
Ammonium Excess Leads to Ca Restrictions, Morphological Changes, and Nutritional Imbalances in Tomato Plants, Which Can Be Monitored by the N/Ca Ratio

 ,
,  ,
,  and
and 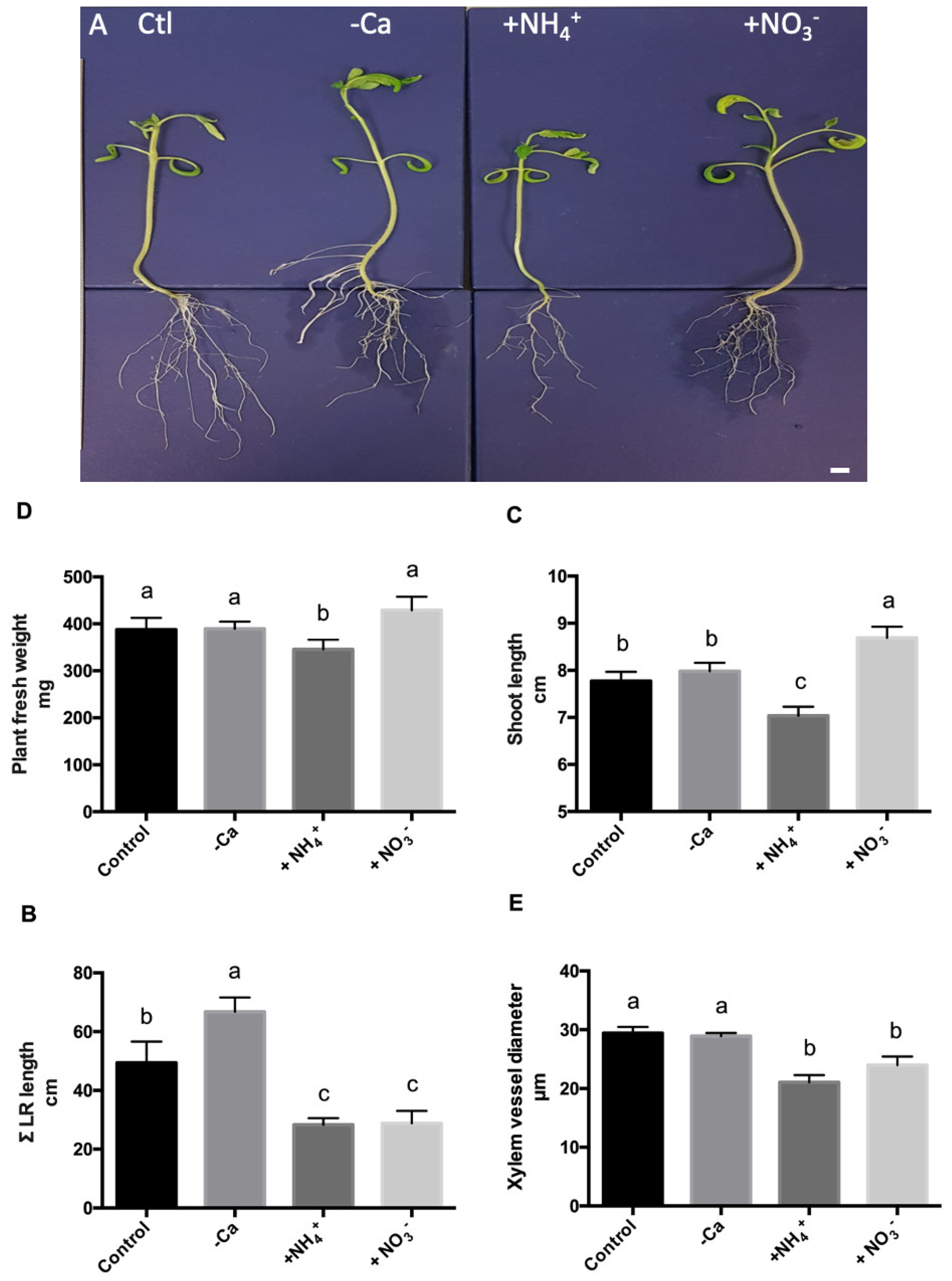{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material and Growth Conditions
2.2. Phenotypic Analysis
2.3. Microscopy Analysis
2.4. Mineral Analysis
2.5. RNA Extraction and cDNA Synthesis
2.6. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.7. Statistical Analysis
3. Results and Discussion
3.1. Calcium Deficiency and Nitrogen Excess Affects Plant Development In Vitro
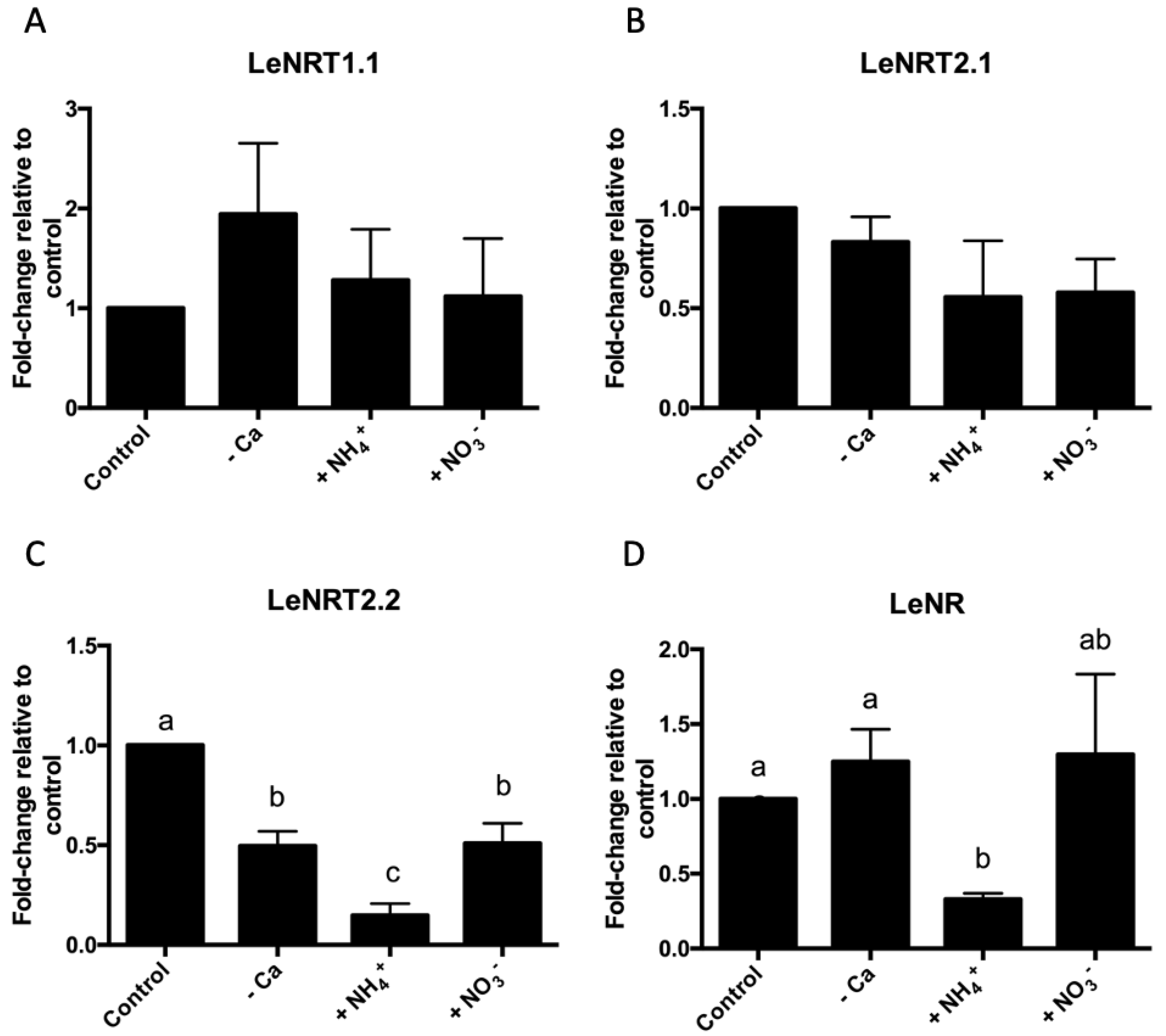
3.2. Calcium Deficit and Ammonium Excess Triggers Changes in Nitrate Transporters Expression and Nitrogen Assimilation
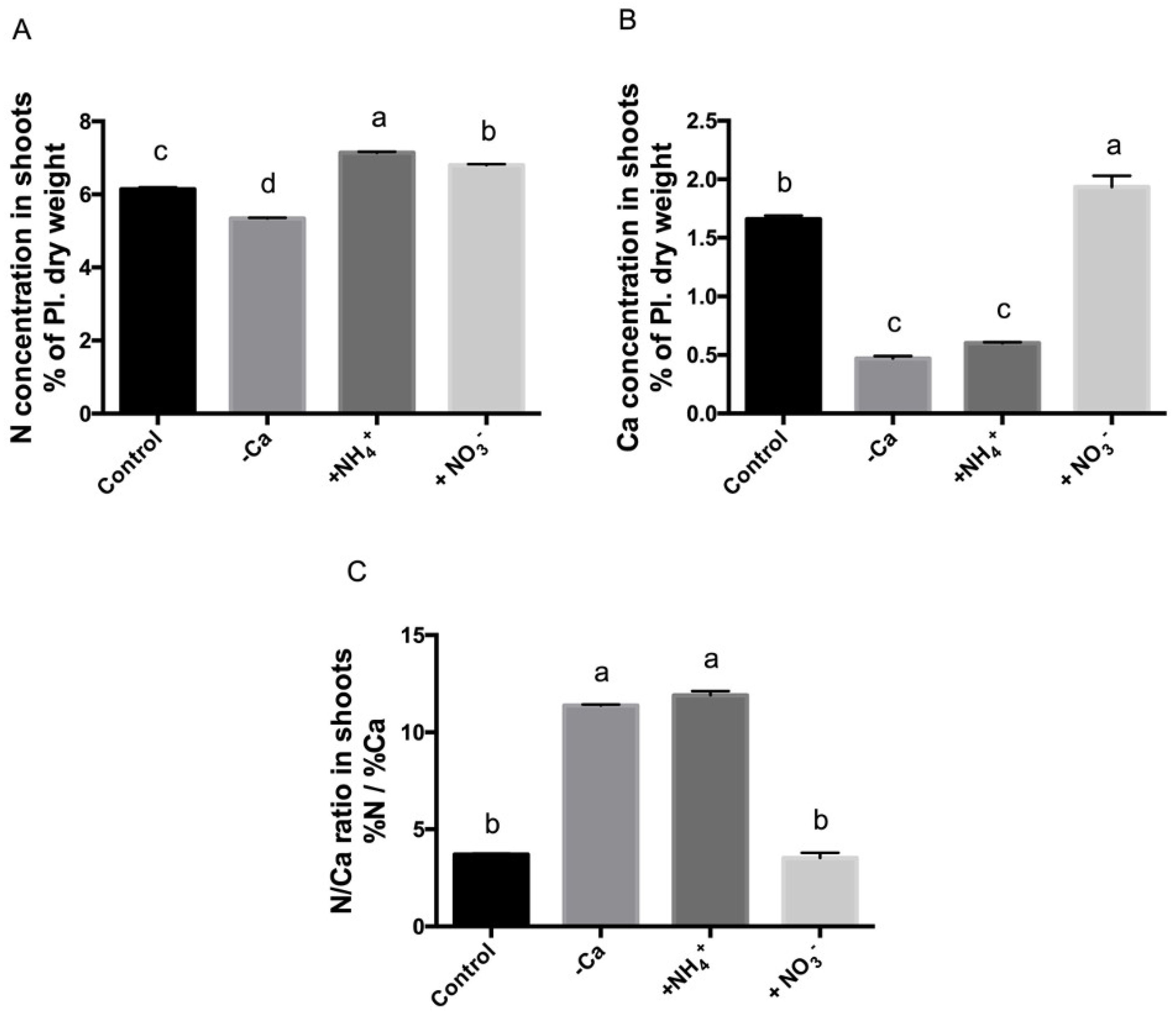
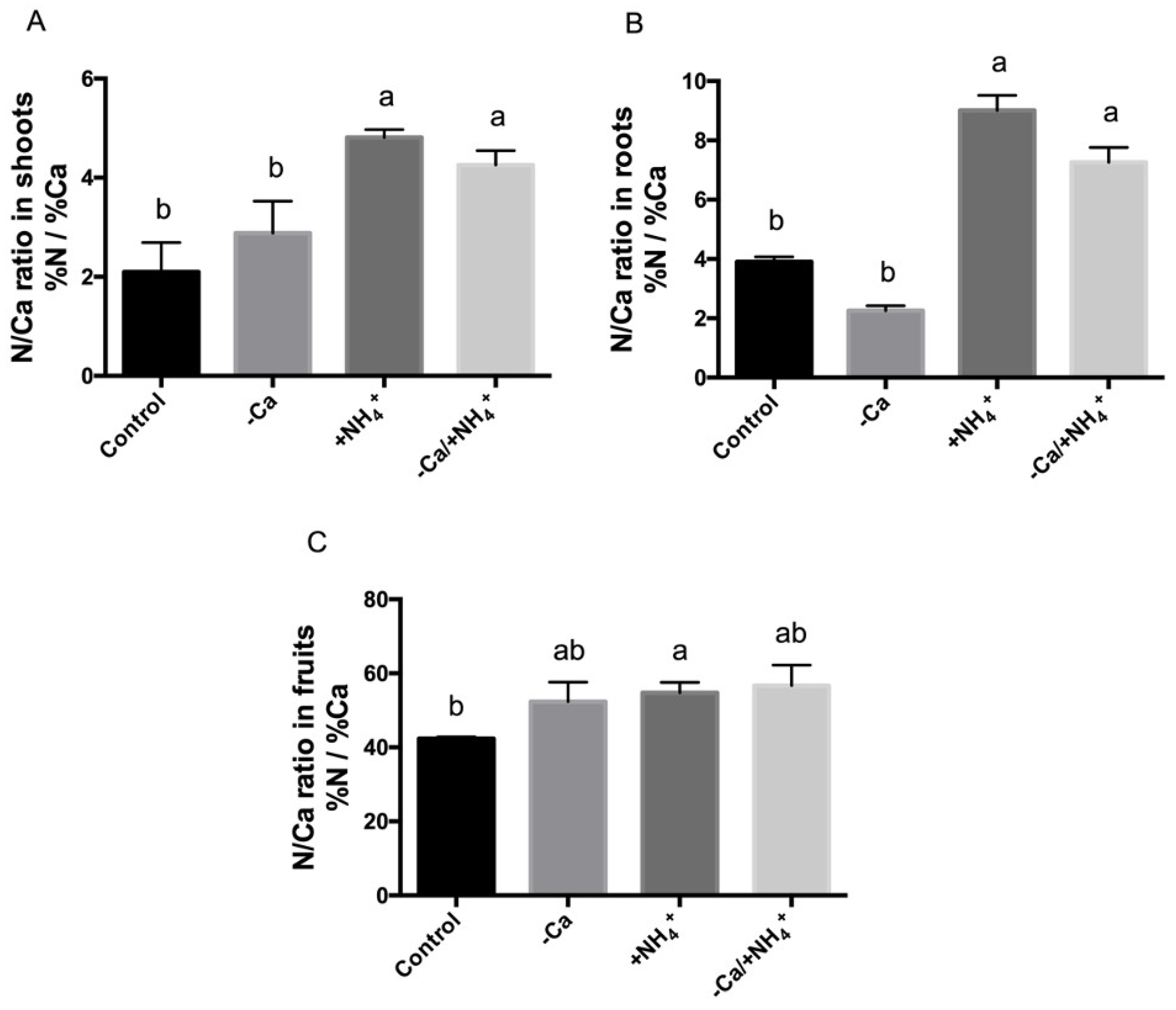
3.3. Calcium Deficit and Ammonium Excess Results in Nutrient Imbalance and an Altered N/Ca Ratio
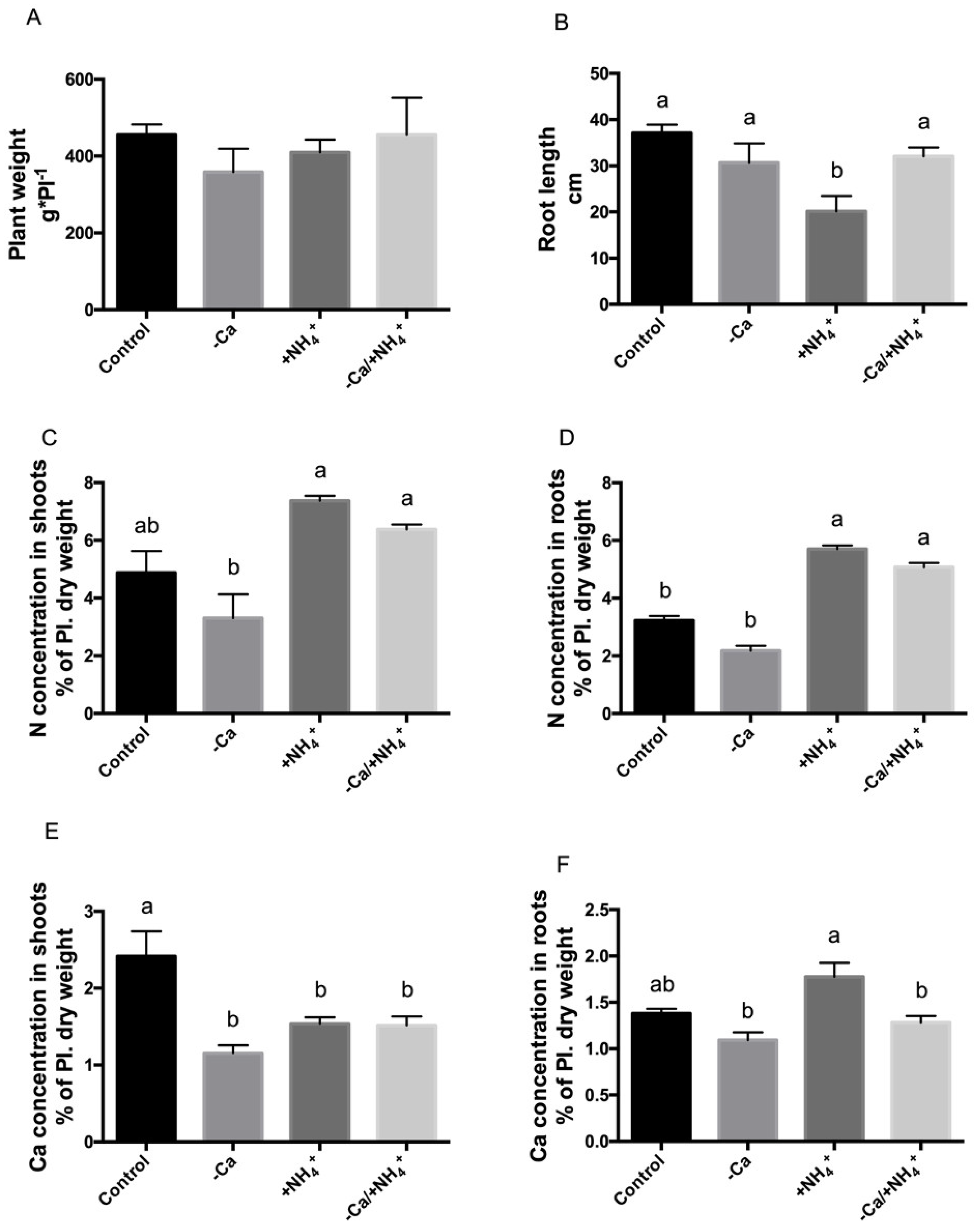
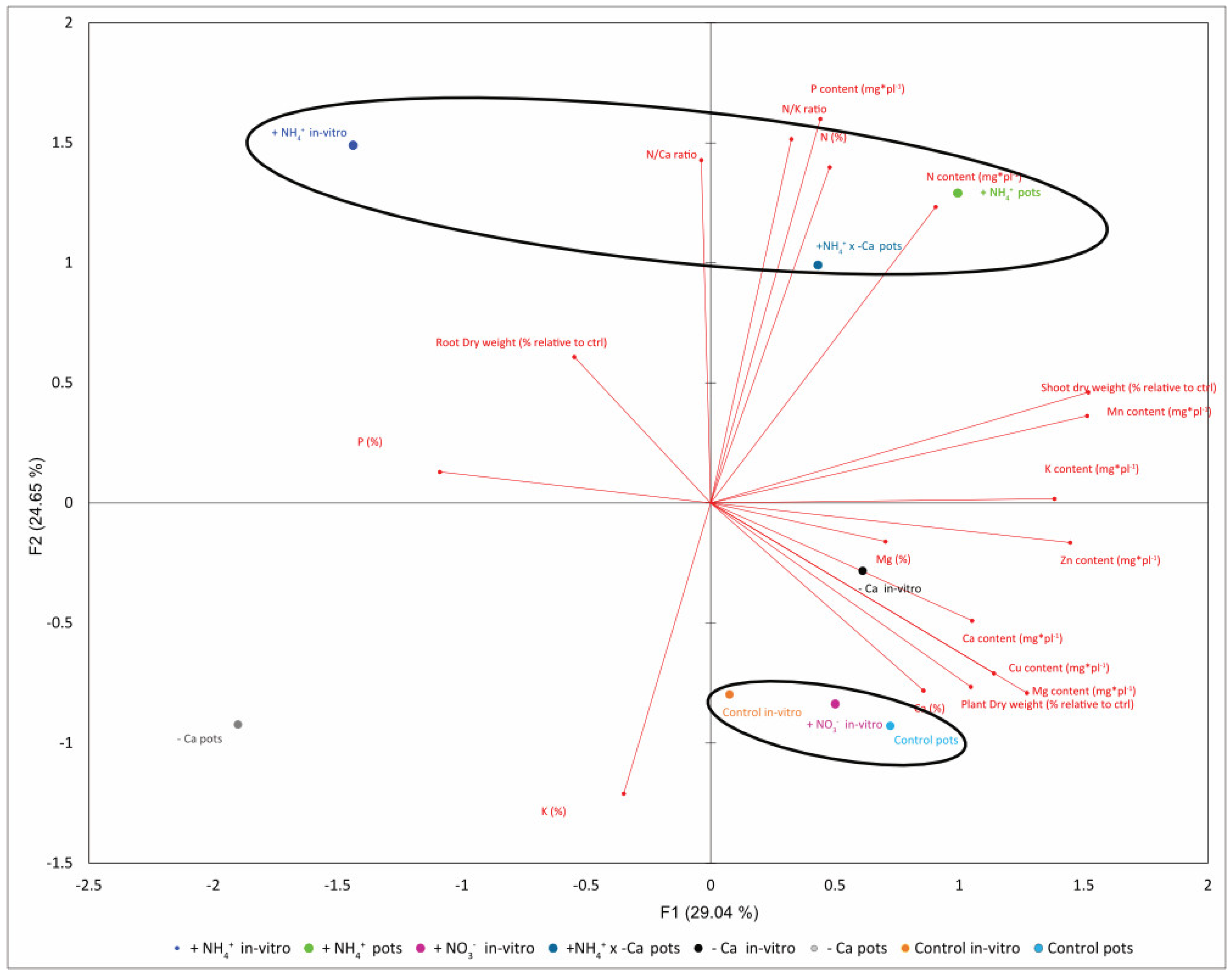
3.4. Physiological Changes in Response to Calcium Deficit and Ammonium Excess in Potted Plants Correlate with Treatments In Vitro
3.5. Ammonium Excess Correlates with N/Ca Ratio and N Content
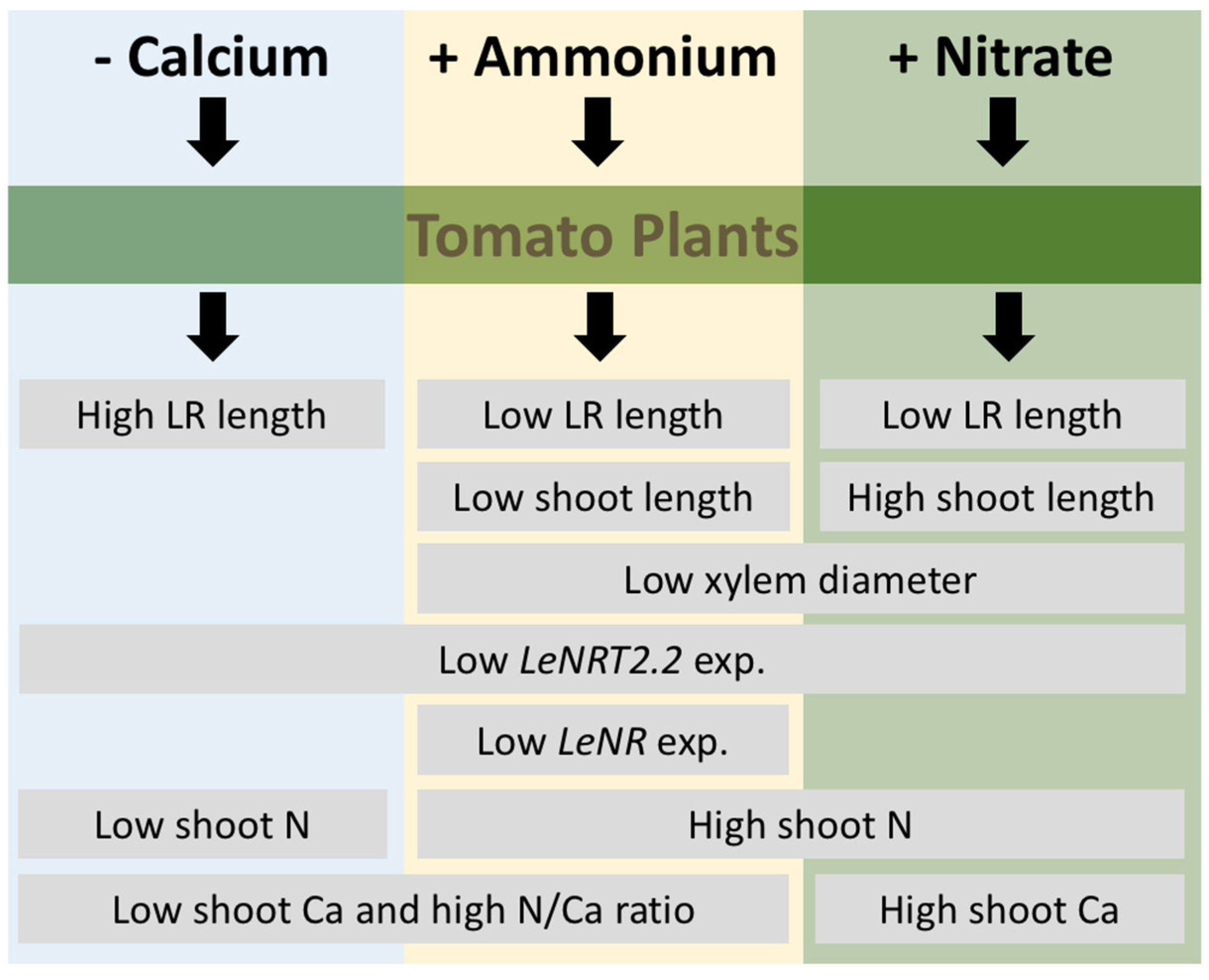
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bénard, C.; Gautier, H.; Bourgaud, F.; Grasselly, D.; Navez, B.; Caris-Veyrat, C.; Weiss, M.; Génard, M. Effects of Low Nitrogen Supply on Tomato (Solanum Lycopersicum) Fruit Yield and Quality with Special Emphasis on Sugars, Acids, Ascorbate, Carotenoids, and Phenolic Compounds. J. Agric. Food Chem. 2009, 57, 4112–4123. [Google Scholar] [CrossRef]
