
State of the Art Management of the Central American Locust Schistocerca piceifrons piceifrons (Walker, 1870)

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Life History of the Central American Locust
3. Habitat and Ecology
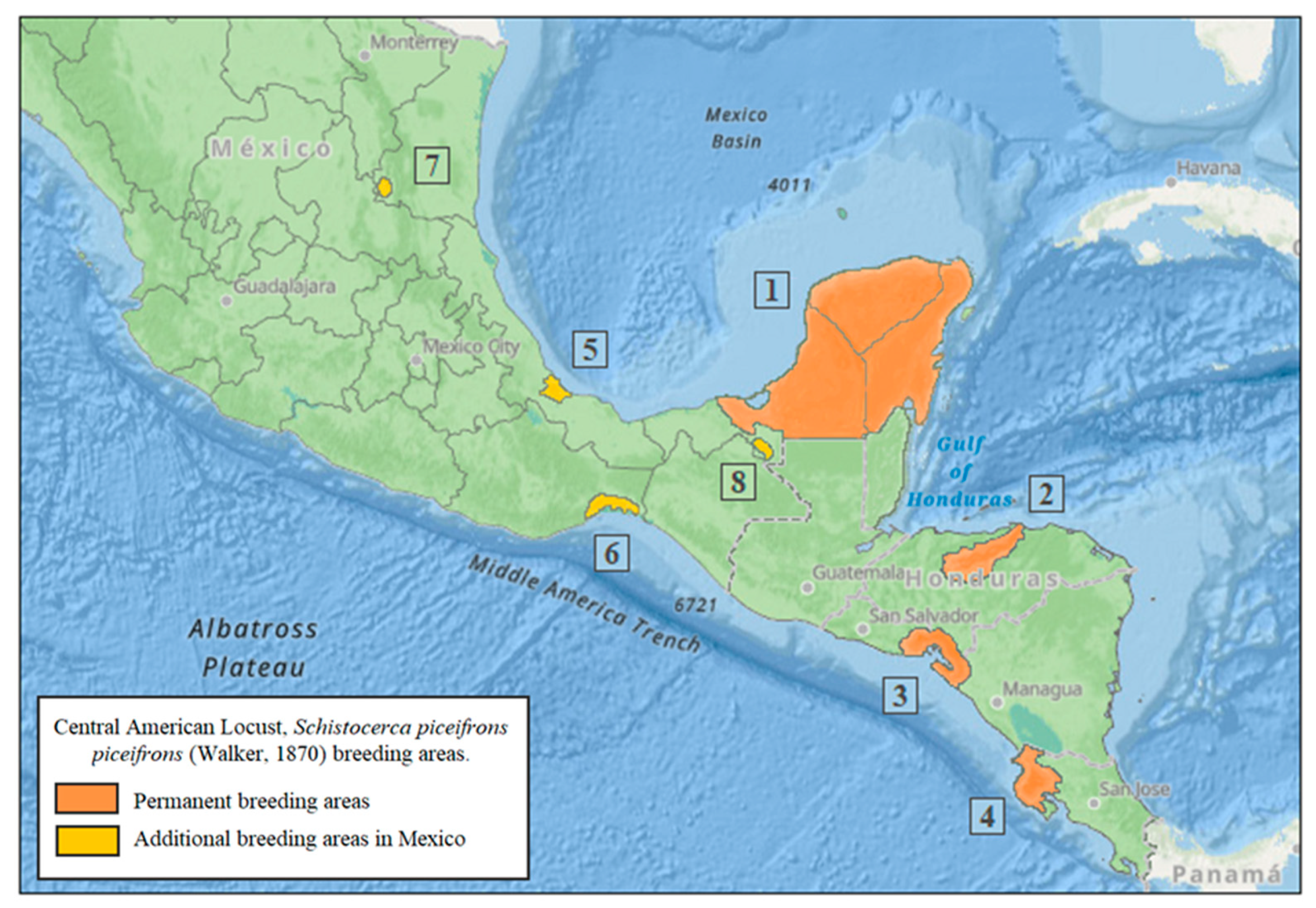
4. Permanent Breeding Areas
5. Band and Swarm Formation
6. Phase Polyphenism, Gregarization, and Migration
7. Central American Locust Outbreaks
8. Central American Locust Management Strategies
9. Control Measures
10. Current Research
11. Future Work
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flores-Granados, F. Las Plagas de Langosta en el Área Maya: Ambiente e Historia de Una Calamidad en la Época Prehispánica. Península 2011, 6, 27–46. Available online: https://www.redalyc.org/pdf/3583/358333208002.pdf (accessed on 10 January 2021).
