwards, I.; O’Hara, G.; Islam, S.; Ma, W. Developing Wheat for Improved Yield and Adaptation Under a Changing Climate: Optimization of a Few Key Genes. Engineering 2018, 4, 514–522. [Google Scholar] [CrossRef]
- Morgounov, A.; Ozdemir, F.; Keser, M.; Akin, B.; Payne, T.; Braun, H.-J. International Winter Wheat Improvement Program: History, activities, impact and future. Front. Agric. Sci. Eng. 2019, 6, 240. [Google Scholar] [CrossRef] [Green Version]
- Hanson, H.; Borlaug, N.E.; Anderson, R.G. Wheat in the Third World; Westview Press: Boulder, CO, USA, 1982; p. 174. [Google Scholar]
- Graf, R.; Thomas, J.; Gaudet, D.; Laroche, A.; Beres, B. Broadview hard red winter wheat. Can. J. Plant Sci. 2012, 92, 177–181. [Google Scholar] [CrossRef] [Green Version]
- International Winter Wheat Improvement Program. Available online: http://www.iwwip.org/Nursery/Details/1009 (accessed on 18 February 2021).
- International Maize and Wheat Improvement Center. Available online: https://data.cimmyt.org/dataset.xhtml?persistentId=hdl:11529/10548147 (accessed on 18 February 2021).
- Pask, A.J.D.; Pietragalla, J.; Mullan, D.M.; Reynolds, M.P. Physiological Breeding II: A Field Guide to Wheat Phenotyping; CIMMYT: Mexico City, Mexico, 2012; Available online: https://repository.cimmyt.org/handle/10883/1288?show=full (accessed on 18 February 2021).
- Andeden, E.E.; Yediay, F.; Baloch, F.S.; Shaaf, S.; Kilian, B.; Nachit, M.; Ozkan, H. Distribution of vernalization and photoperiod genes (Vrn-A1, Vrn-B1, Vrn-D1, Vrn-B3, Ppd-D1)in Turkish bread wheat cultivars and landraces. Cereal Res. Commun. 2011, 39, 352–364. [Google Scholar] [CrossRef]
- Dubcovsky, J.; Loukoianov, A.; Fu, D.; Valarik, M.; Sanchez, A.; Yan, L. Effect of Photoperiod on the Regulation of Wheat Vernalization Genes VRN1 and VRN2. Plant Mol. Biol. 2006, 60, 469–480. [Google Scholar] [CrossRef] [Green Version]
- McMaster, G.S. Development of the wheat plant. In Wheat: Science and Trade; Carver, B.F., Ed.; Wiley-Blackwell: Ames, IA, USA, 2009; pp. 31–55. Available online: https://doi.org/10.1002/9780813818832 (accessed on 18 February 2021).
- Almeida, A.S.; Macas, B.; Rodrigues, V.L.G.; Torrao, M.M. The history of wheat breeding in Portugal. In The World Wheat Book: The History of Wheat Breeding; Bonjean, A.P., Angus, W.S., van Ginkel, M., Eds.; Lavoisier: Paris, France, 2016; Volume 3, pp. 93–126. Available online: https://www.lavoisier.fr/livre/agriculture/the-world-wheat-book/bonjean/descriptif-9782743020910 (accessed on 18 February 2021).
- Lipkovich, I.A.; Smith, E.P. Biplot and Singular Value Decomposition Macros forExcel. J. Stat. Softw. 2002, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Spring Freeze Injury to Kansas Wheat. Available online: https://bookstore.ksre.ksu.edu/pubs/C646.PDF (accessed on 18 February 2021).