- Kusano, M.; Fukushima, A.; Redestig, H.; Saito, K. Metabolomic Approaches toward Understanding Nitrogen Metabolism in Plants. J. Exp. Bot. 2011, 62, 1439–1453. [Google Scholar] [CrossRef]
- Mengel, K.; Kirkby, E.A. Principles of Plant Nutrition, 3rd ed.; International Potash Institute: Bern, Switzerland, 1978. [Google Scholar]
- Krapp, A. Plant Nitrogen Assimilation and Its Regulation: A Complex Puzzle with Missing Pieces. Curr. Opin. Plant Biol. 2015, 25, 115–122. [Google Scholar] [CrossRef]
- Nacry, P.; Bouguyon, E.; Gojon, A. Nitrogen Acquisition by Roots: Physiological and Developmental Mechanisms Ensuring Plant Adaptation to a Fluctuating Resource. Plant Soil 2013, 370, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Pal’ove-Balang, P. Role of Nitrogen Metabolites on the Regulation of Nitrate Uptake Inmaize Seedlings. Acta Biol. Szeged. 2002, 46, 177–178. [Google Scholar]
- Issawi, M.; Guillaumot, D.; Sol, V.; Riou, C. Responses of an Adventitious Fast-Growing Plant to Photodynamic Stress: Comparative Study of Anionic and Cationic Porphyrin Effect on Arabidopsis Thaliana. Physiol. Plant. 2018, 162, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Jampeetong, A.; Brix, H.; Kantawanichkul, S. Effects of Inorganic Nitrogen Form on Growth, Morphology, N Uptake, and Nutrient Allocation in Hybrid Napier Grass (Pennisetum Purpureum × Pennisetum Americanum Cv. Pakchong1). Ecol. Eng. 2014, 73, 653–658. [Google Scholar] [CrossRef]
- Martínez-Andújar, C.; Ghanem, M.E.; Albacete, A.; Pérez-Alfocea, F. Response to Nitrate/Ammonium Nutrition of Tomato (Solanum Lycopersicum L.) Plants Overexpressing a Prokaryotic NH4+-Dependent Asparagine Synthetase. J. Plant Physiol. 2013, 170, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Ben-Oliel, G.; Kant, S.; Naim, M.; Rabinowitch, H.D.; Takeoka, G.R.; Buttery, R.G.; Kafkafi, U. Effects of Ammonium to Nitrate Ratio and Salinity on Yield and Fruit Quality of Large and Small Tomato Fruit Hybrids. J. Plant Nutr. 2005, 27, 1795–1812. [Google Scholar] [CrossRef]