- García-Quintanilla, A. La Langosta, los Mayas y el Colonialismo en Yucatán, México, 1883. Relaciones 2012, 129, 215–249. Available online: http://www.scielo.org.mx/pdf/rz/v33n129/v33n129a7.pdf (accessed on 10 January 2021).
- García-Quintanilla, A. Saak’ y el Retorno del Fin del Mundo: La Plaga de Langosta en las Profecías del Katun 13 Ahau. Ancient Mesoamerica 2005, 16, 327–344. Available online: https://www.jstor.org/stable/26309188?seq=1 (accessed on 10 January 2021). [CrossRef]
- Barrientos-Lozano, L.; Astacio-Cabrera, O.; Poot-Pech, M.; Cruz-Martínez, O. Manual Técnico Sobre la Langosta Voladora (Schistocerca Piceifrons Walker 1870) y Otros Acridoideos de Centro América y Sureste de México, 1st ed.; Fao-Oirsa: San Salvador, El Salvador, 1992; pp. 1–162. [Google Scholar]
- Song, H.; Weissman, D.B.; Barrientos-Lozano, L.; Cano-Santana, Z. The Locust Island. Am. Entomol. 2009, 52, 168–181. [Google Scholar] [CrossRef] [Green Version]
- Song, H. Density-dependent phase polyphenism in nonmodel locusts: A minireview. Psyche 2011. [Google Scholar] [CrossRef] [Green Version]
- Senasica-DGSV. Langosta Centroamericana [Schistocerca piceifrons piceifrons (Walker, 1870)] (Orthoptera: Acrididae), Ficha Técnica; National Service of Agrifood Health, Safety and Quality-General Directorate of Plant Health-National Center for Phytosanitary Reference-Phytosanitary Specialist Group: Ciudad de México, Mexico, 2016; pp. 1–18. Available online: https://www.gob.mx/cms/uploads/attachment/file/157817/Ficha_t_cnica_langosta.pdf (accessed on 11 January 2021).
- Cullen, D.A.; Cease, A.J.; Latchininsky, A.V.; Ayali, A.; Berry, K.; Buhl, J.; de Keyser, R.; Foquet, B.; Hadrich, J.C.; Matheson, T.; et al. From molecules to management: Mechanisms and consequences of locust phase polyphenism. Adv. Insect Phys. 2017, 53, 169–285. [Google Scholar] [CrossRef]
- Ávila-Valdez, J.; Barrientos-Lozano, L.; García-Salazar, P. Aspectos Bio-Ecológicos de la Langosta Centroamericana en el Sur de Tamaulipas. In Simposio Control Biológico y Manejo de la Langosta Centroamericana Schistocerca piceifrons piceifrons (Walker); Instituto Tecnológico de Cuidad Victoria: Ciudad Victoria, Tamaulipas, Mexico, 2007; pp. 1–21. [Google Scholar]
- Pflüger, H.J.; Bräunig, P. One hundred years of phase polymorphism research in locusts. J. Comp. Physiol. 2021, 207, 321–326. [Google Scholar] [CrossRef]
- Astacio-Cabrera, O. La Langosta Voladora o Chapulín Schistocerca piceifrons piceifrons (Walker) en Centroamérica, 1st ed.; Organismo Internacional Regional de Sanidad Agropecuaria (OIRSA): San Salvador, El Salvador, 1990; pp. 1–42. [Google Scholar]
- Hunter-Jones, P. Life history of the Central American Locust, Schistocerca sp. (Orthoptera: Acrididae), in the laboratory. Ann. Entomol. Soc. Am. 1967, 60, 468–477. [Google Scholar] [CrossRef]
- Harvey, A.W. Schistocerca piceifrons (Walker) (Orthoptera: Acrididae), the swarming locust of tropical America: A review. Bull. Entomol. Res. 1983, 73, 171–184. [Google Scholar] [CrossRef]