- Ochagavía, H.; Prieto, P.; Zikhali, M.; Griffiths, S.; Slafer, G.A. Earliness Per Se by Temperature Interaction on Wheat Development. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cook, J.P.; Heo, H.-Y.; Varella, A.C.; Lanning, S.P.; Blake, N.K.; Sherman, J.D.; Martín, J.M.; See, D.; Chao, S.; Talbert, L.E. Evaluation of a QTL Mapping Population Composed of Hard Red Spring and Winter Wheat Alleles Using Various Marker Platforms. Crop. Sci. 2018, 58, 701–712. [Google Scholar] [CrossRef]
- Huang, M.; Mheni, N.; Brown-Guedira, G.; McKendry, A.; Griffey, C.; Van Sanford, D.; Costa, J.; Sneller, C. Genetic analysis of heading date in winter and spring wheat. Euphytica 2018, 214, 128. [Google Scholar] [CrossRef]
- Würschum, T.; Langer, S.M.; Longin, C.F.H. Genetic control of plant height in European winter wheat cultivars. Theor. Appl. Genet. 2015, 128, 865–874. [Google Scholar] [CrossRef]
- Fiore, M.C.; Mercati, F.; Spina, A.; Blangiforti, S.; Venora, G.; Dell’Acqua, M.; Lupini, A.; Preiti, G.; Monti, M.; Pè, M.E.; et al. High-Throughput Genotype, Morphology, and Quality Traits Evaluation for the Assessment of Genetic Diversity of Wheat Landraces from Sicily. Plants 2019, 8, 116. [Google Scholar] [CrossRef] [Green Version]
- Nuttall, J.G.; Barlow, K.M.; Delahunty, A.J.; Christy, B.P.; O’Leary, G.J. Acute High Temperature Response in Wheat. Agron. J. 2018, 110, 1296–1308. [Google Scholar] [CrossRef] [Green Version]
- Ren, S.; Qin, Q.; Ren, H.; Sui, J.; Zhang, Y. Heat and Drought Stress Advanced Global Wheat Harvest Timing from 1981–2014. Remote Sens. 2019, 11, 971. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Asseng, S.; Müller, C.; Ewert, F.; Elliott, J.; Lobell, D.B.; Martres, P.; Ruane, A.C.; Wallach, D.; Jones, J.W.; et al. Similar estimates of temperature impacts on global wheat yield by three independent methods. Nat. Clim. Chang. 2016, 6, 1130–1136. [Google Scholar] [CrossRef]
- Yan, L.; Li, G.; Yu, M.; Fang, T.; Cao, S.; Carver, B.F. Genetic Mechanisms of Vernalization Requirement Duration in Winter Wheat Cultivars. In Advances in Wheat Genetics: From Genome to Field, 1st ed.; Ogihara, Y., Takumi, S., Handa, H., Eds.; Springer: Tokyo, Japan, 2015; pp. 117–125. [Google Scholar]
- Gol, L.; Tomé, F.; Von Korff, M. Floral transitions in wheat and barley: Interactions between photoperiod, abiotic stresses, and nutrient status. J. Exp. Bot. 2017, 68, 1399–1410. [Google Scholar] [CrossRef]
- El Hassouni, K.; Belkadi, B.; Filali-Maltouf, A.; Tidiane-Sall, A.; Al-Abdallat, A.; Nachit, M.; Bassi, F.M. Loci Controlling Adaptation to Heat Stress Occurring at the Reproductive Stage in Durum Wheat. Agronomy 2019, 9, 414. [Google Scholar] [CrossRef] [Green Version]
- Byrns, B.M.; Greer, K.J.; Fowler, D.B. Modeling winter survival in cereals: An interactive tool. Crop. Sci. 2020, 60, 2408–2419. [Google Scholar] [CrossRef]
- Bloomfield, M.T.; Hunt, J.R.; Trevaskis, B.; Ramm, K.; Hyles, J. Ability of alleles of PPD1 and VRN1 genes to predict flowering time in diverse Australian wheat (Triticum aestivum) cultivars in controlled environments. Crop. Pasture Sci. 2018, 69, 1061–1075. [Google Scholar] [CrossRef]
- Deng, W.; Casao, M.C.; Wang, P.; Sato, K.; Hayes, P.M.; Finnegan, E.J.; Trevaskis, B. Direct links between the vernalization response and other key traits of cereal crops. Nat. Commun. 2015, 6, 5882. [Google Scholar] [CrossRef]
- Wilcox, J.; Makowski, D. A meta-analysis of the predicted effects of climate change on wheat yields using simulation studies. Field Crop. Res. 2014, 156, 180–190. [Google Scholar] [CrossRef]
- Ugarte, C.; Calderini, D.F.; Slafer, G.A. Grain weight and grain number responsiveness to pre-anthesis temperature in wheat, barley and triticale. Field Crop. Res. 2007, 100, 240–248. [Google Scholar] [CrossRef]
- Feng, F.; Dang, P.; Pu, X.; Wen, X.; Qin, X.; Chen, Y.; Siddique, K.H.M. Contribution of Proximal and Distal Grains Within Spikelets in Relation to Yield and Yield Components in the Winter Wheat Production Region of China From 1948 to 2012. Agronomy 2019, 9, 850. [Google Scholar] [CrossRef] [Green Version]
- Blum, A. Plant Breeding for Water-Limited Environments; Springer: New York, NY, USA, 2011; pp. 53–152. [Google Scholar]
- General Directorate of Agricultural Research and Policies for Turkey. Available online: https://www.tarimorman.gov.tr/TAGEM/Sayfalar/EN/Anasayfa.aspx (accessed on 18 February 2021).
