Winter Wheat Adaptation to Climate Change in Turkey


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Wheat Genotypes
2.3. Plant Characteristcs
2.4. Soil Properties
2.5. Crop Management
2.6. Vernalization, Photoperiod and Growing Degree Days
2.7. Comparing Winter Wheat with Spring Wheat: Relative Differences between Plant Characteristics
2.8. Statistical Analyses
3. Results
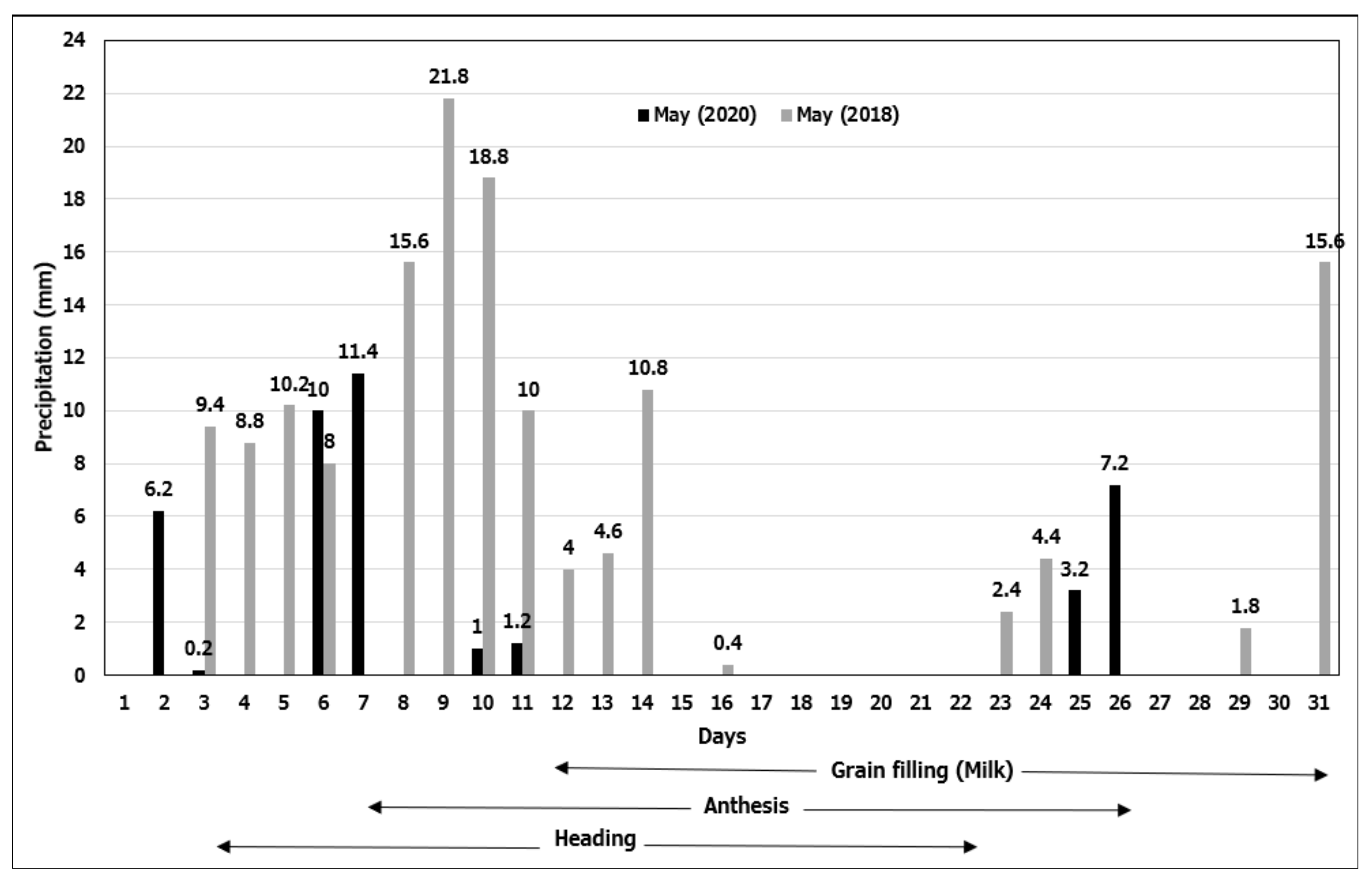
3.1. Precipitation
3.2. Temperature
3.3. Grain Yield
3.4. Days to Heading
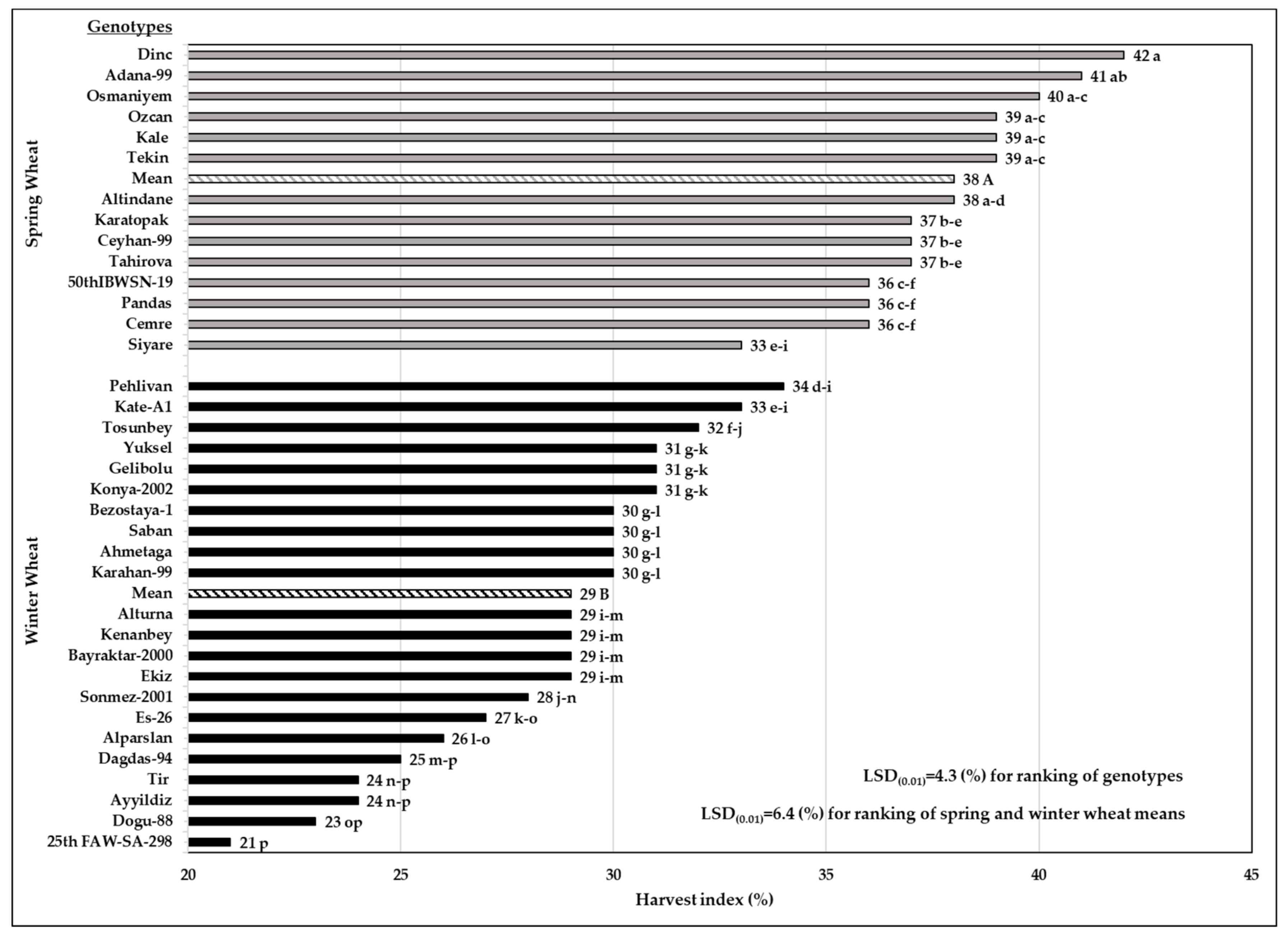
3.5. Harvest Index
3.6. Plant Height
3.7. Thousand Kernel Weight
3.8. Grains per Spike
3.9. Spike Sterility
3.10. Vernalization, Photoperiod, and Growing Degree Days
3.11. Multivariate Analyses for Plant Characteristics and Genotypes
4. Discussion
4.1. Heat and Drought Tolerance
4.2. Suggestions to Mitigate Possible Effects of Climate Change on Winter Wheat
4.2.1. Wheat Breeding Program
4.2.2. Crossing Block
4.2.3. Early Generations
4.2.4. Advanced Generations
4.2.5. Plant Characteristics Essential for Adapting to Climate Change
4.2.6. New Approaches in Wheat Agronomy for Adapting to Climate Change
4.2.7. A Particular Topic: Genetically Modified Wheat

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Characteristics | Current | Future | Reference |
|---|---|---|---|
| Vernalization | +++ | + | [53,74] |
| Photoperiod | ++ | + | [40,56] |
| Cold tolerance | +++ | + | [16,75] |
| Grain yield (Food security) | ++++ | +++++ | [55] |
| Grain quality | +++ | ++++ | [65] |
| Phenology | + | ++++ | [76,77] |
| Thermal time | +++ | + | [41,78] |
| Earliness | + | ++++ | [45] |
| Heat tolerance | ++ | +++++ | [9,10,13,17] |
| Drought tolerance | ++++ | +++++ | [14,17,19] |
| Waterlogging | + | ++ | [79] |
| Disease resistance | +++ | ++++ | [27,28] |
| Insect pest resistance | + | +++ | [80,81] |
| Water use efficiency | + | ++++ | [82] |
| CO2 use efficiency | + | +++ | [11] |
| Genetically modified wheat | ? | ? | [73] |