- Ortiz-Yam, I.; Zuleta, M.C. Asuntos de vecinos: Langosta, defensa agrícola y la construcción de la sanidad vegetal en México y Centroamérica, siglo XX. Historia Méxicana 2019, 70, 313–373. [Google Scholar] [CrossRef]
- Conteras-Servín, C.; Magaña-Ortíz, C. Ficha Técnica de la Langosta Centroamericana, Schistocerca piceifrons piceifrons (Walker). In La Plaga de la Langosta (Schistocerca piceifrons piceifrons Walker) Una Visión Multidisciplinaria Desde la Perspectiva del Desastre Fitosanitario en México, 1st ed.; Galindo Mendoza, G., Contreras Servín, C., Ibarra Zapata, E., Eds.; Universidad Autónoma de San Luis Potosí: San Luis Potosí, Mexico, 2013; Volume 1, pp. 17–36. [Google Scholar]
- Barrientos-Lozano, L. Taller Sobre Control. In Biológico y Manejo de Langosta Centroamericana (Schistocerca Piceifrons Piceifrons, Walker), 1st ed.; Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria-Instituto Tecnológico de Ciudad Victoria: Ciudad Victoria, Tamaulipas, Mexico, 2007; pp. 1–197. [Google Scholar]
- Rodríguez-Absi, J.; Almaguer-Sierra, P.; Barrientos-Lozano, L.; Rodríguez-Fuentes, H. Thermal time clock for estimating phenological development of Schistocerca piceifrons piceifrons Walker (Orthoptera: Acrididae) in northeastern Mexico. J. Orthoptera Res. 2009, 18, 65–73. [Google Scholar] [CrossRef]
- Hernández-Zul, M.I.; Quijano-Carranza, J.A.; Yañez-López, R.; Ocampo-Velazquez, R.V.; Torres-Pacheco, I.; Guevara-Gonzalez, R.G.; Castro-Ramírez, E. Dynamic simulation model of Central American Locust Schistocerca piceifrons (Orthoptera: Acrididae). Fla. Entomol. 2013, 96, 1274–1283. [Google Scholar] [CrossRef]
- Galindo-Mendoza, M.G.; Ibarra-Zapata, E.; Hipólito-Cruz, G.; Aldama-Aguilera, C. Sistema de Alerta Temprana Para el Control de la Langosta Centroamericana (Schistocerca piceifrons piceifrons (Walker) en la Península De Yucatán Aplicando Modelos Multicriteria e Imágenes de Satélite NOAA-AVHRR. In La Plaga de la Langosta, Schistocerca piceifrons piceifrons (Walker), una Visión Multidisciplinaria Desde la Perspectiva del Desastre Fitosanitario en México, 1st ed.; Universidad Autónoma de San Luis Potosí: San Luis Potosí, Mexico, 2013; pp. 173–215. [Google Scholar]
- Senasica-DGSV. Manual Operativo de la Campaña Contra la Langosta Centroamericana [Schistocerca piceifrons piceifrons (Walker, 1870)] (Orthoptera: Acrididae), 1st ed.; Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria–Dirección General de Sanidad Vegetal: Ciudad de México, Mexico, 2019; pp. 1–80. Available online: https://www.gob.mx/cms/uploads/attachment/file/483901/Manual_Langosta_Junio_2019.pdf (accessed on 15 January 2021).
- Organismo Internacional Regional de Sanidad Agropecuaria (OIRSA). Plan de Acción de Manejo de la Langosta Centroamericana, 1st ed.; Secretaría General del Sistema de la Integración Centroamericana: San Salvador, El Salvador, 2020; pp. 1–80. Available online: https://www.oirsa.org/contenido/2020/(17%20marzo2020)%20Plan%20de%20accio%CC%81n%20y%20atencio%CC%81n%20de%20brotes%20corregido%202.pdf (accessed on 15 January 2021).