- Stoskopf, N.C.; Nathaniel, R.K.; Reinbergs, E. Comparison of Spring Wheat and Barley with Winter Wheat: Yield Components in Ontario 1. Agron. J. 1974, 66, 748–750. [Google Scholar] [CrossRef]
- Asseng, S.; Martre, P.; Maiorano, A.; Rötter, R.P.; O’Leary, G.J.; Fitzgerald, G.J.; Girousse, C.; Motzo, R.; Giunta, F.; Babar, M.A.; et al. Climate change impact and adaptation for wheat protein. Glob. Chang. Biol. 2018, 25, 155–173. [Google Scholar] [CrossRef] [Green Version]
- Turkish Statistical Institute. Available online: https://turkstatweb.tuik.gov.tr/Start.do (accessed on 18 February 2021).
- Reynolds, M.; Tattaris, M.; Cossani, C.M.; Ellis, M.; Yamaguchi-Shinozaki, K.; Pierre, C.S. Exploring Genetic Resources to Increase Adaptation of Wheat to Climate Change. In Advances in Wheat Genetics: From Genome to Field, 1st ed.; Ogihara, Y., Takumi, S., Handa, H., Eds.; Springer: Tokyo, Japan, 2015; pp. 355–368. [Google Scholar]
- Tshikunde, N.M.; Mashilo, J.; Shimelis, H.; Odindo, A. Agronomic and Physiological Traits, and Associated Quantitative Trait Loci (QTL) Affecting Yield Response in Wheat (Triticum aestivum L.): A Review. Front. Plant Sci. 2019, 10, 1428. [Google Scholar] [CrossRef] [Green Version]
- Ding, D.Y.; Feng, H.; Zhao, Y.; He, J.Q.; Zou, Y.F.; Jin, J.M. Modifying Winter Wheat Sowing Date as an Adaptation to Climate Change on the Loess Plateau. Agron. J. 2016, 108, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Zahid, A.; Ali, S.; Ahmed, M.; Iqbal, N. Improvement of Soil Health through Residue Management and Conservation Tillage in Rice-Wheat Cropping System of Punjab, Pakistan. Agronomy 2020, 10, 1844. [Google Scholar] [CrossRef]
- Zampieri, M.; Toreti, A.; Ceglar, A.; Naumann, G.; Turco, M.; Tebaldi, C. Climate resilience of the top ten wheat producers in the Mediterranean and the Middle East. Reg. Environ. Chang. 2020, 20, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Anwar, S.; Yu, S.; Sun, M.; Yang, Z.; Gao, Z.-Q. Development of Drought-Tolerant Transgenic Wheat: Achievements and Limitations. Int. J. Mol. Sci. 2019, 20, 3350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, F.G.; Rigalli, N.; Miranda, P.V.; Romagnoli, M.; Ribichich, K.F.; Trucco, F.; Portapila, M.; Otegui, M.E.; Chan, R.L. An Interdisciplinary Approach to Study the Performance of Second-generation Genetically Modified Crops in Field Trials: A Case Study With Soybean and Wheat Carrying the Sunflower HaHB4 Transcription Factor. Front. Plant Sci. 2020, 11, 178. [Google Scholar] [CrossRef] [PubMed]
- Fowler, D.B.; Byrns, B.M.; Greer, K.J. Overwinter Low-Temperature Responses of Cereals: Analyses and Simulation. Crop. Sci. 2014, 54, 2395–2405. [Google Scholar] [CrossRef] [Green Version]
- Olsen, A.K.B.; Persson, T.; De Wit, A.; Nkurunziza, L.; Sindhøj, E.; Eckersten, H. Estimating winter survival of winter wheat by simulations of plant frost tolerance. J. Agron. Crop. Sci. 2018, 204, 62–73. [Google Scholar] [CrossRef]
- Slafer, G. Wheat development: Its role in phenotyping and improving crop adaptation. In Physiological Breeding I: Interdisciplinary Approaches to Improve Crop Adaptation; Reynolds, M.P., Pask, A.J.D., Mullan, D.M., Eds.; CIMMYT: Mexico City, Mexico, 2012; pp. 109–123. Available online: https://repository.cimmyt.org/handle/10883/1287 (accessed on 18 February 2021).