- Gerendás, J.; Zhu, Z.; Bendixen, R.; Ratcliffe, R.G.; Sattelmacher, B. Physiological and Biochemical Processes Related to Ammonium Toxicity in Higher Plants. J. Plant Nutr. Soil Sci. 1997, 160, 239–251. [Google Scholar] [CrossRef]
- Fallahi, E.; Fallahi, B.; Peryea, F.J.; Neilsen, G.H.; Neilsen, D. Effects of Mineral Nutrition on Fruit Quality and Nutritional Disorders in Apples. Acta Hortic. 2010, 868, 49–59. [Google Scholar] [CrossRef]
- Paradiković, N.; Teklić, T.; Horvat, D.; Vinković, T.; Gumze, A. Influence of Different Nitrogen Form Application on the Incidence of BER and Elementary Composition of Bell Peppers. In Cereal Research Communications; Springer: Berlin/Heidelberg, Germany, 2007; Volume 35, pp. 897–900. [Google Scholar] [CrossRef]
- Taylor, M.D.; Locascio, S.J. Blossom-End Rot: A Calcium Deficiency. J. Plant Nutr. 2004, 27, 123–139. [Google Scholar] [CrossRef]
- Masaharu, K.; Araki, T.; Yoshida, S.; Eguchi, T. Dependence of Calcium Uptake on Water and Respiration in Roots of Tomato Plants (Lycopersicon Esculentum Mill). Biotronics 1999, 28, 121–130. [Google Scholar]
- Hirschi, K.D. The Calcium Conundrum. Both Versatile Nutrient and Specific Signal. Plant Physiology. Am. Soc. Plant Biol. 2004, 2348–2442. [Google Scholar] [CrossRef] [Green Version]
- Montanaro, G.; Dichio, B.; Lang, A.; Mininni, A.N.; Nuzzo, V.; Clearwater, M.J.; Xiloyannis, C. Internal versus External Control of Calcium Nutrition in Kiwifruit. J. Plant Nutr. Soil Sci. 2014, 177, 819–830. [Google Scholar] [CrossRef]