- Poot-Pech, M.A.; Ruiz-Sánchez, E.; Ballina-Gómez, H.S.; Gamboa-Angulo, M.M.; Reyes-Ramírez, A. Olfactory response and host plant feeding of the Central American Locust Schistocerca piceifrons piceifrons Walker, to common plants in a gregarious zone. Neotrop. Entomol. 2016, 45, 382–388. [Google Scholar] [CrossRef]
- Poot-Pech, M.A.; Ruiz-Sánchez, E.; Gamboa-Angulo, M.; Ballina-Gómez, H.S.; Reyes-Ramírez, A. Population fluctuation of Schistocerca piceifrons piceifrons (Orthoptera: Acrididae) in the Yucatán peninsula and its relation with the environmental conditions. Rev. Biol. Trop. 2018, 66, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Ramírez, R.; Torres-Castillo, J.A.; Barrientos-Lozano, L.; Almaguer-Sierra, P.; Torres-Acosta, R.I. Schistocerca piceifrons piceifrons (Orthoptera: Acrididae) as a source of compounds of biotechnological and nutritional interest. J. Insect Sci. 2019, 19, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viesca-González, F.C.; Romero-Contreras, A.T. La entomofagia en México. Algunos aspectos culturales. El Periplo Sustentable 2009, 16, 57–83. Available online: https://www.redalyc.org/articulo.oa?id=193414421003 (accessed on 16 January 2021). [CrossRef] [Green Version]
- Garza-Sánchez, J. Evaluación del Potencial Alimenticio de la Langosta Centroamericana, Schistocerca piceifrons piceifrons, (Walker, 1870), (Orthoptera: Acrididae). Master’s Thesis, Tecnológico Nacional de México-Instituto Tecnológico de Cd. Victoria, Victoria, Tamaulipas, Mexico, 21 October 2020. [Google Scholar]
- Barrientos-Lozano, L. Population Dynamics, Biology and Ecology of the Central American Locust (Schistocerca piceifrons piceifrons, Walker) in Southern Mexico. In Proceedings of the Metaleptea—Eight International Meeting of the Orthopterists’ Society, Montpellier, France, 19–22 August 2001; International Conference of Orthopteroid Insects. p. 75. [Google Scholar]
- Barrientos-Lozano, L. Ecología, Manejo y Control de la Langosta Voladora, Schistocerca piceifrons piceifrons (Walker). In Proceedings of the Primer Curso Internacional: Ecología, Manejo y Control de la Langosta Voladora, Schistocerca piceifrons piceifrons (Walker), Altamira, Tamaulipas, México, 5–7 November 2001; Barrientos-Lozano, L., Ed.; Dinámica Impresa S.A.: Cuidad Victoria, Tamaulipas, Mexico, 2001; pp. 1–232. [Google Scholar]
- Ávila-Valdez, J.; Barrientos-Lozano, L.; García-Salazar, P. Biología y comportamiento de la langosta centroamericana, Schistocerca piceifrons piceifrons (Walker, 1870), en el Sur de Tamaulipas. In Proceedings of the 2do Curso Internacional: Manejo Integrado de la Langosta Centroamericana y Acridoideos Plaga en América Latina, Cuidad Victoria, Tamaulipas, Mexico, 27–29 June 2005; Barrientos-Lozano, L., Almaguer-Sierra, P., Eds.; Dinámica Impresa S.A.: Cuidad Victoria, Tamaulipas, Mexico, 2005; pp. 31–35. [Google Scholar]
- Cano-Santana, Z. Ecología e Historia Natural de Schistocerca americana socorro y S. piceifrons piceifrons en Isla Socorro; Informe Final Proyecto BS007; Universidad Nacional Autónoma de México: Ciudad de México, México, 2006; pp. 1–138. Available online: http://www.conabio.gob.mx/institucion/proyectos/resultados/InfBS007.pdf (accessed on 1 February 2021).
- Poot-Pech, M.A. La Langosta Voladora Schistocerca piceifrons (Orthoptera: Acrididae), Hacia un Manejo Sustentable. In El Patrimonio, su Importancia y Conservación: Conociendo el Patrimonio, 1st ed.; TECCIS: Ciudad de Campeche, Campeche, Mexico, 2016; pp. 58–66. Available online: https://www.researchgate.net/profile/Marvel-Valencia-Gutierrez-publication/329761423GUAYA_Talisia_olivaeformis_R_AL_RESCATE_DEUN_PATRIMONIO_NATURAL/links/5c1985e7a6fdccfc70587065/GUAYA-Talisia-olivaeformis-R-AL-RESCATE-DEUN-PATRIMONIO-NATURAL.pdf (accessed on 4 February 2021).