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Intergovernmental Panel on Climate Change. Available online: https://www.ipcc.ch/site/assets/uploads/sites/2/2019/05/SR15_Chapter2_Low_Res.pdf (accessed on 18 February 2021).
- National Centers for Environmental Information. Available online: https://www.ncdc.noaa.gov/sotc/global/202013 (accessed on 18 February 2021).
- Environmental Protection Agency. Available online: https://www.epa.gov/climate-indicators/climate-change-indicators-us-and-global-precipitation (accessed on 18 February 2021).
- Dai, A.; Zhao, T.; Chen, J. Climate Change and Drought: A Precipitation and Evaporation Perspective. Curr. Clim. Chang. Rep. 2018, 4, 301–312. [Google Scholar] [CrossRef]
- Turkish State Meteorological Service. Available online: https://www.mgm.gov.tr/eng/yearly-climate.aspx (accessed on 18 February 2021).
- Climate Change and Its Projections with New Scenarios in Turkey (in Turkish). Available online: https://mgm.gov.tr/FILES/iklim/iklim-degisikligi-projeksiyon2015.pdf (accessed on 18 February 2021).
- Demircan, M.; Gürkan, H.; Eskioğlu, O.; Arabacı, H.; Coşkun, M. Climate Change Projections for Turkey: Three Models and Two Scenarios. Turk. J. Water Sci. Manag. 2017, 1, 22–43. [Google Scholar] [CrossRef]
- Hütsch, B.W.; Jahn, D.; Schubert, S. Grain yield of wheat (Triticum aestivum L.) under long-term heat stress is sink-limited with stronger inhibition of kernel setting than grain filling. J. Agron. Crop. Sci. 2018, 205, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Balla, K.; Karsai, I.; Bónis, P.; Kiss, T.; Berki, Z.; Horváth, Á.; Mayer, M.; Bencze, S.; Veisz, O. Heat stress responses in a large set of winter wheat cultivars (Triticum aestivum L.) depend on the timing and duration of stress. PLoS ONE 2019, 14, e0222639. [Google Scholar] [CrossRef]
- Thistlethwaite, R.J.; Tan, D.K.; Bokshi, A.I.; Ullah, S.; Trethowan, R.M. A phenotyping strategy for evaluating the high-temperature tolerance of wheat. Field Crop. Res. 2020, 255, 107905. [Google Scholar] [CrossRef]
- Sabella, E.; Aprile, A.; Negro, C.; Nicolì, F.; Nutricati, E.; Vergine, M.; Luvisi, A.; De Bellis, L. Impact of Climate Change on Durum Wheat Yield. Agronomy 2020, 10, 793. [Google Scholar] [CrossRef]