- Hyles, J.; Bloomfield, M.T.; Hunt, J.R.; Trethowan, R.M.; Trevaskis, B. Phenology and related traits for wheat adaptation. Heredity 2020, 125, 417–430. [Google Scholar] [CrossRef]
- Fatima, Z.; Ahmed, M.; Hussain, M.; Abbas, G.; Ul-Allah, S.; Ahmad, S.; Ahmed, N.; Ali, M.A.; Sarwar, G.; Haque, E.U.; et al. The fingerprints of climate warming on cereal crops phenology and adaptation options. Sci. Rep. 2020, 10, 1–21. [Google Scholar] [CrossRef]
- Ploschuk, R.A.; Miralles, D.J.; Colmer, T.D.; Ploschuk, E.L.; Striker, G.G. Waterlogging of Winter Crops at Early and Late Stages: Impacts on Leaf Physiology, Growth and Yield. Front. Plant Sci. 2018, 9, 1863. [Google Scholar] [CrossRef] [Green Version]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef] [Green Version]
- Crespo-Herrera, L.A.; Singh, R.P.; Sabraoui, A.; El-Bouhssini, M. Resistance to insect pests in wheat—rye and Aegilops speltoides Tausch translocation and substitution lines. Euphytica 2019, 215, 123. [Google Scholar] [CrossRef] [Green Version]
- Basso, B.; Ritchie, J.T. Evapotranspiration in High-Yielding Maize and under Increased Vapor Pressure Deficit in the US Midwest. Agric. Environ. Lett. 2018, 3, 170039. [Google Scholar] [CrossRef]
Figure 1.
Daily precipitations recorded in May of both seasons, excluding June because of lack of precipitation (0.2 and 2.8 mm, respectively) (Table S3).
Figure 1.
Daily precipitations recorded in May of both seasons, excluding June because of lack of precipitation (0.2 and 2.8 mm, respectively) (Table S3).
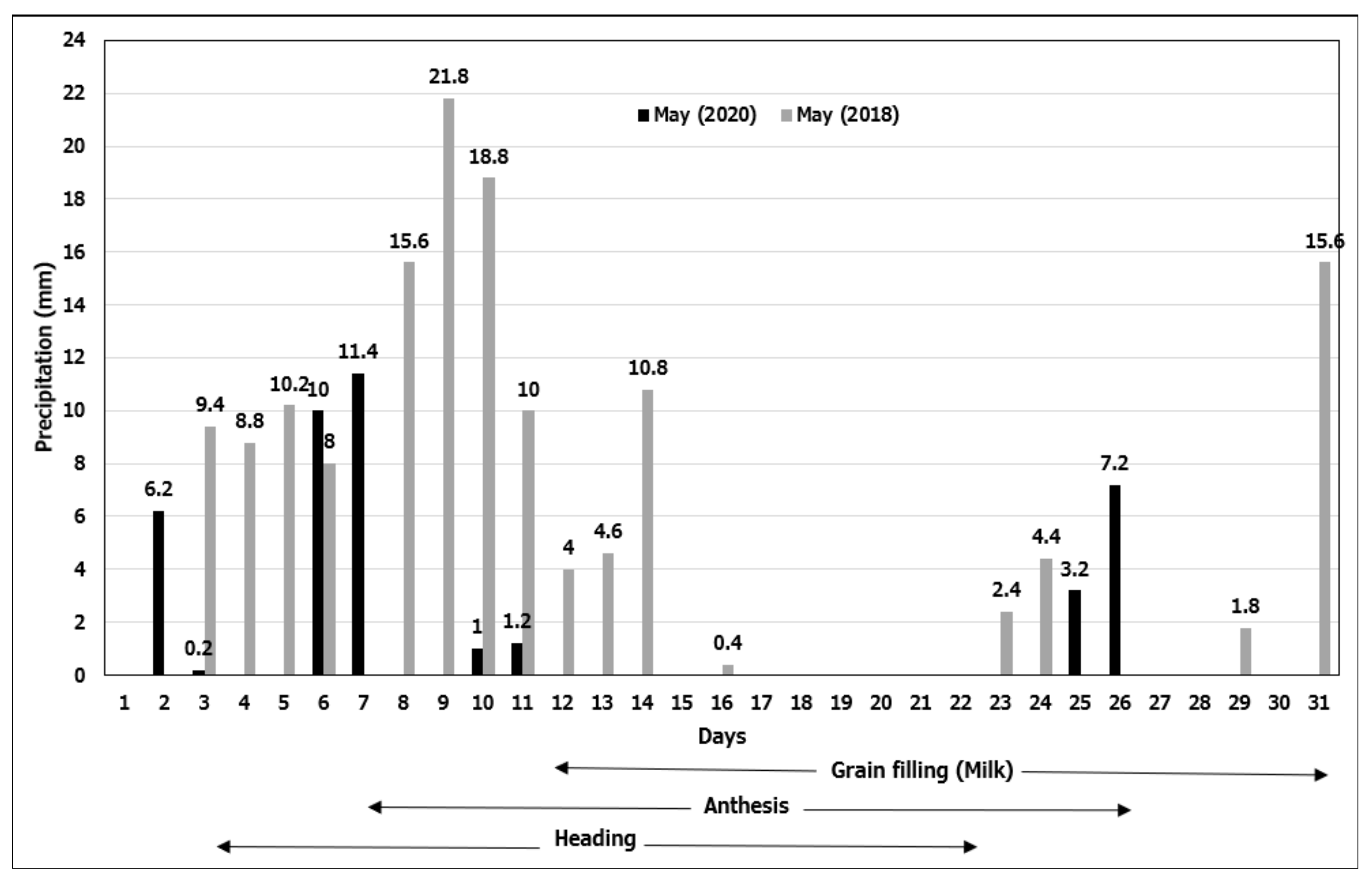
Figure 2.
Daily maximum temperatures measured in May and June of both seasons.

Figure 3.