- Karley, A.J.; White, P.J. Moving Cationic Minerals to Edible Tissues: Potassium, Magnesium, Calcium. In Current Opinion in Plant Biology; Elsevier: Amsterdam, The Netherlands, 2009; pp. 291–298. [Google Scholar] [CrossRef]
- White, P.J. The Pathways of Calcium Movement to the Xylem. J. Exp. Bot. 2001, 52, 891–899. [Google Scholar] [CrossRef]
- Kirkby, E.A.; Pilbeam, D.J. Calcium as a Plant Nutrient. In Plant, Cell & Environment; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1984; pp. 397–405. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Calcium in Plants. In Annals of Botany; Oxford Academic: Oxford, UK, 2003; pp. 487–511. [Google Scholar] [CrossRef]
- Kanayama, Y.; Kochetov, A. Abiotic Stress Biology in Horticultural Plants; Springer: Japan, 2015; Available online: https://www.springer.com/gp/book/9784431552505 (accessed on 19 March 2021). [CrossRef]
- Fenn, L.B.; Taylor, R.M. Calcium Stimulation of Ammonium Absorption in Plants; Elsevier: Amsterdam, The Netherlands, 1991; Volume 24, pp. 39–47. [Google Scholar] [CrossRef]
- Hernández-Gómez, E.; Valdez-Aguilar, L.A.; Cartmill, D.L.; Cartmill, A.D.; Alia-Tajacal, I. Supplementary Calcium Ameliorates Ammonium Toxicity by Improving Water Status in Agriculturally Important Species. AoB Plants 2015, 7, plv105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Z.L.; Alva, A.K.; Calvert, D.V.; Li, Y.C.; Banks, D.J. Effects of Nitrogen Fertilization of Grapefruit Trees on Soil Acidification and Nutrient Availability in a Riviera Fine Sand. Plant Soil 1999, 206, 11–19. [Google Scholar] [CrossRef]