- Galindo-Mendoza, M.G.; Contreras Servín, C. Alerta Climática-Fitosanitaria Para: Langosta Centroamericana, Schistocerca piceifrons piceifrons (Walker, 1870), Zona Huasteca, San Luis Potosí. In Sistemas de Alerta Temprana en Actividades Agropecuarias con Tecnología Espacial; SENASICA-UASL: San Luis Potosí, Mexico, 2021. [Google Scholar]
- Barrientos-Lozano, L. Ortópteros Plaga de México y Centro América: Guía de Campo, 1st ed.; Instituto Tecnológico de Cd. Victoria, COSNET, SEP-CONACYT: Cuidad Victoria, Tamaulipas, Mexico, 2003; pp. 1–114. [Google Scholar]
- Garza-Urbina, E. La langosta Schistocerca piceifrons piceifrons y su Manejo en la Planicie Huasteca; Folleto Técnico No. 12; Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), CIRNE: San Luís Potosí, Mexico, 2005; pp. 1–23. Available online: http://www.inifapcirne.gob.mx/Biblioteca/Publicaciones/149.pdf (accessed on 4 February 2021).
- Senasica-DGSV. Memoria del Curso Regional Sobre la Langosta (Schistocerca piceifrons piceifrons); Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria-Dirección General de Sanidad Vegetal, CIRNE, INIFAP, SAGARPA: San Luis Potosí, Mexico, 2003; pp. 1–93.
- Retana, B.J.A. Relación Entre Algunos Aspectos Climatológicos y el Desarrollo de la Langosta Centroamericana Schistocerca piceifrons piceifrons en el Pacífico Norte de Costa Rica Durante la Fase Cálida del Fenómeno El Niño-Oscilación Sur (ENOS). Tóp. Meteorol. Oceanogr. 2000, 7, 73–87. Available online: http://www.bio-nica.info/biblioteca/retana2000acrididae.pdf (accessed on 4 February 2021).
- Poot-Pech, M.A.; Ruiz-Sánchez, E.; Ballina-Gómez, H. Indicators Plants in Solitary Phase and Migration Behavior of Schistocerca Piceifrons in Yucatán, México. Metaleptea 2017, 37, 21. [Google Scholar]
- Rainey, R. Meteorology and the Migration of Desert Locusts: Applications of Synoptic Meteorology in Locust Control; Technical Note No. 54; World Meteorological Organization (WMO): Geneva, Switzerland, 1988; pp. 1–115. Available online: https://library.wmo.int/doc_num.php?explnum_id=1746 (accessed on 4 February 2021).
- Márquez-Delgado, A. La Lucha Contra la Langosta en México, 1st ed.; Colegio de Ingenieros Agrónomos: Ciudad de México, Mexico, 1963; pp. 1–220. Available online: https://agris.fao.org/agris-search/search.do?recordID=US201300278309 (accessed on 4 February 2021).
- Hoffmann, C.C.; Dampf, A.; Varela, G. Informe de la Comisión Científica Exploradora de la Plaga de la Langosta en el Estado de Veracruz, 1st ed.; Talleres Gráficos de México: Ciudad de México, Mexico, 1925; Volume 3, pp. 1–140.
- Ávila-Valdez, J.; Barrientos-Lozano, L.; García-Salazar, P. Aspectos de la Biología y hábitos de la langosta centroamericana, Schistocerca piceifrons piceifrons (Walker) (Orthoptera: Acrididae) bajo las condiciones del Sur de Tamaulipas. Entomol. Mex. 2007, 3, 177–181. [Google Scholar]
- Sword, G.A.; Simpson, S.J.; El Hadi, O.M.; Wilps, H. Density-dependent aposematism in the desert locust. Proc. Roy. Soc. B Biol. Sci. 2000, 267, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Pener, M.P.; Simpson, S.J. Locust phase polyphenism: An update. Adv. Insect Physiol. 2009, 36, 1–286. [Google Scholar] [CrossRef]
- Song, H.; Foquet, B.; Mariño-Pérez, R.; Woller, D.A. Phylogeny of locusts and grasshoppers reveals complex evolution of density-dependent phenotypic plasticity. Sci. Rep. 2017, 7, 6606. [Google Scholar] [CrossRef] [Green Version]
- Simpson, S.J.; Sword, G.A. Phase Polyphenism in Locusts: Mechanisms, Population Consequences, Adaptive Significance and Evolution. In Phenotypic Plasticity of Insects Mechanisms and Consequences; Whitman, D.W., Ananthakrishnan, T.N., Eds.; Science Publishers: Enfield, NH, USA, 2009; pp. 147–189. [Google Scholar]
- Contreras-Servín, C. Conexión Climática del Fenómeno del Niño con la Plaga de la Langosta en Centroamérica (Schistocerca piceifrons piceifrons, Walker) Localizada en el Estado de Yucatán y la Huasteca Potosina. Entomol. Mex. 2009, 8, 34–351. Available online: http://www.entomologia.socmexent.org/revista/2009/BHN/347-351.pdf (accessed on 5 February 2021).