Comparing winter wheat genotypes with spring wheat genotypes for grain yield (t ha−1) over two cropping seasons. Genotypes with the same letter are not significantly different.
Figure 3.
Comparing winter wheat genotypes with spring wheat genotypes for grain yield (t ha−1) over two cropping seasons. Genotypes with the same letter are not significantly different.

Figure 4.
Comparing winter wheat genotypes with spring wheat genotypes for days to heading over two cropping seasons. Genotypes with the same letter are not significantly different.
Figure 4.
Comparing winter wheat genotypes with spring wheat genotypes for days to heading over two cropping seasons. Genotypes with the same letter are not significantly different.

Figure 5.
Comparing winter wheat genotypes with spring wheat genotypes for harvest index (%) over two cropping seasons. Genotypes with the same letter are not significantly different.
Figure 5.
Comparing winter wheat genotypes with spring wheat genotypes for harvest index (%) over two cropping seasons. Genotypes with the same letter are not significantly different.
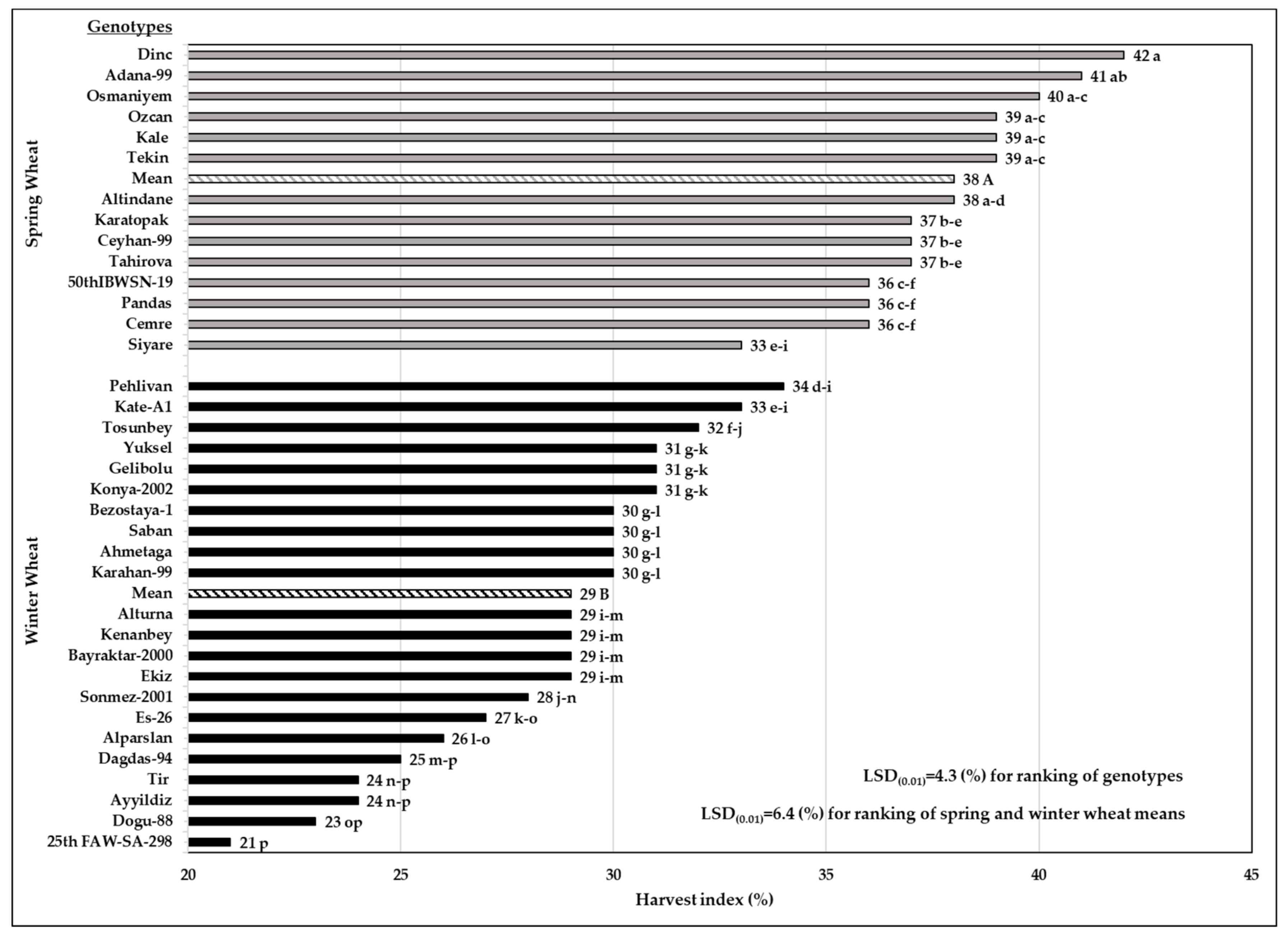
Figure 6.
Comparing winter wheat genotypes with spring wheat genotypes for plant height (cm) over two cropping seasons. Genotypes with the same letter are not significantly different.
Figure 6.
Comparing winter wheat genotypes with spring wheat genotypes for plant height (cm) over two cropping seasons. Genotypes with the same letter are not significantly different.

Figure 7.
Comparing winter wheat genotypes with spring wheat genotypes for thousand kernel weight (g) over two cropping seasons. Genotypes with the same letter are not significantly different.
Figure 7.