- Sattar, A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Rizwan, M.S.; Hussain, M.; Jabran, K.; Cheema, M.A. Terminal drought and heat stress alter physiological and biochemical attributes in flag leaf of bread wheat. PLoS ONE 2020, 15, e0232974. [Google Scholar] [CrossRef]
- Schmidt, J.; Claussen, J.; Wörlein, N.; Eggert, A.; Fleury, D.; Garnett, T.; Gerth, S. Drought and heat stress tolerance screening in wheat using computed tomography. Plant Methods 2020, 16, 1–12. [Google Scholar] [CrossRef]
- Schmidt, J.; Garcia, M.; Brien, C.; Kalambettu, P.; Garnett, T.; Fleury, D.; Tricker, P.J. Transcripts of wheat at a target locus on chromosome 6B associated with increased yield, leaf mass and chlorophyll index under combined drought and heat stress. PLoS ONE 2020, 15, e0241966. [Google Scholar] [CrossRef]
- Wang, H.; Lemke, R.; Goddard, T.; Sprout, C. Tillage and root heat stress in wheat in central Alberta. Can. J. Soil Sci. 2007, 87, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Braun, H.J.; Payne, T. Mega-Environment breeding. In Physiological Breeding I: Interdisciplinary Approaches to Improve Crop Adaptation; Reynolds, M.P., Pask, A.J.D., Mullan, D.M., Eds.; CIMMYT: Mexico City, Mexico, 2012; pp. 6–17. Available online: https://repository.cimmyt.org/handle/10883/1287 (accessed on 18 February 2021).
- Snowdon, R.J.; Wittkop, B.; Chen, T.-W.; Stahl, A. Crop adaptation to climate change as a consequence of long-term breeding. Theor. Appl. Genet. 2020. [Google Scholar] [CrossRef]
- Hochman, Z.; Gobbett, D.L.; Horan, H. Climate trends account for stalled wheat yields in Australia since 1990. Glob. Chang. Biol. 2017, 23, 2071–2081. [Google Scholar] [CrossRef]
- Fábián, A.; Sáfrán, E.; Szabó-Eitel, G.; Barnabás, B.; Jäger, K. Stigma Functionality and Fertility Are Reduced by Heat and Drought Co-stress in Wheat. Front. Plant Sci. 2019, 10, 244. [Google Scholar] [CrossRef]