- Wieczorek, P.; Wrzesińska, B.; Obrepalska-Steplowska, A. Assessment of Reference Gene Stability Influenced by Extremely Divergent Disease Symptoms in Solanum Lycopersicum L. J. Virol. Methods 2013, 194, 161–168. [Google Scholar] [CrossRef]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.H.; Karlen, Y.; Bakker, O.; van den Hoff, M.J.B.; Moorman, A.F.M. Amplification Efficiency: Linking Baseline and Bias in the Analysis of Quantitative PCR Data. Nucleic Acids Res. 2009, 37. [Google Scholar] [CrossRef] [Green Version]
- Swietlik, D. The Interaction between Calcium Chloride and Ammonium-Nitrogen on Growth, Nitrogen Uptake and Translocation in Apple and Sour Orange. Acta Hortic. 2006, 721, 159–164. [Google Scholar] [CrossRef]
- Mokhele, B.; Zhan, X.; Yang, G.; Zhang, X. Review: Nitrogen Assimilation in Crop Plants and Its Affecting Factors. Can. J. Plant Sci. 2012, 399–405. [Google Scholar] [CrossRef]
- Sun, H.; Li, J.; Song, W.; Tao, J.; Huang, S.; Chen, S.; Hou, M.; Xu, G.; Zhang, Y. Nitric Oxide Generated by Nitrate Reductase Increases Nitrogen Uptake Capacity by Inducing Lateral Root Formation and Inorganic Nitrogen Uptake under Partial Nitrate Nutrition in Rice. J. Exp. Bot. 2015, 66, 2449–2459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rančić, D.; Quarrie, S.P.; Pećinar, I. Anatomy of Tomato Fruit and Fruit Pedicel during Fruit Development. Microsc. Sci. Technol. Appl. Educ. 2010, 2, 851–861. [Google Scholar]
- Andrews, M.; Raven, J.A.; Lea, P.J. Do Plants Need Nitrate? The Mechanisms by Which Nitrogen Form Affects Plants. Ann. Appl. Biol. 2013, 163, 174–199. [Google Scholar] [CrossRef]
- Saure, M.C. Why Calcium Deficiency Is Not the Cause of Blossom-End Rot in Tomato and Pepper Fruit—A Reappraisal. Sci. Hortic. 2014, 174, 151–154. [Google Scholar] [CrossRef]