Comparing winter wheat genotypes with spring wheat genotypes for thousand kernel weight (g) over two cropping seasons. Genotypes with the same letter are not significantly different.

Figure 8.
Comparing winter wheat genotypes with spring wheat genotypes for grains per spike over two cropping seasons. Genotypes with the same letter are not significantly different.
Figure 8.
Comparing winter wheat genotypes with spring wheat genotypes for grains per spike over two cropping seasons. Genotypes with the same letter are not significantly different.

Figure 9.
Comparing winter wheat genotypes with spring wheat genotypes for spike sterility (%) over two cropping seasons. Genotypes with the same letter are not significantly different.
Figure 9.
Comparing winter wheat genotypes with spring wheat genotypes for spike sterility (%) over two cropping seasons. Genotypes with the same letter are not significantly different.

Figure 10.
Biplot depicting relationships between plant characteristics and genotypes. G stands for genotype codes given in Table S1.
Figure 10.
Biplot depicting relationships between plant characteristics and genotypes. G stands for genotype codes given in Table S1.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kaya, Y. Winter Wheat Adaptation to Climate Change in Turkey. Agronomy 2021, 11, 689. https://doi.org/10.3390/agronomy11040689
AMA Style
Kaya Y. Winter Wheat Adaptation to Climate Change in Turkey. Agronomy. 2021; 11(4):689. https://doi.org/10.3390/agronomy11040689
Chicago/Turabian StyleKaya, Yuksel. 2021. "Winter Wheat Adaptation to Climate Change in Turkey" Agronomy 11, no. 4: 689. https://doi.org/10.3390/agronomy11040689
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.