- Vanli, Ö.; Ustundag, B.B.; Ahmad, I.; Hernandez-Ochoa, I.M.; Hoogenboom, G. Using crop modeling to evaluate the impacts of climate change on wheat in southeastern turkey. Environ. Sci. Pollut. Res. 2019, 26, 29397–29408. [Google Scholar] [CrossRef]
- Yeşilköy, S.; Şaylan, L. Yields and Water Footprints of Sunflower and Winter Wheat under Different Climate Projections. J. Clean. Prod. 2021, 298, 126780. [Google Scholar] [CrossRef]
- Yang, C.; Fraga, H.; Van Ieperen, W.; Trindade, H.; Santos, J.A. Effects of climate change and adaptation options on winter wheat yield under rainfed Mediterranean conditions in southern Portugal. Clim. Chang. 2019, 154, 159–178. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Villegas, J.; Milan, A.M.; Alexandrov, N.; Asseng, S.; Challinor, A.J.; Crossa, J.; Van Eeuwijk, F.; Ghanem, M.E.; Grenier, C.; Heinemann, A.B.; et al. CGIAR modeling approaches for resource-constrained scenarios: I. Accelerating crop breeding for a changing climate. Crop. Sci. 2020, 60, 547–567. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, H.; Bentley, A. Changing times: Opportunities for altering winter wheat phenology. Plants People Planet 2021, 3, 113–123. [Google Scholar] [CrossRef]
- Hunt, J.R.; Lilley, J.M.; Trevaskis, B.; Flohr, B.M.; Peake, A.; Fletcher, A.; Zwart, A.B.; Gobbett, D.; Kirkegaard, J.A. Early sowing systems can boost Australian wheat yields despite recent climate change. Nat. Clim. Chang. 2019, 9, 244–247. [Google Scholar] [CrossRef]
- Karapinar, B.; Özertan, G. Yield implications of date and cultivar adaptation to wheat phenological shifts: A survey of farmers in Turkey. Clim. Chang. 2020, 158, 453–472. [Google Scholar] [CrossRef]
- Turgay, E.B.; Buyuk, O.; Olmez, F.; Yildirim, A.F. Determination of prevalence of the Septoria leaf blotch disease of wheat [Zymoseptoria tritici (Desm. Quaedvlieg & Crous)] in Central Anatolia and its molecular identification. Plant Prot. Bull. 2016, 56, 283–293. [Google Scholar]
- Impact of Climate Change on Wheat Production in Kentucky. Available online: http://dx.doi.org/10.13023/PSSRR.2014.3 (accessed on 18 February 2021).