- Nunes Júnior, E.S.; De Medeiros, J.F.; De Oliveira, F.D.A.; Lima, L.A.; Bezerra, F.M.S.; De, R.; Alves, C. Nitrogen and Potassium Fertigation in Bell Pepper Cultivated in Greenhouse Using Fertigation Managements 1. Rev. Bras. Eng. Agrícola Ambient. 2017, 21, 186–190. [Google Scholar] [CrossRef] [Green Version]
- Hagassou, D.; Francia, E.; Ronga, D.; Buti, M. Blossom End-Rot in Tomato (Solanum Lycopersicum L.): A Multi-Disciplinary Overview of Inducing Factors and Control Strategies. Sci. Hortic. 2019, 249, 49–58. [Google Scholar] [CrossRef]
- Liu, Y.; Lai, N.; Gao, K.; Chen, F.; Yuan, L.; Mi, G. Ammonium Inhibits Primary Root Growth by Reducing the Length of Meristem and Elongation Zone and Decreasing Elemental Expansion Rate in the Root Apex in Arabidopsis Thaliana. PLoS ONE 2013, 8, 61031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, P.; Carvajal, M.; Cerdá, A.; Martínez, V. Salinity and Ammonium/Nitrate Interactions on Tomato Plant Development, Nutrition, and Metabolites. J. Plant Nutr. 2001, 24, 1561–1573. [Google Scholar] [CrossRef]
- Rossato, L.; Lainé, P.; Ourry, A. Nitrogen Storage and Remobilization in Brassica Napus L. during the Growth Cycle: Nitrogen Fluxes within the Plant and Changes in Soluble Protein Patterns. J. Exp. Bot. 2001, 52, 1655–1663. [Google Scholar] [CrossRef] [Green Version]
- Trapani, N.; Hall, A.J.; Weber, M. Effects of Constant and Variable Nitrogen Supply on Sunflower (Helianthus Annuus L.) Leaf Cell Number and Size. Ann. Bot. 1999, 84, 599–606. [Google Scholar] [CrossRef] [Green Version]
- Aloni, R. Role of Hormones in Controlling Vascular Differentiation and the Mechanism of Lateral Root Initiation. Planta 2013, 238, 819–830. [Google Scholar] [CrossRef]