- Senasica-DGSV. Central American Locust; Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria-Dirección General de Sanidad Vegetal: Ciudad de México, México, 2018; pp. 1–2. Available online: https://issuu.com/senasica/docs/info_senasica_no_2 (accessed on 8 February 2021).
- Hernández-Velázquez, V.M.; Berlanga-Padilla, A.M.; Barrientos-Lozano, L. Vegetable and mineral oil formulations of Metarhizium anisopliae var. acridum to control the Central American Locust Schistocerca piceifrons piceifrons (Walker, 1870) (Orthoptera: Acrididae). J. Orthop. Res. 2000, 9, 223–227. [Google Scholar] [CrossRef]
- Song, H.; Poot-Pech, M. Taller regional de manejo de la langosta centroamericana, Schistocerca Piceifrons. Metaleptea 2017, 37, 16–17. [Google Scholar]
- Organismo Internacional Regional de Sanidad Agropecuaria (OIRSA). Situación Actual y Recomendaciones para el Control de la Langosta Centroamericana en los Países de la Región OIRSA, 1st ed.; Secretaría General del Sistema de la Integración Centroamericana: San Salvador, El Salvador, 2020; pp. 1–14. Available online: https://www.sica.int/documentos/situacion-actual-y-recomendaciones-para-el-control-de-la-langosta-centroamericana-en-los-paises-de-la-region-oirsa_1_122911.html (accessed on 9 February 2021).
- Galván-Martínez, E.; Almaguer-Sierra, P.; Barrientos-Lozano, L. Dinámica poblacional de la langosta centroamericana, Schistocerca piceifrons piceifrons (Walker 1870), usando un modelo de simulación. Entomol. Mex. 2008, 7, 285–289. Available online: http://www.socmexent.org/entomologia/revista/2008/BHN/285-289.pdf (accessed on 10 February 2021).
- Senasica-DGSV. Langosta Centroamericana. Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria-Dirección General de Sanidad Vegetal. 2020. Available online: https://www.gob.mx/senasica/documentos/langosta-centroamericana-1109199 (accessed on 10 February 2021).
- Barrientos-Lozano, L.; Hernández-Velázquez, V.M.; Milner, R.J.; Hunter, D.M. Advances in biological control of locusts and grasshoppers in México. J. Orthop. Res. 2002, 11, 77–82. [Google Scholar] [CrossRef]
- Hernández-Velázquez, V.M.; Berlanga-Padilla, A.M.; Garza-González, E. Detección de Metarhizium flavoviride sobre Schistocerca piceifrons piceifrons (Orthoptera: Acrididae) en la Isla Socorro, Archipiélago de Revillagigedo, México. Vedalia 1997, 4, 45–46. [Google Scholar]
- Barrientos-Lozano, L.; Milner, R.J. Comparison of the Pathogenicity of Three Isolates of Metarhizium anisopliae var. acridum, Effect of Dose and Temperature. In Proceedings of the XXI International Congress of Entomology, Foz de Iguaçu, Brazil, 20–26 August 2000; Sociedade Entomológica do Brasil: Londrina, PR, Brazil, 2000; pp. 513–514. [Google Scholar]
- Hernández-Velázquez, V.M.; Hunter, D.M.; Barrientos-Lozano, L.; Lezama-Gutiérrez, R.; Reyes-Villanueva, F. Susceptibility of Schistocerca piceifrons (Orthoptera: Acrididae) to Metarhizium anisopliae var. acridum (Deuteromycotina: Hyphomycetes): Laboratory and field trials. J. Orthop. Res. 2003, 12, 89–92. [Google Scholar] [CrossRef]
- Milner, R.J.; Barrientos-Lozano, L.; Driver, F.; Hunter, D.M. A comparative study of two Mexican isolates with an Australian isolate of Metarhizium anisopliae var. acridum strain characterization, temperature profile and virulence for wingless grasshopper, Phaulacridium vittatum. BioControl 2003, 48, 335–348. [Google Scholar] [CrossRef]
- Barrientos-Lozano, L.; Hunter, D.M.; Ávila-Valdéz, J.; García-Salazar, P.; Hernández-Velázquez, V.M. Control Biológico de la Langosta Voladora (Schistocerca piceifrons Walker) con Metarhizum anisopliae var. acridum en el Noreste de México. In Proceedings of the XXVII Congreso Nacional de Control Biológico, Los Mochis, Sinaloa, México, 8–13 November 2004; Sociedad Mexicana de Control Biológico: Sinaloa, Mexico, 2004; pp. 137–141. Available online: https://www.cabi.org/wp-content/uploads/Barrientos-Lozano-2004-Biological-control-of-Schistocerca.pdf (accessed on 12 February 2021).