- Zhao, Z.; Rebetzke, G.J.; Zheng, B.; Chapman, S.C.; Wang, E. Modelling impact of early vigour on wheat yield in dryland regions. J. Exp. Bot. 2019, 70, 2535–2548. [Google Scholar] [CrossRef] [PubMed]
- Zeleke, K. Simulating Agronomic Adaptation Strategies to Mitigate the Impacts of Climate Change on Wheat Yield in South-Eastern Australia. Agronomy 2021, 11, 337. [Google Scholar] [CrossRef]
- Dubey, R.; Pathak, H.; Chakrabarti, B.; Singh, S.; Gupta, D.K.; Harit, R. Impact of terminal heat stress on wheat yield in India and options for adaptation. Agric. Syst. 2020, 181, 102826. [Google Scholar] [CrossRef]
- Dowla, M.N.U.; Edwards, I.; O’Hara, G.; Islam, S.; Ma, W. Developing Wheat for Improved Yield and Adaptation Under a Changing Climate: Optimization of a Few Key Genes. Engineering 2018, 4, 514–522. [Google Scholar] [CrossRef]
- Morgounov, A.; Ozdemir, F.; Keser, M.; Akin, B.; Payne, T.; Braun, H.-J. International Winter Wheat Improvement Program: History, activities, impact and future. Front. Agric. Sci. Eng. 2019, 6, 240. [Google Scholar] [CrossRef] [Green Version]
- Hanson, H.; Borlaug, N.E.; Anderson, R.G. Wheat in the Third World; Westview Press: Boulder, CO, USA, 1982; p. 174. [Google Scholar]
- Graf, R.; Thomas, J.; Gaudet, D.; Laroche, A.; Beres, B. Broadview hard red winter wheat. Can. J. Plant Sci. 2012, 92, 177–181. [Google Scholar] [CrossRef] [Green Version]
- International Winter Wheat Improvement Program. Available online: http://www.iwwip.org/Nursery/Details/1009 (accessed on 18 February 2021).
- International Maize and Wheat Improvement Center. Available online: https://data.cimmyt.org/dataset.xhtml?persistentId=hdl:11529/10548147 (accessed on 18 February 2021).
- Pask, A.J.D.; Pietragalla, J.; Mullan, D.M.; Reynolds, M.P. Physiological Breeding II: A Field Guide to Wheat Phenotyping; CIMMYT: Mexico City, Mexico, 2012; Available online: https://repository.cimmyt.org/handle/10883/1288?show=full (accessed on 18 February 2021).
- Andeden, E.E.; Yediay, F.; Baloch, F.S.; Shaaf, S.; Kilian, B.; Nachit, M.; Ozkan, H. Distribution of vernalization and photoperiod genes (Vrn-A1, Vrn-B1, Vrn-D1, Vrn-B3, Ppd-D1)in Turkish bread wheat cultivars and landraces. Cereal Res. Commun. 2011, 39, 352–364. [Google Scholar] [CrossRef]
- Dubcovsky, J.; Loukoianov, A.; Fu, D.; Valarik, M.; Sanchez, A.; Yan, L. Effect of Photoperiod on the Regulation of Wheat Vernalization Genes VRN1 and VRN2. Plant Mol. Biol. 2006, 60, 469–480. [Google Scholar] [CrossRef] [Green Version]
- McMaster, G.S. Development of the wheat plant. In Wheat: Science and Trade; Carver, B.F., Ed.; Wiley-Blackwell: Ames, IA, USA, 2009; pp. 31–55. Available online: https://doi.org/10.1002/9780813818832 (accessed on 18 February 2021).