- Dražeta, L.; Lang, A.; Cappellini, C.; Hall, A.J.; Volz, R.K.; Jameson, P.E. Vessel Differentiation in the Pedicel of Apple and the Effects of Auxin Transport Inhibition. Physiol. Plant. 2004, 120, 162–170. [Google Scholar] [CrossRef]
- O’Brien, J.A.; Vega, A.; Bouguyon, E.; Krouk, G.; Gojon, A.; Coruzzi, G.; Gutiérrez, R.A. Nitrate Transport, Sensing, and Responses in Plants. Mol. Plant 2016, 9, 837–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li-Li, G.; Sheng-Feng, W.; Zi-Fei, L.; Jin-Sheng, H.; Rong-Le, L.; Hong, W. Two Cultivars of Peanut (Arachis Hypogaea) Seedlings Show Different Tolerance to Calcium Deficiency. J. Plant Nutr. 2015, 4167. [Google Scholar] [CrossRef]
- Rietra, R.P.J.J.; Heinen, M.; Dimkpa, C.O.; Bindraban, P.S. Effects of Nutrient Antagonism and Synergism on Yield and Fertilizer Use Efficiency. Commun. Soil Sci. Plant Anal. 2017, 48, 1895–1920. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, I.; Sady, W.; Leja, M. Effect of Nitrogen Form and Type of Polyethylene Film Covering Tunnel on Nutrient Content of Hydroponically Grown Sweet Pepper. Veg. Crop. Res. Bull. 2009, 71, 69–78. [Google Scholar] [CrossRef]
- White, P.J. Ion Uptake Mechanisms of Individual Cells and Roots: Short-Distance Transport; Elsevier Ltd.: Amsterdam, The Netherlands, 2011. [Google Scholar] [CrossRef]
- Wang, H.-Y.; Wang, Y.-H.; Du, C.-W.; Xu, F.-S.; Yang, Y.-H. Effects of Boron and Calcium Supply on Calcium Fractionation in Plants and Suspension Cells of Rape Cultivars with Different Boron Efficiency. J. Plant Nutr. 2003, 26, 789–806. [Google Scholar] [CrossRef]
- Bonomelli, C.; Mogollón, R.; de Freitas, S.T.; Zoffoli, J.P.; Contreras, C. Nutritional Relationships in Bitter Pit-Affected Fruit and the Feasibility of Vis-NIR Models to Determine Calcium Concentration in “Fuji” Apples. Agronomy 2020, 10, 1476. [Google Scholar] [CrossRef]