- Barrientos-Lozano, L.; Hunter, D.M.; Ávila-Valdéz, J.; García-Salazar, P.; Horta-Vega, J.V. Control Biológico de la Langosta centroamericana Schistocerca piceifrons piceifrons Walker (Orthoptera: Acrididae) en el Noreste de México. Vedalia 2005, 12, 119–128. [Google Scholar]
- Williams, T.; Arredondo-Bernal, H.C.; Rodríguez-del-Bosque, L.A. Biological Pest Control in Mexico. Annu. Rev. Entomol. 2013, 58, 119–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrientos-Lozano, L. Manejo Integrado de la Langosta Centroamericana (Schistocerca piceifrons piceifrons, Walker) y Acridoideos Plaga en América Latina, 1st ed.; Tecnológico Nacional de México-Instituto Tecnológico de Cuidad Victoria: Cuidad Victoria, Tamaulipas, Mexico, 2005; pp. 1–303. [Google Scholar]
- Barrientos-Lozano, L. Memoria Simposio Control. In Biológico y Manejo de la Langosta Centroamericana Schistocerca piceifrons piceifrons (Walker); Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria-Instituto Tecnológico de Cuidad Victoria: Cuidad Victoria, Tamaulipas, Mexico, 2007; pp. 1–112. [Google Scholar]
- Montelongo-Ruíz, G.; Olazarán-Santibáñez, F.E.; Reyes-Zepeda, F.; Torres-Castillo, J.A.; De la Rosa-Manzano, E.; Martínez-Ávalos, J.G.; Arellano-Méndez, L.U.; Mora-Olivo, A. Inhibition of intestinal proteolytic activity of Schistocerca piceifrons piceifrons (Walker) and Pterophylla beltrani (Bolívar and Bolívar) using extracts from Chamaedorea radicalis Mart. Fruit. Southwest Entomol. 2020, 45, 137–146. [Google Scholar] [CrossRef]
- Le Gall, M.; Overson, R.; Cease, A. A global review on locusts (Orthoptera: Acrididae) and their interactions with livestock grazing practices. Front. Ecol. Evol. 2019, 7, 1–24. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrientos-Lozano, L.; Song, H.; Rocha-Sánchez, A.Y.; Torres-Castillo, J.A. State of the Art Management of the Central American Locust Schistocerca piceifrons piceifrons (Walker, 1870). Agronomy 2021, 11, 1024. https://doi.org/10.3390/agronomy11061024
Barrientos-Lozano L, Song H, Rocha-Sánchez AY, Torres-Castillo JA. State of the Art Management of the Central American Locust Schistocerca piceifrons piceifrons (Walker, 1870). Agronomy. 2021; 11(6):1024. https://doi.org/10.3390/agronomy11061024
Chicago/Turabian StyleBarrientos-Lozano, Ludivina, Hojun Song, Aurora Y. Rocha-Sánchez, and Jorge Ariel Torres-Castillo. 2021. "State of the Art Management of the Central American Locust Schistocerca piceifrons piceifrons (Walker, 1870)" Agronomy 11, no. 6: 1024. https://doi.org/10.3390/agronomy11061024
APA StyleBarrientos-Lozano, L., Song, H., Rocha-Sánchez, A. Y., & Torres-Castillo, J. A. (2021). State of the Art Management of the Central American Locust Schistocerca piceifrons piceifrons (Walker, 1870). Agronomy, 11(6), 1024. https://doi.org/10.3390/agronomy11061024