- Almeida, A.S.; Macas, B.; Rodrigues, V.L.G.; Torrao, M.M. The history of wheat breeding in Portugal. In The World Wheat Book: The History of Wheat Breeding; Bonjean, A.P., Angus, W.S., van Ginkel, M., Eds.; Lavoisier: Paris, France, 2016; Volume 3, pp. 93–126. Available online: https://www.lavoisier.fr/livre/agriculture/the-world-wheat-book/bonjean/descriptif-9782743020910 (accessed on 18 February 2021).
- Lipkovich, I.A.; Smith, E.P. Biplot and Singular Value Decomposition Macros forExcel. J. Stat. Softw. 2002, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Spring Freeze Injury to Kansas Wheat. Available online: https://bookstore.ksre.ksu.edu/pubs/C646.PDF (accessed on 18 February 2021).
- Ochagavía, H.; Prieto, P.; Zikhali, M.; Griffiths, S.; Slafer, G.A. Earliness Per Se by Temperature Interaction on Wheat Development. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cook, J.P.; Heo, H.-Y.; Varella, A.C.; Lanning, S.P.; Blake, N.K.; Sherman, J.D.; Martín, J.M.; See, D.; Chao, S.; Talbert, L.E. Evaluation of a QTL Mapping Population Composed of Hard Red Spring and Winter Wheat Alleles Using Various Marker Platforms. Crop. Sci. 2018, 58, 701–712. [Google Scholar] [CrossRef]
- Huang, M.; Mheni, N.; Brown-Guedira, G.; McKendry, A.; Griffey, C.; Van Sanford, D.; Costa, J.; Sneller, C. Genetic analysis of heading date in winter and spring wheat. Euphytica 2018, 214, 128. [Google Scholar] [CrossRef]
- Würschum, T.; Langer, S.M.; Longin, C.F.H. Genetic control of plant height in European winter wheat cultivars. Theor. Appl. Genet. 2015, 128, 865–874. [Google Scholar] [CrossRef]
- Fiore, M.C.; Mercati, F.; Spina, A.; Blangiforti, S.; Venora, G.; Dell’Acqua, M.; Lupini, A.; Preiti, G.; Monti, M.; Pè, M.E.; et al. High-Throughput Genotype, Morphology, and Quality Traits Evaluation for the Assessment of Genetic Diversity of Wheat Landraces from Sicily. Plants 2019, 8, 116. [Google Scholar] [CrossRef] [Green Version]
- Nuttall, J.G.; Barlow, K.M.; Delahunty, A.J.; Christy, B.P.; O’Leary, G.J. Acute High Temperature Response in Wheat. Agron. J. 2018, 110, 1296–1308. [Google Scholar] [CrossRef] [Green Version]
- Ren, S.; Qin, Q.; Ren, H.; Sui, J.; Zhang, Y. Heat and Drought Stress Advanced Global Wheat Harvest Timing from 1981–2014. Remote Sens. 2019, 11, 971. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Asseng, S.; Müller, C.; Ewert, F.; Elliott, J.; Lobell, D.B.; Martres, P.; Ruane, A.C.; Wallach, D.; Jones, J.W.; et al. Similar estimates of temperature impacts on global wheat yield by three independent methods. Nat. Clim. Chang. 2016, 6, 1130–1136. [Google Scholar] [CrossRef]
- Yan, L.; Li, G.; Yu, M.; Fang, T.; Cao, S.; Carver, B.F. Genetic Mechanisms of Vernalization Requirement Duration in Winter Wheat Cultivars. In Advances in Wheat Genetics: From Genome to Field, 1st ed.; Ogihara, Y., Takumi, S., Handa, H., Eds.; Springer: Tokyo, Japan, 2015; pp. 117–125. [Google Scholar]
- Gol, L.; Tomé, F.; Von Korff, M. Floral transitions in wheat and barley: Interactions between photoperiod, abiotic stresses, and nutrient status. J. Exp. Bot. 2017, 68, 1399–1410. [Google Scholar] [CrossRef]
- El Hassouni, K.; Belkadi, B.; Filali-Maltouf, A.; Tidiane-Sall, A.; Al-Abdallat, A.; Nachit, M.; Bassi, F.M. Loci Controlling Adaptation to Heat Stress Occurring at the Reproductive Stage in Durum Wheat. Agronomy 2019, 9, 414. [Google Scholar] [CrossRef] [Green Version]