- Do Amarante, C.V.T.; Miqueloto, A.; de Freitas, S.T.; Steffens, C.A.; Silveira, J.P.G.; Corrêa, T.R. Fruit Sampling Methods to Quantify Calcium and Magnesium Contents to Predict Bitter Pit Development in “Fuji” Apple: A Multivariate Approach. Sci. Hortic. 2013, 157, 19–23. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonomelli, C.; de Freitas, S.T.; Aguilera, C.; Palma, C.; Garay, R.; Dides, M.; Brossard, N.; O’Brien, J.A. Ammonium Excess Leads to Ca Restrictions, Morphological Changes, and Nutritional Imbalances in Tomato Plants, Which Can Be Monitored by the N/Ca Ratio. Agronomy 2021, 11, 1437. https://doi.org/10.3390/agronomy11071437
Bonomelli C, de Freitas ST, Aguilera C, Palma C, Garay R, Dides M, Brossard N, O’Brien JA. Ammonium Excess Leads to Ca Restrictions, Morphological Changes, and Nutritional Imbalances in Tomato Plants, Which Can Be Monitored by the N/Ca Ratio. Agronomy. 2021; 11(7):1437. https://doi.org/10.3390/agronomy11071437
Chicago/Turabian StyleBonomelli, Claudia, Sergio Tonetto de Freitas, Camila Aguilera, Carola Palma, Rebeca Garay, Maximiliano Dides, Natalia Brossard, and José Antonio O’Brien. 2021. "Ammonium Excess Leads to Ca Restrictions, Morphological Changes, and Nutritional Imbalances in Tomato Plants, Which Can Be Monitored by the N/Ca Ratio" Agronomy 11, no. 7: 1437. https://doi.org/10.3390/agronomy11071437
APA StyleBonomelli, C., de Freitas, S. T., Aguilera, C., Palma, C., Garay, R., Dides, M., Brossard, N., & O’Brien, J. A. (2021). Ammonium Excess Leads to Ca Restrictions, Morphological Changes, and Nutritional Imbalances in Tomato Plants, Which Can Be Monitored by the N/Ca Ratio. Agronomy, 11(7), 1437. https://doi.org/10.3390/agronomy11071437