- Byrns, B.M.; Greer, K.J.; Fowler, D.B. Modeling winter survival in cereals: An interactive tool. Crop. Sci. 2020, 60, 2408–2419. [Google Scholar] [CrossRef]
- Bloomfield, M.T.; Hunt, J.R.; Trevaskis, B.; Ramm, K.; Hyles, J. Ability of alleles of PPD1 and VRN1 genes to predict flowering time in diverse Australian wheat (Triticum aestivum) cultivars in controlled environments. Crop. Pasture Sci. 2018, 69, 1061–1075. [Google Scholar] [CrossRef]
- Deng, W.; Casao, M.C.; Wang, P.; Sato, K.; Hayes, P.M.; Finnegan, E.J.; Trevaskis, B. Direct links between the vernalization response and other key traits of cereal crops. Nat. Commun. 2015, 6, 5882. [Google Scholar] [CrossRef]
- Wilcox, J.; Makowski, D. A meta-analysis of the predicted effects of climate change on wheat yields using simulation studies. Field Crop. Res. 2014, 156, 180–190. [Google Scholar] [CrossRef]
- Ugarte, C.; Calderini, D.F.; Slafer, G.A. Grain weight and grain number responsiveness to pre-anthesis temperature in wheat, barley and triticale. Field Crop. Res. 2007, 100, 240–248. [Google Scholar] [CrossRef]
- Feng, F.; Dang, P.; Pu, X.; Wen, X.; Qin, X.; Chen, Y.; Siddique, K.H.M. Contribution of Proximal and Distal Grains Within Spikelets in Relation to Yield and Yield Components in the Winter Wheat Production Region of China From 1948 to 2012. Agronomy 2019, 9, 850. [Google Scholar] [CrossRef] [Green Version]
- Blum, A. Plant Breeding for Water-Limited Environments; Springer: New York, NY, USA, 2011; pp. 53–152. [Google Scholar]
- General Directorate of Agricultural Research and Policies for Turkey. Available online: https://www.tarimorman.gov.tr/TAGEM/Sayfalar/EN/Anasayfa.aspx (accessed on 18 February 2021).
- Stoskopf, N.C.; Nathaniel, R.K.; Reinbergs, E. Comparison of Spring Wheat and Barley with Winter Wheat: Yield Components in Ontario 1. Agron. J. 1974, 66, 748–750. [Google Scholar] [CrossRef]
- Asseng, S.; Martre, P.; Maiorano, A.; Rötter, R.P.; O’Leary, G.J.; Fitzgerald, G.J.; Girousse, C.; Motzo, R.; Giunta, F.; Babar, M.A.; et al. Climate change impact and adaptation for wheat protein. Glob. Chang. Biol. 2018, 25, 155–173. [Google Scholar] [CrossRef] [Green Version]
- Turkish Statistical Institute. Available online: https://turkstatweb.tuik.gov.tr/Start.do (accessed on 18 February 2021).
- Reynolds, M.; Tattaris, M.; Cossani, C.M.; Ellis, M.; Yamaguchi-Shinozaki, K.; Pierre, C.S. Exploring Genetic Resources to Increase Adaptation of Wheat to Climate Change. In Advances in Wheat Genetics: From Genome to Field, 1st ed.; Ogihara, Y., Takumi, S., Handa, H., Eds.; Springer: Tokyo, Japan, 2015; pp. 355–368. [Google Scholar]
- Tshikunde, N.M.; Mashilo, J.; Shimelis, H.; Odindo, A. Agronomic and Physiological Traits, and Associated Quantitative Trait Loci (QTL) Affecting Yield Response in Wheat (Triticum aestivum L.): A Review. Front. Plant Sci. 2019, 10, 1428. [Google Scholar] [CrossRef] [Green Version]
- Ding, D.Y.; Feng, H.; Zhao, Y.; He, J.Q.; Zou, Y.F.; Jin, J.M. Modifying Winter Wheat Sowing Date as an Adaptation to Climate Change on the Loess Plateau. Agron. J. 2016, 108, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Zahid, A.; Ali, S.; Ahmed, M.; Iqbal, N. Improvement of Soil Health through Residue Management and Conservation Tillage in Rice-Wheat Cropping System of Punjab, Pakistan. Agronomy 2020, 10, 1844. [Google Scholar] [CrossRef]
- Zampieri, M.; Toreti, A.; Ceglar, A.; Naumann, G.; Turco, M.; Tebaldi, C. Climate resilience of the top ten wheat producers in the Mediterranean and the Middle East. Reg. Environ. Chang. 2020, 20, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Anwar, S.; Yu, S.; Sun, M.; Yang, Z.; Gao, Z.-Q. Development of Drought-Tolerant Transgenic Wheat: Achievements and Limitations. Int. J. Mol. Sci. 2019, 20, 3350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, F.G.; Rigalli, N.; Miranda, P.V.; Romagnoli, M.; Ribichich, K.F.; Trucco, F.; Portapila, M.; Otegui, M.E.; Chan, R.L. An Interdisciplinary Approach to Study the Performance of Second-generation Genetically Modified Crops in Field Trials: A Case Study With Soybean and Wheat Carrying the Sunflower HaHB4 Transcription Factor. Front. Plant Sci. 2020, 11, 178. [Google Scholar] [CrossRef] [PubMed]
- Fowler, D.B.; Byrns, B.M.; Greer, K.J. Overwinter Low-Temperature Responses of Cereals: Analyses and Simulation. Crop. Sci. 2014, 54, 2395–2405. [Google Scholar] [CrossRef] [Green Version]
- Olsen, A.K.B.; Persson, T.; De Wit, A.; Nkurunziza, L.; Sindhøj, E.; Eckersten, H. Estimating winter survival of winter wheat by simulations of plant frost tolerance. J. Agron. Crop. Sci. 2018, 204, 62–73. [Google Scholar] [CrossRef]
- Slafer, G. Wheat development: Its role in phenotyping and improving crop adaptation. In Physiological Breeding I: Interdisciplinary Approaches to Improve Crop Adaptation; Reynolds, M.P., Pask, A.J.D., Mullan, D.M., Eds.; CIMMYT: Mexico City, Mexico, 2012; pp. 109–123. Available online: https://repository.cimmyt.org/handle/10883/1287 (accessed on 18 February 2021).
- Hyles, J.; Bloomfield, M.T.; Hunt, J.R.; Trethowan, R.M.; Trevaskis, B. Phenology and related traits for wheat adaptation. Heredity 2020, 125, 417–430. [Google Scholar] [CrossRef]
- Fatima, Z.; Ahmed, M.; Hussain, M.; Abbas, G.; Ul-Allah, S.; Ahmad, S.; Ahmed, N.; Ali, M.A.; Sarwar, G.; Haque, E.U.; et al. The fingerprints of climate warming on cereal crops phenology and adaptation options. Sci. Rep. 2020, 10, 1–21. [Google Scholar] [CrossRef]
- Ploschuk, R.A.; Miralles, D.J.; Colmer, T.D.; Ploschuk, E.L.; Striker, G.G. Waterlogging of Winter Crops at Early and Late Stages: Impacts on Leaf Physiology, Growth and Yield. Front. Plant Sci. 2018, 9, 1863. [Google Scholar] [CrossRef] [Green Version]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef] [Green Version]
- Crespo-Herrera, L.A.; Singh, R.P.; Sabraoui, A.; El-Bouhssini, M. Resistance to insect pests in wheat—rye and Aegilops speltoides Tausch translocation and substitution lines. Euphytica 2019, 215, 123. [Google Scholar] [CrossRef] [Green Version]
- Basso, B.; Ritchie, J.T. Evapotranspiration in High-Yielding Maize and under Increased Vapor Pressure Deficit in the US Midwest. Agric. Environ. Lett. 2018, 3, 170039. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaya, Y. Winter Wheat Adaptation to Climate Change in Turkey. Agronomy 2021, 11, 689. https://doi.org/10.3390/agronomy11040689
Kaya Y. Winter Wheat Adaptation to Climate Change in Turkey. Agronomy. 2021; 11(4):689. https://doi.org/10.3390/agronomy11040689
Chicago/Turabian StyleKaya, Yuksel. 2021. "Winter Wheat Adaptation to Climate Change in Turkey" Agronomy 11, no. 4: 689. https://doi.org/10.3390/agronomy11040689
APA StyleKaya, Y. (2021). Winter Wheat Adaptation to Climate Change in Turkey. Agronomy, 11(4), 689. https://doi.org/10.3390/agronomy11040689